

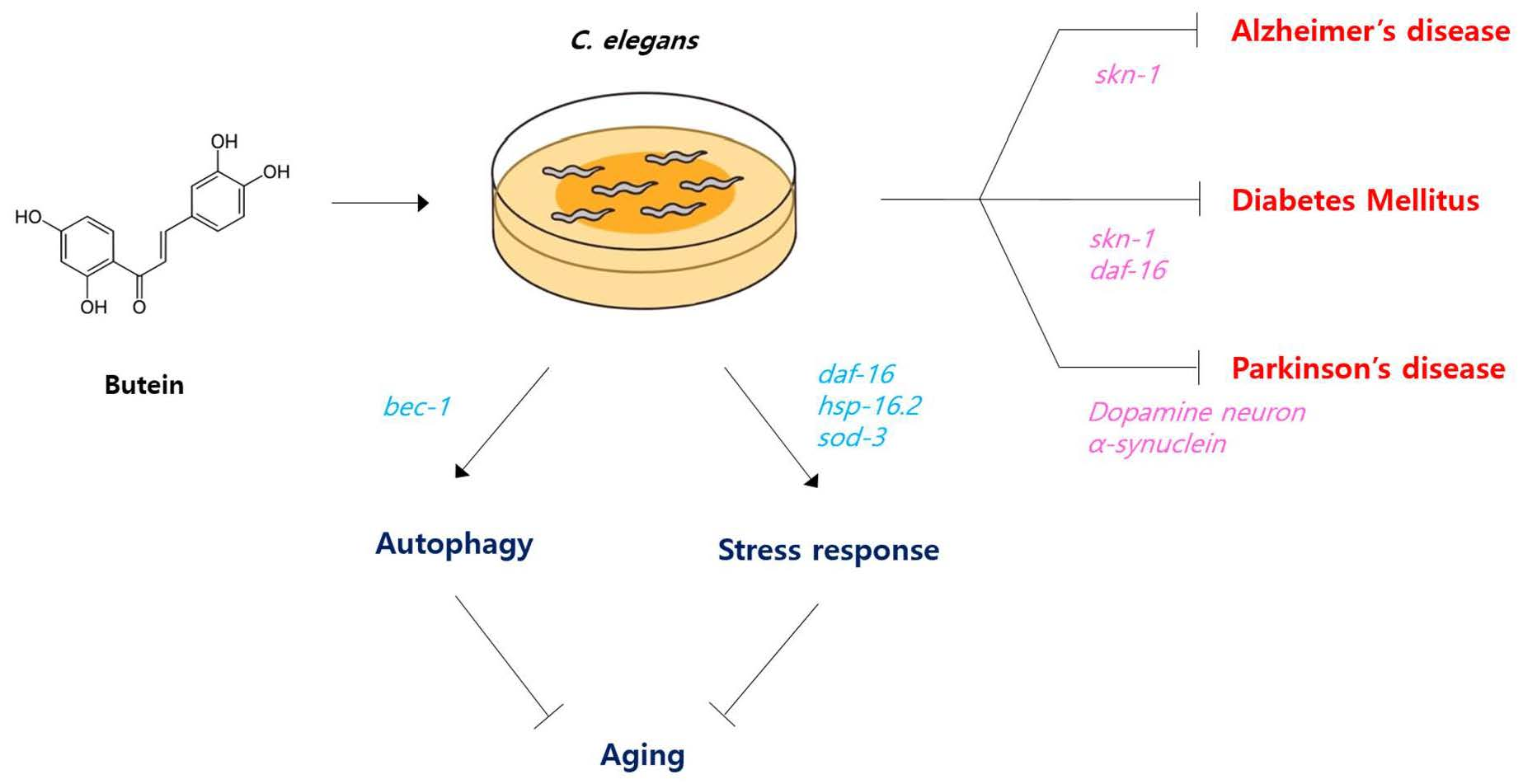
Butein Increases Resistance to Oxidative Stress and Lifespan with Positive Effects on the Risk of Age-Related Diseases in Caenorhabditis elegans


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Worm Strains and Maintenance
2.2. In Vitro Antioxidant Activity
2.3. Resistance to Environmental Stressors
2.4. Lifespan
2.5. Fertility
2.6. Motility
2.7. RNAi
2.8. Subcellular Localization of DAF-16
2.9. Expression of Downstream Targets of DAF-16
2.10. Aβ-Induced Paralysis
2.11. Toxicity Caused by High-Glucose Diet (HGD)
2.12. Genetic Model of PD
3. Results
3.1. Butein Scavenges Free Radicals and Increases Resistance to Oxidative Stress
3.2. Butein Extends the Lifespan of C. elegans but Reduces Fertility
3.3. Butein Delays the Age-Related Decline in Motility
3.4. The Lifespan-Extending Effect of Butein Requires DAF-16 and BEC-1
3.5. DAF-16-Regulated Stress-Responsive Genes Are Induced by Butein
3.6. Butein Prevents Aβ-Induced Toxicity in an AD Model
3.7. Butein Reduces HGD-Induced Mortality in C. elegans
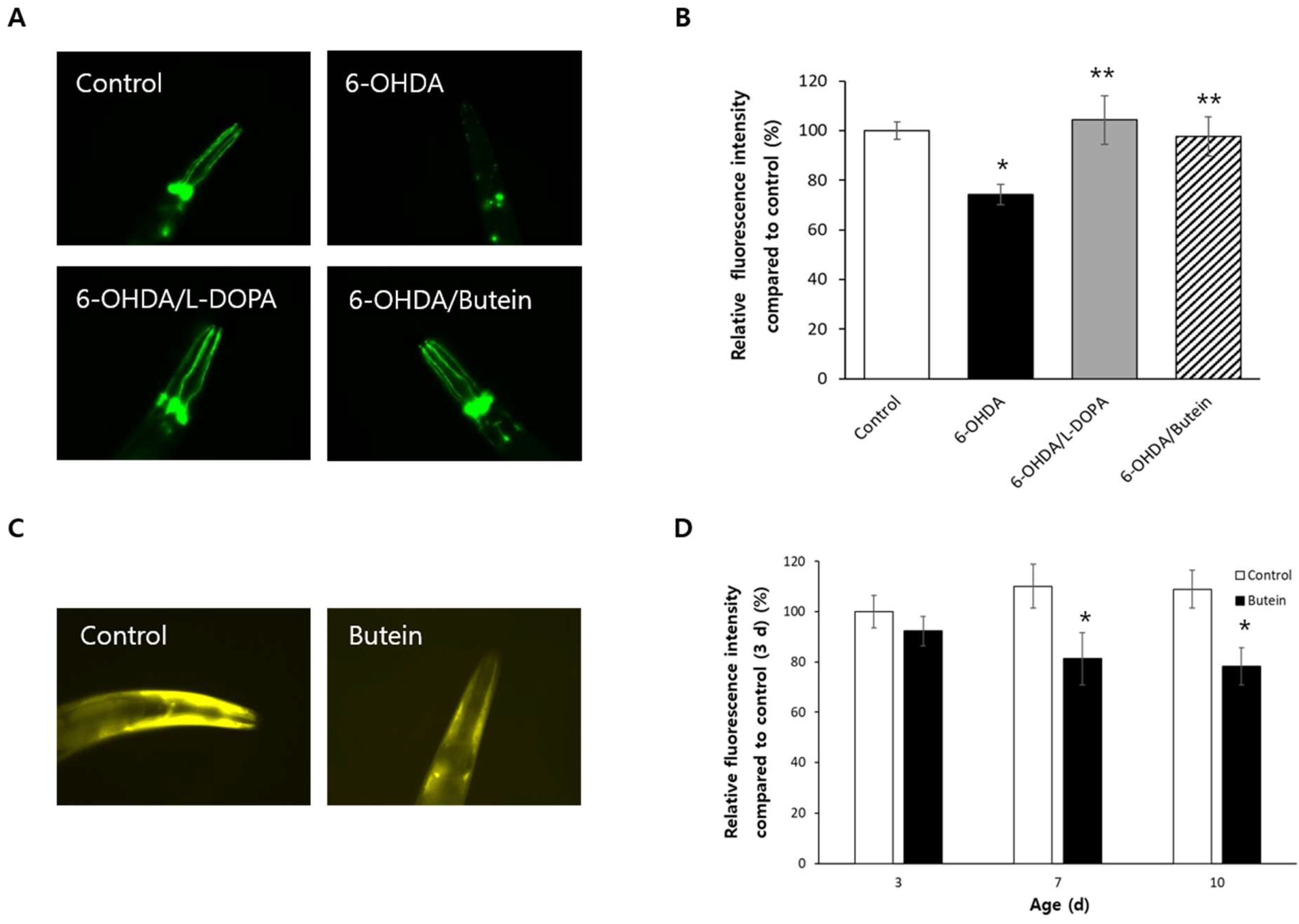
3.8. Butein Inhibits Dopaminergic Neurodegeneration and Accumulation of α-Synuclein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kudryavtseva, A.V.; Krasnov, G.S.; Dmitriev, A.A.; Alekseev, B.Y.; Kardymon, O.L.; Sadritdinova, A.F.; Fedorova, M.S.; Pokrovsky, A.V.; Melnikova, N.V.; Kaprin, A.D.; et al. Mitochondrial Dysfunction and Oxidative Stress in Aging and Cancer. Oncotarget 2016, 7, 44879–44905. [Google Scholar] [CrossRef]
- Finkel, T. Signal Transduction by Reactive Oxygen Species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef]
- Harman, D. Aging: A Theory Based on Free Radical and Radiation Chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef]
- Boengler, K.; Kosiol, M.; Mayr, M.; Schulz, R.; Rohrbach, S. Mitochondria and Ageing: Role in Heart, Skeletal Muscle and Adipose Tissue. J. Cachexia Sarcopenia Muscle 2017, 8, 349–369. [Google Scholar] [CrossRef]
- Aggarwal, V.; Tuli, H.S.; Varol, A.; Thakral, F.; Yerer, M.B.; Sak, K.; Varol, M.; Jain, A.; Khan, M.A.; Sethi, G. Role of Reactive Oxygen Species in Cancer Progression: Molecular Mechanisms and Recent Advancements. Biomolecules 2019, 9, 735. [Google Scholar] [CrossRef]
- Shin, D.H.; Dier, U.; Melendez, J.A.; Hempel, N. Regulation of MMP-1 Expression in Response to Hypoxia Is Dependent on the Intracellular Redox Status of Metastatic Bladder Cancer Cells. Biochim. Biophys. Acta 2015, 1852, 2593–2602. [Google Scholar] [CrossRef]
- Mendez, E.F.; Sattler, R. Biomarker Development for C9orf72 Repeat Expansion in ALS. Brain Res. 2015, 1607, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, B.K.; Park, S.K. Effects of Fisetin, a Plant-Derived Flavonoid, on Response to Oxidative Stress, Aging, and Age-Related Diseases in Caenorhabditis elegans. Pharmaceuticals 2022, 15, 1528. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Park, S.K. Anti-Oxidant and Anti-Aging Effects of Phlorizin Are Mediated by DAF-16-Induced Stress Response and Autophagy in Caenorhabditis elegans. Antioxidants 2022, 11, 1996. [Google Scholar] [CrossRef]
- Zhu, Q.; Qu, Y.; Zhou, X.G.; Chen, J.N.; Luo, H.R.; Wu, G.S. A Dihydroflavonoid Naringin Extends the Lifespan of C. elegans and Delays the Progression of Aging-Related Diseases in PD/AD Models via DAF-16. Oxid. Med. Cell. Longev. 2020, 2020, 6069354. [Google Scholar] [CrossRef]
- Okoro, N.O.; Odiba, A.S.; Osadebe, P.O.; Omeje, E.O.; Liao, G.; Fang, W.; Jin, C.; Wang, B. Bioactive Phytochemicals with Anti-Aging and Lifespan Extending Potentials in Caenorhabditis elegans. Molecules 2021, 26, 7323. [Google Scholar] [CrossRef]
- Zhou, D.D.; Luo, M.; Huang, S.U.; Saimaiti, A.; Shang, A.; Gan, R.U.; Li, H.B. Effects and Mechanisms of Resveratrol on Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef]
- Büchter, C.; Ackermann, D.; Havermann, S.; Honnen, S.; Chovolou, Y.; Fritz, G.; Kampkötter, A.; Wätjen, W. Myricetin-Mediated Lifespan Extension in Caenorhabditis elegans Is Modulated by DAF-16. Int. J. Mol. Sci. 2013, 14, 11895–11914. [Google Scholar] [CrossRef]
- Kumar, J.; Park, K.C.; Awasthi, A.; Prasad, B. Silymarin Extends Lifespan and Reduces Proteotoxicity in C. elegans Alzheimer’s Model. CNS Neurol. Disord. Drug Targets 2015, 14, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Jun, C.Y.; Lee, S.; Park, C.H.; Cho, K.; Ko, S.G. Rhus verniciflua Stokes Extract: Radical Scavenging Activities and Protective Effects on H2O2-Induced Cytotoxicity in Macrophage RAW 264.7 Cell Lines. Biol. Pharm. Bull. 2006, 29, 1603–1607. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Kim, J.H.; Hong, M.H.; Seog, H.M.; Oh, S.H.; Lee, P.J.; Kim, G.J.; Kim, H.M.; Um, J.Y.; Ko, S.G. Phenolic-Rich Fraction from Rhus verniciflua Stokes (RVS) Suppress Inflammatory Response via NF-kappaB and JNK Pathway in Lipopolysaccharide-Induced RAW 264.7 Macrophages. J. Ethnopharmacol. 2007, 110, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Szuster-Ciesielska, A.; Mizerska-Dudka, M.; Daniluk, J.; Kandefer-Szerszeń, M. Butein Inhibits Ethanol-Induced Activation of Liver Stellate Sells through TGF-beta, NFkappaB, p38, and JNK Signaling Pathways and Inhibition of Oxidative Stress. J. Gastroenterol. 2013, 48, 222–237. [Google Scholar] [CrossRef] [PubMed]
- Gay, N.H.; Suwanjang, W.; Ruankham, W.; Songtawee, N.; Wongchitrat, P.; Prachayasittikul, V.; Prachayasittikul, S.; Phopin, K. Butein, Isoliquiritigenin, and Scopoletin Attenuate Neurodegeneration via Antioxidant Enzymes and SIRT1/ADAM10 Signaling Pathway. RSC Adv. 2020, 10, 16593–16606. [Google Scholar] [CrossRef] [PubMed]
- Farias-Pereira, R.; Zhang, Z.; Park, C.S.; Kim, D.; Kim, K.H.; Park, Y. Butein Inhibits Lipogenesis in Caenorhabditis elegans. Biofactors 2020, 46, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Butein Activates Autophagy Through AMPK/TSC2/ULK1/mTOR Pathway to Inhibit IL-6 Expression in IL-1β Stimulated Human Chondrocytes. Cell. Physiol. Biochem. 2018, 49, 932–946. [Google Scholar] [CrossRef]
- Chen, Y.H.; Yeh, C.W.; Lo, H.C.; Su, S.L.; Hseu, Y.C.; Hsu, L.S. Generation of Reactive Oxygen Species Mediates Butein-Induced Apoptosis in Neuroblastoma Cells. Oncol. Rep. 2012, 27, 1233–1237. [Google Scholar] [CrossRef]
- Mendonca, P.; Horton, A.; Bauer, D.; Messeha, S.; Soliman, K.F.A. The Inhibitory Effects of Butein on Cell Proliferation and TNF-alpha-Induced CCL2 Release in Racially Different Triple Negative Breast Cancer Cells. PLoS ONE 2019, 14, e0215269. [Google Scholar] [CrossRef] [PubMed]
- Alshammari, G.M.; Balakrishnan, A.; Chinnasamy, T. Butein Protects the Nonalcoholic Fatty Liver through Mitochondrial Reactive Oxygen Species Attenuation in Rats. Biofactors 2018, 44, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Pan, Q. Butein Inhibits Oxidative Stress Injury in Rats with Chronic Heart Failure via ERK/Nrf2 Signaling. Cardiovasc. Ther. 2022, 2022, 8684014. [Google Scholar] [CrossRef] [PubMed]
- Peto, R.; Peto, J. Asymptotically Efficient Rank Invariant Test Procedures. J. R. Statist. Soc. A 1972, 135, 185–207. [Google Scholar] [CrossRef]
- Kamath, R.S.; Fraser, A.G.; Dong, Y.; Poulin, G.; Durbin, R.; Gotta, M.; Kanapin, A.; Le Bot, N.; Moreno, S.; Sohrmann, M.; et al. Systematic Functional Analysis of the Caenorhabditis elegans Genome Using RNAi. Nature 2003, 421, 231–237. [Google Scholar] [CrossRef]
- Gruber, J.; Tang, S.Y.; Halliwell, B. Evidence for a Trade-Off between Survival and Fitness Caused by Resveratrol Treatment of Caenorhabditis elegans. Ann. N. Y. Acad. Sci. 2007, 1100, 530–542. [Google Scholar] [CrossRef]
- Hughes, S.E.; Evason, K.; Xiong, C.; Kornfeld, K. Genetic and Pharmacological Factors that Influence Reproductive Aging in Nematodes. PLoS Genet. 2007, 3, e25. [Google Scholar] [CrossRef]
- Schlotterer, A.; Kukudov, G.; Bozorgmehr, F.; Hutter, H.; Du, X.; Oikonomou, D.; Ibrahim, Y.; Pfisterer, F.; Rabbani, N.; Thornalley, P.; et al. Elegans as Model for the Study of High Glucose- Mediated Life Span Reduction. Diabetes 2009, 58, 2450–2456. [Google Scholar] [CrossRef]
- Pietsch, K.; Saul, N.; Menzel, R.; Stürzenbaum, S.R.; Steinberg, C.E.W. Quercetin Mediated Lifespan Extension in Caenorhabditis elegans Is Modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology 2009, 10, 565–578. [Google Scholar] [CrossRef]
- Kampkotter, A.; Nkwonkam, C.G.; Zurawski, R.F.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. Effects of the Flavonoids Kaempferol and Fisetin on Thermotolerance, Oxidative Stress and FoxO Transcription Factor DAF-16 in the Model Organism Caenorhabditis elegans. Arch. Toxicol. 2007, 81, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Gems, D. The Hyperfunction Theory: An Emerging Paradigm for the Biology of Aging. Ageing Res. Rev. 2022, 74, 101557. [Google Scholar] [CrossRef] [PubMed]
- Fulle, S.; Protasi, F.; Tano, G.D.; Pietrangelo, T.; Beltramin, A.; Boncompagni, S.; Vecchiet, L.; Fanò, G. The Contribution of Reactive Oxygen Species to Sarcopenia and Muscle Ageing. Exp. Gerontol. 2004, 39, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Toth, M.L.; Sigmond, T.; Borsos, E.; Barna, J.; Erdélyi, P.; Takács-Vellai, K.; Orosz, L.; Kovács, A.L.; Csikós, G.; Sass, M.; et al. Longevity Pathways Converge on Autophagy Genes to Regulate Life Span in Caenorhabditis elegans. Autophagy 2008, 4, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.J.; Willcox, D.C.; Donlon, T.A.; Willcox, B.J. FOXO3: A Major Gene for Human Longevity—A Mini-Review. Gerontology 2015, 61, 515–525. [Google Scholar] [CrossRef]
- Burnaevskiy, N.; Sands, B.; Yun, S.; Tedesco, P.M.; Johnson, T.E.; Kaeberlein, M.; Brent, R.; Mendenhall, A. Chaperone Biomarkers of Lifespan and Penetrance Track the Dosages of Many Other Proteins. Nat. Commun. 2019, 10, 5725. [Google Scholar] [CrossRef]
- Sanchez-Blanco, A.; Kim, S.K. Variable Pathogenicity Determines Individual Lifespan in Caenorhabditis elegans. PLoS Genet. 2011, 7, e1002047. [Google Scholar] [CrossRef]
- Caruana, M.; Cauchi, R.; Vassallo, N. Putative Role of Red Wine Polyphenols against Brain Pathology in Alzheimer’s and Parkinson’s Disease. Front. Nutr. 2016, 3, 31. [Google Scholar] [CrossRef]
- Akaishi, T.; Morimoto, T.; Shibao, M.; Watanabe, S.; Sakai-Kato, K.; Utsunomiya-Tate, N.; Abe, K. Structural Requirements for the Flavonoid Fisetin in Inhibiting Fibril Formation of Amyloid Beta Protein. Neurosci. Lett. 2008, 444, 280–285. [Google Scholar] [CrossRef]
- Dash, R.; Emran, T.B.; Uddin, M.M.N.; Islam, A.; Junaid, M. Molecular Docking of Fisetin with AD Associated AChE, ABAD and BACE1 Proteins. Bioinformation 2014, 10, 562–568. [Google Scholar] [CrossRef]
- Yang, J.; Huang, X.B.; Wan, Q.L.; Ding, A.J.; Yang, Z.L.; Qiu, M.H.; Sun, H.Y.; Qi, S.H.; Luo, H.R. Otophylloside B Protects against Abeta Toxicity in Caenorhabditis elegans Models of Alzheimer’s Disease. Nat. Prod. Bioprospect. 2017, 7, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Li, H.; Dong, J.; Yang, W.; Liu, T.; Wang, Y.; Wang, X.; Wang, M.; Zhi, D. Rose Essential Oil Delayed Alzheimer’s Disease-Like Symptoms by SKN-1 Pathway in C. elegans. J. Agric. Food Chem. 2017, 65, 8855–8865. [Google Scholar] [CrossRef] [PubMed]
- Okoye, I.; Yu, S.; Caruso, F.; Rossi, M. X-ray Structure Determination, Antioxidant Voltammetry Studies of Butein and 2′,4′-Dihydroxy-3,4-dimethoxychalcone. Computational Studies of 4 Structurally Related 2′,4′-diOH Chalcones to Examine Their Antimalarial Activity by Binding to Falcipain-2. Molecules 2021, 26, 6511. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.N.; Braga, R.C.; Grzelak, E.M.; Neves, B.J.; Muratov, E.; Ma, R.; Klein, L.L.; Cho, S.; Oliveira, G.R.; Franzblau, S.G.; et al. QSAR-Driven Design, Synthesis and Discovery of Potent Chalcone Derivatives with Antitubercular Activity. Eur. J. Med. Chem. 2017, 137, 126–138. [Google Scholar] [CrossRef]
- Chen, Z.; Li, P.; Hu, D.; Dong, L.; Pan, J.; Luo, L.; Zhang, W.; Xue, W.; Jin, L.; Song, B. Synthesis, Antiviral Activity, and 3D-QSAR Study of Novel Chalcone Derivatives Containing Malonate and Pyridine Moieties. Arab. J. Chem. 2019, 12, 2685–2696. [Google Scholar] [CrossRef]
- Xue, C.X.; Cui, S.Y.; Liu, M.C.; Hu, Z.D.; Fan, B.T. 3D QSAR Studies on Antimalarial Alkoxylated and Hydroxylated Chalcones by CoMFA and CoMSIA. Eur. J. Med. Chem. 2004, 39, 745–753. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Yoon, H.; Park, S.-K. Butein Increases Resistance to Oxidative Stress and Lifespan with Positive Effects on the Risk of Age-Related Diseases in Caenorhabditis elegans. Antioxidants 2024, 13, 155. https://doi.org/10.3390/antiox13020155
Kim S, Yoon H, Park S-K. Butein Increases Resistance to Oxidative Stress and Lifespan with Positive Effects on the Risk of Age-Related Diseases in Caenorhabditis elegans. Antioxidants. 2024; 13(2):155. https://doi.org/10.3390/antiox13020155
Chicago/Turabian StyleKim, Seona, Hyemin Yoon, and Sang-Kyu Park. 2024. "Butein Increases Resistance to Oxidative Stress and Lifespan with Positive Effects on the Risk of Age-Related Diseases in Caenorhabditis elegans" Antioxidants 13, no. 2: 155. https://doi.org/10.3390/antiox13020155
APA StyleKim, S., Yoon, H., & Park, S. -K. (2024). Butein Increases Resistance to Oxidative Stress and Lifespan with Positive Effects on the Risk of Age-Related Diseases in Caenorhabditis elegans. Antioxidants, 13(2), 155. https://doi.org/10.3390/antiox13020155