
Validating the Health Benefits of Coffee Berry Pulp Extracts in Mice with High-Fat Diet-Induced Obesity and Diabetes

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
2.2. High-Performance Liquid Chromatography Analyses (HPLC)
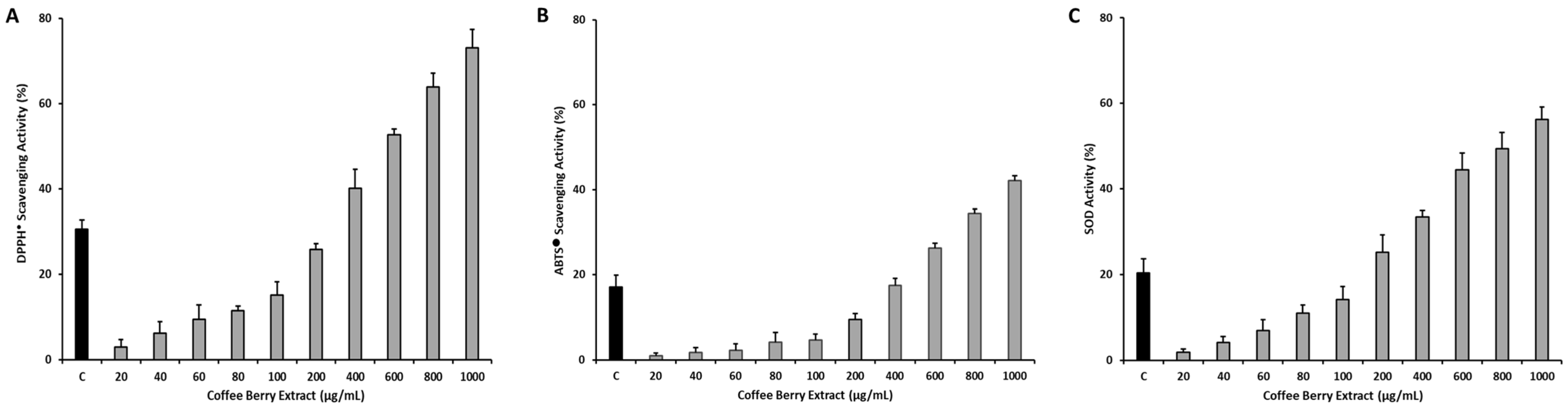
2.3. In Vitro Assessment of Antioxidant Activity
2.3.1. DPPH● Scavenging Activity
2.3.2. ABTS● Scavenging Activity
2.3.3. SOD Activity
2.4. Animal Model
- NFD control = 10 mL/kg of vehicle (distilled water)-administered mice with NFD supply
- HFD control = 10 mL/kg of vehicle (distilled water)-administered mice with HFD supply
- MET250 = 250 mg/kg of metformin-administered mice with HFD supply
- CBP400 = 400 mg/kg of CBP extract-administered mice with HFD supply
- CBP200 = 200 mg/kg of CBP extract-administered mice with HFD supply
- CBP100 = 100 mg/kg of CBP extract-administered mice with HFD supply.
2.5. Test Substance Dosage and Administration
2.6. Observation Items
2.7. Statistical Analysis
3. Results
3.1. Content of Chlorogenic Acid in the CBP Extracts
3.2. Evaluation of In Vitro Antioxidant Activity
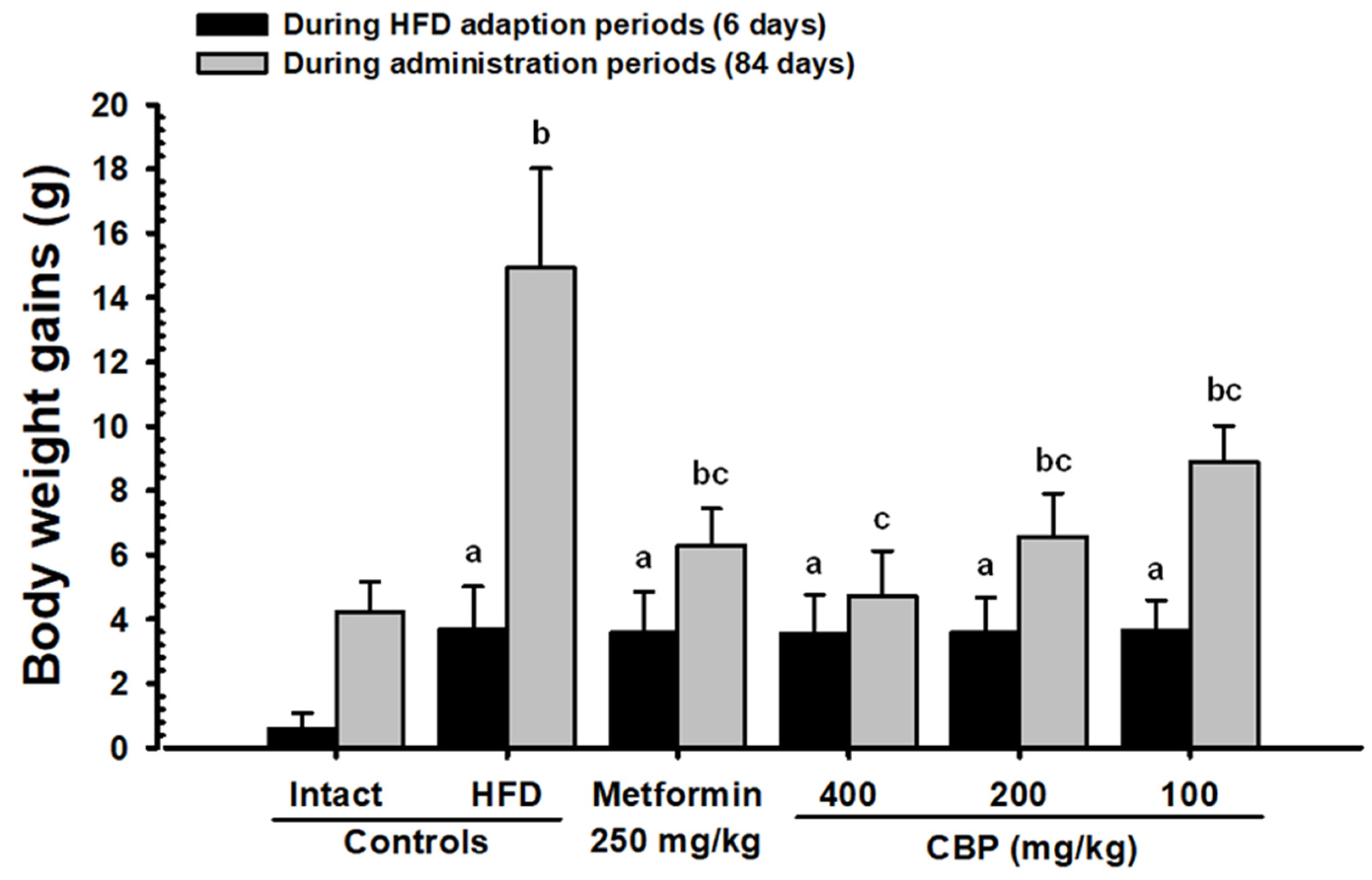
3.3. Anti-Obesity Properties
3.3.1. Effects on Body Weights and Average Feed Intake
3.3.2. Effects on Abdominal and Body Fat Volume
3.3.3. Change in Fat Weight
3.3.4. Histopathological Observations for Accumulated Fats around the Abdominal Wall and the Ovaries
3.3.5. Histopathological Observations for Pancreatic Exocrine Zymogen Granules
3.4. Anti-Diabetic Properties
3.4.1. Effects on Insulin Content, Blood Sugar, and HbA1c Ratio
3.4.2. Effects on Pancreatic Weight
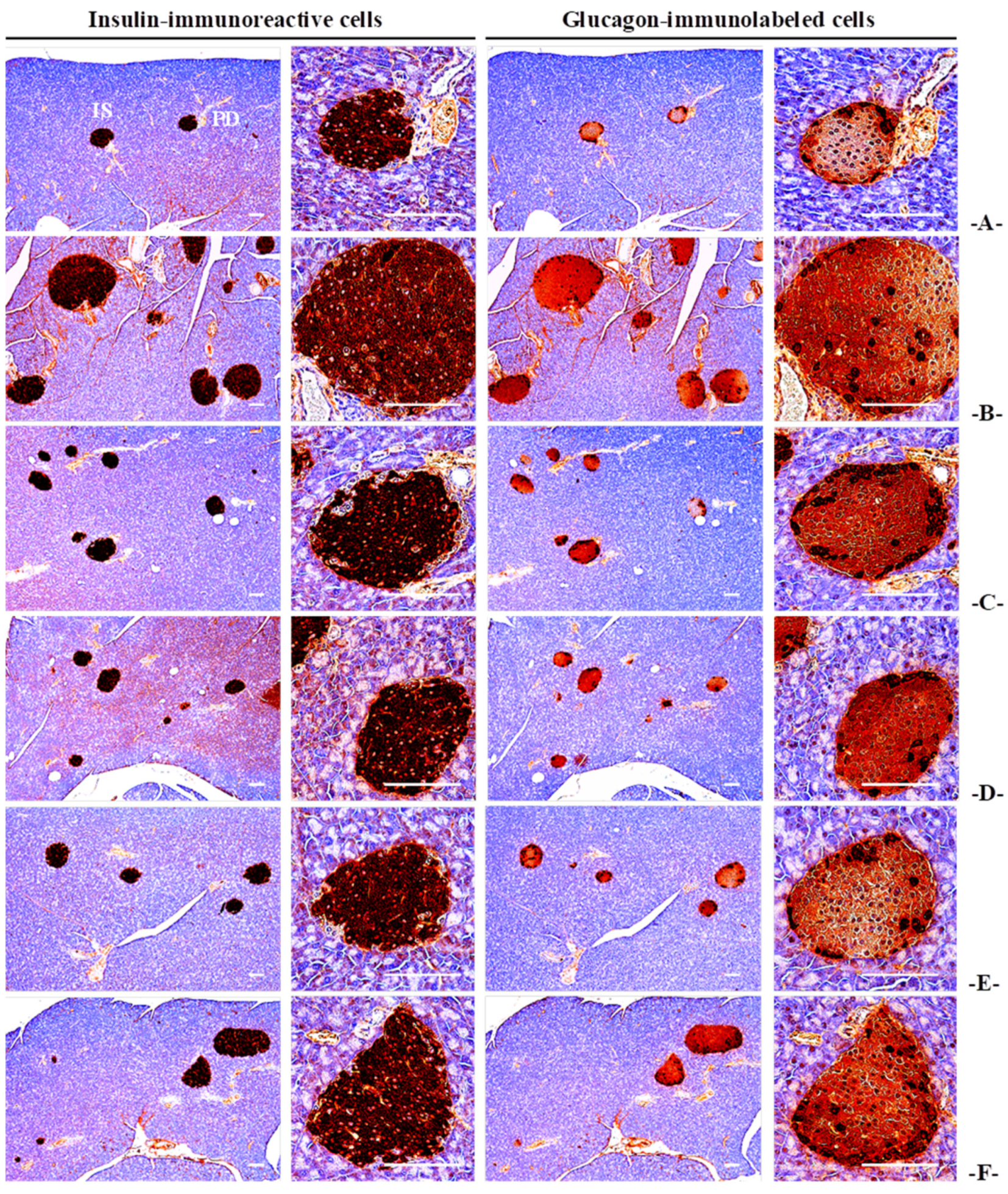
3.4.3. Effects on the General Histopathology of Pancreatic Islands
3.4.4. Effects on the Immunohistochemistry of Pancreatic Islands
3.5. Effects on Hyperlipidemia
3.5.1. Effects on Blood TC, TG, and LDL Content
3.5.2. Effects on HDL Content in Blood
3.5.3. Effects on Fecal Lipid Content
3.6. Liver Damage
3.6.1. Liver Weight
3.6.2. Changes in Serum ALT, AST, ALP, GGT, and LDH Content
3.6.3. Effects on Histopathology of Hepatic Fat Change Rate and Hepatocellular Diameter
3.7. Kidney Damage
3.7.1. Effects on Kidney Weight
3.7.2. Effects on Blood BUN and Creatinine Content
3.7.3. Histopathology of Kidney
3.8. Effect on the Hepatic Antioxidant Defense Systems
3.8.1. Lipid Peroxidation
3.8.2. Changes in Hepatic SOD, CAT, and GSH Activity
3.9. Sugar Metabolism-Related Enzyme Activity
3.9.1. Effects on Hepatic GK Activity
3.9.2. Changes in Liver PEPCK and G6phase Activity
3.10. Changes in Lipid Metabolism-Related Gene Expressions
3.10.1. Changes in Gene Expressions in Liver Tissue
3.10.2. Changes in Gene Expressions in Adipose Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wendel, A.A.; Purushotham, A.; Liu, L.F.; Belury, M.A. Conjugated linoleic acid fails to worsen insulin resistance but induces hepatic steatosis in the presence of leptin in ob/ob mice. J. Lipid Res. 2008, 49, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.S. Obesity, cardiovascular disease, and role of vitamin C on inflammation: A review of facts and underlying mechanisms. Inflammopharmacology 2017, 25, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Carrera, I.; Corzo, L.; Naidoo, V.; Martínez-Iglesias, O.; Cacabelos, R. Cardiovascular and lipid-lowering effects of a marine lipoprotein extract in a high-fat diet-induced obesity mouse model. Int. J. Med. Sci. 2023, 20, 292–306. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. Insulin resistance: Definition and consequences. Exp. Clin. Endocrinol. Diabetes 2001, 109, S135–S148. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.J. Insulin resistance as the core defect in type 2 diabetes mellitus. Am. J. Cardiol. 2002, 90, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Angulo, P. Nonalcoholic fatty liver disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef]
- Inzucchi, S.E. Oral antihyperglycemic therapy for type 2 diabetes: Scientific review. JAMA 2002, 287, 360–372. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, J.W.; Park, J.B.; Pyo, S.E.; Hong, Y.K.; Ku, S.K.; Kim, M.R. Blood glycemia-modulating effects of melanian snail protein hydrolysates in mice with type II diabetes. Int. J. Mol. Med. 2017, 39, 1437–1451. [Google Scholar] [CrossRef]
- Choi, B.R.; Kim, H.J.; Lee, Y.J.; Ku, S.K. Anti-diabetic obesity effects of Wasabia japonica Matsum leaf extract on 45% Kcal high-fat diet-fed mice. Nutrients 2020, 12, 2837. [Google Scholar] [CrossRef]
- Choi, E.H.; Chun, Y.S.; Kim, J.K.; Ku, S.K.; Jeon, S.W.; Park, T.S.; Shim, S.M. Modulating lipid and glucose metabolism by glycosylated kaempferol rich roasted leaves of Lycium chinense via upregulating adiponectin and AMPK activation in obese mice-induced type 2 diabetes. J. Funct. Foods 2020, 72, 104072. [Google Scholar] [CrossRef]
- Kang, H.G.; Bashir, K.M.I.; Kim, K.Y.; Shin, S.; Choi, M.W.; Hong, E.J.; Choi, S.H.; Kim, J.W.; Choi, J.S.; Ku, S.-K. Evaluation of dose-dependent obesity and diabetes-related complications of water chestnut (fruit of Trapa japonica) extracts in type ii obese diabetic mice induced by 45% Kcal high-fat diet. Medicina 2022, 58, 189. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.M.I.; Kim, J.W.; Kim, J.-K.; Chun, Y.-S.; Choi, J.-S.; Ku, S.-K. Efficacy confirmation test of black cumin (Nigella sativa L.) seeds extract using a high-fat diet mouse model. Metabolites 2023, 13, 501. [Google Scholar] [CrossRef] [PubMed]
- Shokouh, P.; Jeppesen, P.B.; Hermansen, K.; Nørskov, N.P.; Laustsen, C.; Jacques Hamilton-Dutoit, S.; Qi, H.; Stødkilde-Jørgensen, H.; Gregersen, S. A combination of coffee compounds shows insulin-sensitizing and hepatoprotective effects in a rat model of diet-induced metabolic syndrome. Nutrients 2017, 10, 6. [Google Scholar] [CrossRef]
- Heeger, A.; Kosińska-Cagnazzo, A.; Cantergiani, E.; Andlauer, W. Bioactives of coffee cherry pulp and its utilisation for production of Cascara beverage. Food Chem. 2017, 221, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Oliva, A.; Uscanga-Sosa, D.P.; González-Rios, O.; Morales-Ramos, V. The use of coffee (Coffea arabica L.) pulp in the preparation of a beverage with antioxidant properties. Int. Food Res. J. 2022, 29, 274–282. [Google Scholar] [CrossRef]
- Handayani, B.R. Study and Characterization of Antibacterial Compounds of Arabica Coffee Berry Pulp. Ph.D. Dissertation, Oklahoma State University, Stillwater, OK, USA, 2009; pp. 1–97. [Google Scholar]
- Jackson, K.M.P.; Rathinasabapathy, T.; Esposito, D.; Komarnytsky, S. Structural constraints and importance of caffeic acid moiety for anti-hyperglycemic effects of caffeoylquinic acids from chicory. Mol. Nutr. Food Res. 2017, 61, 1601118. [Google Scholar] [CrossRef]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef]
- Santos, R.M.; Lima, D.R. Coffee consumption, obesity and type 2 diabetes: A mini-review. Eur. J. Nutr. 2016, 55, 1345–1358. [Google Scholar] [CrossRef]
- Ding, M.; Bhupathiraju, S.N.; Chen, M.; van Dam, R.M.; Hu, F.B. Caffeinated and decaffeinated coffee consumption and risk of type 2 diabetes: A systematic review and a dose-response meta-analysis. Diabetes Care 2014, 37, 569–586. [Google Scholar] [CrossRef]
- Song, S.J.; Choi, S.; Park, T. Decaffeinated green coffee bean extract attenuates diet-induced obesity and insulin resistance in mice. eCAM 2014, 2014, 718379. [Google Scholar] [CrossRef] [PubMed]
- Mellbye, F.B.; Jeppesen, P.B.; Hermansen, K.; Gregersen, S. Cafestol, a bioactive substance in coffee, stimulates insulin secretion and increases glucose uptake in muscle cells: Studies in vitro. J. Nat. Prod. 2015, 78, 2447–2451. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.N.; Moon, S.J.; Ko, S.K.; Im, B.O.; Chung, S.H. Wild ginseng prevents the onset of high-fat diet induced hyperglycemia and obesity in ICR mice. Arch. Pharm. Res. 2004, 27, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Lee, Y.S.; Seol, D.J.; Cho, I.J.; Ku, S.K.; Choi, J.-S.; Lee, H.J. Anti-obesity and fatty liver-preventing activities of Lonicera caerulea in high-fat diet-fed mice. Int. J. Mol. Med. 2018, 42, 3047–3064. [Google Scholar] [CrossRef] [PubMed]
- Kuda, T.; Tsunekawa, M.; Goto, H.; Araki, Y. Antioxidant properties of four edible algae harvested in the Noto Peninsula, Japan. J. Food Compost. Anal. 2005, 18, 625–633. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-R.; Kim, J.W.; Park, J.B.; Hong, Y.-K.; Ku, S.-K.; Choi, J.-S. Anti-obesity effects of yellow catfish protein hydrolysate on mice fed a 45% kcal high fat diet. Int. J. Mol. Med. 2017, 40, 784–800. [Google Scholar] [CrossRef]
- US FDA. Guidance for Industry: Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers. 2005. Available online: https://www.fda.gov/media/72309/download (accessed on 23 April 2023).
- KFDA. Standard for Toxicity Study of Pharmaceuticals. 2017. Available online: https://www.mfds.go.kr/eng/brd/m_18/view.do?seq=71459&srchFr=&srchTo=&srchWord=&srchTp=&itm_seq_1=0&itm_seq_2=0&multi_itm_seq=0&company_cd=&company_nm=&page=1 (accessed on 11 December 2022).
- Jamall, I.S.; Smith, J.C. Effects of cadmium on glutathione peroxidase, superoxidase dismutase and lipid peroxidation in the rat heart: A possible mechanism of cadmium cardiotoxicity. Toxicol. Appl. Pharmacol. 1985, 80, 33–42. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Bolzán, A.D.; Bianchi, M.S.; Bianchi, N.O. Superoxide dismutase, catalase and glutathione peroxidase activities in human blood: Influence of sex, age and cigarette smoking. Clin. Biochem. 1997, 30, 449–454. [Google Scholar] [CrossRef]
- Assady, M.; Farahnak, A.; Golestani, A.; Esharghian, M. Superoxide dismutase (SOD) enzyme activity assay in Fasciola spp. parasites and liver tissue extract. Iran. J. Parasitol. 2011, 6, 17–22. [Google Scholar] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.; Wang, J.; Yang, L.; An, Y.; Zhu, H. AMPK activation enhances the anti-atherogenic effects of high density lipoproteins in apoE(−/−) mice. J. Lipid Res. 2017, 58, 1536–1547. [Google Scholar] [CrossRef] [PubMed]
- Mottillo, E.P.; Desjardins, E.M.; Fritzen, A.M.; Zou, V.Z.; Crane, J.D.; Yabut, J.M.; Kiens, B.; Erion, D.M.; Lanba, A.; Granneman, J.G.; et al. FGF21 does not require adipocyte AMP-activated protein kinase (AMPK) or the phosphorylation of acetyl-CoA carboxylase (ACC) to mediate improvements in whole-body glucose homeostasis. Mol. Metab. 2017, 6, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.H.; Kim, T.W.; Kim, C.G.; Kim, T.W.; Park, N.H. Antioxidant activity of various solvent fractions from edible brown alga, Eisenia bicyclcis and its active compounds. J. Food Sci. 2013, 15, C679–C684. [Google Scholar]
- Bashir, K.M.I.; Park, Y.-J.; An, J.H.; Choi, S.-J.; Kim, J.-H.; Baek, M.-K.; Kim, A.; Sohn, J.-H.; Choi, J.S. Antioxidant properties of Scomber japonicus hydrolysates prepared by enzymatic hydrolysis. J. Aquat. Food Prod. Technol. 2018, 27, 107–121. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Sohn, J.-H.; Kim, J.S.; Choi, J.-S. Identification and characterization of novel antioxidant peptides from mackerel (Scomber japonicus) muscle protein hydrolysates. Food Chem. 2020, 323, 126809. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.W.; Cho, C.H.; Lee, J.S.; Lee, S.H.; Kim, T.; Kim, D.O. Deastringent peel extracts of persimmon (Diosyros kaki thumb. cv. Cheondo-bansi) protect neuronal PC-12 and SH-SY5Y cells against oxidative stress. J. Microbiol. Biotechnol. 2018, 28, 1094–1104. [Google Scholar] [CrossRef]
- Thyagarajan, A.; Sahu, R.P. Potential contributions of antioxidants to cancer therapy: Immunomodulation and radiosensitization. Integr. Cancer Ther. 2018, 17, 210–216. [Google Scholar] [CrossRef]
- Haider, K.; Haider, M.R.; Neha, K.; Yar, M.S. Free radical scavengers: An overview on heterocyclic advances and medicinal prospects. Eur. J. Med. Chem. 2020, 204, 112607. [Google Scholar] [CrossRef]
- Kim, N.Y.; Yim, T.B.; Lee, H.Y. Skin anti-aging activities of bacteriochlorophyll a from photosynthetic bacteria, Rhodobacter sphaeroides. J. Microbiol. Biotechnol. 2015, 25, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Geremu, M.; Tola, Y.B.; Sualeh, A. Extraction and determination of total polyphenols and antioxidant capacity of red coffee (Coffea arabica L.) pulp of wet processing plants. Chem. Biol. Technol. Agric. 2016, 3, 25. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Shih, C.-H.; Lin, T.-C.; Zheng, J.-H.; Hsu, C.-C.; Chen, K.-M.; Lin, Y.-S.; Wu, C.-T. Antioxidation and tyrosinase inhibitory ability of coffee pulp extract by ethanol. J. Chem. 2021, 2021, 8649618. [Google Scholar] [CrossRef]
- Eswari, R.; Geetha, S.; Kalpana, V.K. Antimicrobial and antioxidant activity of coffee pulp extract. IOSR J. Pharm. Biol. Sci. 2021, 16, 27–32. [Google Scholar]
- Pieme, C.A.; Penlap, V.N.; Ngogang, J.; Costache, M. In vitro cytotoxicity and antioxidant activities of five medicinal plants of Malvaceae family from Cameroon. Environ. Toxicol. Pharmacol. 2010, 29, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Mohd Esa, N.; Abdul Kadir, K.K.; Amom, Z.; Azlan, A. Antioxidant activity of white rice, brown rice and germinated brown rice (in vivo and in vitro) and the effects on lipid peroxidation and liver enzymes in hyperlipidaemic rabbits. Food Chem. 2013, 141, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.; Salam, S.K.; Salleh, M.S.; Gurtu, S. Comparison of antioxidant effects of honey, glibenclamide, metformin, and their combinations in the kidneys of streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2011, 12, 829–843. [Google Scholar] [CrossRef]
- Wu, D.; Wen, W.; Qi, C.L.; Zhao, R.X.; Lü, J.H.; Zhong, C.Y.; Chen, Y.Y. Ameliorative effect of berberine on renal damage in rats with diabetes induced by high-fat diet and streptozotocin. Phytomedicine 2012, 19, 712–718. [Google Scholar] [CrossRef]
- Saewan, N. Effect of coffee berry extract on anti-aging for skin and hair—In vitro approach. Cosmetics 2022, 9, 66. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.J.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. J. Agric. Food Chem. 2004, 52, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [PubMed]
- Chakniramol, S.; Wierschem, A.; Cho, M.-G.; Bashir, K.M.I. Physiological and clinical aspects of bioactive peptides from marine animals. Antioxidants 2022, 11, 1021. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Sone, H.; Suzuki, H.; Takahashi, A.; Yamada, N. Disease model: Hyperinsulinemia and insulin resistance. Part A-targeted disruption of insulin signaling or glucose transport. Trends Mol. Med. 2001, 7, 320–322. [Google Scholar] [CrossRef]
- Park, S.H.; Ko, S.K.; Chung, S.H. Euonymus alatus prevents the hyperglycemia and hyperlipidemia induced by high-fat diet in ICR mice. J. Ethnopharmacol. 2005, 102, 326–335. [Google Scholar] [CrossRef]
- Rustenbeck, I.; Lier-Glaubitz, V.; Willenborg, M.; Eggert, F.; Engelhardt, U.; Jörns, A. Effect of chronic coffee consumption on weight gain and glycaemia in a mouse model of obesity and type 2 diabetes. Nutr. Diabetes 2014, 4, e123. [Google Scholar] [CrossRef]
- Terzo, S.; Caldara, G.F.; Ferrantelli, V.; Puleio, R.; Cassata, G.; Mulè, F.; Amato, A. Pistachio consumption prevents and improves lipid dysmetabolism by reducing the lipid metabolizing gene expression in diet-induced obese mice. Nutrients 2018, 10, 1857. [Google Scholar] [CrossRef]
- Bookchin, R.M.; Gallop, P.M. Structure of hemoglobin AIc: Nature of the N-terminal beta chain blocking group. Biochem. Biophys. Res. Commun. 1968, 32, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.L.; Hørder, M.; Mogensen, E.F. Effect of long-term monitoring of glycosylated hemoglobin levels in insulin-dependent diabetes mellitus. N. Engl. J. Med. 1990, 323, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.I.; Rico, C.W.; Kang, M.Y. Comparative study on the hypoglycemic and antioxidative effects of fermented paste (doenjang) prepared from soybean and brown rice mixed with rice bran or red ginseng marc in mice fed with high fat diet. Nutrients 2014, 6, 4610–4624. [Google Scholar] [CrossRef] [PubMed]
- Noriega-López, L.; Tovar, A.R.; Gonzalez-Granillo, M.; Hernández-Pando, R.; Escalante, B.; Santillán-Doherty, P.; Torres, N. Pancreatic insulin secretion in rats fed a soy protein high fat diet depends on the interaction between the amino acid pattern and isoflavones. J. Biol. Chem. 2007, 282, 20657–20666. [Google Scholar] [CrossRef] [PubMed]
- Terauchi, Y.; Takamoto, I.; Kubota, N.; Matsui, J.; Suzuki, R.; Komeda, K.; Hara, A.; Toyoda, Y.; Miwa, I.; Aizawa, S.; et al. Glucokinase and IRS-2 are required for compensatory beta cell hyperplasia in response to high-fat diet-induced insulin resistance. J. Clin. Investig. 2007, 117, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qu, Z.; Fu, L.; Dong, P.; Zhang, X. Physicochemical properties and antioxidant capacity of 3 polysaccharides from green tea, oolong tea, and black tea. J. Food Sci. 2009, 74, C469–C474. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.J.; Wang, S.S.; Jiang, Y.; Wang, Y.; Shen, H.; Xu, P.; Xiang, H.; Xiao, H. Hypolipidemic effect of XH601 on hamsters of hyperlipidemia and its potential mechanism. Lipids Health Dis. 2017, 16, 85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chen, L.; Zhou, H.; Deng, C.; Han, Q.; Chen, Y.; Wu, Q.; Li, S. Protective effect of carvacrol on liver injury in type 2 diabetic db/db mice. Mol. Med. Rep. 2021, 24, 741. [Google Scholar] [CrossRef]
- Sodikoff, C.H. Laboratory Profiles of Small Animal Diseases: A Guide to Laboratory Diagnosis, 2nd ed.; Mosby: St. Louise, MO, USA, 1995; pp. 1–36. [Google Scholar]
- George, J.; Zhang, Y.; Sloan, J.; Sims, J.M.; Imig, J.D.; Zhao, X. Tim-1 deficiency aggravates high-fat diet-induced steatohepatitis in mice. Front. Immunol. 2021, 12, 747794. [Google Scholar] [CrossRef]
- Chi, P.J.; Lee, C.J.; Hsieh, Y.J.; Lu, C.W.; Hsu, B.G. Dapagliflozin ameliorates lipopolysaccharide related acute kidney injury in mice with streptozotocin-induced diabetes mellitus. Int. J. Med. Sci. 2022, 19, 729–739. [Google Scholar] [CrossRef]
- Garg, M.C.; Singh, K.P.; Bansal, D.D. Effect of vitamin C supplementation on oxidative stress in experimental diabetes. Indian. J. Exp. Biol. 1997, 35, 264–266. [Google Scholar]
- Comporti, M. Lipid peroxidation and cellular damage in toxic liver injury. Lab. Investig. 1985, 53, 599–623. [Google Scholar] [PubMed]
- Wu, D.; Zheng, N.; Qi, K.; Cheng, H.; Sun, Z.; Gao, B.; Zhang, Y.; Pang, W.; Huangfu, C.; Ji, S.; et al. Exogenous hydrogen sulfide mitigates the fatty liver in obese mice through improving lipid metabolism and antioxidant potential. Med. Gas Res. 2015, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Ferre, T.; Riu, E.; Bosch, F.; Valera, A. Evidence from transgenic mice that glucokinase is rate limiting for glucose utilization in the liver. FASEB J. 1996, 10, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Coope, G.J.; Atkinson, A.M.; Allott, C.; McKerrecher, D.; Johnstone, C.; Pike, K.G.; Holme, P.C.; Vertigan, H.; Gill, D.; Coghlan, M.P.; et al. Predictive blood glucose lowering efficacy by glucokinase activators in high fat fed female Zucker rats. Br. J. Pharmacol. 2006, 149, 328–335. [Google Scholar] [CrossRef] [PubMed]
- She, P.; Shiota, M.; Shelton, K.D.; Chalkley, R.; Postic, C.; Magnuson, M.A. Phosphoenolpyruvate carboxykinase is necessary for the integration of hepatic energy metabolism. Mol. Cell. Biol. 2000, 20, 6508–6517. [Google Scholar] [CrossRef] [PubMed]
- van Schaftingen, E.; Gerin, I. The glucose-6-phosphatase system. Biochem. J. 2002, 362, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef]
- Lin, C.H.; Kuo, Y.H.; Shih, C.C. Effects of Bofu-Tsusho-San on diabetes and hyperlipidemia associated with AMP-activated protein kinase and glucose transporter 4 in high-fat-fed mice. Int. J. Mol. Sci. 2014, 15, 20022–20044. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Kim, D.S.; Kim, S.H.; Kim, H.K. Anti-obesity activity, acute toxicity, and chemical constituents of aqueous and ethanol Viola mandshurica extracts. BMC Complement. Med. Ther. 2017, 17, 297. [Google Scholar] [CrossRef]
- Park, J.E.; Lee, H.; Rho, H.; Hong, S.M.; Kim, S.Y.; Lim, Y. Effect of Quamoclit angulata extract supplementation on oxidative stress and inflammation on hyperglycemia-induced renal damage in type 2 diabetic mice. Antioxidants 2020, 9, 459. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Zhang, L. Resveratrol provides benefits in mice with type II diabetes-induced chronic renal failure through AMPK signaling pathway. Exp. Ther. Med. 2018, 16, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Torres-Valenzuela, L.S.; Serna-Jiménez, J.A.; Martínez, K. Coffee by-products: Nowadays and perspectives. In Coffee—Production and Research; Dalyse, T.C., Ed.; IntechOpen Publishers: London, UK, 2020; pp. 1–18. [Google Scholar]
- Junior, M.P.F.; de Siqueira, J.C.; dos Reis Souza, A.; de Matos, M.P.; Fia, R. Use of by-products generated in the processing of coffee berries: A review. Coffee Sci. 2023, 18, e182101. [Google Scholar]
- Manasa, V.; Padmanabhan, A.; Appaiah, K.A. Utilization of coffee pulp waste for rapid recovery of pectin and polyphenols for sustainable material recycle. Waste Manag. 2021, 120, 762–771. [Google Scholar] [CrossRef]
- Bertrand, B.; Davis, A.P.; Maraval, I.; Forestier, N.; Mieulet, D. Potential beverage quality of three wild coffee species (Coffea brevipes, C. congensis and C. stenophylla) and consideration of their agronomic use. J. Sci. Food Agric. 2023, 103, 3602–3612. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, B.; Mun, E.G.; Jeong, S.Y.; Cha, Y.S. The antioxidant activity of steamed ginger and its protective effects on obesity induced by high-fat diet in C57BL/6J mice. Nutr. Res. Pract. 2018, 12, 503–511. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Absolute Organ Weights (g) | ||||
|---|---|---|---|---|---|
| Liver | Kidney | Pancreas | Periovarian Fat Pads | Abdominal Wall Fat Pads | |
| Controls | |||||
| NFD | 1.238 ± 0.031 | 0.208 ± 0.011 | 0.264 ± 0.039 | 0.062 ± 0.035 | 0.035 ± 0.026 |
| HFD | 1.906 ± 0.094 a | 0.297 ± 0.020 a | 0.255 ± 0.021 | 0.452 ± 0.125 a | 0.387 ± 0.051 a |
| Reference | |||||
| MET250 | 1.526 ± 0.102 ac | 0.241 ± 0.012 ac | 0.259 ± 0.026 | 0.237 ± 0.072 ac | 0.207 ± 0.024 ac |
| Test materials | |||||
| CBP400 | 1.418 ± 0.144 bc | 0.227 ± 0.009 ac | 0.262 ± 0.026 | 0.163 ± 0.022 ac | 0.135 ± 0.039 ac |
| CBP200 | 1.436 ± 0.081 ac | 0.241 ± 0.009 ac | 0.258 ± 0.012 | 0.236 ± 0.053 ac | 0.215 ± 0.068 ac |
| CBP100 | 1.638 ± 0.097 ac | 0.258 ± 0.009 ac | 0.259 ± 0.023 | 0.301 ± 0.023 ad | 0.264 ± 0.043 ac |
| Groups | Relative Organ Weights (% of Body Weights) | ||||
|---|---|---|---|---|---|
| Liver | Kidney | Pancreas | Periovarian Fat Pads | Abdominal Wall Fat Pads | |
| Controls | |||||
| NFD | 4.223 ± 0.219 | 0.709 ± 0.036 | 0.900 ± 0.129 | 0.211 ± 0.119 | 0.119 ± 0.091 |
| HFD | 4.393 ± 0.389 | 0.685 ± 0.071 | 0.589 ± 0.081 c | 1.030 ± 0.229 a | 0.889 ± 0.117 c |
| Reference | |||||
| MET250 | 4.381 ± 0.238 | 0.692 ± 0.055 | 0.744 ± 0.076 e | 0.678 ± 0.109 ab | 0.595 ± 0.082 ce |
| Test materials | |||||
| CBP400 | 4.259 ± 0.357 | 0.685 ± 0.059 | 0.789 ± 0.073 e | 0.491 ± 0.085 ab | 0.402 ± 0.105 ce |
| CBP200 | 4.423 ± 0.235 | 0.693 ± 0.024 | 0.744 ± 0.047 de | 0.677 ± 0.134 ab | 0.618 ± 0.188 cf |
| CBP100 | 4.400 ± 0.244 | 0.693 ± 0.025 | 0.694 ± 0.049 cf | 0.808 ± 0.074 ab | 0.710 ± 0.122 cf |
| Groups | Periovarian Fat Pads | Abdominal Wall Fat Pads | ||
|---|---|---|---|---|
| Thickness (mm) | Adipocyte Diameters (μm) | Thickness (mm) | Adipocyte Diameters (μm) | |
| Controls | ||||
| NFD | 1.52 ± 0.34 | 41.48 ± 11.19 | 1.70 ± 0.37 | 39.48 ± 10.14 |
| HFD | 4.76 ± 0.89 c | 120.55 ± 17.20 a | 5.97 ± 1.03 c | 105.97 ± 12.31 a |
| Reference | ||||
| MET250 | 2.80 ± 0.37 ce | 64.00 ± 12.72 ab | 3.35 ± 0.57 ce | 60.26 ± 12.61 ab |
| Test materials | ||||
| CBP400 | 2.29 ± 0.49 de | 47.58 ± 10.66 b | 2.71 ± 0.69 de | 52.35 ± 10.04 b |
| CBP200 | 2.82 ± 0.38 ce | 64.01 ± 10.72 ab | 3.39 ± 0.57 ce | 61.86 ± 13.38 ab |
| CBP100 | 3.18 ± 0.37 ce | 82.65 ± 10.10 ab | 4.13 ± 0.30 ce | 81.08 ± 11.37 ab |
| Groups | Glucose (mg/dL) | Total Cholesterol (mg/dL) | Triglyceride (mg/dL) | Low Density Lipoprotein (mg/dL) | High Density Lipoprotein (mg/dL) |
|---|---|---|---|---|---|
| Controls | |||||
| NFD | 85.60 ± 12.13 | 88.10 ± 16.72 | 74.50 ± 14.38 | 20.10 ± 3.96 | 94.90 ± 12.64 |
| HFD | 253.50 ± 36.18 c | 259.60 ± 38.58 c | 221.10 ± 40.06 c | 97.50 ± 14.40 a | 19.80 ± 10.71 a |
| Reference | |||||
| MET250 | 161.00 ± 17.11 cd | 167.10 ± 26.04 cd | 148.30 ± 19.11 cd | 62.70 ± 12.15 ab | 52.20 ± 11.78 ab |
| Test materials | |||||
| CBP400 | 115.10 ± 17.24 cd | 114.60 ± 22.17 d | 103.60 ± 27.19 d | 44.50 ± 11.13 ab | 68.60 ± 16.77 ab |
| CBP200 | 161.80 ± 13.86 cd | 165.00 ± 21.41 cd | 148.60 ± 12.85 cd | 62.80 ± 11.79 ab | 51.80 ± 10.34 ab |
| CBP100 | 192.90 ± 15.21 cd | 196.20 ± 13.55 cd | 171.40 ± 11.83 ce | 74.60 ± 11.46 ab | 43.90 ± 10.12 ab |
| Groups | AST (IU/L) | ALT (IU/L) | ALP (IU/L) | LDH (×10 IU/L) | GGT (IU/L) | BUN (mg/dL) | Creatinine (mg/dL) |
|---|---|---|---|---|---|---|---|
| Controls | |||||||
| NFD | 70.90 ± 13.77 | 40.90 ± 11.62 | 70.10 ± 16.93 | 69.95 ± 14.19 | 5.50 ± 1.27 | 32.60 ± 10.96 | 0.55 ± 0.24 |
| HFD | 189.50 ± 14.24 a | 146.30 ± 11.28 a | 199.10 ± 27.10 c | 359.41 ± 78.70 c | 20.10 ± 1.37 a | 132.10 ± 15.14 a | 2.06 ± 0.24 a |
| Reference | |||||||
| MET250 | 129.40 ± 14.26 ab | 93.20 ± 11.71 ab | 125.20 ± 14.53 cd | 186.30 ± 28.93 cd | 12.00 ± 1.63 ab | 82.00 ± 19.50 ab | 1.32 ± 0.23 ab |
| Test materials | |||||||
| CBP400 | 103.20 ± 10.46 ab | 76.90 ± 16.54 ab | 95.60 ± 20.19 d | 136.41 ± 30.98 cd | 8.90 ± 1.66 ab | 63.00 ± 17.83 ab | 0.93 ± 0.16 ab |
| CBP200 | 129.50 ± 14.35 ab | 91.50 ± 18.11 ab | 124.10 ± 7.91 cd | 194.03 ± 34.42 cd | 11.90 ± 1.66 ab | 82.20 ± 19.97 ab | 1.31 ± 0.19 ab |
| CBP100 | 147.20 ± 13.10 ab | 115.30 ± 10.17 ab | 145.80 ± 8.12 cd | 219.80 ± 25.99 cd | 15.40 ± 1.58 ab | 101.90 ± 14.61 ab | 1.57 ± 0.12 ab |
| Groups | Liver Steatosis (%/mm2 of Hepatic Tissues) | Mean Hepatocyte Diameters (μm/cell) | Degenerative Renal Tubule Numbers (%) |
|---|---|---|---|
| Controls | |||
| NFD | 3.08 ± 1.91 | 14.26 ± 1.39 | 4.90 ± 2.56 |
| HFD | 78.64 ± 10.66 c | 28.12 ± 2.46 a | 78.60 ± 10.13 c |
| Reference | |||
| MET250 | 48.80 ± 10.31 cd | 21.18 ± 2.33 ab | 45.60 ± 11.89 cd |
| Test materials | |||
| CBP400 | 27.54 ± 15.48 cd | 18.48 ± 2.24 ab | 28.10 ± 11.86 cd |
| CBP200 | 47.86 ± 10.14 cd | 21.20 ± 2.00 ab | 45.90 ± 12.72 cd |
| CBP100 | 55.34 ± 11.63 cd | 22.86 ± 1.72 ab | 55.50 ± 11.66 cd |
| Groups | Lipid Peroxidation | Antioxidant Defense System | ||
|---|---|---|---|---|
| Malondialdehyde (nM/mg Tissue) | Glutathione (μM/mg Tissue) | Catalase (U/mg Tissue) | SOD (U/mg Tissue) | |
| Controls | ||||
| NFD | 10.52 ± 4.69 | 76.34 ± 10.49 | 70.53 ± 14.53 | 7.28 ± 1.14 |
| HFD | 87.95 ± 10.42 a | 11.21 ± 2.78 a | 11.42 ± 2.08 d | 1.04 ± 0.28 a |
| Reference | ||||
| MET250 | 53.31 ± 12.12 ab | 34.16 ± 13.01 ab | 36.02 ± 11.70 df | 3.17 ± 0.90 ab |
| Test materials | ||||
| CBP400 | 38.58 ± 11.52 ab | 46.61 ± 13.29 ab | 46.86 ± 10.86 ef | 4.21 ± 0.85 ab |
| CBP200 | 53.62 ± 11.68 ab | 34.64 ± 13.86 ab | 35.86 ± 11.95 df | 3.16 ± 1.04 ab |
| CBP100 | 64.47 ± 11.53 ab | 24.73 ± 11.88 a | 26.16 ± 10.20 dg | 2.33 ± 0.65 ac |
| Groups | Glucokinase (nM/min/mg Protein) | Glucose-6-Phosphatase (nM/min/mg Protein) | PEPCK (nM/min/mg Protein) |
|---|---|---|---|
| Controls | |||
| NFD | 5.99 ± 1.57 | 119.25 ± 26.51 | 1.87 ± 0.67 |
| HFD | 1.65 ± 0.20 a | 350.53 ± 104.60 a | 5.82 ± 1.17 a |
| Reference | |||
| MET250 | 3.16 ± 0.71 ac | 206.90 ± 28.45 ad | 3.39 ± 0.65 ac |
| Test materials | |||
| CBP400 | 3.99 ± 1.09 ac | 155.51 ± 20.98 bc | 2.48 ± 0.42 c |
| CBP200 | 3.13 ± 0.57 ac | 204.28 ± 27.26 ad | 3.37 ± 0.49 ac |
| CBP100 | 2.55 ± 0.67 ad | 239.16 ± 32.57 a | 4.01 ± 0.33 ad |
| Groups | Hepatic Tissue (Relative to Control/GAPDH) | ||
|---|---|---|---|
| ACC1 | AMPKα1 | AMPKα2 | |
| Controls | |||
| NFD | 1.00 ± 0.05 | 1.00 ± 0.05 | 1.01 ± 0.04 |
| HFD | 5.46 ± 0.81 a | 0.24 ± 0.05 a | 0.23 ± 0.05 a |
| Reference | |||
| MET250 | 3.02 ± 0.68 ac | 0.50 ± 0.13 ac | 0.46 ± 0.09 ac |
| Test materials | |||
| CBP400 | 2.07 ± 0.85 bc | 0.75 ± 0.15 ac | 0.68 ± 0.15 ac |
| CBP200 | 3.04 ± 0.81 ac | 0.50 ± 0.12 ac | 0.46 ± 0.13 ac |
| CBP100 | 4.09 ± 0.35 ac | 0.43 ± 0.12 ad | 0.39 ± 0.11 ad |
| Groups | Control | Reference | Test materials—CBP | |||
|---|---|---|---|---|---|---|
| NFD | HFD | Metformin | 400 mg/kg | 200 mg/kg | 100 mg/kg | |
| Adipose tissue (Relative to control/GAPDH) | ||||||
| Leptin | 1.00 ± 0.08 | 8.24 ± 0.74 d | 4.38 ± 0.73 de | 2.96 ± 0.79 de | 4.40 ± 0.65 de | 6.21 ± 0.78 de |
| UCP2 | 1.00 ± 0.07 | 0.24 ± 0.07 d | 0.52 ± 0.14 de | 0.70 ± 0.16 de | 0.52 ± 0.13 de | 0.43 ± 0.08 de |
| Adiponectin | 1.00 ± 0.08 | 0.18 ± 0.05 a | 0.40 ± 0.13 ab | 0.59 ± 0.12 ab | 0.40 ± 0.13 ab | 0.33 ± 0.07 ac |
| C/EBPα | 1.00 ± 0.04 | 3.95 ± 1.25 d | 2.03 ± 0.33 de | 1.49 ± 0.24 de | 2.04 ± 0.19 df | 2.39 ± 0.09 df |
| C/EBPβ | 1.00 ± 0.05 | 4.34 ± 0.85 d | 2.39 ± 0.54 de | 1.68 ± 0.27 de | 2.38 ± 0.54 de | 3.10 ± 0.29 df |
| SREBP1c | 1.00 ± 0.05 | 3.01 ± 0.45 d | 1.88 ± 0.25 de | 1.46 ± 0.24 de | 1.89 ± 0.30 de | 2.27 ± 0.19 de |
| PPARα | 1.00 ± 0.06 | 0.20 ± 0.04 d | 0.36 ± 0.07 de | 0.53 ± 0.14 de | 0.37 ± 0.10 de | 0.31 ± 0.05 de |
| PPARγ | 1.00 ± 0.05 | 7.26 ± 1.17 d | 4.33 ± 0.83 de | 2.66 ± 0.79 de | 4.34 ± 0.71 de | 5.04 ± 0.53 de |
| FAS | 1.00 ± 0.06 | 17.03 ± 2.26 d | 9.53 ± 2.12 de | 6.57 ± 2.51 de | 9.59 ± 1.46 de | 12.17 ± 1.23 de |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashir, K.M.I.; Kim, J.W.; Park, H.-R.; Lee, J.-K.; Choi, B.-R.; Choi, J.-S.; Ku, S.-K. Validating the Health Benefits of Coffee Berry Pulp Extracts in Mice with High-Fat Diet-Induced Obesity and Diabetes. Antioxidants 2024, 13, 10. https://doi.org/10.3390/antiox13010010
Bashir KMI, Kim JW, Park H-R, Lee J-K, Choi B-R, Choi J-S, Ku S-K. Validating the Health Benefits of Coffee Berry Pulp Extracts in Mice with High-Fat Diet-Induced Obesity and Diabetes. Antioxidants. 2024; 13(1):10. https://doi.org/10.3390/antiox13010010
Chicago/Turabian StyleBashir, Khawaja Muhammad Imran, Joo Wan Kim, Hye-Rim Park, Jae-Kyoung Lee, Beom-Rak Choi, Jae-Suk Choi, and Sae-Kwang Ku. 2024. "Validating the Health Benefits of Coffee Berry Pulp Extracts in Mice with High-Fat Diet-Induced Obesity and Diabetes" Antioxidants 13, no. 1: 10. https://doi.org/10.3390/antiox13010010
APA StyleBashir, K. M. I., Kim, J. W., Park, H. -R., Lee, J. -K., Choi, B. -R., Choi, J. -S., & Ku, S. -K. (2024). Validating the Health Benefits of Coffee Berry Pulp Extracts in Mice with High-Fat Diet-Induced Obesity and Diabetes. Antioxidants, 13(1), 10. https://doi.org/10.3390/antiox13010010