
Quinces (Cydonia oblonga, Chaenomeles sp., and Pseudocydonia sinensis) as Medicinal Fruits of the Rosaceae Family: Current State of Knowledge on Properties and Use


Abstract
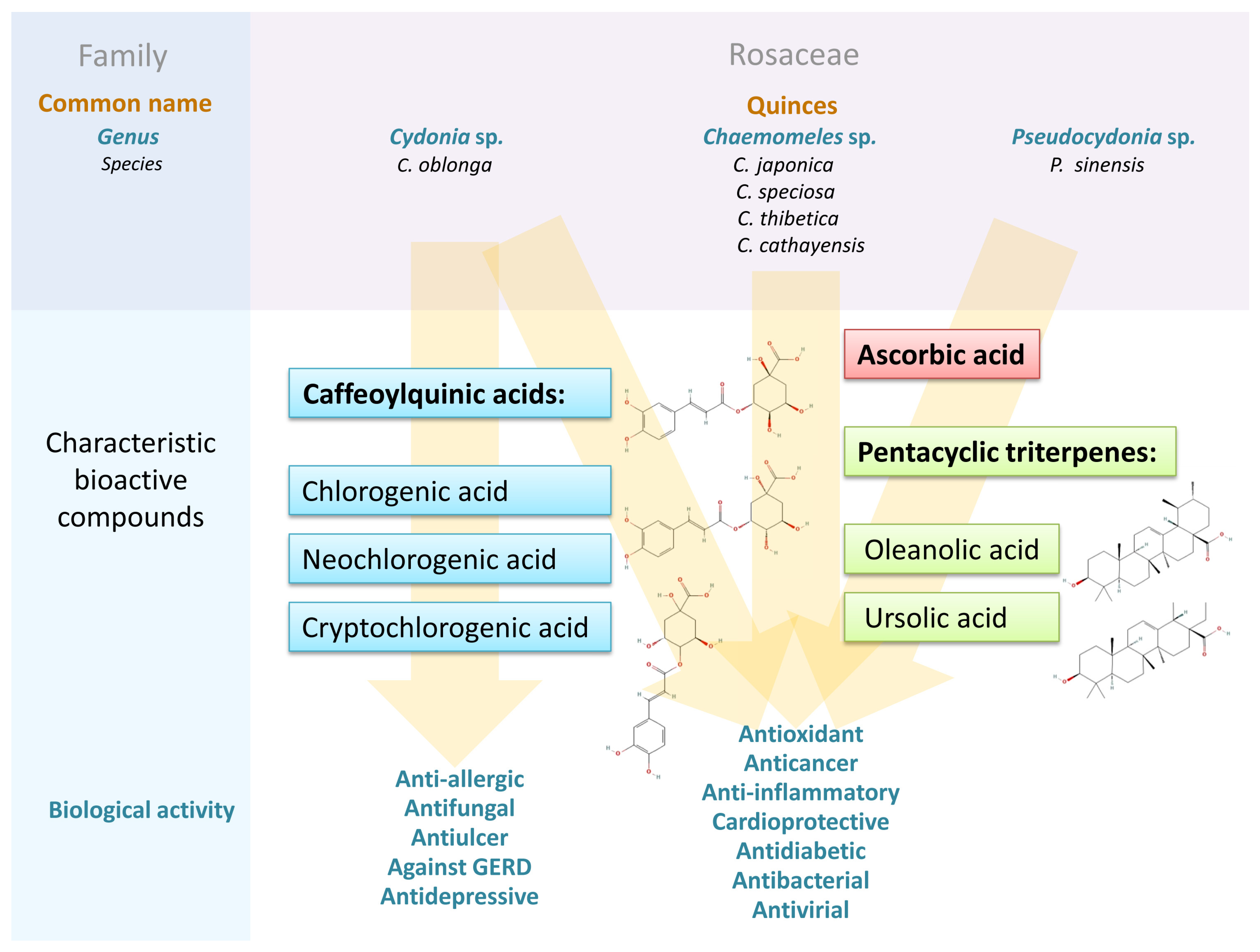
:1. Introduction
2. Characteristics of Quince Plants, Their Biology, Cultivation and Culinary Use
3. Chemical Composition of Quince Fruits and Seeds
3.1. Phenolic Compounds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Species | Content | Ref. | |
|---|---|---|---|---|
| Apigenin | C. japonica | 19.66 | mg/100 g dm | [28] |
| 3-CQA (5-O-caffeoylquinic acid) neochlorogenic acid | P. sinensis | 5.00 | mg/100 g fm | [53] |
| 4-CQA (5-O-caffeoylquinic acid) cryptochlorogenic acid | P. sinensis | 1.20 | mg/100 g fm | [53] |
| 5-CQA (5-O-caffeoylquinic acid) chlorogenic acid | C. japonica | 818.55 | mg/100 g dm | [28] |
| 10.00 | mg/100 g fm ** | [62] | ||
| 12.17 | mg/100 g dm * | [72] | ||
| C. speciosa | 182.00 | mg/100 g fm ** | [62] | |
| C. thiberica | 117.00 | mg/100 g fm ** | [62] | |
| C. cathayensis | 119.00 | mg/100 g fm ** | [62] | |
| P. sinensis | 9.00 | mg/100 g fm ** | [62] | |
| 0.50 | mg/100 g fm | [53] | ||
| (+)-Catechin | C. japonica | 15.75 | mg/100 g dm * | [72] |
| Catechin | C. japonica | 121.12 | mg/100 g dm | [28] |
| C. speciosa | 54.00 | mg/100 g fm ** | [62] | |
| C. thiberica | 156.00 | mg/100 g fm ** | [62] | |
| C. cathayensis | 113.00 | mg/100 g fm ** | [62] | |
| P. sinensis | 5.00 | mg/100 g fm ** | [62] | |
| 2.90 | mg/100 g fm | [53] | ||
| trans-Cinnamic acid | C. japonica | 18.72 | mg/100 g dm | [28] |
| p-Coumaric acid | C. japonica | 5.72 | mg/100 g dm | [28] |
| (–)-Epicatechin | C. japonica | 348.44 | mg/100 g dm * | [72] |
| Epicatechin | C. japonica | 102.00 | mg/100 g fm ** | [62] |
| C. speciosa | 235.00 | mg/100 g fm ** | [62] | |
| P. sinensis | 54.00 | mg/100 g fm ** | [62] | |
| 11.90 | mg/100 g fm | [53] | ||
| 2,5-di-Hydroxybenzoic acid | C. japonica | 2.01 | mg/100 g dm | [28] |
| 4-Hyrdoxybenzoic acid | C. japonica | 1.92 | mg/100 g dm | [28] |
| Ferulic acid | C. japonica | 2.17 | mg/100 g dm | [28] |
| Isoquercitrin | C. japonica | 3.82 | mg/100 g dm | [72] |
| Naringenin | C. japonica | 5.92 | mg/100 g dm | [28] |
| Procyanidin B1 | C. speciosa | 83.00 | mg/100 g fm ** | [62] |
| C. thiberica | 222.00 | mg/100 g fm ** | [62] | |
| C. cathayensis | 145.00 | mg/100 g fm ** | [62] | |
| P. sinensis | 13.00 | mg/100 g fm ** | [62] | |
| 9.80 | mg/100 g fm | [53] | ||
| Procyanidin B2 | C. japonica | 98.00 | mg/100 g dm ** | [62] |
| C. speciosa | 296.00 | mg/100 g fm ** | [62] | |
| P. sinensis | 16.80 | mg/100 g fm | [53] | |
| 40.00 | mg/100 g fm ** | [62] | ||
| Quercitin | C. japonica | 5.03 | mg/100 g dm | [28] |
| Q-3-R (quercetin-3-O-rutinoside) rutin | C. japonica | 107.09 | mg/100 g dm | [28] |
| 5.40 | mg/100 g dm * | [72] | ||
| Sinapic acid | C. japonica | 27.99 | mg/100 g dm * | [72] |
| Syringic acid | C. japonica | 0.03 | mg/100 g dm | [28] |
| Vanilic acid | C. japonica | 13.69 | mg/100 g dm | [28] |
3.2. Ascorbic Acid, Carotenoids, and Other Antioxidants
3.3. Minerals
3.4. Carboxylic Acids
3.5. Carbohydrates Including Fiber
4. Biological Activity of Quince Fruits
4.1. Antioxidant Properties
4.2. Anti-Inflammatory, Anti-Allergic, and Various Immunomodulatory Effects
4.3. Anticancer Activity
4.4. Cardioprotective Effects
4.5. Antidiabetic Activity
4.6. Antiviral and Antibacterial Activity
4.7. Other Health-Promoting Properties
5. Conclusions
6. Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Rop, O.; Balik, J.; Řezniček, V.; Jurikova, T.; Škardova, P.; Salaš, P.; Sochor, J.; Mlček, J.; Kramařova, D. Chemical characteristics of fruits of some selected quince (Cydonia oblonga Mill.) cultivars. Czech J. Food Sci. 2011, 29, 65–73. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. The medical importance of Cydonia oblonga—A review. IOSR J. Pharm. 2016, 6, 87–99. [Google Scholar]
- Nahorska, A.; Dzwoniarska, M.; Thiem, B. Fruits of Japanese quince (Chaenomeles japonica Thunb. Lindl. ex Spach) as a sourcen of bioactive compounds. Post. Fitoter. 2014, 4, 239–246. (In Polish) [Google Scholar]
- Gao, R.; Xiong, S.; Zhang, T.; Deng, X.; Li, J.; Liao, M. Two new quinic acid derivatives from the fruits of Chaenomeles speciose. Biochem. Syst. Ecol. 2020, 93, e104167. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Z.; Ma, W.; Liu, J.; Tian, L.; Zhou, Y.; Shang, F.; Guo, P. Comparative Pollen Morphology of the Genus Chaenomeles Lindl. (Rosaceae): Diagnostic Features and Implications for Taxonomy. Diversity 2023, 15, 960. [Google Scholar] [CrossRef]
- Mabberley, D.J. The Plant Book, a Portable Dictionary of the Higher Plants, 4th ed.; Cambridge University Press: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Martins, R.C.; Seabra, R.M.; Ferreira, M.A. Principal component analysis as tool of characterization of quince (Cydonia oblonga Miller) jam. Food Chem. 2006, 94, 504–512. [Google Scholar] [CrossRef]
- Trigueros, L.; Pérez-Alvarez, J.A.; Viuda-Martos, M.; Sendra, E. Production of low-fat yogurt with quince (Cydonia oblonga Mill.) scalding water. LWT—Food Sci. Technol. 2011, 44, 1388–1395. [Google Scholar] [CrossRef]
- Wojdyło, A.; Teleszko, M.; Oszmiański, J. Antioxidant property and storage stability of quince juice phenolic compounds. Food Chem. 2014, 152, 261–270. [Google Scholar] [CrossRef]
- Mir, S.A.; Wani, S.M.; Wani, T.A.; Ahmad, M.; Gani, A.; Masoodi, F.A.; Nazir, A. Comparative Evaluation of the Proximate Composition and Antioxidant Properties of Processed Products of Quince (Cydonia oblonga). Int. J. Food Res. 2016, 23, 816–821. [Google Scholar] [CrossRef]
- De Almeida Lopes, M.M.; Guimarães Sanches, A.; de Souza, K.O.; de Oliveira Silva, E. Quince—Cydonia oblonga. In Exotic Fruits; Rodrigues, S., de Oliveira Silva, E., de Brito, E.S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 363–368. [Google Scholar] [CrossRef]
- Cascales, E.V.; García, J.M.R. Characteristics of the raw fruit, industrial pulp, and commercial jam elaborated with Spanish quince (Cydonia oblonga Miller). Emir. J. Food Agric. 2020, 32, 623–633. [Google Scholar] [CrossRef]
- Gheisari, H.R.; Abhari, K.H. Drying method effects on the antioxidant activity of quince (Cydonia oblonga Miller) tea. Acta Sci. Pol. Technol. Aliment. 2014, 13, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Carbonell-Barrachina, A.A.; Szychowski, P.J.; Vásquez, M.V.; Hernández, F.; Wojdyło, A. Technological aspects as the main impact on quality of quince liquors. Food Chem. 2015, 167, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Zapata, P.J.; Martínez-Esplá, A.; Gironés-Vilaplana, A.; Santos-Lax, D.; Noguera-Artiaga, L.; Carbonell-Barrachina, A.A. Phenolic, volatile, and sensory profiles of beer enriched by macerating quince fruits. LWT—Food Sci. Technol. 2019, 103, 139–146. [Google Scholar] [CrossRef]
- Baranowska-Bosiacka, I.; Bosiacka, B.; Rast, J.; Gutowska, I.; Wolska, J.; Rębacz-Maron, E.; Dębia, K.; Janda, K.; Korbecki, J.; Chlubek, D. Macro- and Microelement Content and Other Properties of Chaenomeles japonica L. Fruit and Protective Effects of Its Aqueous Extract on Hepatocyte Metabolism. Biol. Trace Elem. Res. 2017, 178, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Nawirska-Olszańska, A.; Kucharska, A.Z.; Sokół-Łętowska, A.; Biesiada, A. Quality Assasment of pumpkin jams enriched with Japanese quince, Cornellian cherry and strawberries. Acta Sci. Pol. Technol. Aliment. 2010, 1, 40–48. (In Polish) [Google Scholar]
- Wojdyło, A.; Oszmiański, J.; Teleszko, M.; Sokół-Łętowska, A. Composition and quantification of major polyphenolic compounds, antioxidant activity and colour properties of quince and mixed quince jams. Int. J. Food Sci. Nutr. 2013, 64, 749–756. [Google Scholar] [CrossRef]
- Nowicka, P.; Wojdyło, A.; Teleszko, M.; Samoticha, J. Sensory attributes and changes of physicochemical properties during storage of smoothies prepared from selected fruit. LWT—Food Sci Technol. 2016, 71, 102–109. [Google Scholar] [CrossRef]
- Seglina, D.; Krasnova, I.; Heidemane, G.; Ruisa, S. Influence of drying technology on the quality of dried candied Chaenomeles japonica during storage. Lat. J. Agron. 2009, 12, 113–118. [Google Scholar]
- Krasnova, I.; Seglina, D.; Pole, V. The effect of pre-treatment methods on the quality of dehydrated candied Japanese quince fruits during storage. J. Food Sci. Technol. 2018, 55, 4468–4476. [Google Scholar] [CrossRef]
- Antoniewska, A.; Rutkowska, J.; Martinez Pineda, M. Antioxidative, sensory and volatile profiles of cookies enriched with freeze-dried Japanese quince (Chaenomeles japonica) fruits. Food Chem. 2019, 286, 376–387. [Google Scholar] [CrossRef]
- Nawirska-Olszańska, A.; Biesiada, A.; Sokół-Łętowska, A.; Kucharska, A.Z. Content of biactive compounds and antioxidant capacity of pumpkin puree enriched with Japanese quince Cornellian cherry, strawberry and apples. Acta Sci. Pol. Technol. Aliment. 2011, 10, 51–60. [Google Scholar] [PubMed]
- Hamauzu, Y.; Mizuno, Y. Non-extractable Procyanidins and Lignin are Important Factors in the Bile Acid Binding and Radical Scavenging Properties of Cell Wall Material in some Fruits. Plant Foods Hum. Nutr. 2011, 66, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Hamauzu, Y.; Takedachi, N.; Miyasaka, R.; Makabe, H. Heat treatment of Chinese quince polyphenols increases rat plasma levels of protocatechuic and vanillic acids. Food Chem. 2010, 118, 757–763. [Google Scholar] [CrossRef]
- Cheng, X.-C.; Guo, X.-R.; Qin, Z.; Wang, X.-D.; Liu, H.-M.; Liu, Y.-L. Structural features and antioxidant activities of Chinese quince (Chaenomeles sinensis) fruits lignin during auto-catalyzed ethanol organosolv pretreatment. Int. J. Biol. Macromol. 2020, 164, 4348–4358. [Google Scholar] [CrossRef] [PubMed]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P. Comprehensive characterization of Chaenomeles seeds as a potential source of nutritional and biologically active compounds. J. Food Comp. Anal. 2021, 102, e104065. [Google Scholar] [CrossRef]
- Byczkiewicz, S.; Szwajgier, D.; Kobus-Cisowska, J.; Szczepaniak, O.; Szulc, P. Comparative examination of bioactive phytochemicals in quince (Chaenomeles) fruits and their in vitro antioxidant activity. EJFA 2021, 3, 293–302. [Google Scholar] [CrossRef]
- Al-Zughbi, I.; Krayem, M. Quince fruit Cydonia oblonga Mill nutritional composition, antioxidative properties, health benefits and consumers preferences towards some industrial quince products: A review. Food Chem. 2022, 393, e133362. [Google Scholar] [CrossRef]
- Mierina, I.; Seržane, R.; Strele, M.; Moskaluka, J.; Seglina, D.; Jure, M. Extracts of Japanese quince seeds—Potential source of antioxidants. In Proceedings of 6th Baltic Conference on Food Science and Technology: Innovations for Food Science and Production; Latvia University of Agriculture: Jelgava, Latvia, 2011; p. 75. [Google Scholar]
- Mierina, I.; Seržane, R.; Strele, M.; Moskaļuka, J.; Ivdre, E.; Jure, M. Investigation of the oil and Meal of Japanese Quince (Chaenomeles japonica) Seeds. Proc. Latv. Acad. Sci. Sect. B 2013, 67, 405–410. [Google Scholar] [CrossRef]
- Bolarinwa, I.F.; Orfila, C.; Morgan, M.R. Determination of amygdalin in apple seeds, fresh apples and processed apple juices. Food Chem. 2015, 170, 437–442. [Google Scholar] [CrossRef]
- Górnaś, P.; Siger, A.; Juhņeviča, K.; Lācis, G.; Šnē, E.; Segliņa, D. Cold-pressed Japanese quince (Chaenomeles japonica (Thunb.) Lindl. ex Spach) seed oil as a rich source of α-tocopherol, carotenoids and phenolics: A comparison of the composition and antioxidant activity with nine other plant oils. Eur. J. Lipid Sci. Technol. 2014, 116, 563–570. [Google Scholar] [CrossRef]
- Ben-Othman, S.; Bleive, U.; Kaldmäe, H.; Aluvee, A.; Rätsep, R.; Karp, K.; Maciel, L.S.; Herodes, K.; Rinken, T. Phytochemical characterization of oil and protein fractions isolated from Japanese quince (Chaenomeles japonica) wine by-product, LWT—Food Sci. Technol. 2023, 178, e114632. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Ferreres, F.; Domingues, A.L.; Seabra, R.M.; Ferreira, M.A. Phenolic Profile of Quince Fruit (Cydonia oblonga Miller) (Pulp and Peel). J. Agric. Food Chem. 2002, 50, 4615–4618. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Valentão, P.; Ferreres, F.; Seabra, R.M.; Ferreira, M.A. Quince (Cydonia oblonga Miller) Fruit (Pulp, Peel, and Seed) and Jam: Antioxidant Activity. J. Agric. Food Chem. 2004, 52, 4705–4712. [Google Scholar] [CrossRef] [PubMed]
- Fattouch, S.; Caboni, P.; Coroneo, V.; Tuberoso, C.I.G.; Angioni, A.; Dessi, S.; Marzouki, N.; Cabras, P. Antimicrobial activity of tunisian quince (Cydonia oblonga Miller) pulp and peel polyphenolic extracts. J. Agric. Food Chem. 2007, 55, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.; Silva, B.M.; Silva, R.; Valentão, P.; Andrade, P.B.; Bastos, M.L. First report on Cydonia oblonga Miller anticancer potential: Differential antiproliferative effect against human kidney and colon cancer cells. J. Agric. Food Chem. 2010, 58, 3366–3370. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, B.T.; Mitić, S.S.; Stojanović, G.S.; Mitić, M.N.; Kostić, D.A.; Paunović, D.D.; Arsić, B.B.; Pavlović, A.N. Phenolic profiles and metal ions analyses of pulp and peel of fruits and seeds of quince (Cydonia oblonga Mill.). Food Chem. 2017, 232, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Sut, S.; Dall’Acqua, S.; Poloniato, G.; Maggi, F.; Malagoli, M. Preliminary evaluation of quince (Cydonia oblonga Mill.) fruit as extraction source of antioxidant phytoconstituents for nutraceutical and functional food applications. J. Sci. Food Agric. 2019, 99, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.S.; Silva, B.M.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Carvalho, M. Protective effect of quince (Cydonia oblonga Miller) fruit against oxidative hemolysis of human erythrocytes. Food Chem. Toxicol. 2009, 47, 1372–1377. [Google Scholar] [CrossRef]
- Alesiani, D.; Canini, A.; D’Abrosca, B.; DellaGreca, M.; Fiorentino, A.; Mastellone, C.; Pacifico, S. Antioxidant and antiproliferative activities of phytochemicals from Quince (Cydonia vulgaris) peels. Food Chem. 2010, 118, 199–207. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Ferreres, F.; Seabra, R.M.; Beatriz, M.; Oliveira, P.P.; Ferreira, M.A. Composition of Quince (Cydonia oblonga Miller) seeds: Phenolics, organic acids and free amino acids. Nat. Prod. Res. 2005, 19, 275–281. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Seabra, R.M.; Silva, B.M. Organic acids composition of Cydonia oblonga Miller leaf. Food Chem. 2008, 111, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.; Costa, R.M.; Magalhães, A.S.; Pereira, J.A.; Carvalho, M.; Valentão, P.; Andrade, P.B.; Silva, B.M. Targeted metabolites and biological activities of Cydonia oblonga Miller leaves. Food Res. Int. 2012, 46, 496–504. [Google Scholar] [CrossRef]
- Costa, R.M.; Magalhães, A.S.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Carvalho, M.; Silva, B.M. Evaluation of free radical-scavenging and antihemolytic activities of quince (Cydonia oblonga) leaf: A comparative study with green tea (Camellia sinensis). Food Chem. Toxicol. 2009, 47, 860–865. [Google Scholar] [CrossRef]
- Teleszko, M.; Wojdyło, A. Comparison of phenolic compounds and antioxidant potential between selected edible fruits and their leaves, J. Funct. Foods 2015, 14, 736–746. [Google Scholar] [CrossRef]
- Ashrafi, H.; Ghabili, K.; Alihemmati, A.; Jouyban, A.; Shoja, M.M.; Aslanabadi, S.; Adl, F.H.; Ghavimi, H.; Hajhosseini, L. The effect of quince leaf (Cydonia oblonga Miller) decoction on testes in hypercholesterolemic rabbits: A pilot study. Afr. J. Tradit. Complement. Altern. Med. AJTCAM 2013, 10, 277–282. [Google Scholar] [CrossRef]
- Szychowski, P.J.; Lech, K.; Sendra-Nadal, E.; Hernández, F.; Figiel, A.; Wojdyło, A.; Carbonell-Barrachina, A.A. Kinetics, biocompounds, antioxidant activity, and sensory attributes of quinces as affected by drying metod. Food Chem. 2018, 255, 157–164. [Google Scholar] [CrossRef]
- Blanda, G.; Rodriguez-Roque, M.J.; Comandini, P.; Flores-Cordova, M.A.; Salas-Salazar, N.A.; Cruz-Alvarez, O.; Soto-Caballero, M.C. Phenolic profile and physicochemical characterization of quince (Cydonia oblonga Mill) fruits at different maturity index. Not. Bot. Horti. Agrobot. Cluj Napoca 2020, 48, 2306–2315. [Google Scholar] [CrossRef]
- De Bellis, R.; Chiarantini, L.; Potenza, L.; Gorassini, A.; Verardo, G.; De Marco, R.; Benayada, L.; Stocchi, V.; Albertini, M.C.; Fraternale, D. High Production of Secondary Metabolites and Biological Activities of C. oblonga Mill. Pulp Fruit Callus. J. Funct. Foods 2022, 94, 105133. [Google Scholar] [CrossRef]
- Andrade, P.B.; Carvalho, A.R.F.; Seabra, R.M.; Ferreira, M.A. A previous study of phenolic profiles of quince, pear, and apple purees by HPLC diode array detection for the evaluation of quince puree genuineness. J. Agric. Food Chem. 1998, 46, 968–972. [Google Scholar] [CrossRef]
- Hamauzu, Y.; Yasui, H.; Inno, T.; Kume, C.; Omanyuda, M. Phenolic profile, antioxidant property, and anti-influenza viral activity of Chinese quince (Pseudocydonia sinensis Schneid.), quince (Cydonia oblonga Mill.), and apple (Malus domestica Mill.) fruits. J. Agric. Food Chem. 2005, 53, 928–934. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Gonçalves, A.C.; Seabra, R.M.; Oliveira, M.B.; Ferreira, M.A. Influence of jam processing upon the contents of phenolics, organic acids and free amino acids in quince fruit (Cydonia oblonga Miller). Eur. Food Res. Technol. 2004, 218, 385–389. [Google Scholar] [CrossRef]
- Baroni, M.V.; Fabani, M.P.; Adan, F.; Podio, N.S.; Wunderlin, D.A. Effect of geographical location, processing and simulated digestion on antioxidant characteristics of quince (Cydonia oblonga). Heliyon 2022, 8, e11435. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.A.; Romero, L.A.; Diaz, R.I. Quality and sensory attributes of apple and quince leathers made without preservatives and with enhanced antioxidant activity. LWT—Food Sci. Technol. 2015, 62, 996–1003. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Bielicki, P. Polyphenolic Composition, Antioxidant Activity, and Polyphenol Oxidase (PPO) Activity of Quince (Cydonia oblonga Miller) Varieties. J. Agric. Food Chem. 2013, 61, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Joshi, V.K.; Rana, J.C. Nutritional Composition and Processed Products of Quince (Cydonia oblonga). J. Asian Nat. Prod. Res. 2011, 2, 354–357. [Google Scholar]
- Urbanavičiūtė, I.; Viškelis, P. Biochemical Composition of Japanese Quince (Chaenomeles japonica) and Its Promising Value for Food, Cosmetic, and Pharmaceutical Industries. In Fruit Industry; Kahramanoğlu, İ., Wan, C., Eds.; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Satora, P.; Sroka, P.; Pogoń, P.; Machalica, J. Chaenomeles japonica, Cornus mas, Morus nigra fruits characteristics and their processing potential. J. Food Sci. Technol. 2014, 51, 3934–3941. [Google Scholar] [CrossRef]
- Hamauzu, Y.; Inno, T.; Kume, C.; Irie, M.; Hiramatsu, K. Antioxidant and Antiulcerative Properties of Phenolics from Chinese Quince, Quince, and Apple Fruits. J. Agric. Food Chem. 2006, 54, 765–772. [Google Scholar] [CrossRef]
- Du, H.; Wu, J.; Li, H.; Zhong, P.-X.; Xu, Y.-J.; Li, C.-H.; Ji, K.-X.; Wang, L.-S. Polyphenols and triterpenes from Chaenomeles fruits: Chemical analysis and antioxidant activities assessment. Food Chem. 2013, 141, 4260–4268. [Google Scholar] [CrossRef]
- Huang, G.-H.; Xi, Z.-X.; Li, J.-L.; Chen, C.; Yang, G.-J.; Sun, L.; Chen, W.-S.; Zhu, H.-Y. Isolation of two new phenolic compounds from the fruit of Chaenomeles speciosa (Sweet) Nakai. Phytochem. Lett. 2013, 6, 526–530. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Lech, K.; Tkacz, K.; Nowicka, P. Influence of different drying methods on the quality of Japanese quince fruit. LWT—Food Sci. Technol. 2019, 114, e108416. [Google Scholar] [CrossRef]
- Lee, M.H.; Son, Y.K.; Han, Y.N. Tissue factor inhibitory flavonoids from the fruits of Chaenomeles sinensis. Arch. Pharm. Res. 2002, 25, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Subedi, L.; Kim, S.Y.; Choi, S.U.; Kim, K.H.; Lee, K.R. Lignan glycosides from the twigs of Chaenomeles sinensis and their biological activities. J. Nat. Prod. 2015, 78, 1174–1178. [Google Scholar] [CrossRef] [PubMed]
- Suh, W.S.; Park, K.J.; Kim, D.H.; Subedi, L.; Kim, S.Y.; Choi, S.U.; Lee, K.R. A biphenyl derivative from the twigs of Chaenomeles speciosa. Bioorg. Chem. 2017, 72, 156–160. [Google Scholar] [CrossRef]
- Xie, X.; Cai, X.; Zhu, S.; Zou, G. Chemical composition and antimicrobial activity of essential oils of Chaenomeles speciosa from China. Food Chem. 2007, 100, 1312–1315. [Google Scholar] [CrossRef]
- Hellín, P.; Vila, R.; Jordán, M.J.; Laencina, J.; Rumpunen, K.; Ros, J.M. Characteristics and Composition of Chaenomeles Fruit Juice. In Japanese Quince—Potential Fruit Crop for Northern Europe. Department of Crop Science; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2003; pp. 127–139. [Google Scholar]
- Gardner, P.T.; White, T.A.C.; McPhail, D.B.; Duthie, G.G. The relative contributions of vitamin C, carotenoids and phenolics to the antioxidant potential of fruit juices. Food Chem. 2000, 68, 471–474. [Google Scholar] [CrossRef]
- Vila, R.; Granados, M.V.; Hellín, P.; Kauppinen, S.; Laencina, J.; Rumpunen, K.; Ros, J.M. Biochemical changes in chaenomeles fruits and fruit juice during ripening and storage. In Japanese Quince—Potential Fruit Crop for Northern Europe; Department of Crop Science, Swedish University of Agricultural Sciences: Uppsala, Sweden, 2003; pp. 159–168. [Google Scholar]
- Urbanavičiūtė, I.; Liaudanskas, M.; Bobinas, Č.; Šarkinas, A.; Rezgienė, A.; Viskelis, P. Japanese Quince (Chaenomeles japonica) as a Potential Source of Phenols: Optimization of the Extraction Parameters and Assessment of Antiradical and Antimicrobial Activities. Foods 2020, 9, 1132. [Google Scholar] [CrossRef]
- Osman, A. Chapter 49—Cold pressed Japanese quince (Chaenomeles japonica (Thunb.) Lindl. ex Spach) seed oil. In Cold Pressed Oils; Ramadan, M.F., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 541–546. [Google Scholar]
- Górnaś, P.; Siger, A.; Segliņa, D. Physicochemical characteristics of the cold-pressed Japanese quince seed oil: New promising unconventional bio-oil from by-products for the pharmaceutical and cosmetic industry. Ind. Crops Prod. 2013, 48, 178–182. [Google Scholar] [CrossRef]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P. UPLC/ESI-Q-TOF-MS analysis of (poly)phenols, tocols and amino acids in Chaenomeles leaves versus in vitro anti-enzyme activities. Ind. Crop. Prod. 2022, 181, e114829. [Google Scholar] [CrossRef]
- Marat, N.; Danowska-Oziewicz, M.; Narwojsz, A. Chaenomeles Species-Characteristics of Plant, Fruit and Processed Products: A Review. Plants 2022, 11, 3036. [Google Scholar] [CrossRef]
- Souci, S.W.; Fachmann, W.; Kraut, H. Food Composition and Nutrition Tables, 7th ed.; MedPharm: Stuttgart, Germany, 2008; pp. 743–1238. [Google Scholar]
- Zhang, H.; Geng, Y.L.; Wang, D.J.; Liu, J.H.; Wang, X.; Du, J.H.; Li, S.B. Research on nutrient components of different species of Chaenomeles speciosa Nakai. Shandong Sc. 2011, 24, 24–27. [Google Scholar]
- Bieniasz, M.; Dziedzic, E.; Kaczmarczyk, E. The effect of storage and processing on vitamin C content in Japanese quince fruit. Folia Hortic. 2017, 29, 83–93. [Google Scholar] [CrossRef]
- Hallmann, E.; Orpel, E.; Rembiałkowska, E. The content of biologically active compounds in some fruits from natural state. Veg. Crop Res. Bull. 2011, 75, 81–90. [Google Scholar] [CrossRef]
- Mezhenskij, V.L. Research, Cultivation and Processing of Japanese Quince, Chaenomeles spp. in Ukraine. Verksamhetsberättelse 1992–1994: Report; Balsgård-Deparment of Horticultural Plant Breeding, Swedish University of Agricultural Sciences: Uppsala, Sweden, 1996; pp. 193–195. [Google Scholar]
- Legua, P.; Serrano, M.; Melgarejo, P.; Valero, D.; Martínez, J.J.; Martínez, R.; Hernández, F. Quality parameters, biocompounds and antioxidant activity in fruits of nine quince (Cydonia oblonga Miller) accessions. Sci. Hortic. 2013, 154, 61–65. [Google Scholar] [CrossRef]
- Najman, K.; Adrian, S.; Hallmann, E.; Sadowska, A.; Buczak, K.; Waszkiewicz-Robak, B.; Szterk, A. Effect of Various Drying Methods on Physicochemical and Bioactive Properties of Quince Fruit (Cydonia oblonga Mill.). Agriculture 2023, 13, 446. [Google Scholar] [CrossRef]
- Najman, K.; Adrian, S.; Sadowska, A.; Świąder, K.; Hallmann, E.; Buczak, K.; Waszkiewicz-Robak, B.; Szterk, A. Changes in Physicochemical and Bioactive Properties of Quince (Cydonia oblonga Mill.) and Its Products. Molecules 2023, 28, 3066. [Google Scholar] [CrossRef] [PubMed]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P. Carotenoids, chlorophylls, vitamin E and amino acid profile in fruits of nineteen Chaenomeles cultivars. J. Food Comp. Anal. 2020, 93, e103608. [Google Scholar] [CrossRef]
- Zhang, R.; Li, S.; Zhu, Z.; He, J. Recent advances in valorization of Chaenomeles fruit: A review of botanical profile, phytochemistry, advanced extraction technologies and bioactivities. Trend. Food Sci. Technol. 2019, 91, 467–482. [Google Scholar] [CrossRef]
- Xu, N.Y.; Kim, J.S.; Kang, S.S.; Son, K.H.; Kim, H.P.; Chang, H.W.; Bae, K. A new acylated triterpene from the roots of Chaenomeles japonica. Chem. Pharm. Bull. 2002, 50, 1124–1125. [Google Scholar] [CrossRef]
- Lesińska, E. Zawartość składników mineralnych w owocach pigwowca. Zesz. Nauk. AR W Krakowie Rol. 1985, 25, 175–183. (In Polish) [Google Scholar]
- Watychowicz, K.; Janda, K.; Jakubczyk, K.; Wolska, J. Chaenomeles—Health Promoting Benefits. Rocz. Panstw. Zakl. Hig. 2017, 68, 217–227. [Google Scholar]
- Lesińska, E. Characteristics of sugars and acids in the fruits of East Asian quince. Die Nahrung 1987, 31, 763–765. [Google Scholar] [CrossRef]
- Rasheed, M.; Hussain, I.; Rafiq, S.; Hayat, I.; Qayyum, A.; Ishaq, S.; Awan, M.S. Chemical composition and antioxidant activity of quince fruit pulp collected from different locations. Int. J. Food Propert. 2018, 21, 2320–2327. [Google Scholar] [CrossRef]
- Hopur, H.; Asrorov, A.M.; Qingling, M.; Yili, A.; Ayupbek, A.; Nannan, P.; Aisa, H.A. HPLC Analysis of Polysaccharides in Quince (Cydonia Oblonga Mill. var. maliformis) Fruit and PTP1B Inhibitory Activity. Nat. Prod. J. 2011, 1, 146–150. [Google Scholar] [CrossRef]
- Ashraf, M.U.; Muhammad, G.; Hussain, M.A.; Bukhari, S.N.A. Cydonia oblonga M., A Medicinal Plant Rich in Phytonutrients for Pharmaceuticals. Front. Pharmacol. 2016, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, A.; Nora, P.; Eva, S.B. A Review of Nutritional Value and Putative Health—Effects of Quince (Cydonia oblonga) Fruit. Int. J. Hortic. Sci. 2013, 3, 29–32. [Google Scholar] [CrossRef]
- Thomas, M.; Crépeau, M.J.C.; Rumpunen, K.; Thibault, J.-F. Dietary fibre and cell-wall polysaccharides in the fruits of Japanese quince (Chaenomeles japonica). LWT—Food Sci. Technol. 2000, 33, 124–131. [Google Scholar] [CrossRef]
- Thomas, M.; Thibault, J.-F. Cell-wall polysaccharides in the fruits of Japanese quince (Chaenomeles japonica): Extraction and preliminary characterisation. Carbohydr. Polym. 2002, 49, 345–355. [Google Scholar] [CrossRef]
- Thomas, M.; Guillemin, F.; Guillon, F.; Thibault, J.F. Pectins in the fruits of Japanese Quince (Chaenomeles japonica). Carbohyd. Polym. 2003, 53, 361–372. [Google Scholar] [CrossRef]
- Hellín, P.; Jordán, M.J.; Vila, R.; Gustafsson, M.; Göransson, E.; Ĺkesson, B.; Gröön, I.; Laencina, J.; Ros, J.M. Processing and products of Japanese quince (Chaenomeles japonica) fruits. In Japanese Quince—Potential Fruit Crop for Northern Europe; Department of Crop Science, Swedish University of Agricultural Sciences: Uppsala, Sweden, 2003; pp. 169–175. [Google Scholar]
- Qin, Z.; Liu, H.-M.; Lv, T.-T.; Wang, X.-D. Structure, rheological, thermal and antioxidant properties of cell wall polysaccharides from Chinese quince fruits. Int. J. Biol. Macromol. 2019, 147, 1146–1155. [Google Scholar] [CrossRef]
- Aliasl, F.; Toliyat, T.; Mohammadi, A.; Minaee, B.; Samadi, N.; Aliasl, J.; Sadeghpour, O. Medicinal Properties of Cydonia Oblonga Mill Fruit (Pulp and Peel) in Iranian Traditional Medicine and Modern Phytothrapy. Trad Integr. Med. 2016, 1, 122–128. [Google Scholar]
- Khoubnasabjafari, M.; Jouyban, A. A review of phytochemistry and bioactivity of quince (Cydonia oblonga Mill.). J. Med. Plant. Res. 2011, 5, 3577–3594. [Google Scholar] [CrossRef]
- Yang, G.; Fen, W.; Lei, C.; Xiao, W.; Sun, H. Study on determination of pentacyclic triterpenoids in Chaenomeles by HPLC-ELSD. J. Chromatogr. Sci. 2009, 47, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-J.; Jin, D.-N.; Zhou, Y.; Sang, X.-Y.; Zhu, Y.-Y.; He, Y.-J.; Xie, T.-Z.; Dai, Z.; Zhao, Y.-L.; Luo, X.-D. Bioactivity Ingredients of Chaenomeles speciosa against Microbes: Characterization by LC-MS and Activity Evaluation. J. Agric. Food Chem. 2021, 69, 4686–4696. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Zheng, H.; Fu, G.; Yin, M.; Jiang, L.; Zhao, Y.; Zha, L.; Chu, S.; Peng, H.; Huang, L. Integrated untargeted metabolome, full-length sequencing, and transcriptome analyses reveal insights into the fruit quality at different harvest times of Chaenomeles speciosa. Food Res. Int. 2023, 164, 112314. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.M.; Kim, D.H.; Subedi, L.; Khan, Z.; Choi, S.U.; Kim, S.Y.; Kim, C.S. Chemical constituents of Chaenomeles sinensis twigs and their biological activity. Beilstein J. Org. Chem. 2020, 16, 3078–3085. [Google Scholar] [CrossRef] [PubMed]
- Oku, H.; Ueda, Y.; Ishiguro, K. Antipruritic effects of the fruits of Chaenomeles sinensis. Biol. Pharm. Bull. 2003, 26, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Lee, J.-S.; Yun, C.-Y.; Kim, D.-H.; Kim, I.S. Chinese quince (Chaenomeles sinensis) extract inhibits cell migration and cytokine release in HMC-1 cells. Food Sci. Biotechnol. 2013, 22, 501–506. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 1794427, Chlorogenic Acid; CID 5280633, Neochlorogenic Acid; CID 9798666, Cryptochlorogenic Acid; CID 64945, Ursolic Acid, and CID 10494, Oleanolic Acid. 2023. Available online: http://pubchem.ncbi.nlm.nih.gov (accessed on 21 December 2023).
- Baroni, M.V.; Gastaminza, J.; Podio, N.S.; Lingua, M.S.; Wunderlin, D.A.; Rovasio, J.L.; Dotti, R.; Rosso, J.C.; Ghione, S.; Ribotta, P.D. Changes in the antioxidant properties of quince fruit (Cydonia oblonga Miller) during jam production at industrial scale. J. Food Qual. 2018, 2018, e1460758. [Google Scholar] [CrossRef]
- Essafi-Benkhadir, K.; Refai, A.; Riahi, I.; Fattouch, S.; Karoui, H.; Essafi, M. Quince (Cydonia oblonga Miller) peel polyphenols modulate LPS-induced inflammation in human THP-1-derived macrophages through NF-κB, p38MAPK and Akt inhibition. Biochem. Biophys. Res. Commun. 2012, 418, 180–185. [Google Scholar] [CrossRef]
- García-Alonso, M.; de Pascual-Teresa, S.; Santos-Buelga, C.; Rivas-Gonzalo, J. Evaluation of the antioxidant properties of fruits. Food Chem. 2004, 84, 13–18. [Google Scholar] [CrossRef]
- Yildirim, A.; Oktay, M.; Bilaloglu, V. The antioxidant activity of the leaves of Cydonia vulgaris. Turk. J. Med. Sci. 2001, 31, 23–27. [Google Scholar]
- Pacifico, S.; Gallicchio, M.; Fiorentino, A.; Fischer, A.; Meyer, U.; Stintzing, F.C. Antioxidant properties and cytotoxic effects on human cancer cell lines of aqueous fermented and lipophilic quince (Cydonia oblonga Mill.) preparations. Food Chemical Toxicol. 2012, 50, 4130–4135. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.T.; Kim, B.J.; Kim, J.H.; Heo, M.Y.; Kim, H.P. Biological screening of 100 plant extracts for cosmetic use (I): Inhibitory activities of tyrosinase and DOPA auto-oxidation. Int. J. Cosmet. Sci. 1997, 19, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yu, X.; Mi, M.; Zhao, J.; Wang, J.; Zhang, T. Antioxidative property and antiatherosclerotic effects of the powder processed from Chaenomeles speciosa in APOE-/- mice. J. Food Biochem. 2010, 34, 535–548. [Google Scholar] [CrossRef]
- Deng, Y.; Huang, L.; Zhang, C.; Xie, P.; Cheng, J.; Wang, X.; Liu, L. Skin-care functions of peptides prepared from Chinese quince seed protein: Sequences analysis, tyrosinase inhibition and molecular docking study. Ind. Crops Prod. 2020, 148, e112331. [Google Scholar] [CrossRef]
- Ma, B.; Wang, J.; Tong, J.; Zhou, G.; Chen, Y.; He, J.; Wang, Y. Protective effects of Chaenomeles thibetica extract against carbon tetrachloride-induced damage via the MAPK/Nrf2 pathway. Food Funct. 2016, 7, 1492–1500. [Google Scholar] [CrossRef]
- Sancheti, S.; Sancheti, S.; Bafna, M.; Seo, S.Y. Antihyperglycemic, antihyperlipidemic, and antioxidant effects of Chaenomeles sinensis fruit extract in streptozotocin-induced diabetic rats. Eur. Food Res. Technol. 2010, 231, 415–421. [Google Scholar] [CrossRef]
- Kawahara, T.; Iizuka, T. Inhibitory effect of hotwater extract of quince (Cydonia oblonga) on immunoglobulin E-dependent late-phase immune reactions of mast cells. Cytotechnology 2011, 63, 143–152. [Google Scholar] [CrossRef]
- Shinomiya, F.; Hamauzu, Y.; Kawahara, T. Anti-allergic effect of a hot-water extract of quince (Cydonia oblonga). Biosci. Biotechnol. Biochem. 2009, 73, 1773–1778. [Google Scholar] [CrossRef]
- Gründemann, C.; Papagiannopoulos, M.; Lamy, E.; Mersch-Sundermann, V.; Huber, R. Immunomodulatory properties of a lemon-quince preparation (Gencydo®) as an indicator of anti-allergic potency. Phytomedicine 2011, 18, 760–768. [Google Scholar] [CrossRef]
- Chojnacka, K.; Owczarek, K.; Caban, M.; Caban, M.; Sosnowska, D.; Polka, D.; Koziołkiewicz, M.; Fichna, J.; Lewandowska, U. Japanese quince (Chaenomeles japonica) leaf phenol extract as modulator of the inflammatory response in lipopolysaccharide-triggered murine macrophage raw 264.7 cells. J. Physiol. Pharmacol. 2020, 71, 833–843. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, Y.X.; Liu, A.L.; Wang, H.D.; Wang, Y.L.; Du, G.H. Antioxidant, anti-inflammatory and anti-influenza properties of components from Chaenomeles speciosa. Molecules 2010, 15, 8507–8517. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, Y.-B.; Yang, Q.; Sun, L.-N.; Chen, W.-S. Anti-Inflammatory and Analgesic Activities of Chaenomeles speciosa Fractions in Laboratory Animals. J. Med. Food. 2009, 12, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Strugała, P.; Cyboran-Mikołajczyk, S.; Dudra, A.; Mizgier, P.; Kucharska, A.Z.; Olejniczak, T.; Gabrielska, J. Biological activity of Japanese quince extract and its interactions with lipids, erythrocyte membrane, and human albumin. J. Membr. Biol. 2016, 249, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wei, W. Effects and mechanisms of glucosides of chaenomeles speciosa on collagen-induced arthritis in rats. Int. Immunopharmacol. 2003, 3, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.P.; Dai, M.; Wang, H.; Zhang, L.L.; Wei, W. Antinociceptive effect of glucosides of Chaenomeles speciosa. Chin. J. Pharmacol. Toxicol. 2005, 19, 169–174. [Google Scholar]
- Huang, D.; Jiang, S.; Du, Z.; Chen, Y.; Xue, D.; Wang, X.; Li, M.; Zhang, F.; Chen, W.; Sun, L. Analgesic and AntiArthritic Activities of Polysaccharides in Chaenomeles speciosa. Front. Pharmacol. 2022, 13, e744915. [Google Scholar] [CrossRef]
- Riahi-Chebbi, I.; Haoues, M.; Essafi, M.; Zakraoui, O.; Fattouch, S.; Karoui, H.; Essafi-Benkhadir, K. Quince peel polyphenolic extract blocks human colon adenocarcinoma LS174 cell growth and potentiates 5-fluorouracil efficacy. Cancer Cell Int. 2015, 16, e1. [Google Scholar] [CrossRef]
- Adiban, H.H.; Shirazi, F.; Gholami, S.; Kamalinejad, M.; Hosseini, S.H.; Noubarani, M.; Eskandari, M.R. Chemopreventive effect of quince (Cydonia oblonga Mill.) fruit extract on hepatocellular carcinoma induced by diethylnitrosamine in rats. Int. Pharm. Acta 2019, 2, e2. [Google Scholar] [CrossRef]
- Strek, M.; Gorlach, S.; Podsędek, A.; Sosnowska, D.; Koziołkiewicz, M.; Hrabec, Z.; Hrabec, E. Procyanidin oligomers from Japanese quince (Chaenomeles japonica) fruit inhibit activity of MMP-2 and MMP-9 metalloproteinases. J. Agric. Food Chem. 2007, 55, 6447–6452. [Google Scholar] [CrossRef]
- Lewandowska, U.; Szewczyk, K.; Owczarek, K.; Hrabec, Z.; Podsędek, A.; Koziołkiewicz, M.; Hrabec, E. Flavanols from Japanese quince (Chaenomeles japonica) fruit inhibit human prostate and breast cancer cell line invasiveness and cause favorable changes in Bax/Bcl-2 mRNA Ratio. Nutr. Cancer 2013, 65, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Owczarek, K.; Hrabec, E.; Fichna, J.; Sosnowska, D.; Koziołkiewicz, M.; Szymański, J.; Lewandowska, U. Flavanols from Japanese quince (Chaenomeles japonica) fruit suppress expression of cyclooxygenase-2, metalloproteinase-9, and nuclear factor-kappaB in human colon cancer cells. Acta Biochim. Pol. 2017, 64, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Zvikas, V.; Urbanaviciute, I.; Bernotiene, R.; Kulakauskiene, D.; Morkunaite, U.; Balion, Z.; Majiene, D.; Liaudanskas, M.; Viskelis, P.; Jekabsone, A.; et al. Investigation of Phenolic Composition and Anticancer Properties of Ethanolic Extracts of Japanese Quince Leaves. Foods 2020, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Sosnowska, D.; Polka, D.; Owczarek, K.; Gorlach-Lira, K.; Oliveira de Verasa, B.; Lewandowska, U. Comparison of phenolic compounds, antioxidant and cytotoxic activity of extracts prepared from Japanese quince (Chaenomeles japonica L.) leaves. J. Physiol. Pharmacol. 2020, 71, 213–222. [Google Scholar] [CrossRef]
- Gao, H.Y.; Wu, L.J.; Kuroyanagi, M.; Harada, K.; Kawahara, N.; Nakane, T.; Umehara, K.; Hirasawa, A.; Nakamura, Y. Antitumor-promoting constituents from Chaenomeles sinensis Koehne and their activities in JB6 mouse epidermal cells. Chem. Pharm. Bull. 2003, 51, 1318–1321. [Google Scholar] [CrossRef] [PubMed]
- Abliz, A.; Aji, Q.; Abdusalam, E.; Sun, X.; Abdurahman, A.; Zhou, W.; Moore, N.; Umar, A. Effect of Cydonia oblonga Mill. Leaf extract on serum lipids and liver function in a rat model of hyperlipidaemia. J. Ethnopharmacol. 2014, 151, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Umar, A.; Iskandar, G.; Aikemu, A.; Yiming, W.; Zhou, W.; Berké, B.; Begaud, B.; Moore, N. Effects of Cydonia oblonga Miller leaf and fruit flavonoids on blood lipids and anti-oxydant potential in hyperlipidemia rats. J. Ethnopharmacol. 2015, 169, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Abdusalam, E.; Abliz, P.; Reyim, N.; Tian, S.; Aji, Q.; Issak, M.; Iskandar, G.; Moore, N.; Umar, A. Effect of Cydonia oblonga Mill. fruit and leaf extracts on blood pressure and blood rheology in renal hypertensive rats. J. Ethnopharmacol. 2014, 152, 464–469. [Google Scholar] [CrossRef]
- Zhou, W.; Abdurahman, A.; Abdusalam, E.; Yiming, W.; Abliz, P.; Aji, Q.; Issak, M.; Iskandar, G.; Moore, N.; Umar, A. Effect of Cydonia oblonga Mill. Leaf extracts or captopril on blood pressure and related biomarkers in renal hypertensive rats. J. Ethnopharmacol. 2014, 153, 635–640. [Google Scholar] [CrossRef]
- Zhou, W.; Abdurahman, A.; Umar, A.; Iskander, G.; Abdusalam, E.; Berké, B.; Bégaud, B.; Moore, N. Effects of Cydonia oblonga Miller extracts on blood hemostasis, coagulation and fibrinolysis in mice, and experimental thrombosis in rats. J. Ethnopharmacol. 2014, 154, 163–169. [Google Scholar] [CrossRef]
- Abulizi, A.; Simayi, J.; Nuermaimaiti, M.; Han, M.; Hailati, S.; Talihati, Z.; Maihemuti, N.; Nuer, M.; Khan, N.; Abudurousuli, K.; et al. Quince extract resists atherosclerosis in rats by down-regulating the EGFR/PI3K/Akt/GSK-3β pathway. Biomed. Pharmacother. 2023, 160, 114330. [Google Scholar] [CrossRef]
- Gholami, S.; Hosseini, M.J.; Jafari, L.; Omidvar, F.; Kamalinejad, M.; Mashayekhi, V.; Hosseini, S.H.; Kardan, A.; Pourahmad, J.; Eskandari, M.R. Mitochondria as a Target for the Cardioprotective Effects of Cydonia oblonga Mill. and Ficus carica L. in Doxorubicin-Induced Cardiotoxicity. Drug Res. 2017, 67, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Hanan, E.; Hasan, N.; Zahiruddin, S.; Ahmad, S.; Sharma, V.; Ahmad, F.J. Metabolite profiling and Ameliorative effect of quince (Cydonia oblonga) leaves against doxorubicin induced cardiotoxicity in Wistar rats. Food Biosci. 2023, 53, 102691. [Google Scholar] [CrossRef]
- Lee, M.H.; Han, Y.N. A new in vitro tissue factor inhibitory triterpene from the fruits of Chaenomeles sinensis. Planta Med. 2003, 69, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Xie, L.; Xin, X.; Aisa, H. Anti-diabetic action of Cydonia oblonga seed extract: Improvement of glucose metabolism via activation of PI3K/AKT signaling pathway. J. Pharmacogn. Phytochem. 2016, 4, 7–13. [Google Scholar]
- Lee, H.S.; Jeon, Y.E.; Jung, J.I.; Kim, S.M.; Hong, S.H.; Lee, J.; Hwang, J.S.; Hwang, M.O.; Kwon, K.; Kim, E.J. Anti-obesity effect of Cydonia oblonga Miller extract in high-fat diet-induced obese C57BL/6 mice. J. Funct. Foods 2022, 89, e104945. [Google Scholar] [CrossRef]
- Aslan, M.; Orhan, N.; Orhan, D.D.; Ergun, F. Hypoglycemic activity and antioxidant potential of some medicinal plants traditionally used in Turkey for diabetes. J. Ethnopharmacol. 2010, 128, 384–389. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Pawlik, N. Japanese quince (Chaenomeles japonica L.) fruit polyphenolic extract modulates carbohydrate metabolism in HepG2 cells via AMP-activated protein kinase. Acta Biochim. Pol. 2018, 65, 67–78. [Google Scholar] [CrossRef]
- Sancheti, S.S.; Sancheti, S.; Seo, S. Chaemnomeles sinensis: A potent α- and β-glucosidase inhibitor. Am. J. Pharmacol. Toxicol. 2009, 4, 8–11. [Google Scholar] [CrossRef]
- Sancheti, S.; Sancheti, S.; Seo, S.Y. Antidiabetic and antiacetylcholinesterase effects of ethyl acetate fraction of Chaenomeles sinensis (Thouin) Koehne fruits in streptozotocin-induced diabetic rats. Exp. Toxicol. Pathol. 2013, 65, 55–60. [Google Scholar] [CrossRef]
- Alizadeh, H.; Rahnema, M.; Semnani, S.N.; Hajizadeh, N. Detection of compounds and antibacterial effect of quince (Cydonia oblonga Miller) extracts in vitro and in vivo. J. Biol. Active Prod. Nature 2013, 3, 303–309. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, S.B.; Singh, S. COVID-19: Environment concern and impact of Indian medicinal system. J. Environ. Chem. Eng. 2020, 8, 104–144. [Google Scholar] [CrossRef] [PubMed]
- Naeimi, M.; Kianifar, H.; Memariani, Z.; Kamalinejad, M.; Bijani, A.; Saghebi, R.; Gorji, N. Comparison of the efficacy of ranitidine and quince syrup on gastroesophageal reflux disease in children. BMC Complement. Med. Ther. 2019, 45, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, A.; Hashempur, M.H.; Mojibian, M.; Aliasl, F.; Bioos, S.; Nejatbakhsh, F. A comparative study of ranitidine and quince (Cydonia oblonga mill) sauce on gastroesophageal reflux disease (GERD) in pregnancy: A randomised, open-label, active-controlled clinical trial. J. Obstet. Gynaecol. 2018, 38, 899–905. [Google Scholar] [CrossRef]
- Zohalinezhad, M.E.; Imanieh, M.H.; Samani, S.M.; Mohagheghzadeh, A.; Dehghani, S.M.; Haghighat, M.; Akbarzadeh, A.R. Effects of Quince syrup on clinical symptoms of children with symptomatic gastroesophageal reflux disease: A double-blind randomized controlled clinical trial. Complement. Ther. Clin. Pract. 2015, 21, 268–276. [Google Scholar] [CrossRef]
- Rahi, P.; Mirghafourvand, M.; Mohammad-Alizadeh-Charandabi, S.; Javadzadeh, Y.; Seidi, S. Comparison of the effect of mefenamic acid and quince on the level of menstrual bleeding and hemoglobin: A randomized controlled clinical trial. Eur. J. Integr. Med. 2016, 8, 67–72. [Google Scholar] [CrossRef]
- Din Ganaie, M.U.; Behl, T.; Nijhawan, P.; Sachdeva, M.; Khan, N. Investigation of anti-depressant effect of aqueous and ethanolic extract of Cydonia oblonga in rats. Obes. Med. 2020, 18, e100202. [Google Scholar] [CrossRef]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Villanueva-Cañongo, C.; Hernández-Carlos, B. Antioxidant Compounds and Their Antioxidant Mechanism. In Antioxidants; Shalaby, E., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Mandal, A.; Hossain, U.; Sil, P.C. Chapter 5.2—Antioxidants and cardiovascular diseases. In Antioxidants Effects in Health; Nabavi, S.M., Sanches Silva, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 613–640. [Google Scholar]
- Zekrumah, M.; Begua, P.B.; Razak, A.S.; Wahab, J.; Moffo, N.; Ivane, A.; Oman, M.H.; Elrashied, H.T.; Zou, X.; Zhang, D. Dietary polyphenols—Role in non-communicable chronic disease prevention, and interactions in food systems. An overview. Nutrition 2023, 112, e112034. [Google Scholar] [CrossRef]
- Fiorentino, A.; D’Abrosca, B.; Pacifico, S.; Mastellone, C.; Piscopo, V.; Monaco, P. Spectroscopic identification and antioxidant activity of glucosylated carotenoid metabolites from Cydonia vulgaris fruits. J. Agric. Food Chem. 2006, 54, 9592–9597. [Google Scholar] [CrossRef]
- Fiorentino, A.; D’Abrosca, B.; Pacifico, S.; Mastellone, C.; Piccolella, S.; Monaco, P. Isolation, structure elucidation, and evaluation of cydonioside A, an unusual terpenoid from the fruits of Cydonia vulgaris. Chem. Biodivers. 2007, 4, 973–979. [Google Scholar] [CrossRef]
- Fiorentino, A.; D’Abrosca, B.; Pacifico, S.; Mastellone, C.; Piscopo, V.; Caputo, R.; Monaco, P. Isolation and structure elucidation of antioxidant polyphenols from quince (Cydonia vulgaris) peels. J. Agric. Food Chem. 2008, 56, 2660–2667. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.M.; Valentao, P.; Seabra, R.M.; Andrade, P.P. Quince (Cydonia oblonga Miller): An interesting source of bioactive compounds. In Food Chemistry Research Development; Papadopoulos, K.N., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2008; pp. 243–266. [Google Scholar]
- Devkota, H.P.; Dirar, A.I.; Hassan, M.M.; Logesh, R. Chapter 10—Cydonia oblonga Mill. In Himalayan Fruits and Berries, Bioactive Compounds, Uses and Nutraceutical Potential; Belwal, T., Bhatt, I., Devkota, H.P., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 91–99. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Seabra, R.M.; Silva, B.M. Phenolic profile of Cydonia oblonga Miller leaves. J. Agric. Food Chem. 2007, 55, 7926–7930. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.E.; Frederiksen, H.; Struntze Krogholm, K.; Poulsen, L. Dietary proanthocyanidins: Occurrence, dietary intake, bioavailability, and protection against cardiovascular disease. Mol. Nutr. Food Res. 2005, 49, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Itoh, S.; Yamaguchi, M.; Shigeyama, K.; Sakaguchi, I. The Anti-Aging Potential of Extracts from Chaenomeles sinensis. Cosmetics 2019, 6, 21. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.-M.; Qin, G.-Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, S.P.; Wang, Y.Q.; Shi, L.; Zhang, Z.K.; Dong, F.; Li, H.R.; Zhang, J.Y.; Man, Y.Q. Comparative metabolism study on chlorogenic acid, cryptochlorogenic acid and neochlorogenic acid using UHPLC-Q-TOF MS coupled with network pharmacology. Chin. J. Nat. Med. 2021, 19, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Han, L.Y.; Zhang, H.; Xin, H.L. Chaenomeles speciosa: A review of chemistry and pharmacology. Biomed. Rep. 2014, 2, 12–18. [Google Scholar] [CrossRef]
- Fallon, C.M.; Quach, A.; Lajczak-McGinley, N.; O’Toole, A.; Barrett, K.E.; Sheridan, H.; Keely, S.J. Pentacyclic triterpenes modulate farnesoid X receptor expression in colonic epithelial cells: Implications for colonic secretory function. J. Biol. Chem. 2022, 298, 102569. [Google Scholar] [CrossRef]
- Xie, X.; Zou, G.; Li, C. Antitumor and immunomodulatory activities of a water-soluble polysaccharide from Chaenomeles speciosa. Carbohydr. Polym. 2015, 132, 323–329. [Google Scholar] [CrossRef]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on natural compounds in chemoprevention and treatment of cancer: An update with new promising compounds. Eur. J. Cancer. 2021, 149, 165–183. [Google Scholar] [CrossRef]
- Shao, J.; Fang, Y.; Zhao, R.; Chen, F.; Yang, M.; Jiang, J.; Chen, Z.; Yuan, X.; Jia, L. Evolution from small molecule to nano-drug delivery systems—An emerging approach for cancer therapy of ursolic acid. Asian J. Pharm. Sci. 2020, 15, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.Y.; Li, Y.; Tang, Y.T.; Ma, X.D.; Tang, Z.Y. Anticancer activity of oleanolic acid and its derivatives: Recent advances in evidence, target profiling and mechanisms of action. Biomed. Pharmacother. 2022, 145, e112397. [Google Scholar] [CrossRef] [PubMed]
- Ooi, K.X.; Poo, C.L.; Subramaniam, M.; Cordell, G.A.; Lim, Y.M. Maslinic acid exerts anticancer effects by targeting cancer hallmarks. Phytomedicine 2023, 110, e154631. [Google Scholar] [CrossRef] [PubMed]
- Villani, V.; Di Marco, G.; Iacovelli, F.; Pietrucci, D.; Canini, A.; Gismondi, A. Profile and potential bioactivity of the miRNome and metabolome expressed in Malva sylvestris L. leaf and flower. BMC Plant Biol. 2023, 23, 439. [Google Scholar] [CrossRef]
- Gezici, S.; Sekeroglu, N. Regulation of MicroRNAs by Natural Products and Bioactive Compounds Obtained from Common Medicinal Plants: Novel Strategy in Cancer Therapy. Indian J. Pharm. Educ. Res. 2017, 51, s483–s488. [Google Scholar] [CrossRef]
- Soyturk, A.; Sen, F.; Uncu, A.T.; Celik, I.; Uncu, A.O. De novo assembly and characterization of the first draft genome of quince (Cydonia oblonga Mill.). Sci. Rep. 2021, 11, 3818. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, M.; Li, D. Oleanolic acid suppresses the proliferation of lung carcinoma cells by miR-122/Cyclin G1/MEF2D axis. Mol. Cell. Biochem. 2015, 400, 1–7. [Google Scholar] [CrossRef]
- The Lancet (editorial) Hypertension: Uncontrolled and conquering the world. Lancet 2007, 370, 539. [CrossRef]
- He, H.; Wei, Q.; Chang, J.; Yi, X.; Yu, X.; Luo, G.; Li, X.; Yang, W.; Long, Y. Exploring the hypoglycemic mechanism of chlorogenic acids from Pyrrosia petiolosa (Christ) Ching on type 2 diabetes mellitus based on network pharmacology and transcriptomics strategy. J. Ethnopharmacol. 2023, 322, 117580. [Google Scholar] [CrossRef]
- Loza-Rodríguez, H.; Estrada-Soto, S.; Alarcón-Aguilar, F.J.; Huang, F.; Aquino-Jarquín, G.; Fortis-Barrera, Á.; Giacoman-Martínez, A.; Almanza-Pérez, J.C. Oleanolic acid induces a dual agonist action on PPARγ/α and GLUT4 translocation: A pentacyclic triterpene for dyslipidemia and type 2 diabetes. Eur. J. Pharmacol. 2020, 883, 173252. [Google Scholar] [CrossRef]
- Sawai, R.; Kuroda, K.; Shibata, T.; Gomyou, R.; Osawa, K.; Shimizu, K. Anti-influenza virus activity of Chaenomeles sinensis. J. Ethnopharmacol. 2008, 118, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Sawai-Kuroda, R.; Kikuchi, S.; Shimizu, Y.K.; Sasaki, Y.; Kuroda, K.; Tanaka, T.; Yamamoto, T.; Sakurai, K.; Shimizu, K. A polyphenol-rich extract from Chaenomeles sinensis (Chinese quince) inhibits influenza A virus infection by preventing primary transcription in vitro. J. Ethnopharmacol. 2013, 146, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Karar, M.G.E.; Pletzer, D.; Jaiswal, R.; Weingart, H.; Kuhnert, N. Identification, characterization, isolation and activity against Escherichia coli of quince (Cydonia oblonga) fruit polyphenols. Food Res. Int. 2014, 65, 121–129. [Google Scholar] [CrossRef]
- Armet, A.M.; Deehan, E.C.; O’Sullivan, A.F.; Mota, J.F.; Field, C.J.; Prado, C.M.; Lucey, A.J.; Walter, J. Rethinking healthy eating in light of the gut microbiome. Cell Host Microbe 2022, 30, 764–785. [Google Scholar] [CrossRef]
- Hamauzu, Y.; Kishida, H.; Yamazaki, N. Gastroprotective property of Pseudocydonia sinensis fruit jelly on the ethanol-induced gastric lesions in rats. J. Funct. Foods 2018, 48, 275–282. [Google Scholar] [CrossRef]
- Schloss, J.; Steel, A. Quince fruit compared to Vitamin B6 for treatment of nausea and vomiting in Pregnancy. Adv. Integr. Med. 2017, 4, 80–81. [Google Scholar] [CrossRef]




| Pulp Compound | Main Extractant | Content | Ref. | |
|---|---|---|---|---|
| 3-CQA (3-O-caffeoylquinic acid) neochlorogenic acid | Acetone | 5.68 | mg/100 g fm | [37] |
| 10.89 | mg/100 g fm | [39] | ||
| Methanol | 68.46 | mg/100 g dm | [36] | |
| 50.00 | mg/100 g fm | [38] | ||
| 100.00 | mg/100 g fm | [41] | ||
| 252.00 | mg/100 g fm | [52] | ||
| Water:acetone | 14.10 | mg/100 g fm | [53] | |
| Water:methanol | 2.87 | mg/100 g fm | [35] | |
| 9.46 | mg/100 g fm | [54] | ||
| 4-CQA (4-O-caffeoylquinic acid) cryptochlorogenic acid | Acetone | 2.29 | mg/100 g fm | [39] |
| Acidified methanol | 7.97 | mg/100 g dm | [36] | |
| 3.01 | mg/100 g fm | [55] | ||
| Water:acetone | 1.50 | mg/100 g fm | [53] | |
| Water:methanol | 0.47 | mg/100 g fm | [35] | |
| 0.85 | mg/100 g fm | [54] | ||
| 5-CQA (5-O-caffeoylquinic acid) chlorogenic acid | Acetone | 15.57 | mg/100 g fm | [37] |
| 15.72 | mg/100 g fm | [39] | ||
| Acidified methanol | 12.83 | mg/100 g fm | [55] | |
| Methanol | 64.88 | mg/100 g dm | [36] | |
| 67.00 | mg/100 g fm | [38] | ||
| 142.00 | mg/100 g fm | [41] | ||
| Water:acetone | 12.30 | mg/100 g fm | [53] | |
| Water:ethanol | 14.53 | mg/100 g fm | [40] | |
| Water:methanol | 8.54 | mg/100 g fm | [35] | |
| 9.00 | mg/100 g fm | [54] | ||
| 3,5-di-CQA (3,5-di-O-caffeoylquinic acid) | Acetone | 13.96 | mg/100 g fm | [39] |
| Methanol | 5.63 | mg/100 g dm | [36] | |
| 7.00 | mg/100 g fm | [41] | ||
| Water:methanol | 0.53 | mg/100 g fm | [35] | |
| 0.93 | mg/100 g fm | [54] | ||
| 3-p-Coumaroylquinic acid | Acidified methanol | 0.44 | mg/100 g fm | [55] |
| 5-p-Coumaroylquinic acid | Acidified methanol | 2.27 | mg/100 g fm | [55] |
| p-Coumaroylquinic acid | Acetone | 1.21 | mg/100 g fm | [39] |
| (+)-Catechin | Acetone | 7.20 | mg/100 g fm | [37] |
| Acidified methanol | 0.06 | mg/100 g fm | [55] | |
| (−)-Catechin | Acetone | 0.18 | mg/100 g fm | [37] |
| Catechin | Water:ethanol | 0.02 | mg/100 g fm | [40] |
| (−)-Epicatechin | Acidified methanol | 2.25 | mg/100 g fm | [55] |
| Epicatechin | Water:ethanol | 0.11 | mg/100 g fm | [40] |
| Kaempferol rutinoside | Acidified methanol | 0.08 | mg/100 g fm | [55] |
| Kaempferol hexoside | Acidified methanol | 0.38 | mg/100 g fm | [55] |
| Phloridzin | Water:ethanol | <0.01 | mg/100 g fm | [40] |
| Procyanidin B1 | Methanol | 2.00 | mg/100 g fm | [52] |
| Water:ethanol | 6.52 | mg/100 g fm | [40] | |
| Procyanidin B2 | Water:acetone | 1.40 | mg/100 g fm | [53] |
| Quercetin | Acidified methanol | 0.37 | mg/100 g fm | [55] |
| Q-3-Gal (quercetin-3-O-galactoside) hyperin | Acidified methanol | 5.64 | mg/100 g fm | [55] |
| Water:methanol | 0.06 | mg/100 g fm | [35] | |
| 0.25 | mg/100 g fm | [54] | ||
| Q-3-G (quercetin-3-O-glucoside) | Methanol | 2.00 | mg/100 g fm | [52] |
| Q-3-R (quercetin-3-O-rutinoside) rutin | Acetone | 9.05 | mg/100 g fm | [37] |
| 0.51 | mg/100 g fm | [39] | ||
| Acidified methanol | 2.27 | mg/100 g fm | [55] | |
| Methanol | 3.30 | mg/100 g dm | [36] | |
| 2.00 | mg/100g fm | [41] | ||
| 146.00 | mg/100 g fm | [52] | ||
| Water:ethanol | 0.005 | mg/100 g fm | [40] | |
| Water:methanol | 0.53 | mg/100 g fm | [35] | |
| 0.33 | mg/100 g fm | [54] |
| Peel Compound | Main Extractant | Content | Ref. | |
|---|---|---|---|---|
| 3-CQA (3-O-caffeoylquinic acid) neochlorogenic acid | Acetone | 3.94 | mg/100 g fm | [37] |
| 26.46 | mg/100 g fm | [39] | ||
| Methanol | 196.64 | mg/100 g dm | [36] | |
| 114.00 | mg/100 g fm | [38] | ||
| 128.00 | mg/100 g fm | [41] | ||
| Water:methanol | 5.55 | mg/100 g fm | [35] | |
| 20.99 | mg/100 g fm | [54] | ||
| 4-CQA (4-O-caffeoylquinic acid) cryptochlorogenic acid | Acetone | 0.51 | mg/100 g fm | [37] |
| 4.82 | mg/100 g fm | [39] | ||
| Methanol | 17.44 | mg/100 g dm | [36] | |
| 18.00 | mg/100 g fm | [38] | ||
| Water:methanol | 0.93 | mg/100 g fm | [35] | |
| 1.92 | mg/100 g fm | [54] | ||
| 5-CQA (5-O-caffeoylquinic acid) chlorogenic acid | Acetone | 12.85 | mg/100 g fm | [37] |
| 36.76 | mg/100 g fm | [39] | ||
| Methanol | 182.94 | mg/100 g dm | [36] | |
| 165.00 | mg/100 g fm | [38] | ||
| 184.00 | mg/100 g fm | [41] | ||
| Water:ethanol | 41.13 | mg/100 g fm | [40] | |
| Water:methanol | 17.95 | mg/100 g fm | [35] | |
| 27.98 | mg/100 g fm | [54] | ||
| 3,5-di-CQA (3,5-O-dicaffeoylquinic acid) | Acetone | 13.96 | mg/100 g fm | [39] |
| Methanol | 9.87 | mg/100 g dm | [36] | |
| 13.00 | mg/100 g fm | [41] | ||
| Water:methanol | 1.63 | mg/100 g fm | [35] | |
| 2.43 | mg/100 g fm | [54] | ||
| (+)-Catechin | Acetone | 5.07 | mg/100 g fm | [37] |
| (–)-Catechin | Acetone | 0.10 | mg/100 g fm | [37] |
| Catechin | Acetone | 3.40 | mg/100 g fm | [39] |
| Water:ethanol | 0.20 | mg/100 g fm | [40] | |
| 4-p-Coumaroylquinic acid | Water:ethanol | 0.11 | mg/100 g fm | [40] |
| p-Coumaroylquinic acid | Acetone | 1.39 | mg/100 g fm | [39] |
| Epicatechin | Water:ethanol | 3.50 | mg/100 g fm | [40] |
| Kaempferol | Acetone | 12.60 | mg/100 g fm | [40] |
| K-3-G (kaempferol-3-O-glucoside) | Acetone | 10.65 | mg/100 g fm | [37] |
| 2.48 | mg/100 g fm | [39] | ||
| Methanol | 8.88 | mg/100 g dm | [36] | |
| 55.00 | mg/100 g fm | [38] | ||
| 34.00 | mg/100 g fm | [41] | ||
| Water:methanol | 3.54 | mg/100 g fm | [35] | |
| 2.58 | mg/100 g fm | [54] | ||
| K-3-Gly (kaempferol-3-O-glycoside | Water:methanol | 3.22 | mg/100 g fm | [54] |
| Methanol | 14.00 | mg/100 g fm | [38] | |
| 25.00 | mg/100 g fm | [41] | ||
| Kaempferol glycoside | Methanol | 11.22 | mg/100 g dm | [36] |
| Kaempferol glycoside acylated with p-coumaric acid A1 | Water:methanol | 1.84 | mg/100 g fm | [54] |
| Kaempferol glycoside acylated with p-coumaric acid A1 | Methanol | 5.38 | mg/100 g dm | [36] |
| Kaempferol glycoside acylated with p-coumaric acid A2 | Water:methanol | 3.45 | mg/100 g fm | [54] |
| Kaempferol glycoside acylated with p-coumaric acid A2 | Methanol | 10.95 | mg/100 g dm | [36] |
| Kaempferol glycoside acylated with p-coumaric acid | Methanol | 24.00 | mg/100 g fm | [38] |
| 5.00 | mg/100 g fm | [41] | ||
| K-3-R (kaempferol-3-O-rutinoside) | Acetone | 3.96 | mg/100 g fm | [37] |
| 1.13 | mg/100 g fm | [39] | ||
| Methanol | 15.22 | mg/100 g dm | [36] | |
| 2.10 | mg/100 g fm | [41] | ||
| Water:methanol | 6.11 | mg/100 g fm | [35] | |
| 5.10 | mg/100 g fm | [54] | ||
| Quercetin | Acetone | 7.01 | mg/100 g fm | [37] |
| Q-3-Gal (quercetin-3-O-galactoside) hyperin | Acetone | 12.40 | mg/100 g fm | [37] |
| 4.46 | mg/100 g fm | [39] | ||
| Methanol | 49.12 | mg/100 g dm | [36] | |
| 329.00 | mg/100 g fm | [38] | ||
| Water:methanol | 10.08 | mg/100 g fm | [35] | |
| 6.07 | mg/100 g fm | [54] | ||
| Q-3-G (quercetin-3-O-glucoside) isoquercitrin | Acetone | 9.23 | mg/100 g fm | [37] |
| Quercetin glycoside acylated with p-coumaric acid A1 | Methanol | 16.69 | mg/100 g dm | [36] |
| Quercetin glycoside acylated with p-coumaric acid A2 | Methanol | 6.57 | mg/100 g dm | [36] |
| Quercetin glycoside acylated with p-coumaric acid | Acetone | 5.92 | mg/100 g fm | [37] |
| Methanol | 22.00 | mg/100 g fm | [38] | |
| 11.00 | mg/100 g fm | [41] | ||
| Water:methanol | 5.20 | mg/100 g fm | [54] | |
| 1.77 | mg/100 g fm | [54] | ||
| Quercetin glycosides acylated with p-coumaric acid | Methanol | 4.00 | mg/100 g fm | [41] |
| Q-3-R (quercetin-3-O-rutinoside) rutin | Acetone | 47.21 | mg/100 g fm | [37] |
| 17.59 | mg/100 g fm | [39] | ||
| Methanol | 177.78 | mg/100 g dm | [36] | |
| 329.00 | mg/100 g fm | [38] | ||
| Water:ethanol | 0.93 | mg/100 g fm | [40] | |
| Water:methanol | 51.73 | mg/100 g fm | [35] | |
| 74.08 | mg/100 g fm | [54] | ||
| Phloridzin | Water:ethanol | >0.01 | mg/100 g fm | [40] |
| Procyanidin B1 | Water:ethanol | 11.02 | mg/100 g fm | [40] |
| Seed Compound | Content | Ref. | |
|---|---|---|---|
| 3-CQA (3-O-caffeoylquinic acid) neochlorogenic acid | 2.40 | mg/100 g dm | [36] |
| 1.00 | mg/100 g fm | [38] | |
| 1.00 | mg/100 g fm | [41] | |
| 4-CQA (4-O-caffeoylquinic acid) cryptochlorogenic acid | 2.76 | mg/100 g dm | [36] |
| 5-CQA (5-O-caffeoylquinic acid) chlorogenic acid | 5.44 | mg/100 g dm | [36] |
| 6.00 | mg/100 g fm | [38] | |
| 5.00 | mg/100 g fm | [41] | |
| 3,5-di-CQA (3,5-di-O-caffeoylquinic acid) | 2.99 | mg/100 g dm | [36] |
| 6-C-Glucosyl-8-C-pentosyl chrysoeriol | 1.61 | mg/100 g dm | [36] |
| 5.00 | mg/100 g fm | [38] | |
| 6.00 | mg/100 g fm | [41] | |
| 6-C-Pentosyl-8-C-glucosyl chrysoeriol | 2.18 | mg/100 g dm | [36] |
| 10.00 | mg/100 g fm | [38] | |
| 3.00 | mg/100 g fm | [41] | |
| Isoschaftoside | 1.71 | mg/100 g dm | [36] |
| 8.00 | gm/100 g fm | [41] | |
| Lucenin-2 | 1.02 | mg/100 g dm | [36] |
| 3.00 | mg/100 g fm | [38] | |
| 3.00 | mg/100 g fm | [41] | |
| Schaftoside | 1.14 | mg/100 g dm | [36] |
| 5.00 | mg/100 g fm | [38] | |
| 6.00 | mg/100 g fm | [41] | |
| Stellarin-2 | 2.76 | mg/100 g dm | [36] |
| 15.00 | mg/100 g fm | [38] | |
| 8.00 | mg/100 g fm | [41] | |
| Vicenin-2 | 1.46 | mg/100 g dm | [36] |
| 7.00 | mg/100 g fm | [38] |
| Activity | Species | Details | Ref. |
|---|---|---|---|
| Antioxidative | C. oblonga | prevention of hematotoxic stress | [44] |
| in vitro effects | [46,55,56,109,110,111,112,113] | ||
| C. japonica | in vitro effects | [16,59,62] | |
| C. speciosa | in vitro effects | [62,114] | |
| increase in GSH-Px activity and antioxidant capacity in mice serum | [115] | ||
| in vitro activity of two peptides RHAKF and NNRYE | [116] | ||
| C. thiberica | in vitro effects | [62] | |
| in vitro effects, increased CAT, SOD, and GSH content in rat serum | [117] | ||
| P. sinensis | in vitro effects | [62] | |
| increase in SOD, GSH, and CAT levels in rat serum | [118] | ||
| Anti-inflammatory Immuno-modulatory | C. oblonga | inhibition of NF-κ98980B and p38MAPK, and Akt activation | [110] |
| IgE-dependent late-phase immune reaction modulation in vitro | [119] | ||
| suppressing IgE production in type I allergy | [120] | ||
| various anti-histamine effects | [121] | ||
| C. japonica | reduction in the expression of IL-1β, IL-6, TNF-α, COX-2, iNOS, NF-κB p65, and p-NF-κB p65 in RAW264.7 cells | [122] | |
| C. speciosa | inhibition of TNF-α production in RAW264.7 cells | [123] | |
| anti-inflammatory effects by standard tests in mice/rats | [124] | ||
| inhibition of COX-1 and COX-2 activities | [125] | ||
| reduction in lymphocyte proliferation, and IL-1, IL-2, and TNF-α production in peritoneal macrophages and synoviocytes | [126] | ||
| reduction in PGE2 and TNF-α concentration in synoviocytes | [127] | ||
| inhibition of TNF- α, IL-1β, and COX-2; JNK and ERK1/2 phosphorylation in NR8383 cells | [128] | ||
| P. sinensis | inhibition of scratching induced by serotonin, platelet-activating factor, and prostaglandin E2 | [106] | |
| inhibition of TNF-α expression by blocking ERK, p38(MAPK), and JNK activation in HMC-1 cells | [107] | ||
| Anticancer | C. oblonga | inhibitory activity toward human colon cancer cells | [38] |
| cytotoxic effects on HepG2, A549, and HeLa cells | [113] | ||
| apoptosis of colon cancer LS174 cells | [129] | ||
| reduced liver damage in hepatocellular carcinoma | [130] | ||
| C. japonica | activation of MMP-2 and MMP-9 secreted by leukemia HL-60 cells | [131] | |
| change in Bax/Bcl-2 ratio in DU145 prostate cancer cells; inhibition of MDA-MB-231 breast cancer cells | [132] | ||
| COX-2 and MMP-9 inhibition, NF-κB expression, anti-metastatic activities towards SW-480 colon cancer cells | [133] | ||
| reducing HROG36 glioma cell viability | [134] | ||
| cytotoxic effect on SW-480 and HT-29 colon cancer cells | [135] | ||
| C. speciosa | inhibition of sarcoma 180 cells by promoting secretion of IL-2, TNF-α, and IFN-γ in serum | [68] | |
| C. thiberica | protective effect on chronic hepatic damage via the MAPK/Nrf2 pathway | [117] | |
| P. sinensis | cytotoxic effect on human anaplastic large cell lymphoma JB6 cells | [136] | |
| Cardio-protective | C. oblonga | lipid profile, blood serum parameter improvement | [137,138] |
| essential and renovascular hypertension reduction | [139] | ||
| renal hypertension reduction | [140] | ||
| anti-thromboxane effect | [141] | ||
| improving the degree of aortic injury and hemodynamic indicators | [142] | ||
| DOX-induced cardiotoxicity alleviation | [143,144] | ||
| C. speciosa | reduction in relative atherosclerotic plaque area of aortic sinus and aortic arch | [115] | |
| P. sinensis | increase in thromboplastin (TF) inhibitory activity | [145] | |
| Antidiabetic | C. oblonga | inhibition of tyrosine phosphatase activity | [112] |
| activating PI3K/AKT insulin signaling in vitro | [146] | ||
| multifactorial anti-obesity effects | [147] | ||
| blood glucose level reduction | [148] | ||
| C. japonica | elevation of gluconeogenesis through modulation of PEPCK, PTP1B, FOXO1, and GLUT2/4 expression | [149] | |
| P. sinensis | hypoglycemic effect, modulation of lipid metabolism | [118] | |
| α- and β-galactosidase inhibitory activities | [150] | ||
| inhibition of glucose transporter, α- and β-glucosidase, and amylase | [151] | ||
| Antibacterial Antiviral Antifungal | C. oblonga | against: S. aureus, P. aeruginosa, E. coli, and yeast C. albicans | [37] |
| against E. aerogenes and E. coli | [152] | ||
| against SARS-CoV-2 virus | [153] | ||
| C. japonica | mainly against E. faccalis, B. subtilis, and S. aureus | [72] | |
| Other | C. oblonga | treatment of gastroesophageal reflux (GARD) | [154,155,156] |
| reducing menstrual bleeding and increasing hemoglobin levels | [157] | ||
| antidepressant activity | [158] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostecka-Gugała, A. Quinces (Cydonia oblonga, Chaenomeles sp., and Pseudocydonia sinensis) as Medicinal Fruits of the Rosaceae Family: Current State of Knowledge on Properties and Use. Antioxidants 2024, 13, 71. https://doi.org/10.3390/antiox13010071
Kostecka-Gugała A. Quinces (Cydonia oblonga, Chaenomeles sp., and Pseudocydonia sinensis) as Medicinal Fruits of the Rosaceae Family: Current State of Knowledge on Properties and Use. Antioxidants. 2024; 13(1):71. https://doi.org/10.3390/antiox13010071
Chicago/Turabian StyleKostecka-Gugała, Anna. 2024. "Quinces (Cydonia oblonga, Chaenomeles sp., and Pseudocydonia sinensis) as Medicinal Fruits of the Rosaceae Family: Current State of Knowledge on Properties and Use" Antioxidants 13, no. 1: 71. https://doi.org/10.3390/antiox13010071
APA StyleKostecka-Gugała, A. (2024). Quinces (Cydonia oblonga, Chaenomeles sp., and Pseudocydonia sinensis) as Medicinal Fruits of the Rosaceae Family: Current State of Knowledge on Properties and Use. Antioxidants, 13(1), 71. https://doi.org/10.3390/antiox13010071