



Urolithin A in Health and Diseases: Prospects for Parkinson’s Disease Management



Abstract
:1. Introduction
2. Urolithin A Production and Mechanism of Action
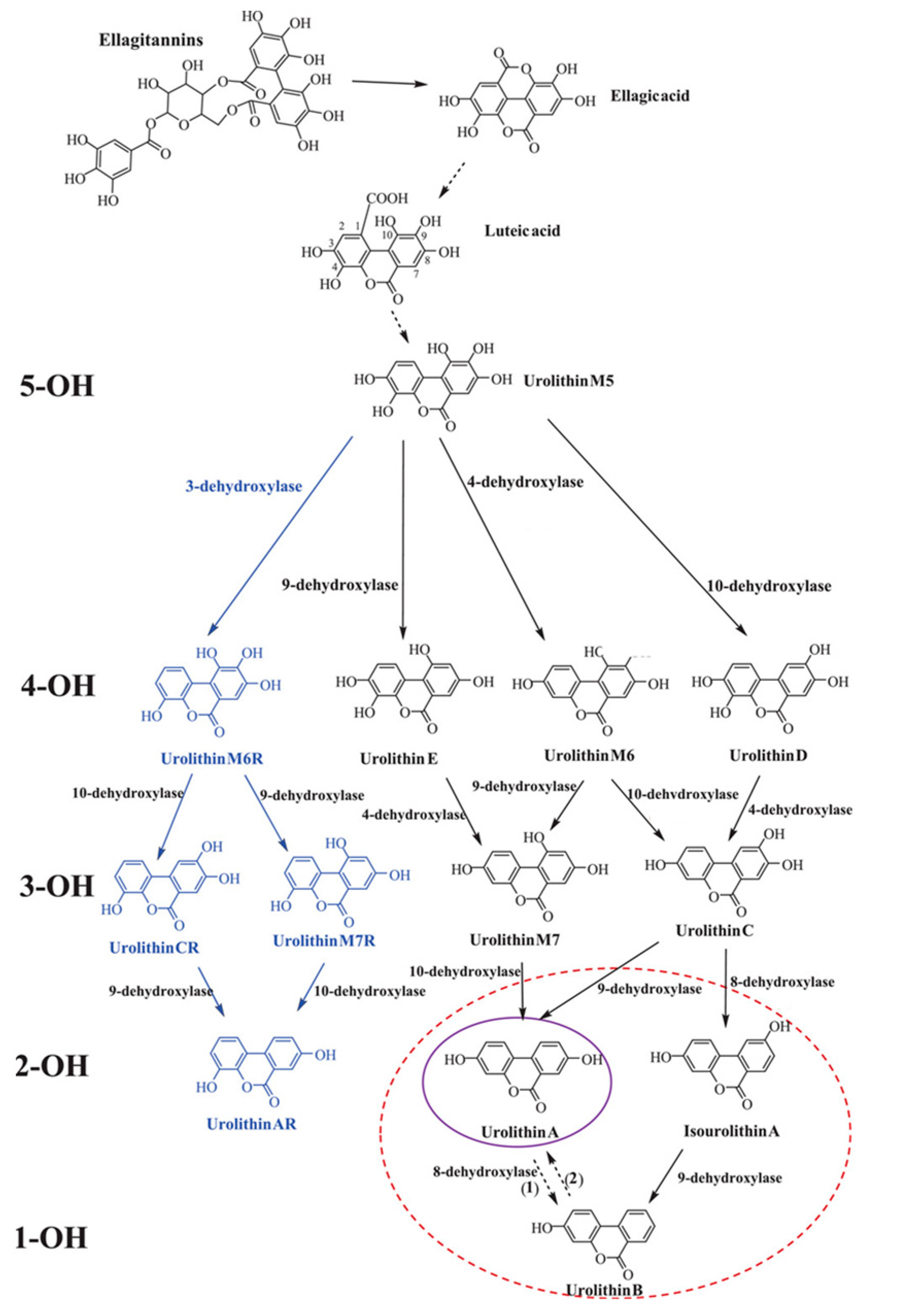
2.1. Urolithins Metabolic Pathways
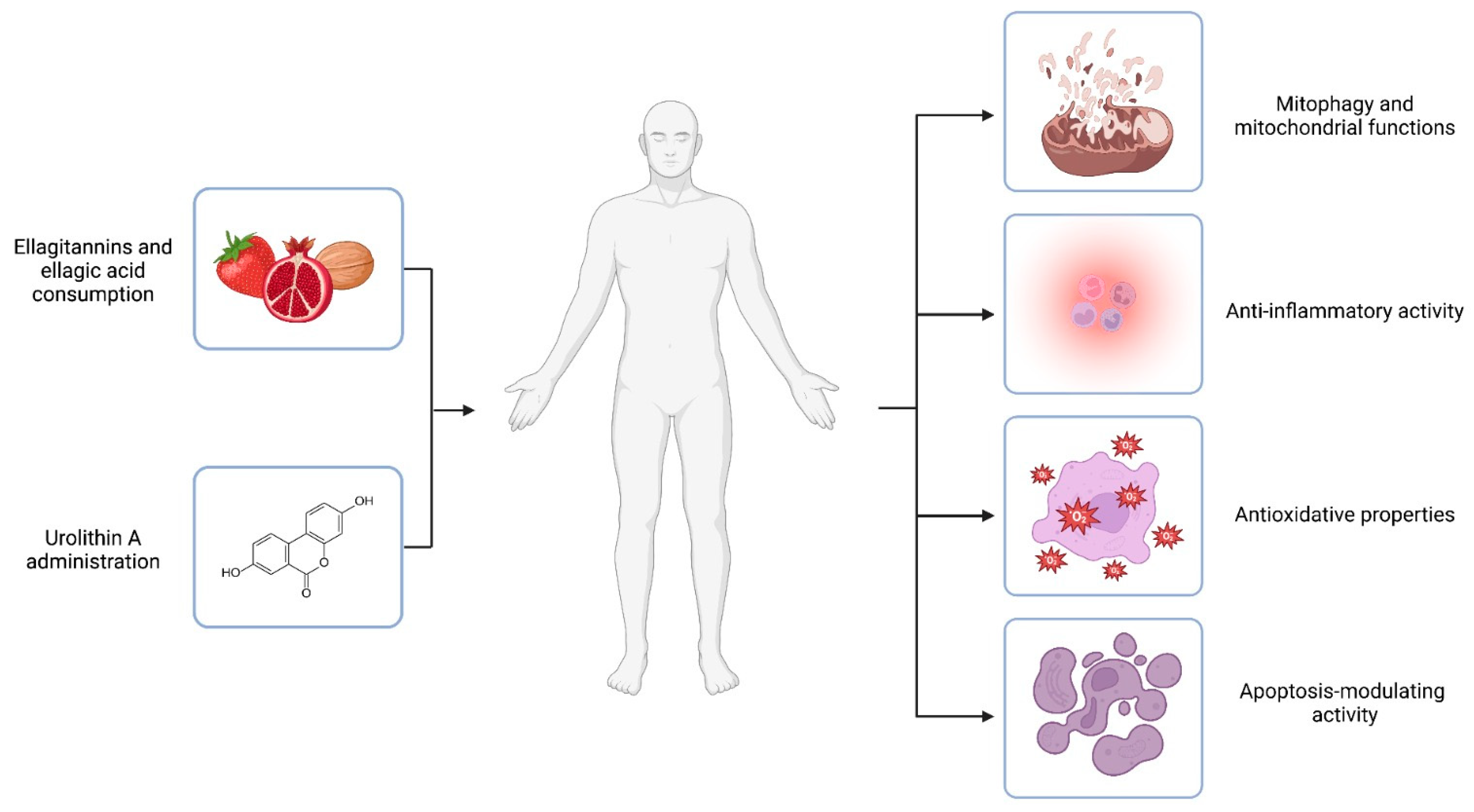
2.2. Mechanism of Action
2.2.1. Mitophagy and Mitochondrial Functions
2.2.2. Anti-Inflammatory Activity
2.2.3. Antioxidant Activity
2.2.4. Apoptosis-Modulating Activity
3. Urolithin A in Health and Diseases
4. Urolithin A and the CNS
4.1. Brain Health
4.2. AD and Brain Injury
4.3. Parkinson’s Disease
5. Summary and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, G.; Liu, J. Autonomic dysfunction in Parkinson’s disease: Implications for pathophysiology, diagnosis, and treatment. Neurobiol. Dis. 2020, 134, 104700. [Google Scholar] [CrossRef] [PubMed]
- Kujawska, M.; Jodynis-Liebert, J. What is the Evidence That Parkinson’s Disease is a Prion Disorder, Which Originates in the Gut? Int. J. Mol. Sci. 2018, 19, 3573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, M.J.; Okun, M.S. Diagnosis and Treatment of Parkinson Disease: A Review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef]
- Kujawska, M.; Jodynis-Liebert, J. Polyphenols in Parkinson’s Disease: A Systematic Review of In Vivo Studies. Nutrients 2018, 10, 642. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Singh, A.K.; Kumar, R.; Jamieson, S.; Pandey, A.K.; Bishayee, A. Neuroprotective Potential of Ellagic Acid: A Critical Review. Adv. Nutr. 2021, 12, 1211–1238. [Google Scholar] [CrossRef]
- Garcia-Villalba, R.; Gimenez-Bastida, J.A.; Cortes-Martin, A.; Avila-Galvez, M.A.; Tomas-Barberan, F.A.; Selma, M.V.; Espin, J.C.; Gonzalez-Sarrias, A. Urolithins: A Comprehensive Update on their Metabolism, Bioactivity, and Associated Gut Microbiota. Mol. Nutr. Food Res. 2022, 66, e2101019. [Google Scholar] [CrossRef]
- D’Amico, D.; Andreux, P.A.; Valdes, P.; Singh, A.; Rinsch, C.; Auwerx, J. Impact of the Natural Compound Urolithin A on Health, Disease, and Aging. Trends Mol. Med. 2021, 27, 687–699. [Google Scholar] [CrossRef]
- Denk, D.; Petrocelli, V.; Conche, C.; Drachsler, M.; Ziegler, P.K.; Braun, A.; Kress, A.; Nicolas, A.M.; Mohs, K.; Becker, C.; et al. Expansion of T memory stem cells with superior anti-tumor immunity by Urolithin A-induced mitophagy. Immunity 2022, 55, 2059–2073.e2058. [Google Scholar] [CrossRef]
- Ghosh, S.; Moorthy, B.; Haribabu, B.; Jala, V.R. Cytochrome P450 1A1 is essential for the microbial metabolite, Urolithin A-mediated protection against colitis. Front. Immunol. 2022, 13, 1004603. [Google Scholar] [CrossRef]
- Singh, A.; D’Amico, D.; Andreux, P.A.; Fouassier, A.M.; Blanco-Bose, W.; Evans, M.; Aebischer, P.; Auwerx, J.; Rinsch, C. Urolithin A improves muscle strength, exercise performance, and biomarkers of mitochondrial health in a randomized trial in middle-aged adults. Cell Rep. Med. 2022, 3, 100633. [Google Scholar] [CrossRef]
- Djedjibegovic, J.; Marjanovic, A.; Panieri, E.; Saso, L. Ellagic Acid-Derived Urolithins as Modulators of Oxidative Stress. Oxid. Med. Cell Longev. 2020, 2020, 5194508. [Google Scholar] [CrossRef]
- Kujawska, M.; Jodynis-Liebert, J. Potential of the ellagic acid-derived gut microbiota metabolite—Urolithin A in gastrointestinal protection. World J. Gastroenterol. 2020, 26, 3170–3181. [Google Scholar] [CrossRef]
- Ahsan, A.; Zheng, Y.R.; Wu, X.L.; Tang, W.D.; Liu, M.R.; Ma, S.J.; Jiang, L.; Hu, W.W.; Zhang, X.N.; Chen, Z. Urolithin A-activated autophagy but not mitophagy protects against ischemic neuronal injury by inhibiting ER stress in vitro and in vivo. CNS Neurosci. Ther. 2019, 25, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Fang, E.F.; Hou, Y.; Palikaras, K.; Adriaanse, B.A.; Kerr, J.S.; Yang, B.; Lautrup, S.; Hasan-Olive, M.M.; Caponio, D.; Dan, X.; et al. Mitophagy inhibits amyloid-beta and tau pathology and reverses cognitive deficits in models of Alzheimer’s disease. Nat. Neurosci. 2019, 22, 401–412. [Google Scholar] [CrossRef]
- Gong, Z.; Huang, J.; Xu, B.; Ou, Z.; Zhang, L.; Lin, X.; Ye, X.; Kong, X.; Long, D.; Sun, X.; et al. Urolithin A attenuates memory impairment and neuroinflammation in APP/PS1 mice. J. Neuroinflammation 2019, 16, 62. [Google Scholar] [CrossRef] [Green Version]
- Busto, R.; Serna, J.; Perianes-Cachero, A.; Quintana-Portillo, R.; Garcia-Seisdedos, D.; Canfran-Duque, A.; Paino, C.L.; Lerma, M.; Casado, M.E.; Martin-Hidalgo, A.; et al. Ellagic acid protects from myelin-associated sphingolipid loss in experimental autoimmune encephalomyelitis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 958–967. [Google Scholar] [CrossRef]
- Senobari, Z.; Karimi, G.; Jamialahmadi, K. Ellagitannins, promising pharmacological agents for the treatment of cancer stem cells. Phytother. Res. 2022, 36, 231–242. [Google Scholar] [CrossRef]
- Zary-Sikorska, E.; Fotschki, B.; Jurgonski, A.; Kosmala, M.; Milala, J.; Kolodziejczyk, K.; Majewski, M.; Ognik, K.; Juskiewicz, J. Protective Effects of a Strawberry Ellagitannin-Rich Extract against Pro-Oxidative and Pro-Inflammatory Dysfunctions Induced by a High-Fat Diet in a Rat Model. Molecules 2020, 25, 5874. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef]
- Garcia-Villalba, R.; Tomas-Barberan, F.A.; Iglesias-Aguirre, C.E.; Gimenez-Bastida, J.A.; Gonzalez-Sarrias, A.; Selma, M.V.; Espin, J.C. Ellagitannins, urolithins, and neuroprotection: Human evidence and the possible link to the gut microbiota. Mol. Aspects Med. 2023, 89, 101109. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villalba, R.; Selma, M.V.; Espin, J.C.; Tomas-Barberan, F.A. Identification of Novel Urolithin Metabolites in Human Feces and Urine after the Intake of a Pomegranate Extract. J. Agric. Food Chem. 2019, 67, 11099–11107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobowska, A.; Granica, S.; Filipek, A.; Melzig, M.F.; Moeslinger, T.; Zentek, J.; Kruk, A.; Piwowarski, J.P. Comparative studies of urolithins and their phase II metabolites on macrophage and neutrophil functions. Eur. J. Nutr. 2021, 60, 1957–1972. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Martin, A.; Garcia-Villalba, R.; Garcia-Mantrana, I.; Rodriguez-Varela, A.; Romo-Vaquero, M.; Collado, M.C.; Tomas-Barberan, F.A.; Espin, J.C.; Selma, M.V. Urolithins in Human Breast Milk after Walnut Intake and Kinetics of Gordonibacter Colonization in Newly Born: The Role of Mothers’ Urolithin Metabotypes. J. Agric. Food Chem. 2020, 68, 12606–12616. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Thomas-Ahner, J.M.; Riedl, K.M.; Bailey, M.T.; Vodovotz, Y.; Schwartz, S.J.; Clinton, S.K. Dietary Black Raspberries Impact the Colonic Microbiome and Phytochemical Metabolites in Mice. Mol. Nutr. Food Res. 2019, 63, e1800636. [Google Scholar] [CrossRef]
- Kujawska, M.; Jourdes, M.; Kurpik, M.; Szulc, M.; Szaefer, H.; Chmielarz, P.; Kreiner, G.; Krajka-Kuzniak, V.; Mikolajczak, P.L.; Teissedre, P.L.; et al. Neuroprotective Effects of Pomegranate Juice against Parkinson’s Disease and Presence of Ellagitannins-Derived Metabolite-Urolithin A-In the Brain. Int. J. Mol. Sci. 2019, 21, 202. [Google Scholar] [CrossRef] [Green Version]
- Toney, A.M.; Fan, R.; Xian, Y.; Chaidez, V.; Ramer-Tait, A.E.; Chung, S. Urolithin A, a Gut Metabolite, Improves Insulin Sensitivity Through Augmentation of Mitochondrial Function and Biogenesis. Obesity (Silver Spring) 2019, 27, 612–620. [Google Scholar] [CrossRef]
- Heilman, J.; Andreux, P.; Tran, N.; Rinsch, C.; Blanco-Bose, W. Safety assessment of Urolithin A, a metabolite produced by the human gut microbiota upon dietary intake of plant derived ellagitannins and ellagic acid. Food Chem. Toxicol. 2017, 108, 289–297. [Google Scholar] [CrossRef]
- Ryu, D.; Mouchiroud, L.; Andreux, P.A.; Katsyuba, E.; Moullan, N.; Nicolet-Dit-Félix, A.A.; Williams, E.G.; Jha, P.; Lo Sasso, G.; Huzard, D.; et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 2016, 22, 879–888. [Google Scholar] [CrossRef]
- Cortes-Martin, A.; Garcia-Villalba, R.; Gonzalez-Sarrias, A.; Romo-Vaquero, M.; Loria-Kohen, V.; Ramirez-de-Molina, A.; Tomas-Barberan, F.A.; Selma, M.V.; Espin, J.C. The gut microbiota urolithin metabotypes revisited: The human metabolism of ellagic acid is mainly determined by aging. Food Funct. 2018, 9, 4100–4106. [Google Scholar] [CrossRef]
- Xian, W.; Yang, S.; Deng, Y.; Yang, Y.; Chen, C.; Li, W.; Yang, R. Distribution of Urolithins Metabotypes in Healthy Chinese Youth: Difference in Gut Microbiota and Predicted Metabolic Pathways. J. Agric. Food Chem. 2021, 69, 13055–13065. [Google Scholar] [CrossRef]
- Singh, A.; D’Amico, D.; Andreux, P.A.; Dunngalvin, G.; Kern, T.; Blanco-Bose, W.; Auwerx, J.; Aebischer, P.; Rinsch, C. Direct supplementation with Urolithin A overcomes limitations of dietary exposure and gut microbiome variability in healthy adults to achieve consistent levels across the population. Eur. J. Clin. Nutr. 2022, 76, 297–308. [Google Scholar] [CrossRef]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Mechanisms of mitophagy in cellular homeostasis, physiology and pathology. Nat. Cell Biol. 2018, 20, 1013–1022. [Google Scholar] [CrossRef]
- Andreux, P.A.; Blanco-Bose, W.; Ryu, D.; Burdet, F.; Ibberson, M.; Aebischer, P.; Auwerx, J.; Singh, A.; Rinsch, C. The mitophagy activator urolithin A is safe and induces a molecular signature of improved mitochondrial and cellular health in humans. Nat. Metab. 2019, 1, 595–603. [Google Scholar] [CrossRef]
- Luan, P.; D’Amico, D.; Andreux, P.A.; Laurila, P.P.; Wohlwend, M.; Li, H.; Imamura de Lima, T.; Place, N.; Rinsch, C.; Zanou, N.; et al. Urolithin A improves muscle function by inducing mitophagy in muscular dystrophy. Sci. Transl. Med. 2021, 13, eabb0319. [Google Scholar] [CrossRef]
- Tuohetaerbaike, B.; Zhang, Y.; Tian, Y.; Zhang, N.N.; Kang, J.; Mao, X.; Zhang, Y.; Li, X. Pancreas protective effects of Urolithin A on type 2 diabetic mice induced by high fat and streptozotocin via regulating autophagy and AKT/mTOR signaling pathway. J. Ethnopharmacol. 2020, 250, 112479. [Google Scholar] [CrossRef]
- Jing, T.; Liao, J.; Shen, K.; Chen, X.; Xu, Z.; Tian, W.; Wang, Y.; Jin, B.; Pan, H. Protective effect of urolithin a on cisplatin-induced nephrotoxicity in mice via modulation of inflammation and oxidative stress. Food Chem. Toxicol. 2019, 129, 108–114. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Hering, N.A.; Luettig, J.; Jebautzke, B.; Schulzke, J.D.; Rosenthal, R. The Punicalagin Metabolites Ellagic Acid and Urolithin A Exert Different Strengthening and Anti-Inflammatory Effects on Tight Junction-Mediated Intestinal Barrier Function In Vitro. Front. Pharmacol. 2021, 12, 610164. [Google Scholar] [CrossRef]
- Shen, P.X.; Li, X.; Deng, S.Y.; Zhao, L.; Zhang, Y.Y.; Deng, X.; Han, B.; Yu, J.; Li, Y.; Wang, Z.Z.; et al. Urolithin A ameliorates experimental autoimmune encephalomyelitis by targeting aryl hydrocarbon receptor. EBioMedicine 2021, 64, 103227. [Google Scholar] [CrossRef]
- Komatsu, W.; Kishi, H.; Yagasaki, K.; Ohhira, S. Urolithin A attenuates pro-inflammatory mediator production by suppressing PI3-K/Akt/NF-kappaB and JNK/AP-1 signaling pathways in lipopolysaccharide-stimulated RAW264 macrophages: Possible involvement of NADPH oxidase-derived reactive oxygen species. Eur. J. Pharmacol. 2018, 833, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Casedas, G.; Les, F.; Choya-Foces, C.; Hugo, M.; Lopez, V. The Metabolite Urolithin-A Ameliorates Oxidative Stress in Neuro-2a Cells, Becoming a Potential Neuroprotective Agent. Antioxidants 2020, 9, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Ma, H.; Frost, L.; Yuan, T.; Dain, J.A.; Seeram, N.P. Pomegranate phenolics inhibit formation of advanced glycation endproducts by scavenging reactive carbonyl species. Food Funct. 2014, 5, 2996–3004. [Google Scholar] [CrossRef] [PubMed]
- Verzelloni, E.; Pellacani, C.; Tagliazucchi, D.; Tagliaferri, S.; Calani, L.; Costa, L.G.; Brighenti, F.; Borges, G.; Crozier, A.; Conte, A.; et al. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol. Nutr. Food Res. 2011, 55 (Suppl. 1), S35–S43. [Google Scholar] [CrossRef]
- Di Stasi, L.C. Natural Coumarin Derivatives Activating Nrf2 Signaling Pathway as Lead Compounds for the Design and Synthesis of Intestinal Anti-Inflammatory Drugs. Pharmaceuticals 2023, 16, 511. [Google Scholar] [CrossRef]
- Lou, L.; Wang, M.; He, J.; Yang, S.; Meng, F.; Wang, S.; Jin, X.; Cai, J.; Cai, C. Urolithin A (UA) attenuates ferroptosis in LPS-induced acute lung injury in mice by upregulating Keap1-Nrf2/HO-1 signaling pathway. Front. Pharmacol. 2023, 14, 1067402. [Google Scholar] [CrossRef]
- Mazumder, M.K.; Choudhury, S.; Borah, A. An in silico investigation on the inhibitory potential of the constituents of Pomegranate juice on antioxidant defense mechanism: Relevance to neurodegenerative diseases. IBRO Rep. 2019, 6, 153–159. [Google Scholar] [CrossRef]
- Zou, D.; Ganugula, R.; Arora, M.; Nabity, M.B.; Sheikh-Hamad, D.; Kumar, M. Oral delivery of nanoparticle urolithin A normalizes cellular stress and improves survival in mouse model of cisplatin-induced AKI. Am. J. Physiol. Renal Physiol. 2019, 317, F1255–F1264. [Google Scholar] [CrossRef]
- Gonzalez-Sarrias, A.; Nunez-Sanchez, M.A.; Tomas-Barberan, F.A.; Espin, J.C. Neuroprotective Effects of Bioavailable Polyphenol-Derived Metabolites against Oxidative Stress-Induced Cytotoxicity in Human Neuroblastoma SH-SY5Y Cells. J. Agric. Food Chem. 2017, 65, 752–758. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.B.; Lee, S.; Kim, J.H. Neuroprotective effects of urolithin A on H(2)O(2)-induced oxidative stress-mediated apoptosis in SK-N-MC cells. Nutr. Res. Pract. 2020, 14, 3–11. [Google Scholar] [CrossRef]
- El-Wetidy, M.S.; Ahmad, R.; Rady, I.; Helal, H.; Rady, M.I.; Vaali-Mohammed, M.A.; Al-Khayal, K.; Traiki, T.B.; Abdulla, M.H. Urolithin A induces cell cycle arrest and apoptosis by inhibiting Bcl-2, increasing p53-p21 proteins and reactive oxygen species production in colorectal cancer cells. Cell Stress. Chaperones 2021, 26, 473–493. [Google Scholar] [CrossRef]
- Mohammed Saleem, Y.I.; Albassam, H.; Selim, M. Urolithin A induces prostate cancer cell death in p53-dependent and in p53-independent manner. Eur. J. Nutr. 2020, 59, 1607–1618. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhou, J.; Zhang, C.; Cheng, Y.; Hu, J.; Zheng, G. Antiproliferative effect of urolithin A, the ellagic acid-derived colonic metabolite, on hepatocellular carcinoma HepG2.2.15 cells by targeting Lin28a/let-7a axis. Braz. J. Med. Biol. Res. 2018, 51, e7220. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, N.; Das, A.; Biswas, N.; Gnyawali, S.; Singh, K.; Gorain, M.; Polcyn, C.; Khanna, S.; Roy, S.; Sen, C.K. Urolithin A augments angiogenic pathways in skeletal muscle by bolstering NAD(+) and SIRT1. Sci. Rep. 2020, 10, 20184. [Google Scholar] [CrossRef]
- Ballesteros-Alvarez, J.; Nguyen, W.; Sivapatham, R.; Rane, A.; Andersen, J.K. Urolithin A reduces amyloid-beta load and improves cognitive deficits uncorrelated with plaque burden in a mouse model of Alzheimer’s disease. Geroscience 2023, 45, 1095–1113. [Google Scholar] [CrossRef]
- Liu, S.; D’Amico, D.; Shankland, E.; Bhayana, S.; Garcia, J.M.; Aebischer, P.; Rinsch, C.; Singh, A.; Marcinek, D.J. Effect of Urolithin A Supplementation on Muscle Endurance and Mitochondrial Health in Older Adults: A Randomized Clinical Trial. JAMA Netw. Open 2022, 5, e2144279. [Google Scholar] [CrossRef]
- Huang, J.R.; Zhang, M.H.; Chen, Y.J.; Sun, Y.L.; Gao, Z.M.; Li, Z.J.; Zhang, G.P.; Qin, Y.; Dai, X.Y.; Yu, X.Y.; et al. Urolithin A ameliorates obesity-induced metabolic cardiomyopathy in mice via mitophagy activation. Acta Pharmacol. Sin. 2023, 44, 321–331. [Google Scholar] [CrossRef]
- Cui, G.H.; Chen, W.Q.; Shen, Z.Y. Urolithin A shows anti-atherosclerotic activity via activation of class B scavenger receptor and activation of Nef2 signaling pathway. Pharmacol. Rep. 2018, 70, 519–524. [Google Scholar] [CrossRef]
- Tang, L.; Mo, Y.; Li, Y.; Zhong, Y.; He, S.; Zhang, Y.; Tang, Y.; Fu, S.; Wang, X.; Chen, A. Urolithin A alleviates myocardial ischemia/reperfusion injury via PI3K/Akt pathway. Biochem. Biophys. Res. Commun. 2017, 486, 774–780. [Google Scholar] [CrossRef]
- Nishimoto, Y.; Fujisawa, K.; Ukawa, Y.; Kudoh, M.; Funahashi, K.; Kishimoto, Y.; Fukuda, S. Effect of urolithin A on the improvement of vascular endothelial function depends on the gut microbiota. Front. Nutr. 2022, 9, 1077534. [Google Scholar] [CrossRef]
- Xia, B.; Shi, X.C.; Xie, B.C.; Zhu, M.Q.; Chen, Y.; Chu, X.Y.; Cai, G.H.; Liu, M.; Yang, S.Z.; Mitchell, G.A.; et al. Urolithin A exerts antiobesity effects through enhancing adipose tissue thermogenesis in mice. PLoS Biol. 2020, 18, e3000688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Guo, Y.; Henning, S.M.; Chan, B.; Long, J.; Zhong, J.; Acin-Perez, R.; Petcherski, A.; Shirihai, O.; Heber, D.; et al. Ellagic Acid and Its Microbial Metabolite Urolithin A Alleviate Diet-Induced Insulin Resistance in Mice. Mol. Nutr. Food Res. 2020, 64, e2000091. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, N.R.; Chandrasekaran, B.; Kolluru, V.; Ankem, M.; Damodaran, C.; Vadhanam, M.V. A natural molecule, urolithin A, downregulates androgen receptor activation and suppresses growth of prostate cancer. Mol. Carcinog. 2018, 57, 1332–1341. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qiu, Z.; Zhou, B.; Liu, C.; Ruan, J.; Yan, Q.; Liao, J.; Zhu, F. In vitro antiproliferative and antioxidant effects of urolithin A, the colonic metabolite of ellagic acid, on hepatocellular carcinomas HepG2 cells. Toxicol. In Vitro 2015, 29, 1107–1115. [Google Scholar] [CrossRef]
- Cheng, F.; Dou, J.; Zhang, Y.; Wang, X.; Wei, H.; Zhang, Z.; Cao, Y.; Wu, Z. Urolithin A Inhibits Epithelial-Mesenchymal Transition in Lung Cancer Cells via P53-Mdm2-Snail Pathway. Onco Targets Ther. 2021, 14, 3199–3208. [Google Scholar] [CrossRef]
- Gimenez-Bastida, J.A.; Avila-Galvez, M.A.; Espin, J.C.; Gonzalez-Sarrias, A. The gut microbiota metabolite urolithin A, but not other relevant urolithins, induces p53-dependent cellular senescence in human colon cancer cells. Food Chem. Toxicol. 2020, 139, 111260. [Google Scholar] [CrossRef]
- Norden, E.; Heiss, E.H. Urolithin A gains in antiproliferative capacity by reducing the glycolytic potential via the p53/TIGAR axis in colon cancer cells. Carcinogenesis 2019, 40, 93–101. [Google Scholar] [CrossRef]
- Zhao, W.; Shi, F.; Guo, Z.; Zhao, J.; Song, X.; Yang, H. Metabolite of ellagitannins, urolithin A induces autophagy and inhibits metastasis in human sw620 colorectal cancer cells. Mol. Carcinog. 2018, 57, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Kshirsagar, S.; Alvir, R.V.; Pradeepkiran, J.A.; Hindle, A.; Vijayan, M.; Ramasubramaniam, B.; Kumar, S.; Reddy, A.P.; Reddy, P.H. A Combination Therapy of Urolithin A+EGCG Has Stronger Protective Effects than Single Drug Urolithin A in a Humanized Amyloid Beta Knockin Mice for Late-Onset Alzheimer’s Disease. Cells 2022, 11, 2660. [Google Scholar] [CrossRef]
- Gong, Q.Y.; Cai, L.; Jing, Y.; Wang, W.; Yang, D.X.; Chen, S.W.; Tian, H.L. Urolithin A alleviates blood-brain barrier disruption and attenuates neuronal apoptosis following traumatic brain injury in mice. Neural Regen. Res. 2022, 17, 2007–2013. [Google Scholar] [CrossRef]
- Lin, X.H.; Ye, X.J.; Li, Q.F.; Gong, Z.; Cao, X.; Li, J.H.; Zhao, S.T.; Sun, X.D.; He, X.S.; Xuan, A.G. Urolithin A Prevents Focal Cerebral Ischemic Injury via Attenuating Apoptosis and Neuroinflammation in Mice. Neuroscience 2020, 448, 94–106. [Google Scholar] [CrossRef]
- Qiu, J.; Chen, Y.; Zhuo, J.; Zhang, L.; Liu, J.; Wang, B.; Sun, D.; Yu, S.; Lou, H. Urolithin A promotes mitophagy and suppresses NLRP3 inflammasome activation in lipopolysaccharide-induced BV2 microglial cells and MPTP-induced Parkinson’s disease model. Neuropharmacology 2022, 207, 108963. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, J.; Qiu, J.; Wang, L.; Zhuo, J.; Wang, B.; Sun, D.; Yu, S.; Lou, H. Urolithin A protects dopaminergic neurons in experimental models of Parkinson’s disease by promoting mitochondrial biogenesis through the SIRT1/PGC-1alpha signaling pathway. Food Funct. 2022, 13, 375–385. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Vemula, P.K.; Haribabu, B.; Jala, V.R. Microbial Metabolite Urolithin B Inhibits Recombinant Human Monoamine Oxidase A Enzyme. Metabolites 2020, 10, 258. [Google Scholar] [CrossRef]
- Cortes-Martin, A.; Selma, M.V.; Tomas-Barberan, F.A.; Gonzalez-Sarrias, A.; Espin, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, e1900952. [Google Scholar] [CrossRef]
- Avila-Galvez, M.A.; Gonzalez-Sarrias, A.; Espin, J.C. In Vitro Research on Dietary Polyphenols and Health: A Call of Caution and a Guide on How To Proceed. J. Agric. Food Chem. 2018, 66, 7857–7858. [Google Scholar] [CrossRef]
- Bookheimer, S.Y.; Renner, B.A.; Ekstrom, A.; Li, Z.; Henning, S.M.; Brown, J.A.; Jones, M.; Moody, T.; Small, G.W. Pomegranate juice augments memory and FMRI activity in middle-aged and older adults with mild memory complaints. Evid. Based Complement. Altern. Med. 2013, 2013, 946298. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, A.; Zelicha, H.; Yaskolka Meir, A.; Rinott, E.; Tsaban, G.; Levakov, G.; Prager, O.; Salti, M.; Yovell, Y.; Ofer, J.; et al. The effect of a high-polyphenol Mediterranean diet (Green-MED) combined with physical activity on age-related brain atrophy: The Dietary Intervention Randomized Controlled Trial Polyphenols Unprocessed Study (DIRECT PLUS). Am. J. Clin. Nutr. 2022, 115, 1270–1281. [Google Scholar] [CrossRef]
- Kujawska, M.; Jourdes, M.; Witucki, L.; Karazniewicz-Lada, M.; Szulc, M.; Gorska, A.; Mikolajczak, P.L.; Teissedre, P.L.; Jodynis-Liebert, J. Pomegranate Juice Ameliorates Dopamine Release and Behavioral Deficits in a Rat Model of Parkinson’s Disease. Brain Sci. 2021, 11, 1127. [Google Scholar] [CrossRef]
- Tan, S.; Tong, W.H.; Vyas, A. Urolithin-A attenuates neurotoxoplasmosis and alters innate response towards predator odor. Brain Behav. Immun. Health 2020, 8, 100128. [Google Scholar] [CrossRef]
- Jayatunga, D.P.W.; Hone, E.; Khaira, H.; Lunelli, T.; Singh, H.; Guillemin, G.J.; Fernando, B.; Garg, M.L.; Verdile, G.; Martins, R.N. Therapeutic Potential of Mitophagy-Inducing Microflora Metabolite, Urolithin A for Alzheimer’s Disease. Nutrients 2021, 13, 3744. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Fellner, A.; Kumar, K.R. Monogenic Parkinson’s Disease: Genotype, Phenotype, Pathophysiology, and Genetic Testing. Genes 2022, 13, 471. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.K.; Tanner, C.M.; Brundin, P. Parkinson Disease Epidemiology, Pathology, Genetics, and Pathophysiology. Clin. Geriatr. Med. 2020, 36, 1–12. [Google Scholar] [CrossRef]
- Yoo, S.M.; Jung, Y.K. A Molecular Approach to Mitophagy and Mitochondrial Dynamics. Mol. Cells 2018, 41, 18–26. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [Green Version]
- Gordon, R.; Albornoz, E.A.; Christie, D.C.; Langley, M.R.; Kumar, V.; Mantovani, S.; Robertson, A.A.B.; Butler, M.S.; Rowe, D.B.; O’Neill, L.A.; et al. Inflammasome inhibition prevents alpha-synuclein pathology and dopaminergic neurodegeneration in mice. Sci. Transl. Med. 2018, 10, eaah4066. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Xu, T.; Fang, Q.; Zhang, H.; Yue, L.; Hu, G.; Sun, L. Quercetin hinders microglial activation to alleviate neurotoxicity via the interplay between NLRP3 inflammasome and mitophagy. Redox Biol. 2021, 44, 102010. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Li, Y.; Yu, S.; Zhao, Y. SIRT1/PGC-1alpha Signaling Promotes Mitochondrial Functional Recovery and Reduces Apoptosis after Intracerebral Hemorrhage in Rats. Front. Mol. Neurosci. 2017, 10, 443. [Google Scholar] [CrossRef] [Green Version]
- Mudo, G.; Makela, J.; Di Liberto, V.; Tselykh, T.V.; Olivieri, M.; Piepponen, P.; Eriksson, O.; Malkia, A.; Bonomo, A.; Kairisalo, M.; et al. Transgenic expression and activation of PGC-1alpha protect dopaminergic neurons in the MPTP mouse model of Parkinson’s disease. Cell Mol. Life Sci. 2012, 69, 1153–1165. [Google Scholar] [CrossRef]
- Ng, J.H.Y.; Andersen, J. Urolithin A: Gut-Brain Dietary Intervention in Parkinson’s Disease. Innov. Aging 2022, 6, 729. [Google Scholar] [CrossRef]
- Rezaee, S.; Jahromy, M.H. Potential Effects of Pomegranate Juice in Attenuating LID in Mice Model of Parkinson Disease. Pharmacogn. J. 2018, 10, 700–704. [Google Scholar] [CrossRef] [Green Version]
- Bar-Ya’akov, I.; Tian, L.; Amir, R.; Holland, D. Primary Metabolites, Anthocyanins, and Hydrolyzable Tannins in the Pomegranate Fruit. Front. Plant Sci. 2019, 10, 620. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Mula, H.M.; Tomas-Barberan, F.A.; Garcia-Villalba, R. Pomegranate Fruit and Juice (cv. Mollar), Rich in Ellagitannins and Anthocyanins, Also Provide a Significant Content of a Wide Range of Proanthocyanidins. J. Agric. Food Chem. 2019, 67, 9160–9167. [Google Scholar] [CrossRef]
- Bellesia, A.; Verzelloni, E.; Tagliazucchi, D. Pomegranate ellagitannins inhibit alpha-glucosidase activity in vitro and reduce starch digestibility under simulated gastro-intestinal conditions. Int. J. Food Sci. Nutr. 2015, 66, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Aichinger, G.; Stevanoska, M.; Beekmann, K.; Sturla, S.J. Physiologically-based pharmacokinetic modeling of the postbiotic supplement urolithin A predicts its bioavailability is orders of magnitudes lower than concentrations that induce toxicity, but also neuroprotective effects. Mol. Nutr. Food Res. 2023, e2300009. [Google Scholar] [CrossRef]
- Yuan, T.; Ma, H.; Liu, W.; Niesen, D.B.; Shah, N.; Crews, R.; Rose, K.N.; Vattem, D.A.; Seeram, N.P. Pomegranate’s Neuroprotective Effects against Alzheimer’s Disease Are Mediated by Urolithins, Its Ellagitannin-Gut Microbial Derived Metabolites. ACS Chem. Neurosci. 2016, 7, 26–33. [Google Scholar] [CrossRef]
- Emami Kazemabad, M.J.; Asgari Toni, S.; Tizro, N.; Dadkhah, P.A.; Amani, H.; Akhavan Rezayat, S.; Sheikh, Z.; Mohammadi, M.; Alijanzadeh, D.; Alimohammadi, F.; et al. Pharmacotherapeutic potential of pomegranate in age-related neurological disorders. Front. Aging Neurosci. 2022, 14, 955735. [Google Scholar] [CrossRef]
- Zaitone, S.A.; Abo-Elmatty, D.M.; Shaalan, A.A. Acetyl-L-carnitine and α-lipoic acid affect rotenone-induced damage in nigral dopaminergic neurons of rat brain, implication for Parkinson’s disease therapy. Pharmacol. Biochem. Behav. 2012, 100, 347–360. [Google Scholar] [CrossRef]
- Khan, E.; Hasan, I.; Haque, M.E. Parkinson’s Disease: Exploring Different Animal Model Systems. Int. J. Mol. Sci. 2023, 24, 9088. [Google Scholar] [CrossRef]
- Guérin, T.; Waterlot, C.; Lipka, E.; Gervois, P.; Bulteel, D.; Betrancourt, D.; Moignard, C.; Nica, A.S.; Furman, C.; Ghinet, A. Ecocatalysed Hurtley reaction: Synthesis of urolithin derivatives as new potential RAGE antagonists with anti-ageing properties. Sustain. Chem. Pharm. 2021, 23, 100518. [Google Scholar] [CrossRef]
- Siderowf, A.; Concha-Marambio, L.; Lafontant, D.E.; Farris, C.M.; Ma, Y.; Urenia, P.A.; Nguyen, H.; Alcalay, R.N.; Chahine, L.M.; Foroud, T.; et al. Assessment of heterogeneity among participants in the Parkinson’s Progression Markers Initiative cohort using alpha-synuclein seed amplification: A cross-sectional study. Lancet Neurol. 2023, 22, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.K.; Chaudhry, S.K.; Singh, H.B.; Gupta, V.K.; Kaushik, A. Perspectives on nano-nutraceuticals to manage pre and post COVID-19 infections. Biotechnol. Rep. 2022, 33, e00712. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Aadil, K.R.; Mondal, K.; Mishra, Y.K.; Oupicky, D.; Ramakrishna, S.; Kaushik, A. Neurodegenerative disorders management: State-of-art and prospects of nano-biotechnology. Crit. Rev. Biotechnol. 2022, 42, 1180–1212. [Google Scholar] [CrossRef]
- Romo-Vaquero, M.; Fernández-Villalba, E.; Gil-Martinez, A.-L.; Cuenca-Bermejo, L.; Espín, J.C.; Herrero, M.T.; Selma, M.V. Urolithins: Potential biomarkers of gut dysbiosis and disease stage in Parkinson’s patients. Food Funct. 2022, 13, 6306–6316. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dose and Route of Administration | Experimental Model | Outcomes | References |
|---|---|---|---|
| Mitophagy and mitochondrial functions | |||
| 50 µM | SH-SY5Y cells | ↑ F-PINK-1, Parkin, Beclin-1, Bcl2L13, AMBRA1, and p-ULK1(Ser555) | [15] |
| 200 mg/kg/d, i.g., for 2 months | 6-month-old APP/PS1 mice | ↑ relative neuronal mitophagy level ↑ mitophagy events ↓ damaged mitochondria ↑ F-PINK-1 in brain tissue | [15] |
| 200 mg/kg/d, i.g., for 2 months | 13-month-old 3 × TgAD mice | ↑ relative neuronal mitophagy level ↑ mitophagy events in brain tissue | [15] |
| 50 µM, ad libitum, for 10 days | Caenorhabditis elegans | ↑ mRNA of the autophagy genes Bec-1, Sqst-1 and Vps-34 ↑ mRNA of the mitophagy genes Pink-1, Dct-1 ↑ mRNA of the mitophagy and biogenesis gene Skn-1 in muscle tissue GFP–LGG-1-positive punctae | [29] |
| 50 µM | C2C12 myoblasts Mode-K intestinal cells | ↑ LC3-II/LC3-I, p-AMPKα ↑ SQSTM-1, Ub in mitochonfrial fraction ↑ autophagosomes ↑ autolysosomes ↓ mt DNA/nDNA ↑ mt chain subunits ↑ CII-driven respiration | [29] |
| 50 mg/kg/d, p.o., for 34 weeks | 16-month-old C57BL/6J mice | ↑ LC3-II/LC3-I ↓ SQSTM-1 ↑ mRNA of autophagy genes Becn1, Ulk1, Pik3c3, Atg8l, p62, Atg5, Atg7, Atg12, Lc3b, LAMP2 ↑ mRNA of mitophagy gene Park2 ↑ p-AMPK ↑ Ub/SDHA, Ub/VDAC in muscle tissue | [29] |
| 50 mg/kg/d, p.o., for 10 weeks | Caenorhabditis elegans/DMD model | ↑ mRNA of mitophagy genes Pink1, Pdr-1, Dct-1 ↑mRNA of autophagy genes Bec-1, Vsp-34 ↑ mitochondrial network ↑ mitochondrial respiration ↑ citrate synthase activity ↑ mtDNA/nDNA in mucle tissue | [35] |
| 50 mg/kg/d, p.o., for 10 weeks | 13-week-old mdx mice/DMD model | ↑ mRNA of mitophagy genes Pink1, Park2, Park7, and Bnip3 ↑ mRNA of autophagy genes Sqstm-1 and Becn1 ↑ p-S65-Ub, BNIP3, PARKIN, VDAC ↑ mt LC3-II in mucle tissue | [35] |
| 25 µM | Primary myoblast cells derived from DMD patients | ↑ mRNA of mitophagy genes PINK1, PARK2, PARK7, and BNIP3 ↑ mRNA of autophagy genes SQSTM-1, BECN1 | [35] |
| 50 mg/kg/d, i.g., for 8 weeks | 10-week-old C57BL/6 mice/STZ-induced model of type 2 diabetes | ↑ LC3II/I, beclin1, ATG5, ↓ SQSTM-1 ↑ p-AKT, mTORC1 ↓ mitochondrial swelling in pancreatic tissue | [36] |
| Anti-inflammatory activity | |||
| 20 mg/kg, p.o., 10-day post-treatment | C57BL/6 mice/DSS-induced acute colitis | ↓ IL-6, IL-1β, and TNF-α in serum ↓ MPO activity ↑ ZO-1, Ocln, Cldn4 ↓ F4/80+ CD11b+, CD4+ cells ↑ CD11c+, T-reg cells in the mesenteric lymph node | [10] |
| 200 mg/kg/d, i.g., for 2 months | 6-month-old APP/PS1 AD mice | ↓ TNF-α, IL-6 ↑ IL-10 in brain tissue ↓ TNF-α, IL-6 ↑ IL-10 in microglia | [15] |
| 300 mg/kg/d, p.o., for 14 days | 28-week-old APP/PS1 AD mice | ↓ mRNA of Tnfα, Il6 and Il1β ↓ TNF-α, IL-6 and IL-1β ↓ IBA1 and GFAP in brain tissue | [16] |
| 20 μg/d, i.p., for 12 weeks | C57BL/6 mice/HFD | ↓ Il1β mRNA ↓ p-eIF2α, p-ERK ↑ IκBα LC3I/II in liver tissue ↓ M1 polarization (mRNA of Cd11c, Tnfα, Il6, Il1β, and Mcp1) ↑ M2 polarization (mRNA of Ch3l3 and Mgl2) in peritoneal macrophages | [27] |
| 50 mg/kg/d, i.g., for 8 weeks | 10-week-old C57BL/6 mice/STZ-induced model of type 2 diabetes | ↓ IL-1β, TNF-α ↑ IL-10 in plasma | [36] |
| 100 mg/kg/d, i.p., for 5 days | C57BL/6 mice, cisplatin-induced nephrotoxicity model | ↓ CD11b positive monocyte/macrophage ↓ mRNA of Tnfα, Il23, Il18, Mip2 in kidney tissue | [37] |
| 20 mg/kg/d, p.o., on 4th and 6th day of DSS cycle | C57BL/6 mice/DSS-, TNBS-induced colitis | ↓ IL-6, IL-1β, and TNF-α in serum ↓ MPO activity ↓ Cldn4 in colon tissue | [38] |
| 150 and 250 μM | Caco-2 and HT-29/B6/TNFα-induced barrier loss models | ↑ TER ↓ claudin-2 | [39] |
| 25 mg/kg/d, p.o., 20 days | C57BL/6 mice/EAE model | ↓ GFP+ cells in the brain and spinal cord ↓ M1-type microglia ↓ CD11c+ cells infiltrated into CNS ↓ CD45high CD11b+, CD45low CD11b+, CD11b+ MHCII+, CD11b+ TNF-α+, CD11b+ CD16/32+ cells | [40] |
| 10, 40 μM, 2 h pretreatment | LPS-stimulated RAW264 macrophages | ↓ TNF-α, IL-6, NO−, iNOS ↓ intracellular peroxides ↓ NADPH oxidase activity ↓ DNA binding activity of NF-κB and AP-1 ↓ NF-κB (p65) ↑ IκBα ↓ c-Jun, p-c-Jun, p-Akt, p-JNK, p-p38 | [41] |
| Antioxidant activity | |||
| 20 μg/d, i.p., for 12 weeks | C57BL/6 mice/HFD | ↑ mRNA of Sod1 and Sod2 in liver tissue | [27] |
| 50 mg/kg/d, i.g., for 8 weeks | 10-week-old C57BL/6 mice/STZ-induced model of type 2 diabetes | ↑ GSH ↓ MDA in pancreas tissue | [36] |
| 100 mg/kg/d, i.p., for 5 days | C57BL/6 mice, cisplatin-induced nephrotoxicity model | ↑ CAT, GPx, SOD activity ↑ GSH ↓ GSSG, HNE, protein nitration, DNA fragmentation, Nox2 mRNA in kidney tissue | [37] |
| 0.5, 1, 2, 4 μM | N2a cells/H2O2 | ↑ CAT, SOD, GR, GPx activity ↓ ROS production, TBARS | [42] |
| Apoptosis-modulating activity | |||
| 300 mg/kg/d, i.g., for 14 days | 28-week-old APP/PS1 AD mice | ↑ NeuN+ cells ↓ TUNEL+ cells in brain tissue | [16] |
| 100 mg/kg/d, i.p., for 5 days | C57BL/6 mice, cisplatin-induced nephrotoxicity model | ↓ caspase 3 activity ↓ DNA fragmentation in kidney tissue | [37] |
| 50 mg/kg, i.g., 3 times/week up to 19 days | C57BL/6J mice/cisplatin-induced AKI | ↓ PARP1, TUNEL+ cells ↑ Bcl-2 in kidney tissue | [48] |
| 10 μM, 6 h pretreatment | SH-SY5Y cells/H2O2 | ↓ apoptotic cells ↓ caspase-3, -9 | [49] |
| 1.25, 2.5, 5 μM, 6 h pretreatment | SK-N-MC cells/H2O2 | ↓ Bax/Bcl-2 ↓ cytochrome c, caspase-3, -9 ↓ PARP | [50] |
| 25, 50, 100 μM | HT29, SW480, SW620 cells | ↑ apoptotic cells ↑ ytochrome c, caspase-3, -9 ↑ p53, p21, XIAP ↓ Bcl-2 G2/M phase arrest | [51] |
| Treated with 40, 80 μM | LNCaP, 22RV1, PC3 cells | ↑ p53, p21, MDM2 ↑ PUMA and NOXA | [52] |
| 80 μM | HepG2.2.15 cells | ↑ caspase-3 ↓ Bax/Bcl-2 | [53] |
| Dose and Route of Administration | Experimental Model | Outcomes | References |
|---|---|---|---|
| Longevity | |||
| 500 mg or 1000 mg/d, p.o. for 4 months | Middle-aged adults between 40 to 65 years old | ↑muscle strength, aerobic-endurance, physical performance ↓ acylcarnitines ↓ CRP, IL-1β, TNF-α, IFNγ in plasma | [11] |
| 50 µM, ad libitum, for 10 days | Caenorhabditis elegans | ↑ lifespan, mobility, ↑ pharyngeal pumping ↑ mitochondrial respiratory capacity ↓ dysfunctional mitochondria in muscle tissue | [29] |
| 50 mg/kg/d, p.o., for 10 weeks | 13-week-old mdx mice DMD model | ↑ improved muscle morphology ↑ motility ↓ muscle fiber degeneration ↑ eMyHC, dystrophin in muscle tissue | [35] |
| 10 mg/kg/d, p.o., for 16 weeks | C57BL/6 mice | ↑muscle strength, mobility, and exercise performance | [54] |
| 1000 mg/d, p.o., for 4 months | Adults aged 65 to 90 years | ↑ muscle endurance, physical performance ↓ acylcarnitines, ceramides ↓ CRP in plasma | [56] |
| 5 mg/kg/d, p.o., 10 months in alternate weeks (1 week on, 1 week off) | Female B6129SF2/J and male C57BL/6NJ/aging model | ↑ lifespan | [55] |
| Cardiovascular health | |||
| 50 mg/kg/d, p.o., for 20 weeks | C57BL/6 mice/HFD | ↑ PINK1/Parkin-dependent mitophagy ↓ mitochondrial defects ↑ cardiac diastolic function | [57] |
| 3 mg/kg/d, p.o., for 3 weeks | Wistar rats/HFD + ballon injury in the aorta | ↓ TC, TG and LDL ↓ Ang II ↓ aortic edema | [58] |
| 1 mg/kg, i.p., 24 h and 1 h pretreatment | C57BL/6 mice/H/R | ↓ myocardial infarct sizes ↓ TUNEL+ cells ↑ ejection fraction, fractional shortening ↓ CK, LDH in serum | [59] |
| 10 mg/d for 12 weeks | Healthy volunteers 40–65 years old, UA non-producer or low producer | ↑ FMD score | [60] |
| Metabolic dysfunctions | |||
| 20 μg/d, i.p., for 12 weeks | C57BL/6 mice/HFD | ↓ hepatic TG ↓ IR, adipocyte hypertrophy ↓ macrophage infiltration into the adipose tissue ↓ M1/M2 | [27] |
| 50 mg/kg/d, i.g., for 8 weeks | 10-week-old C57BL/6 mice/STZ-induced model of type 2 diabetes | ↓ BW ↓ FBG, GHb ↓ pancreatic histopathological damages ↑ HOMA-β | [36] |
| 30 mg/kg/d, i.g., for 10 weeks | C57BL/6 mice/HFD | ↓ BW, fat mass ↓ plasma glucose, insulin ↑ thermogenesis in brown adipose tissue ↑ browning of white adipose tissue | [61] |
| 13 mg/kg/d, p.o., for 8 weeks | DBA2J mice/HFD/HSD | ↓ FBG ↓ serum TG, FFA, adiponectin ↓ IR | [62] |
| Cancer | |||
| 25–100 μM | PC-3 cells, C4-2B cells | ↓ cell proliferation ↑ apoptosis | [63] |
| 50 mg/kg, p.o.; 5 days/week, for 4–5 weeks | C4-2B xenografted mice PC-3 xenografted mice | ↓ tumor volume ↓ Ki67, AR, and pAKT in tumor samples | [63] |
| 50–200 μM | HepG2 cells | ↓ Cell survival ↓ Wnt/β-catenin signaling; β-catenin ↓ c-Myc, cyclin D1, p-c-Jun ↑ TP53, BAX, PUMA, NOXA, p53, p-p53 ↑ Caspase-3, p-p38 | [64] |
| Pretreatment with 5–25 μM | A549, H460, H1299 cells | ↓ EMT ↓ Snail expression | [65] |
| 0.5–10 μM | HCT-116, Caco-2, and HT-29 cells | ↓ Colony formation G2/M arrest (Caco-2, HCT-116) ↑ Senescence-associated β–galactosidase activity (HCT-116) ↑ p53 and p21Cip1/Waf1 expression (HCT-116) | [66] |
| 30 μM | HCT116 cells | ↓ Cell growth ↑ p53, p21, TIGAR expression | [67] |
| 1.5 μM | SW620 cells | ↓ Proliferation, MMP-9 activity ↑ Autophagy, LC3 G2/M arrest ↑ Apoptosis, necrosis | [68] |
| Alzheimer’s disease | |||
| 200 mg/kg/d, i.g., for 2 months | 6-month-old APP/PS1 AD mice | ↑ Cognitive behavior ↓ Aβ 1–40 and 1–42 | [15] |
| 300 mg/kg/d, p.o., for 14 days | 28-week-old APP/PS1 AD mice | ↓ learning and memory deficits ↓ Aβ 42 in the cerebral cortex and hippocampus | [16] |
| 5 mg/kg/d, p.o., 10 months in alternate weeks (1 week on, 1 week off) | 3xTg-AD mice | ↓ Aβ 42 in hippocampus ↑ learning and exploratory behavior | [55] |
| 30 μM | HT22 cells/Aβ oligomers | ↓ Aβ, SQSTM1, LC3 | [55] |
| Intraperitoneal injection, 2.5 mg/kg, 3 times per week for 4 months | 7-month-old hAbKI mice | ↑ Cognitive behavior ↓ Aβ 1–40 and 1–42 ↓ mitochondria fission proteins Drp1, Fis1 ↑ mitochondria fusion proteins Mfn2, Opa1 ↑ mitochondrial biogenesis proteins PGC1α, Nrf1, Nrf2, TFAM ↑ mitophagy proteins PINK1, Parkin ↑ synaptic proteins synaptophysin PSD95 ↑ autophagy proteins ATG5, Beclin, BCL2, LC3B-I, LC3B-II ↓ microglia IBA-1, astrocytes GFAP, neuronal NeuN in brain tissue | [69] |
| Brain injury | |||
| 2.5, 5 mg/kg, i.p., twice s1 h and 24 h before MCAO surgery | C57BL/6 mice/MCAO | ↓ infarct volume, NDS ↓ mRNA of autophagy genes Atf6 and Chop in the brain tissue | [14] |
| 30 μM | N2a cells, primary cultured neurons/OGD/R injury | ↑ cell viability ↓LDH ↑ LC3II, ↓p62 ↓ mRNA of autophagy genes Atf6 and Chop | [14] |
| 2.5 mg/kg, i.p., immediately after controlled injury and every 24 h for 3 days | C57BL/6J/CTI | ↓ NSS score, brain edema ↓ TUNEL+/NeuN+ cells ↓ caspase-3 ↑ bcl-2 ↑ LC3-II/LC3-I ↓ p62 ↓ p-Akt/Akt, p-mTOR/mTOR, p-IKKα/IKKα, p-NFκB/NFκB in the hippocampus | [70] |
| 1.5, 2 mg/kg, i.p., twice 1 h and 24 h before MCAO surgery | Mice/MCAO | ↓ infarct volume, NDS in hippocampus ↓ spatial memory deficits ↑ Nissl+ cells ↓ TUNEL+ cells ↓ Bax, caspase-3, ↑ Bcl-2 in the hippocampus | [71] |
| Parkinson’s disease | |||
| 20 mg/kg/d, i.p., 7-day pretreatment | Mice/MPTP | ↑ Nissl+, TH+neurons ↓ motor deficits ↓ p62 ↑ LC3II/I ↓ NLRP3, caspase-1, IL-1β ↓ IBA1+, GFAP+ cells in the SN ↓ NLRP3 inflammasome activation ↓ neuroinflammation | [72] |
| 10 μM | BV2 cells/LPS | ↓ p62 ↑ LC3II/I ↑ PINK1 and Parkin ↓ Tim23 and Tom20 ↓ mRNA of Il1β, Tnfα, iNos, and Cox2 ↓ NO, NLRP3, caspase-1, IL-1β ↑ MMP, mitochondrial metabolism | [72] |
| 10 μM | PC12 cells/6-OHDA | ↑ cell viability ↓ apoptotic rate ↑ MMP, ↑ Tim23, Tom20 ↑ TFAM, PGC1α, SIRT1 | [73] |
| 10 mg/kg/d, i.p., 7-day pretreatment | C57BL/6J mice/6-OHDA | ↑ Nissl+, TH+neurons ↓ motor deficits ↑ TFAM, PGC1α, SIRT1 In the SN | [73] |
| Diet exposition (exposure details not available) | 9-month-old Thy-1α-syn mice | ↑ Blood colonic γδ T cells ↑ novel object recognition | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojciechowska, O.; Kujawska, M. Urolithin A in Health and Diseases: Prospects for Parkinson’s Disease Management. Antioxidants 2023, 12, 1479. https://doi.org/10.3390/antiox12071479
Wojciechowska O, Kujawska M. Urolithin A in Health and Diseases: Prospects for Parkinson’s Disease Management. Antioxidants. 2023; 12(7):1479. https://doi.org/10.3390/antiox12071479
Chicago/Turabian StyleWojciechowska, Olga, and Małgorzata Kujawska. 2023. "Urolithin A in Health and Diseases: Prospects for Parkinson’s Disease Management" Antioxidants 12, no. 7: 1479. https://doi.org/10.3390/antiox12071479
APA StyleWojciechowska, O., & Kujawska, M. (2023). Urolithin A in Health and Diseases: Prospects for Parkinson’s Disease Management. Antioxidants, 12(7), 1479. https://doi.org/10.3390/antiox12071479