
Rhodanese-Fold Containing Proteins in Humans: Not Just Key Players in Sulfur Trafficking

 and
and 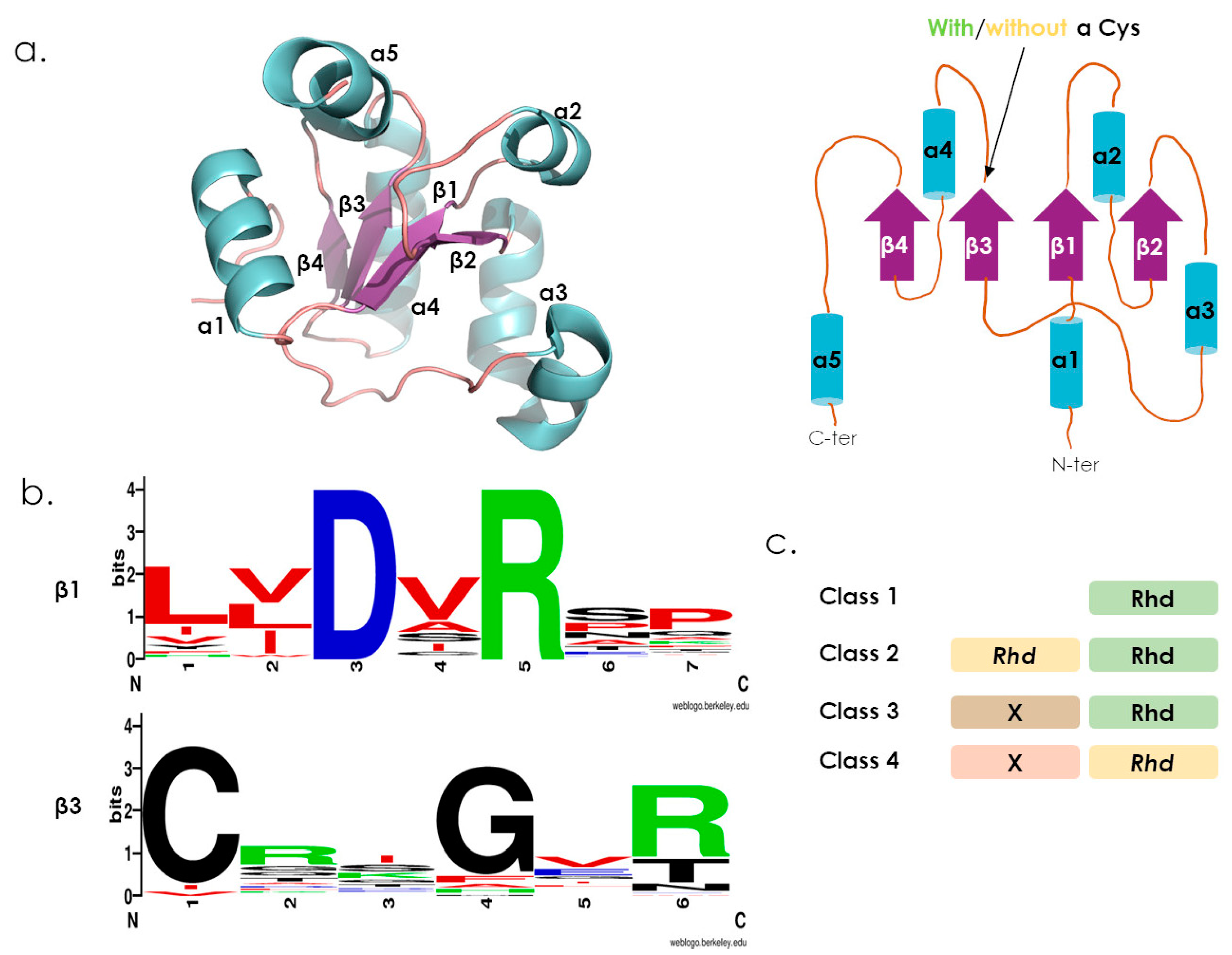{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Rhodanese-Fold and Domains Organization
3. Reactions Catalyzed by Active Rhodanese Domains
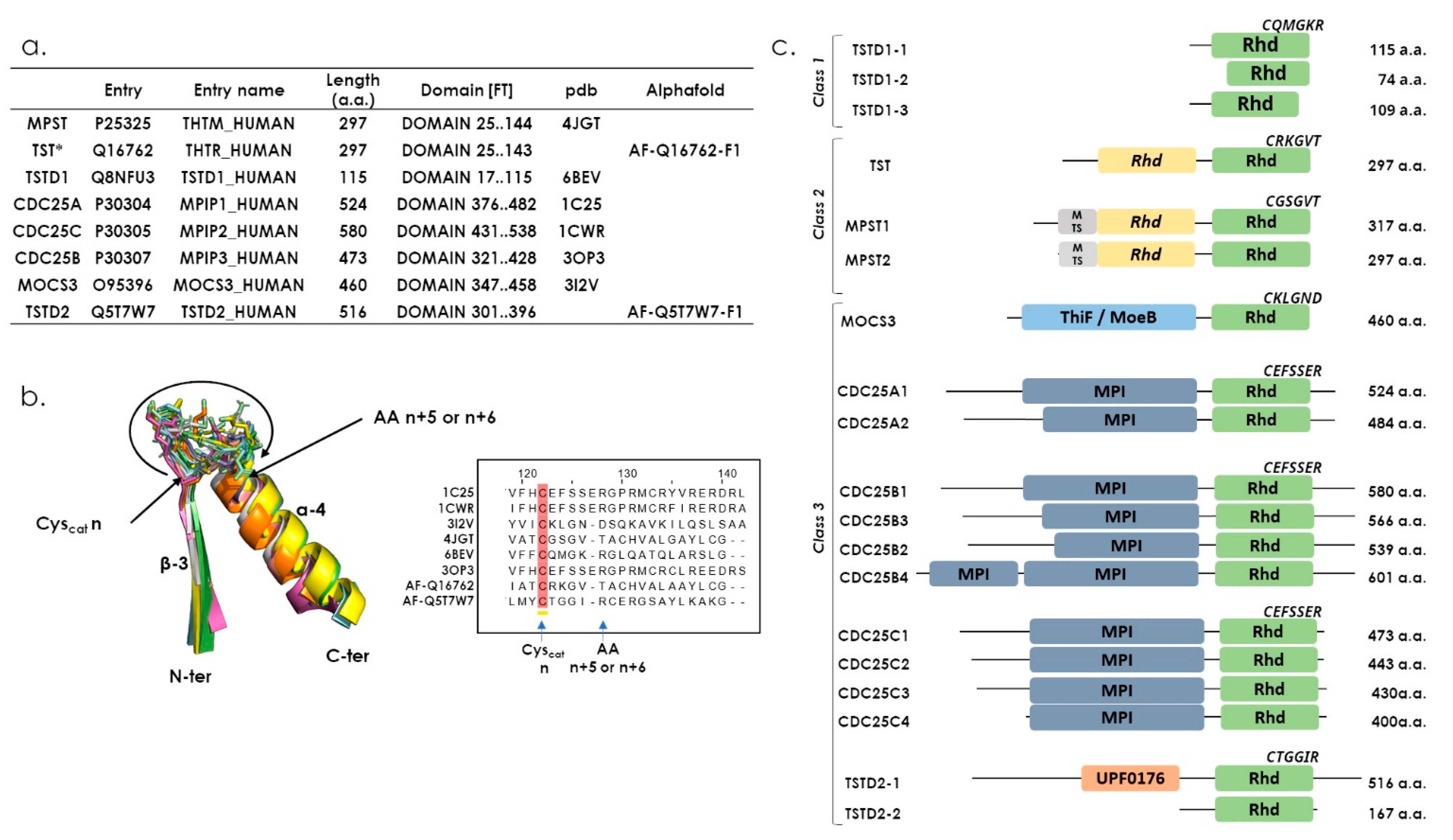
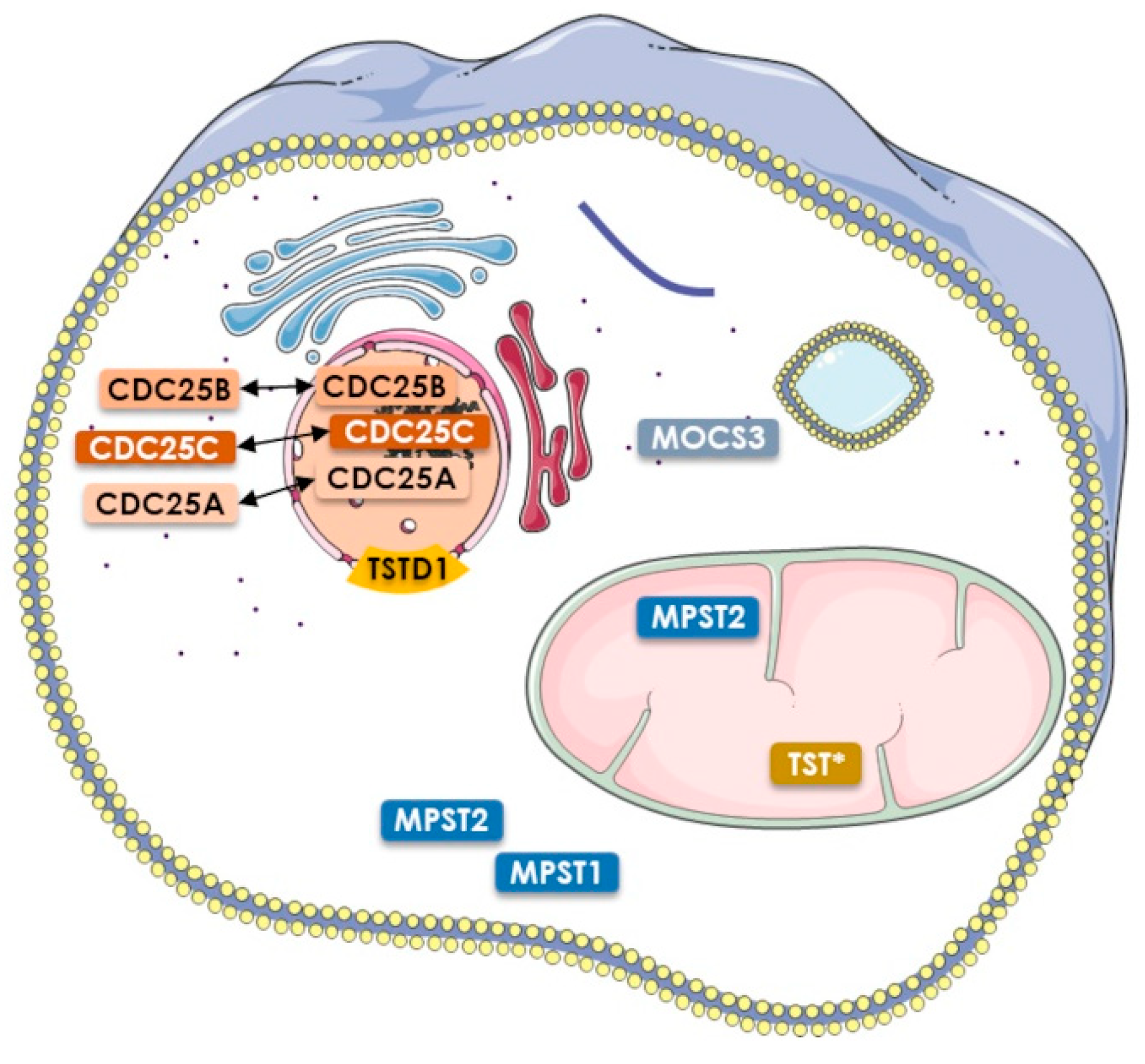
4. The Human Rhodanese Repertoire
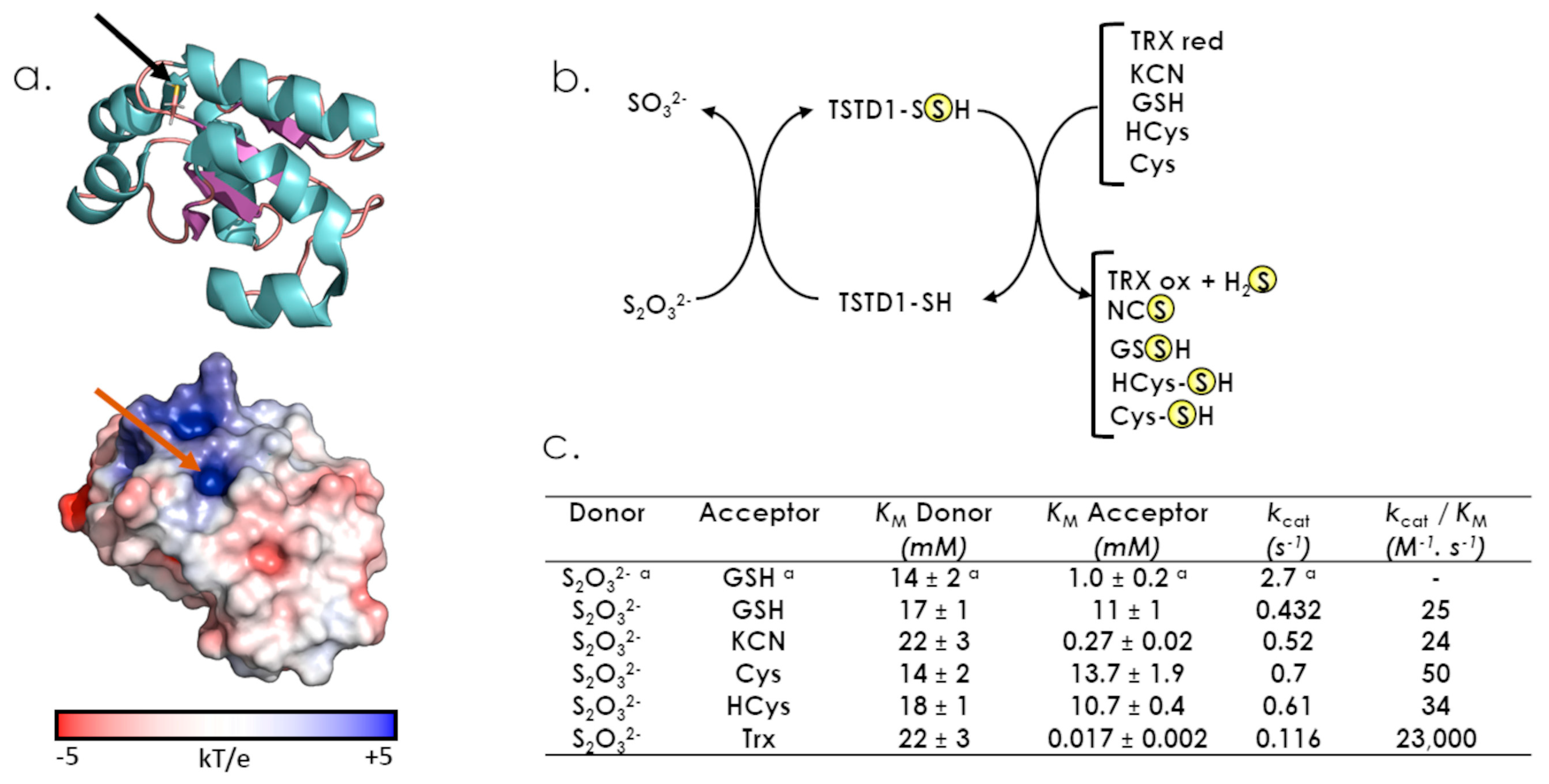
5. TSTD1: The Unique Class One Rhd Enzyme, So Far
6. The Two Rhd Classes, TST and MPST (Two Evolutionarily Related Sulfurtransferases with Different Functions)
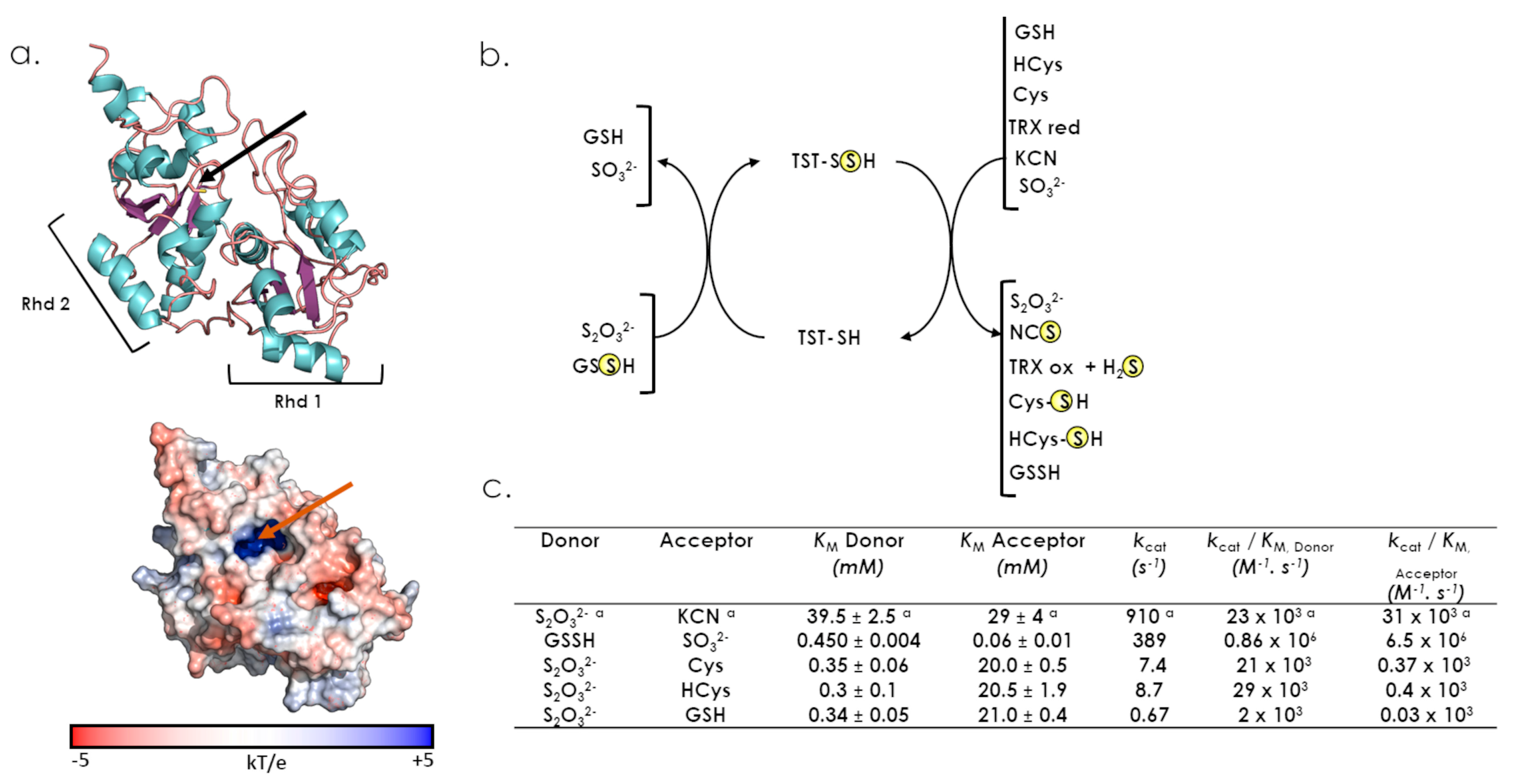
6.1. TST
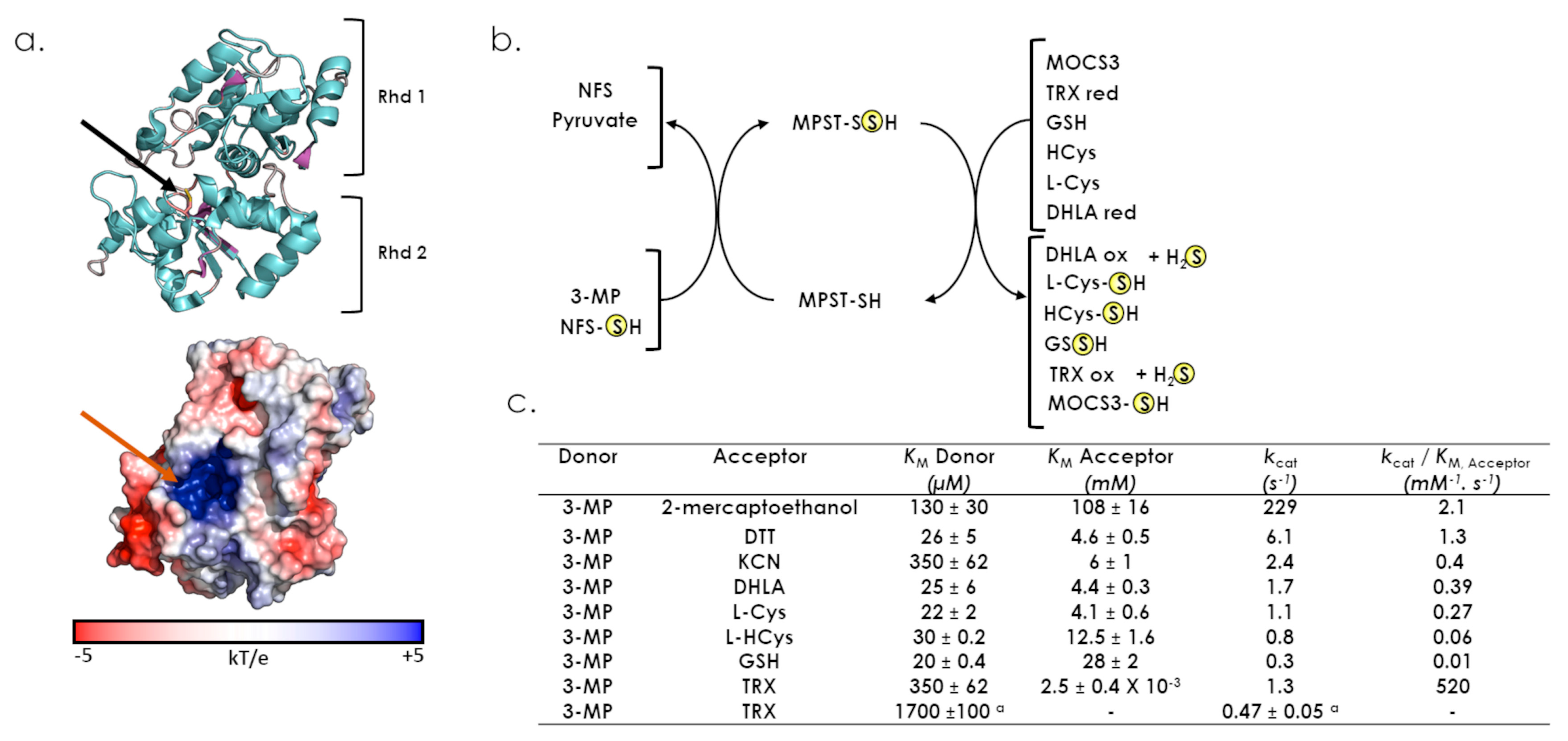
6.2. MPST
7. The Class Three Rhd Enzymes in Humans, the Most Versatile Rhd Domains
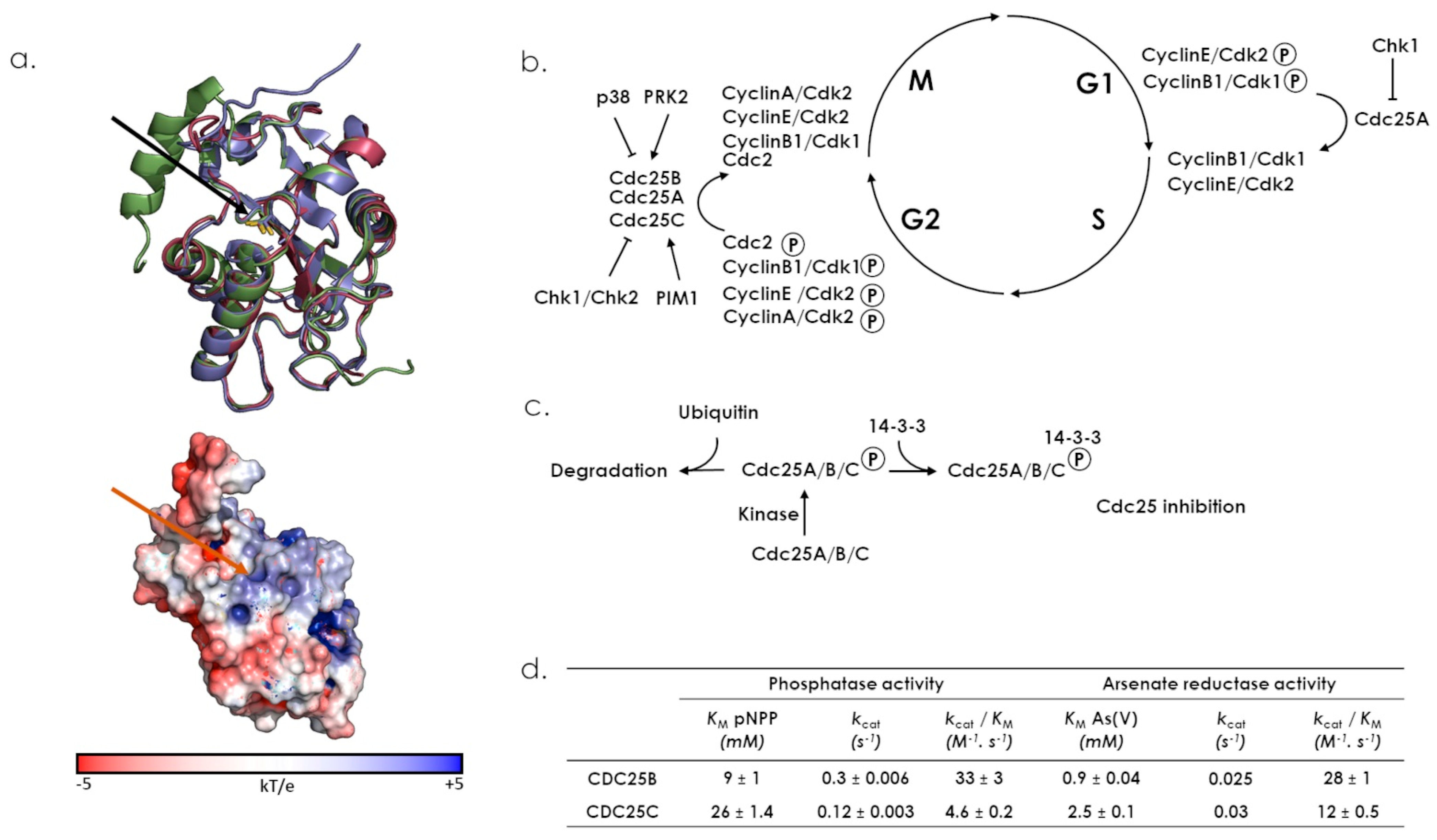
7.1. CDC25s
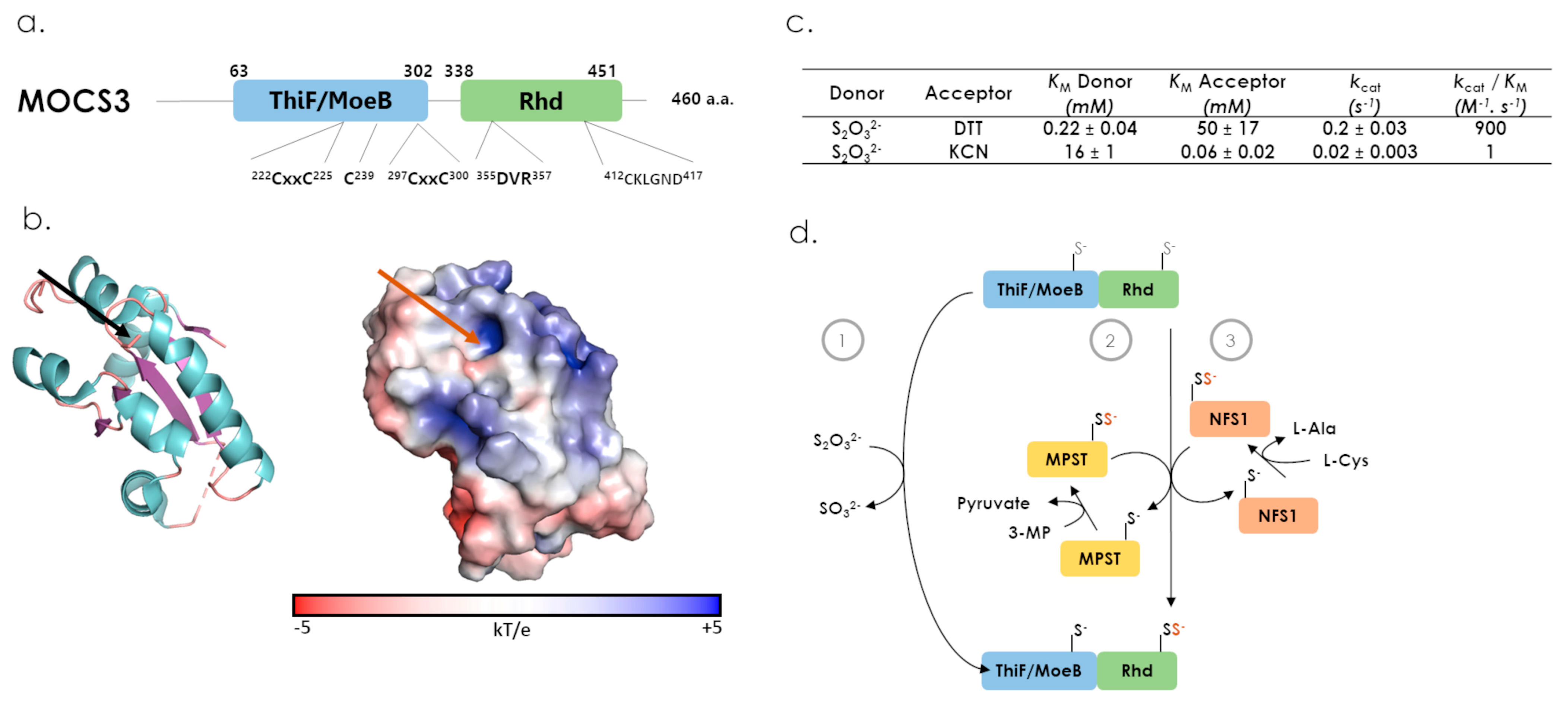
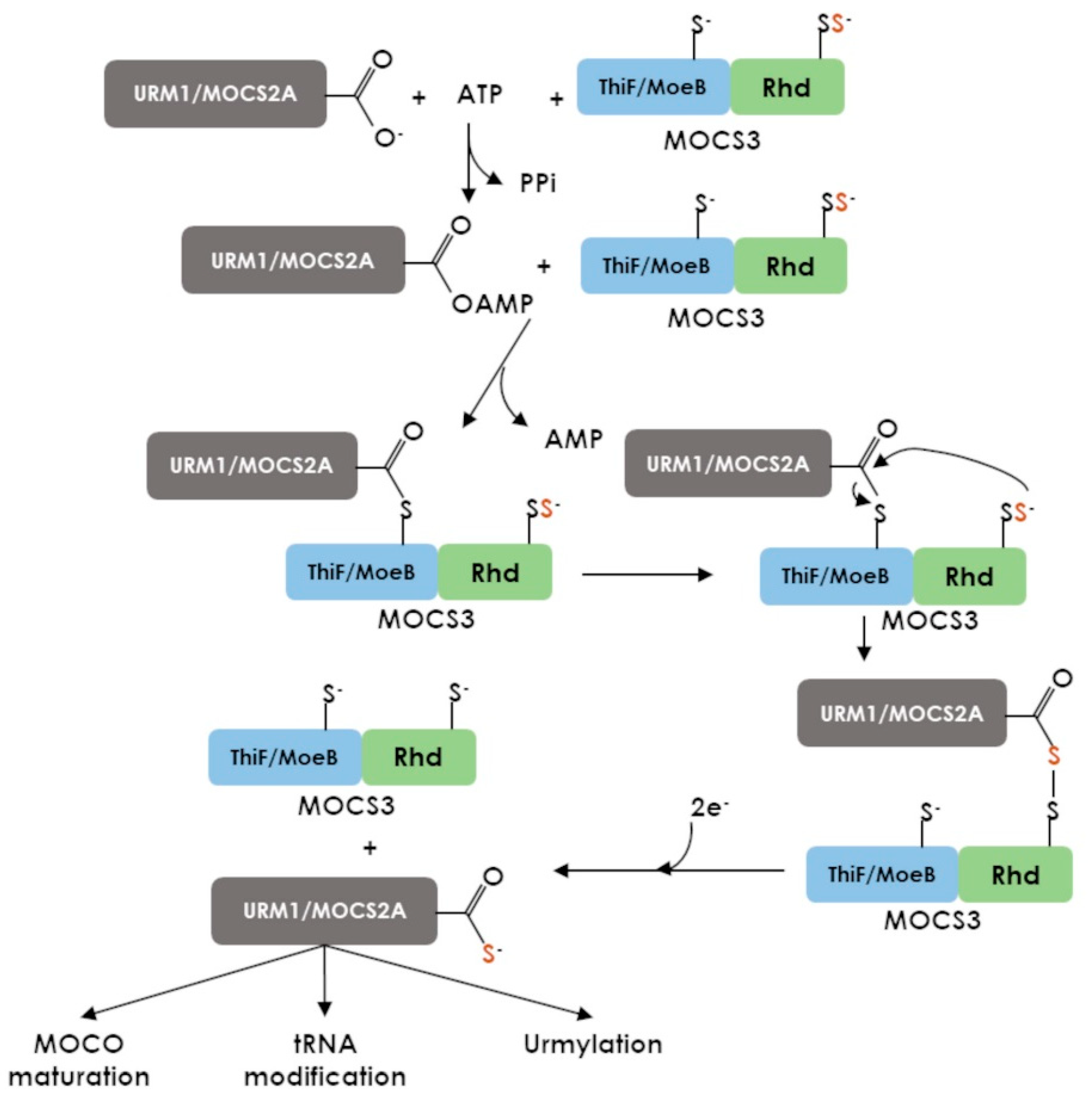
7.2. MOCS3
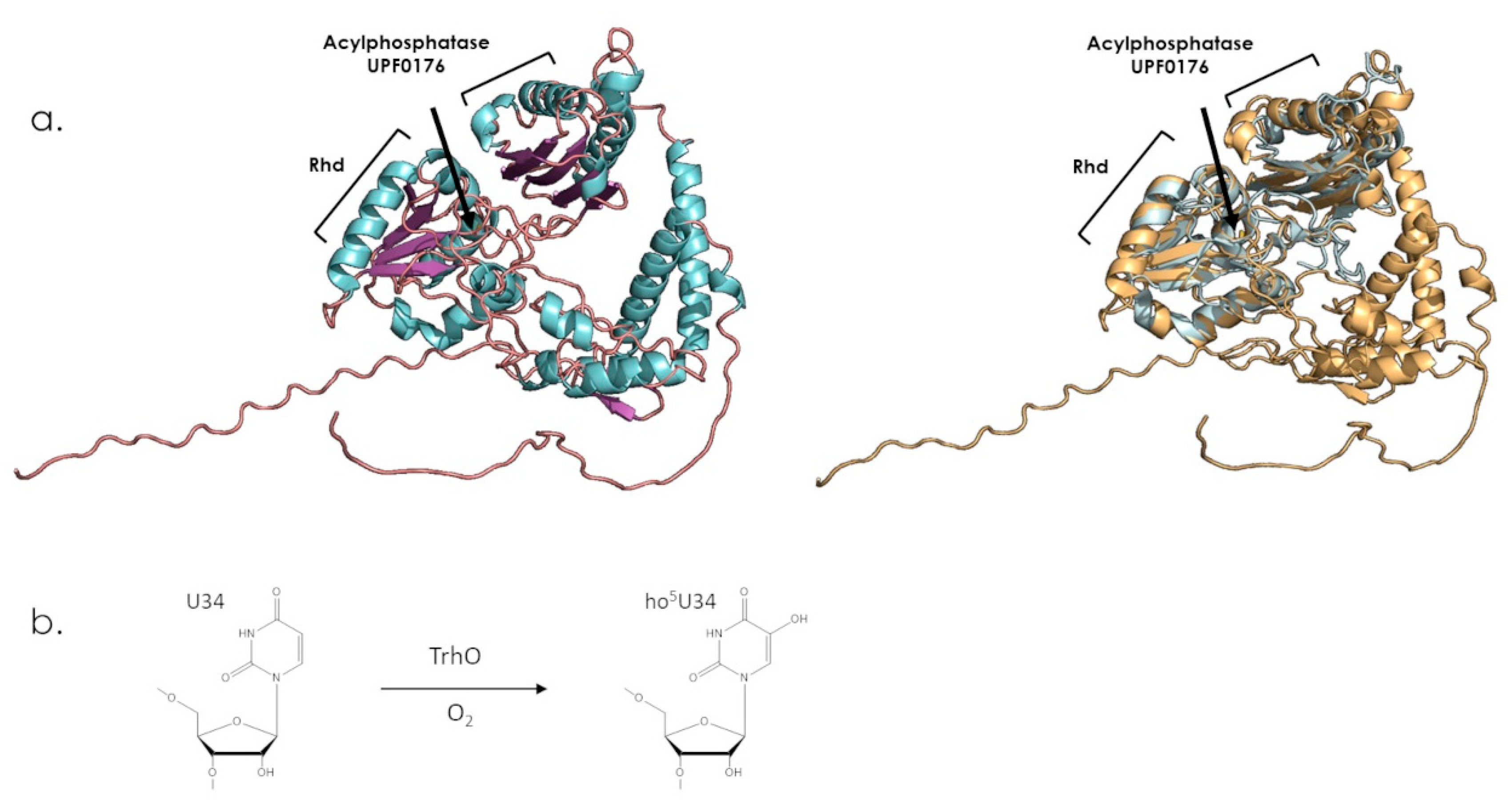
7.3. TSTD2
8. Concluding Remarks
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bordo, D.; Bork, P. The Rhodanese/Cdc25 Phosphatase Superfamily. Sequence-Structure-Function Relations. EMBO Rep. 2002, 3, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Cipollone, R.; Ascenzi, P.; Visca, P. Common Themes and Variations in the Rhodanese Superfamily. IUBMB Life 2007, 59, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Selles, B.; Moseler, A.; Rouhier, N.; Couturier, J. Rhodanese Domain-Containing Sulfurtransferases: Multifaceted Proteins Involved in Sulfur Trafficking in Plants. J. Exp. Bot. 2019, 70, 4139–4154. [Google Scholar] [CrossRef]
- Termathe, M.; Leidel, S.A. Urm1: A Non-Canonical UBL. Biomolecules 2021, 11, 139. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Sun, X.; Wang, R. Hydrogen Sulfide-Induced Apoptosis of Human Aorta Smooth Muscle Cells via the Activation of Mitogen-Activated Protein Kinases and Caspase-3. FASEB J. 2004, 18, 1782–1784. [Google Scholar] [CrossRef] [PubMed]
- Szabó, G.; Veres, G.; Radovits, T.; Gerő, D.; Módis, K.; Miesel-Gröschel, C.; Horkay, F.; Karck, M.; Szabó, C. Cardioprotective Effects of Hydrogen Sulfide. Nitric Oxide 2011, 25, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Calenic, B.; Yaegaki, K.; Ishkitiev, N.; Kumazawa, Y.; Imai, T.; Tanaka, T. P53-Pathway Activity and Apoptosis in Hydrogen Sulfide-Exposed Stem Cells Separated from Human Gingival Epithelium. J. Periodontal Res. 2013, 48, 322–330. [Google Scholar] [CrossRef]
- Talaei, F.; van Praag, V.M.; Henning, R.H. Hydrogen Sulfide Restores a Normal Morphological Phenotype in Werner Syndrome Fibroblasts, Attenuates Oxidative Damage and Modulates MTOR Pathway. Pharmacol. Res. 2013, 74, 34–44. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Bimai, O.; Arragain, S.; Golinelli-Pimpaneau, B. Structure-Based Mechanistic Insights into Catalysis by TRNA Thiolation Enzymes. Curr. Opin. Struct. Biol. 2020, 65, 69–78. [Google Scholar] [CrossRef]
- Schwarz, G. Molybdenum Cofactor and Human Disease. Curr. Opin. Chem. Biol. 2016, 31, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, K.E.; Kinch, L.N.; Schaeffer, R.D.; Grishin, N.V. Functional Analysis of Rossmann-like Domains Reveals Convergent Evolution of Topology and Reaction Pathways. PLoS Comput. Biol. 2019, 15, e1007569. [Google Scholar] [CrossRef] [Green Version]
- Avvakumov, G.V.; Walker, J.R.; Xue, S.; Finerty, P.J.; Mackenzie, F.; Newman, E.M.; Dhe-Paganon, S. Amino-Terminal Dimerization, NRDP1-Rhodanese Interaction, and Inhibited Catalytic Domain Conformation of the Ubiquitin-Specific Protease 8 (USP8). J. Biol. Chem. 2006, 281, 38061–38070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guretzki, S.; Papenbrock, J. Characterization of the Sulfurtransferase Family from Oryza Sativa L. Plant Physiol. Biochem. 2011, 49, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Moseler, A.; Selles, B.; Rouhier, N.; Couturier, J. Novel Insights into the Diversity of the Sulfurtransferase Family in Photosynthetic Organisms with Emphasis on Oak. New Phytol. 2019, 226, 967–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forlani, F.; Carpen, A.; Pagani, S. Evidence That Elongation of the Catalytic Loop of the Azotobacter Vinelandii Rhodanese Changed Selectivity from Sulfur- to Phosphate-containing Substrates. Protein Eng. Des. Sel. 2003, 16, 515–519. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, H.; Sheng, J.; Ajees, A.A.; Mukhopadhyay, R.; Rosen, B.P. Adventitious Arsenate Reductase Activity of the Catalytic Domain of the Human Cdc25B and Cdc25C Phosphatases. Biochemistry 2010, 49, 802–809. [Google Scholar] [CrossRef] [Green Version]
- Spallarossa, A.; Forlani, F.; Carpen, A.; Armirotti, A.; Pagani, S.; Bolognesi, M.; Bordo, D. The “Rhodanese” Fold and Catalytic Mechanism of 3-Mercaptopyruvate Sulfurtransferases: Crystal Structure of SseA from Escherichia Coli. J. Mol. Biol. 2004, 335, 583–593. [Google Scholar] [CrossRef]
- Gottlin, E.B.; Xu, X.; Epstein, D.M.; Burke, S.P.; Eckstein, J.W.; Ballou, D.P.; Dixon, J.E. Kinetic Analysis of the Catalytic Domain of Human Cdc25B. J. Biol. Chem. 1996, 271, 27445–27449. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, R.; Shi, J.; Rosen, B.P. Purification and Characterization of Acr2p, TheSaccharomyces Cerevisiae Arsenate Reductase. J. Biol. Chem. 2000, 275, 21149–21157. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, R.; Bisacchi, D.; Zhou, Y.; Armirotti, A.; Bordo, D. Structural Characterization of the As/Sb Reductase LmACR2 from Leishmania Major. J. Mol. Biol. 2009, 386, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Matthies, A.; Rajagopalan, K.V.; Mendel, R.R.; Leimkühler, S. Evidence for the Physiological Role of a Rhodanese-like Protein for the Biosynthesis of the Molybdenum Cofactor in Humans. Proc. Natl. Acad. Sci. USA 2004, 101, 5946–5951. [Google Scholar] [CrossRef] [Green Version]
- Lec, J.-C.; Boutserin, S.; Mazon, H.; Mulliert, G.; Boschi-Muller, S.; Talfournier, F. Unraveling the Mechanism of Cysteine Persulfide Formation Catalyzed by 3-Mercaptopyruvate Sulfurtransferases. ACS Catal. 2018, 8, 2049–2059. [Google Scholar] [CrossRef]
- Chowdhury, M.M.; Dosche, C.; Löhmannsröben, H.-G.; Leimkühler, S. Dual Role of the Molybdenum Cofactor Biosynthesis Protein MOCS3 in TRNA Thiolation and Molybdenum Cofactor Biosynthesis in Humans. J. Biol. Chem. 2012, 287, 17297–17307. [Google Scholar] [CrossRef] [Green Version]
- Fräsdorf, B.; Radon, C.; Leimkühler, S. Characterization and Interaction Studies of Two Isoforms of the Dual Localized 3-Mercaptopyruvate Sulfurtransferase TUM1 from Humans. J. Biol. Chem. 2014, 289, 34543–34556. [Google Scholar] [CrossRef] [Green Version]
- Smirnov, A.; Comte, C.; Mager-Heckel, A.-M.; Addis, V.; Krasheninnikov, I.A.; Martin, R.P.; Entelis, N.; Tarassov, I. Mitochondrial Enzyme Rhodanese Is Essential for 5 S Ribosomal RNA Import into Human Mitochondria. J. Biol. Chem. 2010, 285, 30792–30803. [Google Scholar] [CrossRef] [Green Version]
- Smirnov, A.; Entelis, N.; Martin, R.P.; Tarassov, I. Biological Significance of 5S RRNA Import into Human Mitochondria: Role of Ribosomal Protein MRP-L18. Genes Dev. 2011, 25, 1289–1305. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, K.; Felix, S.B.; Flachmeier, C.; Heere, P.; Schulze, W.; Grunewald, I.; Pankow, H.; Hewelt, A.; Scherneck, S.; Bauer, D.; et al. Identification and Characterization of KAT, a Novel Gene Preferentially Expressed in Several Human Cancer Cell Lines. Biol. Chem. 2003, 384, 763–775. [Google Scholar] [CrossRef]
- Dalal, S.N.; Schweitzer, C.M.; Gan, J.; DeCaprio, J.A. Cytoplasmic Localization of Human Cdc25C during Interphase Requires an Intact 14-3-3 Binding Site. Mol. Cell Biol. 1999, 19, 4465–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindqvist, A.; Källström, H.; Lundgren, A.; Barsoum, E.; Rosenthal, C.K. Cdc25B Cooperates with Cdc25A to Induce Mitosis but Has a Unique Role in Activating Cyclin B1-Cdk1 at the Centrosome. J. Cell Biol. 2005, 171, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leisser, C.; Rosenberger, G.; Maier, S.; Fuhrmann, G.; Grusch, M.; Strasser, S.; Huettenbrenner, S.; Fassl, S.; Polgar, D.; Krieger, S.; et al. Subcellular Localisation of Cdc25A Determines Cell Fate. Cell Death Differ. 2004, 11, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Tsutsui, T.; Ray, D.; Blomquist, J.F.; Ichijo, H.; Ucker, D.S.; Kiyokawa, H. The Cell Cycle-Regulatory CDC25A Phosphatase Inhibits Apoptosis Signal-Regulating Kinase 1. Mol. Cell Biol. 2001, 21, 4818–4828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melideo, S.L.; Jackson, M.R.; Jorns, M.S. Biosynthesis of a Central Intermediate in Hydrogen Sulfide Metabolism by a Novel Human Sulfurtransferase and Its Yeast Ortholog. Biochemistry 2014, 53, 4739–4753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libiad, M.; Motl, N.; Akey, D.L.; Sakamoto, N.; Fearon, E.R.; Smith, J.L.; Banerjee, R. Thiosulfate Sulfurtransferase-like Domain-Containing 1 Protein Interacts with Thioredoxin. J. Biol. Chem. 2018, 293, 2675–2686. [Google Scholar] [CrossRef] [Green Version]
- Boschi-Muller, S.; Olry, A.; Antoine, M.; Branlant, G. The Enzymology and Biochemistry of Methionine Sulfoxide Reductases. Biochim. Biophys. Acta 2005, 1703, 231–238. [Google Scholar] [CrossRef]
- Schlecker, T.; Comini, M.A.; Melchers, J.; Ruppert, T.; Krauth-Siegel, R.L. Catalytic Mechanism of the Glutathione Peroxidase-Type Tryparedoxin Peroxidase of Trypanosoma Brucei. Biochem. J. 2007, 405, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Selles, B.; Hugo, M.; Trujillo, M.; Srivastava, V.; Wingsle, G.; Jacquot, J.-P.; Radi, R.; Rouhier, N. Hydroperoxide and Peroxynitrite Reductase Activity of Poplar Thioredoxin-Dependent Glutathione Peroxidase 5: Kinetics, Catalytic Mechanism and Oxidative Inactivation. Biochem. J. 2012, 442, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Furne, J.; Springfield, J.; Koenig, T.; DeMaster, E.; Levitt, M.D. Oxidation of Hydrogen Sulfide and Methanethiol to Thiosulfate by Rat Tissues: A Specialized Function of the Colonic Mucosa. Biochem. Pharmacol. 2001, 62, 255–259. [Google Scholar] [CrossRef]
- Lang, K. Die Rhodanbildung Im Tierkörper. Biochem. Z. 1933, 259, 37. [Google Scholar]
- Ploegman, J.H.; Drent, G.; Kalk, K.H.; Hol, W.G.J.; Heinrikson, R.L.; Keim, P.; Weng, L.; Russell, J. The Covalent and Tertiary Structure of Bovine Liver Rhodanese. Nature 1978, 273, 124–129. [Google Scholar] [CrossRef]
- Pallini, R.; Guazzi, G.C.; Cannella, C.; Cacace, M.G. Cloning and Sequence Analysis of the Human Liver Rhodanese: Comparison with the Bovine and Chicken Enzymes. Biochem. Biophys. Res. Commun. 1991, 180, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Aita, N.; Ishii, K.; Akamatsu, Y.; Ogasawara, Y.; Tanabe, S. Cloning and Expression of Human Liver Rhodanese CDNA. Biochem. Biophys. Res. Commun. 1997, 231, 56–60. [Google Scholar] [CrossRef]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the Human Mitochondrial Hydrogen Sulfide Oxidation Pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libiad, M.; Sriraman, A.; Banerjee, R. Polymorphic Variants of Human Rhodanese Exhibit Differences in Thermal Stability and Sulfur Transfer Kinetics. J. Biol. Chem. 2015, 290, 23579–23588. [Google Scholar] [CrossRef] [Green Version]
- Ploegman, J.H.; Drent, G.; Kalk, K.H.; Hol, W.G. Structure of Bovine Liver Rhodanese. I. Structure Determination at 2.5 A Resolution and a Comparison of the Conformation and Sequence of Its Two Domains. J. Mol. Biol. 1978, 123, 557–594. [Google Scholar] [CrossRef]
- Jackson, M.R.; Melideo, S.L.; Jorns, M.S. Human Sulfide: Quinone Oxidoreductase Catalyzes the First Step in Hydrogen Sulfide Metabolism and Produces a Sulfane Sulfur Metabolite. Biochemistry 2012, 51, 6804–6815. [Google Scholar] [CrossRef]
- Motl, N.; Skiba, M.A.; Kabil, O.; Smith, J.L.; Banerjee, R. Structural and Biochemical Analyses Indicate That a Bacterial Persulfide Dioxygenase-Rhodanese Fusion Protein Functions in Sulfur Assimilation. J. Biol. Chem. 2017, 292, 14026–14038. [Google Scholar] [CrossRef] [Green Version]
- Morton, N.M.; Beltram, J.; Carter, R.N.; Michailidou, Z.; Gorjanc, G.; McFadden, C.; Barrios-Llerena, M.E.; Rodriguez-Cuenca, S.; Gibbins, M.T.G.; Aird, R.E.; et al. Genetic Identification of Thiosulfate Sulfurtransferase as an Adipocyte-Expressed Antidiabetic Target in Mice Selected for Leanness. Nat. Med. 2016, 22, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Whitehouse, D.B.; Poole, C.J.; Kind, P.R.; Hopkinson, D.A. Rhodanese Isozymes in Three Subjects with Leber’s Optic Neuropathy. J. Med. Genet. 1989, 26, 113–115. [Google Scholar] [CrossRef] [Green Version]
- Cagianut, B.; Schnebli, H.P.; Rhyner, K.; Furrer, J. Decreased Thiosulfate Sulfur Transferase (Rhodanese) in Leber’s Hereditary Optic Atrophy. Klin. Wochenschr. 1984, 62, 850–854. [Google Scholar] [CrossRef]
- Pedre, B.; Dick, T.P. 3-Mercaptopyruvate Sulfurtransferase: An Enzyme at the Crossroads of Sulfane Sulfur Trafficking. Biol. Chem. 2021, 402, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.K.; Yamada, K.; Chiku, T.; Koutmos, M.; Banerjee, R. Structure and Kinetic Analysis of H2S Production by Human Mercaptopyruvate Sulfurtransferase. J. Biol. Chem. 2013, 288, 20002–20013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; White, R.H.; Cash, V.L.; Dean, D.R. Mechanism for the Desulfurization of L-Cysteine Catalyzed by the NifS Gene Product. Biochemistry 1994, 33, 4714–4720. [Google Scholar] [CrossRef] [PubMed]
- Alphey, M.S.; Williams, R.A.M.; Mottram, J.C.; Coombs, G.H.; Hunter, W.N. The Crystal Structure of Leishmania Major 3-Mercaptopyruvate Sulfurtransferase a Three-Domain Architecture with a Serine Protease-like Triad at the Active Site. J. Biol. Chem. 2003, 278, 48219–48227. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.K.; Vitvitsky, V.; Carballal, S.; Seravalli, J.; Banerjee, R. Thioredoxin Regulates Human Mercaptopyruvate Sulfurtransferase at Physiologically-Relevant Concentrations. J. Biol. Chem. 2020, 295, 6299–6311. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Koike, S.; Nirasawa, T.; Kimura, H.; Ogasawara, Y. Alternative Pathway of H2S and Polysulfides Production from Sulfurated Catalytic-Cysteine of Reaction Intermediates of 3-Mercaptopyruvate Sulfurtransferase. Biochem. Biophys. Res. Commun. 2018, 496, 648–653. [Google Scholar] [CrossRef]
- Crawhall, J.C.; Parker, R.; Sneddon, W.; Young, E.P. Beta-Mercaptolactate-Cysteine Disulfide in the Urine of a Mentally Retarded Patient. Am. J. Dis. Child. 1969, 117, 71–82. [Google Scholar] [CrossRef]
- Crawhall, J.C.; Bir, K.; Purkiss, P.; Stanbury, J.B. Sulfur Amino Acids as Precursors of Beta-Mercaptolactate-Cysteine Disulfide in Human Subjects. Biochem. Med. 1971, 5, 109–115. [Google Scholar] [CrossRef]
- Hannestad, U.; Mårtensson, J.; Sjödahl, R.; Sörbo, B. 3-Mercaptolactate Cysteine Disulfiduria: Biochemical Studies on Affected and Unaffected Members of a Family. Biochem. Med. 1981, 26, 106–114. [Google Scholar] [CrossRef]
- Nilsson, I.; Hoffmann, I. Cell Cycle Regulation by the Cdc25 Phosphatase Family. Prog. Cell Cycle. Res. 2000, 4, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Baldin, V.; Cans, C.; Superti-Furga, G.; Ducommun, B. Alternative Splicing of the Human CDC25B Tyrosine Phosphatase. Possible Implications for Growth Control? Oncogene 1997, 14, 2485–2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrest, A.R.; McCormack, A.K.; DeSouza, C.P.; Sinnamon, J.M.; Tonks, I.D.; Hayward, N.K.; Ellem, K.A.; Gabrielli, B.G. Multiple Splicing Variants of Cdc25B Regulate G2/M Progression. Biochem. Biophys. Res. Commun. 1999, 260, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Wegener, S.; Hampe, W.; Herrmann, D.; Schaller, H.C. Alternative Splicing in the Regulatory Region of the Human Phosphatases CDC25A and CDC25C. Eur. J. Cell Biol. 2000, 79, 810–815. [Google Scholar] [CrossRef]
- Bureik, M.; Rief, N.; Drescher, R.; Jungbluth, A.; Montenarh, M.; Wagner, P. An Additional Transcript of the Cdc25C Gene from A431 Cells Encodes a Functional Protein. Int. J. Oncol. 2000, 17, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, J. Cdc25 Phosphatases: Structure, Specificity, and Mechanism. Biochemistry 2007, 46, 3595–3604. [Google Scholar] [CrossRef]
- Reynolds, R.A.; Yem, A.W.; Wolfe, C.L.; Deibel, M.R.; Chidester, C.G.; Watenpaugh, K.D. Crystal Structure of the Catalytic Subunit of Cdc25B Required for G2/M Phase Transition of the Cell Cycle. J. Mol. Biol. 1999, 293, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.; Kristjánsdóttir, K.; Safi, A.; Parker, B.; Kiburz, B.; Rudolph, J. Remote Hot Spots Mediate Protein Substrate Recognition for the Cdc25 Phosphatase. Proc. Natl. Acad. Sci. USA 2004, 101, 16437–16441. [Google Scholar] [CrossRef] [Green Version]
- Sur, S.; Agrawal, D.K. Phosphatases and Kinases Regulating CDC25 Activity in the Cell Cycle: Clinical Implications of CDC25 Overexpression and Potential Treatment Strategies. Mol. Cell. Biochem. 2016, 416, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Arantes, G.M. The Catalytic Acid in the Dephosphorylation of the Cdk2-PTpY/CycA Protein Complex by Cdc25B Phosphatase. J. Phys. Chem. B 2008, 112, 15244–15247. [Google Scholar] [CrossRef] [PubMed]
- Boutros, R.; Dozier, C.; Ducommun, B. The When and Wheres of CDC25 Phosphatases. Curr. Opin. Cell Biol. 2006, 18, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Wang, Y.; Guan, J.; Wang, H. DNA Damage Checkpoint Control in Cells Exposed to Ionizing Radiation. Oncogene 2003, 22, 5834–5847. [Google Scholar] [CrossRef] [Green Version]
- Karlsson-Rosenthal, C.; Millar, J.B.A. Cdc25: Mechanisms of Checkpoint Inhibition and Recovery. Trends Cell Biol. 2006, 16, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Huang, S. The Role of Cdc25A in the Regulation of Cell Proliferation and Apoptosis. Anticancer Agents Med. Chem. 2012, 12, 631–639. [Google Scholar] [CrossRef]
- Fei, F.; Qu, J.; Liu, K.; Li, C.; Wang, X.; Li, Y.; Zhang, S. The Subcellular Location of Cyclin B1 and CDC25 Associated with the Formation of Polyploid Giant Cancer Cells and Their Clinicopathological Significance. Lab. Investig. 2019, 99, 483–498. [Google Scholar] [CrossRef]
- Liu, K.; Zheng, M.; Lu, R.; Du, J.; Zhao, Q.; Li, Z.; Li, Y.; Zhang, S. The Role of CDC25C in Cell Cycle Regulation and Clinical Cancer Therapy: A Systematic Review. Cancer Cell Int. 2020, 20, 213. [Google Scholar] [CrossRef]
- Lake, M.W.; Wuebbens, M.M.; Rajagopalan, K.V.; Schindelin, H. Mechanism of Ubiquitin Activation Revealed by the Structure of a Bacterial MoeB–MoaD Complex. Nature 2001, 414, 325–329. [Google Scholar] [CrossRef]
- Krepinsky, K.; Leimkühler, S. Site-Directed Mutagenesis of the Active Site Loop of the Rhodanese-like Domain of the Human Molybdopterin Synthase Sulfurase MOCS3. Major Differences in Substrate Specificity between Eukaryotic and Bacterial Homologs. FEBS J. 2007, 274, 2778–2787. [Google Scholar] [CrossRef]
- Marelja, Z.; Stöcklein, W.; Nimtz, M.; Leimkühler, S. A Novel Role for Human Nfs1 in the Cytoplasm: Nfs1 Acts as a Sulfur Donor for MOCS3, a Protein Involved in Molybdenum Cofactor Biosynthesis. J. Biol. Chem. 2008, 283, 25178–25185. [Google Scholar] [CrossRef] [Green Version]
- Leimkühler, S.; Rajagopalan, K.V. A Sulfurtransferase Is Required in the Transfer of Cysteine Sulfur in the in Vitro Synthesis of Molybdopterin from Precursor Z in Escherichia Coli. J. Biol. Chem. 2001, 276, 22024–22031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Mizushima, N.; Noda, T.; Ohsumi, Y. A Protein Conjugation System in Yeast with Homology to Biosynthetic Enzyme Reaction of Prokaryotes. J. Biol. Chem. 2000, 275, 7462–7465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Termathe, M.; Leidel, S.A. The Uba4 Domain Interplay Is Mediated via a Thioester That Is Critical for TRNA Thiolation through Urm1 Thiocarboxylation. Nucleic Acids Res. 2018, 46, 5171–5181. [Google Scholar] [CrossRef] [Green Version]
- Pabis, M.; Termathe, M.; Ravichandran, K.E.; Kienast, S.D.; Krutyhołowa, R.; Sokołowski, M.; Jankowska, U.; Grudnik, P.; Leidel, S.A.; Glatt, S. Molecular Basis for the Bifunctional Uba4-Urm1 Sulfur-Relay System in TRNA Thiolation and Ubiquitin-like Conjugation. EMBO J. 2020, 39, e105087. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R.; Kruse, T. Cell Biology of Molybdenum in Plants and Humans. Biochim. Biophys. Acta 2012, 1823, 1568–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijmans, J.G.M.; Schot, R.; de Klerk, J.B.C.; Williams, M.; de Coo, R.F.M.; Duran, M.; Verheijen, F.W.; van Slegtenhorst, M.; Mancini, G.M.S. Molybdenum Cofactor Deficiency: Identification of a Patient with Homozygote Mutation in the MOCS3 Gene. Am. J. Med. Genet. 2017, 173, 1601–1606. [Google Scholar] [CrossRef]
- Tian, Q.; Cao, Y.; Shu, L.; Chen, Y.; Peng, Y.; Wang, Y.; Chen, Y.; Wang, H.; Mao, X. Case Report: Compound Heterozygous Variants in MOCS3 Identified in a Chinese Infant With Molybdenum Cofactor Deficiency. Front. Genet. 2021, 12, 651878. [Google Scholar] [CrossRef]
- Neukranz, Y.; Kotter, A.; Beilschmidt, L.; Marelja, Z.; Helm, M.; Gräf, R.; Leimkühler, S. Analysis of the Cellular Roles of MOCS3 Identifies a MOCS3-Independent Localization of NFS1 at the Tips of the Centrosome. Biochemistry 2019, 58, 1786–1798. [Google Scholar] [CrossRef]
- Sakai, Y.; Kimura, S.; Suzuki, T. Dual Pathways of TRNA Hydroxylation Ensure Efficient Translation by Expanding Decoding Capability. Nat. Commun. 2019, 10, 2858. [Google Scholar] [CrossRef] [Green Version]
- Weiss, B.; Eberle, B.; Roeth, R.; de Bruin, C.; Lui, J.C.; Paramasivam, N.; Hinderhofer, K.; van Duyvenvoorde, H.A.; Baron, J.; Wit, J.M.; et al. Evidence That Non-Syndromic Familial Tall Stature Has an Oligogenic Origin Including Ciliary Genes. Front. Endocrinol. 2021, 12, 660731. [Google Scholar] [CrossRef]
- Si, T.; Ning, X.; Chen, H.; Hu, Z.; Dun, L.; Zheng, N.; Huang, P.; Yang, L.; Yi, P. ANTXR1 as a Potential Prognostic Biomarker for Hepatitis B Virus-Related Hepatocellular Carcinoma Identified by a Weighted Gene Correlation Network Analysis. J. Gastrointest. Oncol. 2021, 12, 3079–3092. [Google Scholar] [CrossRef] [PubMed]
- Kubota, M.; Zhang, B.-S.; Li, S.-Y.; Yoshida, Y.; Wang, H.; Adachi, A.; Matsutani, T.; Mine, S.; Machida, T.; Kamitsukasa, I.; et al. Serum Anti-TSTD2 Antibody as a Biomarker for Atherosclerosis-induced Ischemic Stroke and Chronic Kidney Disease. Med. Int. 2022, 3, 4. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsohaibani, R.; Claudel, A.-L.; Perchat-Varlet, R.; Boutserin, S.; Talfournier, F.; Boschi-Muller, S.; Selles, B. Rhodanese-Fold Containing Proteins in Humans: Not Just Key Players in Sulfur Trafficking. Antioxidants 2023, 12, 843. https://doi.org/10.3390/antiox12040843
Alsohaibani R, Claudel A-L, Perchat-Varlet R, Boutserin S, Talfournier F, Boschi-Muller S, Selles B. Rhodanese-Fold Containing Proteins in Humans: Not Just Key Players in Sulfur Trafficking. Antioxidants. 2023; 12(4):843. https://doi.org/10.3390/antiox12040843
Chicago/Turabian StyleAlsohaibani, Razan, Anne-Lise Claudel, Romain Perchat-Varlet, Séverine Boutserin, François Talfournier, Sandrine Boschi-Muller, and Benjamin Selles. 2023. "Rhodanese-Fold Containing Proteins in Humans: Not Just Key Players in Sulfur Trafficking" Antioxidants 12, no. 4: 843. https://doi.org/10.3390/antiox12040843
APA StyleAlsohaibani, R., Claudel, A. -L., Perchat-Varlet, R., Boutserin, S., Talfournier, F., Boschi-Muller, S., & Selles, B. (2023). Rhodanese-Fold Containing Proteins in Humans: Not Just Key Players in Sulfur Trafficking. Antioxidants, 12(4), 843. https://doi.org/10.3390/antiox12040843