



The Cerebral Protective Effect of Novel Erinacines from Hericium erinaceus Mycelium on In Vivo Mild Traumatic Brain Injury Animal Model and Primary Mixed Glial Cells via Nrf2-Dependent Pathways

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
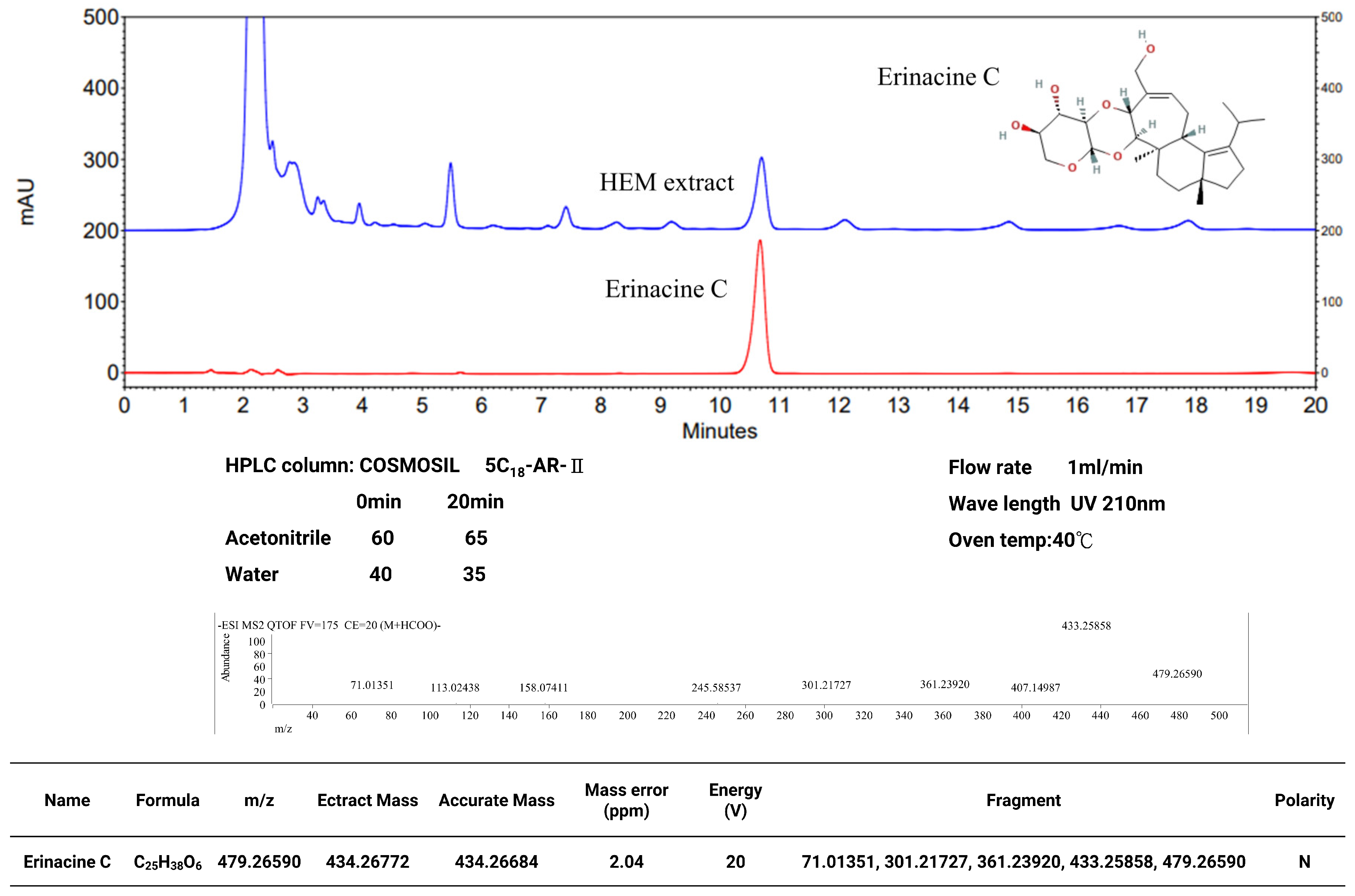
2.1. Hericium Erinaceus Extracts and Analysis of Erinacine C
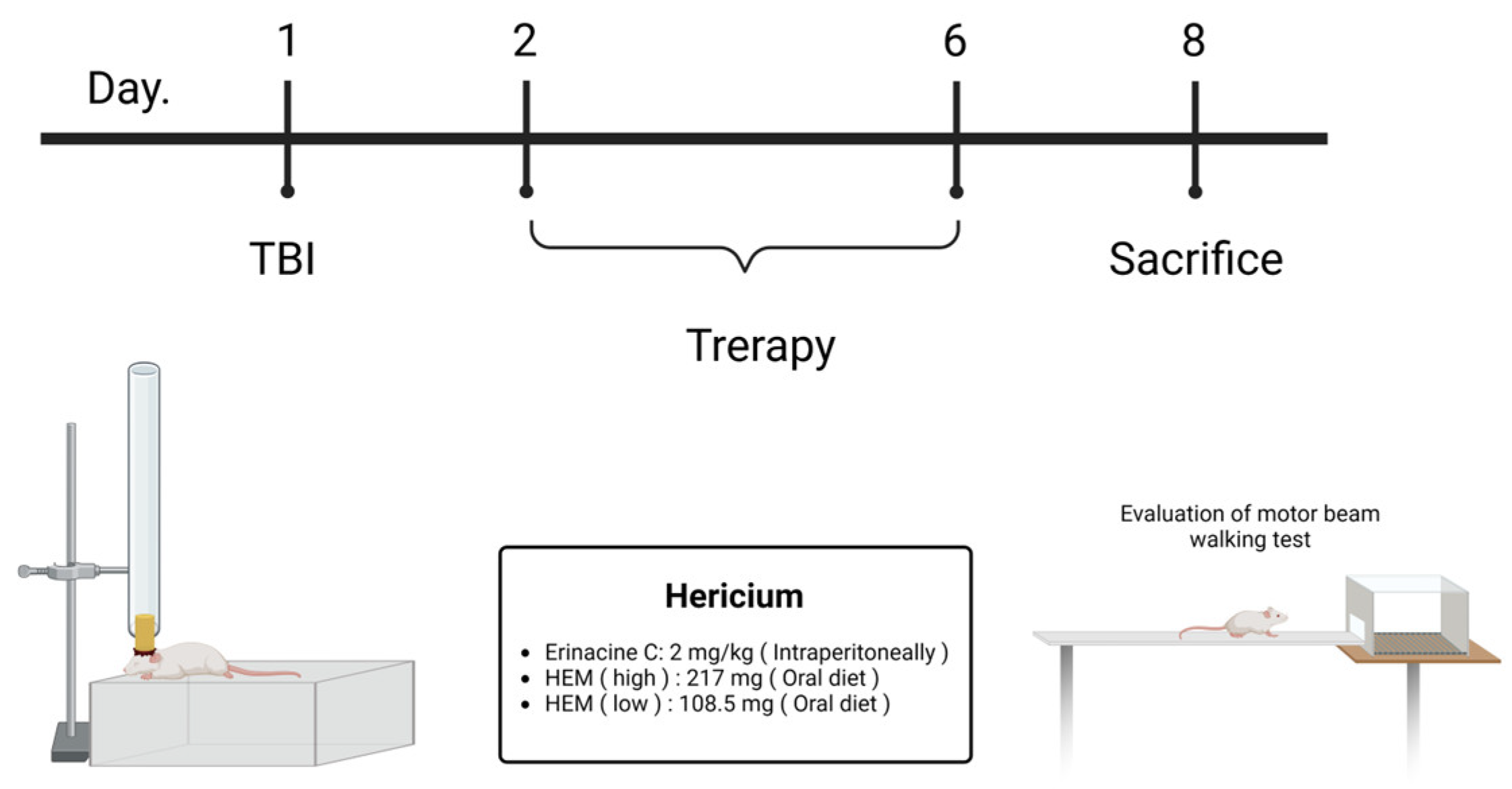
2.2. Induction mTBI Brain Injury and Drug Administration
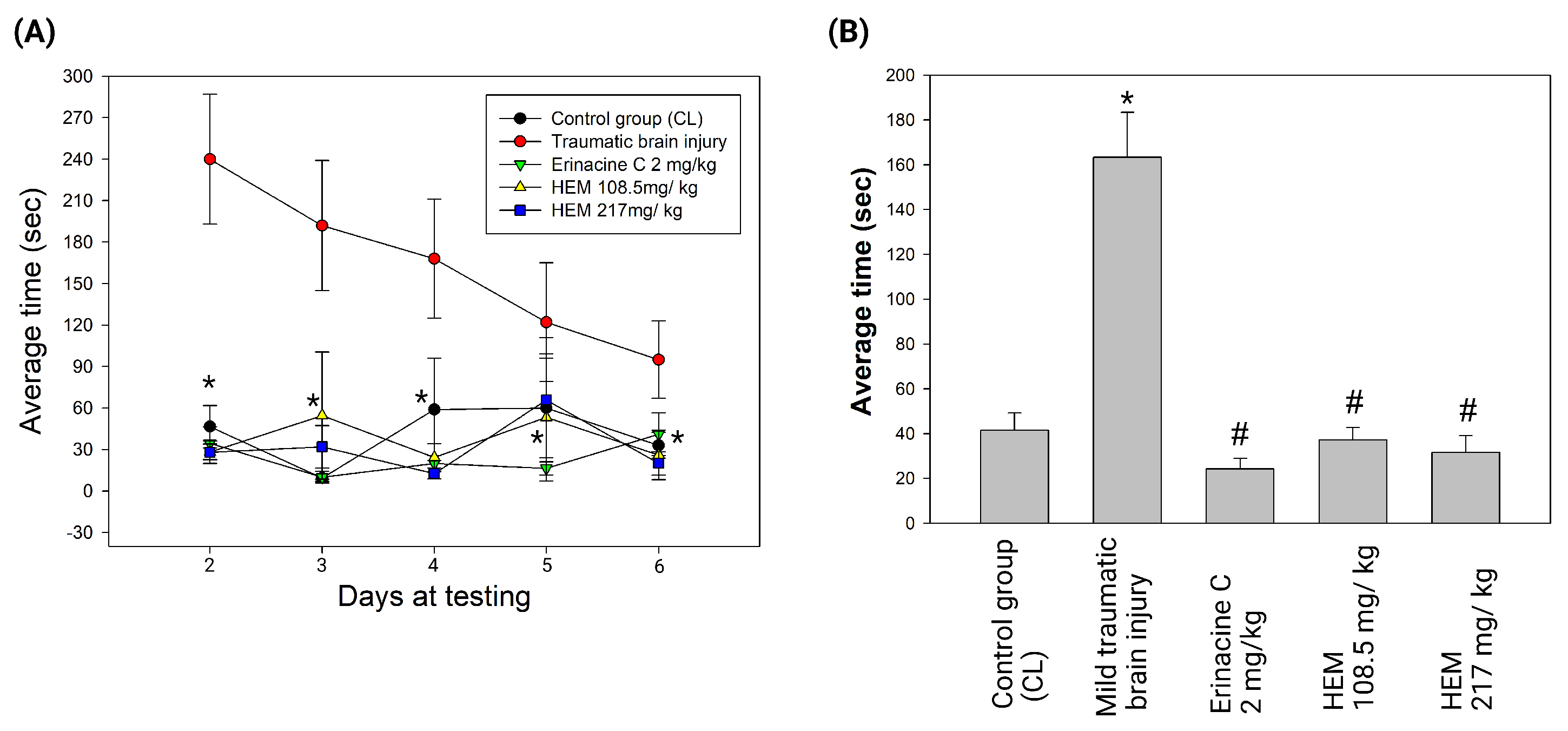
2.3. Verification of mTBI with the Beam Walking Test
2.4. Reagents and Antibodies
2.5. Preparation of Primary Mixed Glia Cultures and Cell Treatment
2.6. Immunohistochemistry
2.7. Histopathological Evaluation
2.8. Preparation of Total Cell Extracts and Immunoblotting Analysis
2.9. Chromatin Immunoprecipitation (ChIP) Analysis
2.10. Statistical Analysis
3. Results
3.1. Hericium Erinaceus Mycelium Treatment Ameliorates Brain Damage in Rat Model with Mild Traumatic Brain Injury
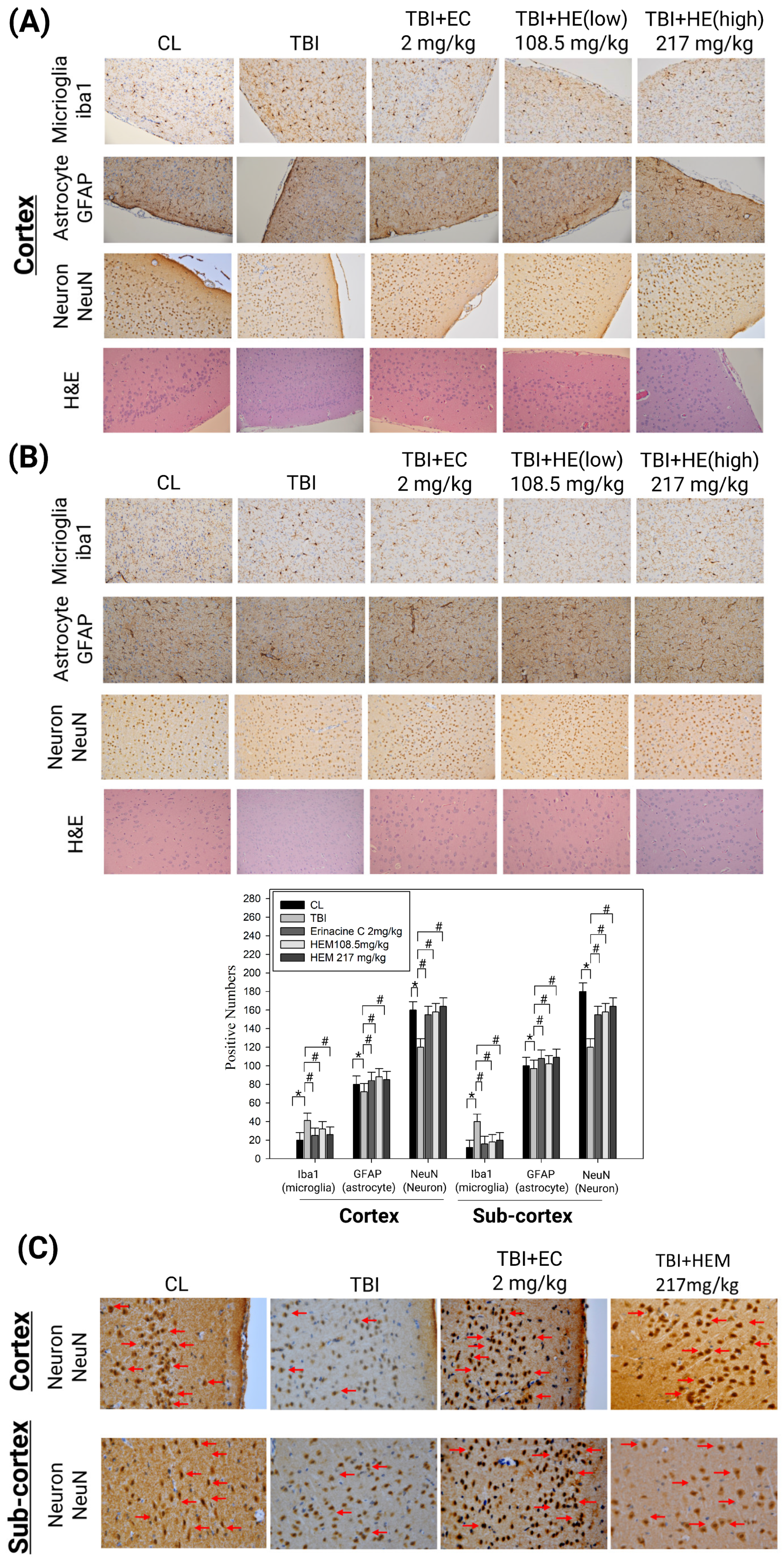
3.2. Hericium Erinaceus Mycelium Prevents Neuron Cell Death and Alleviates Neuroinflammation in the Rat Model with Mild Traumatic Brain Injury
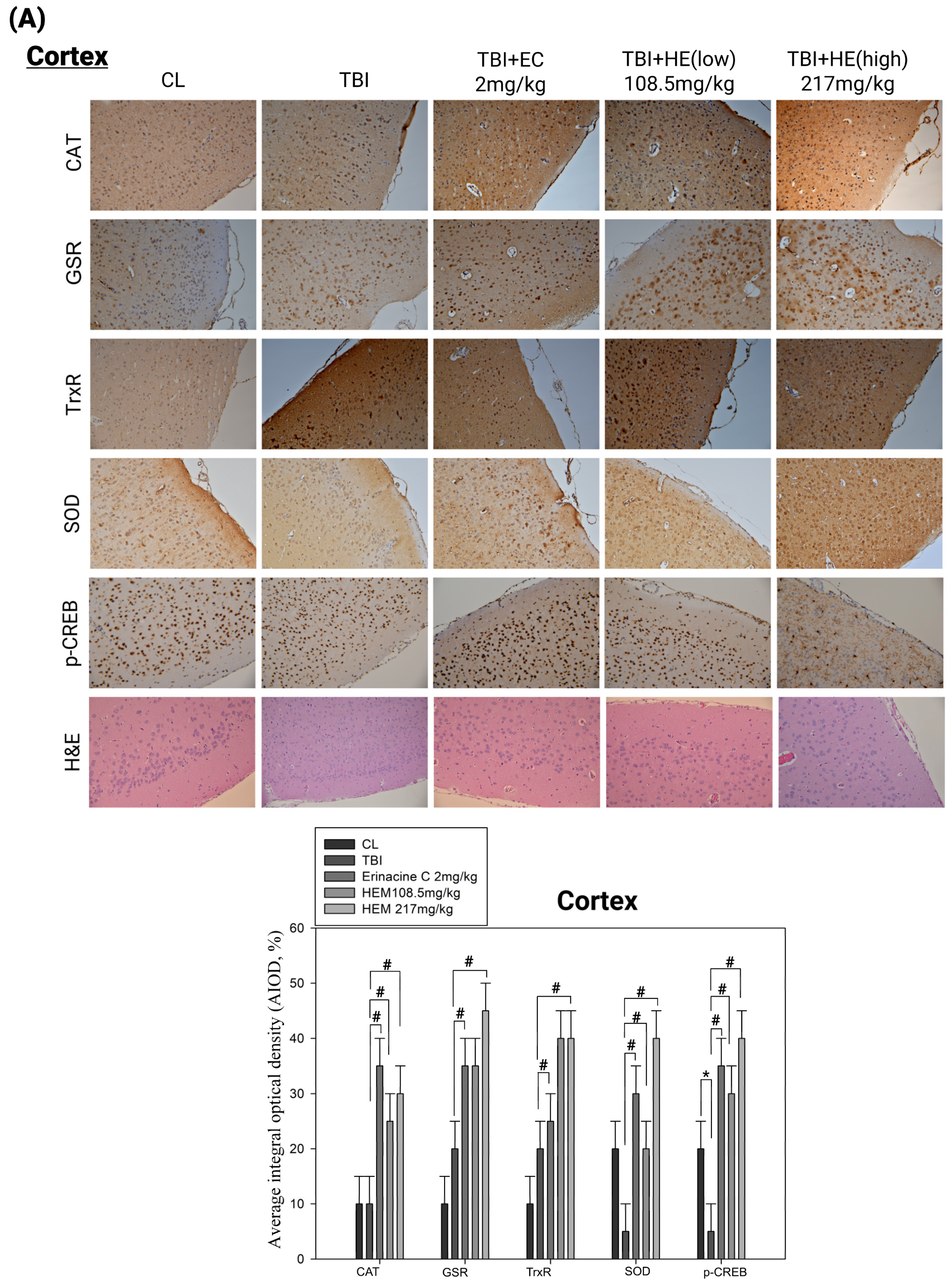
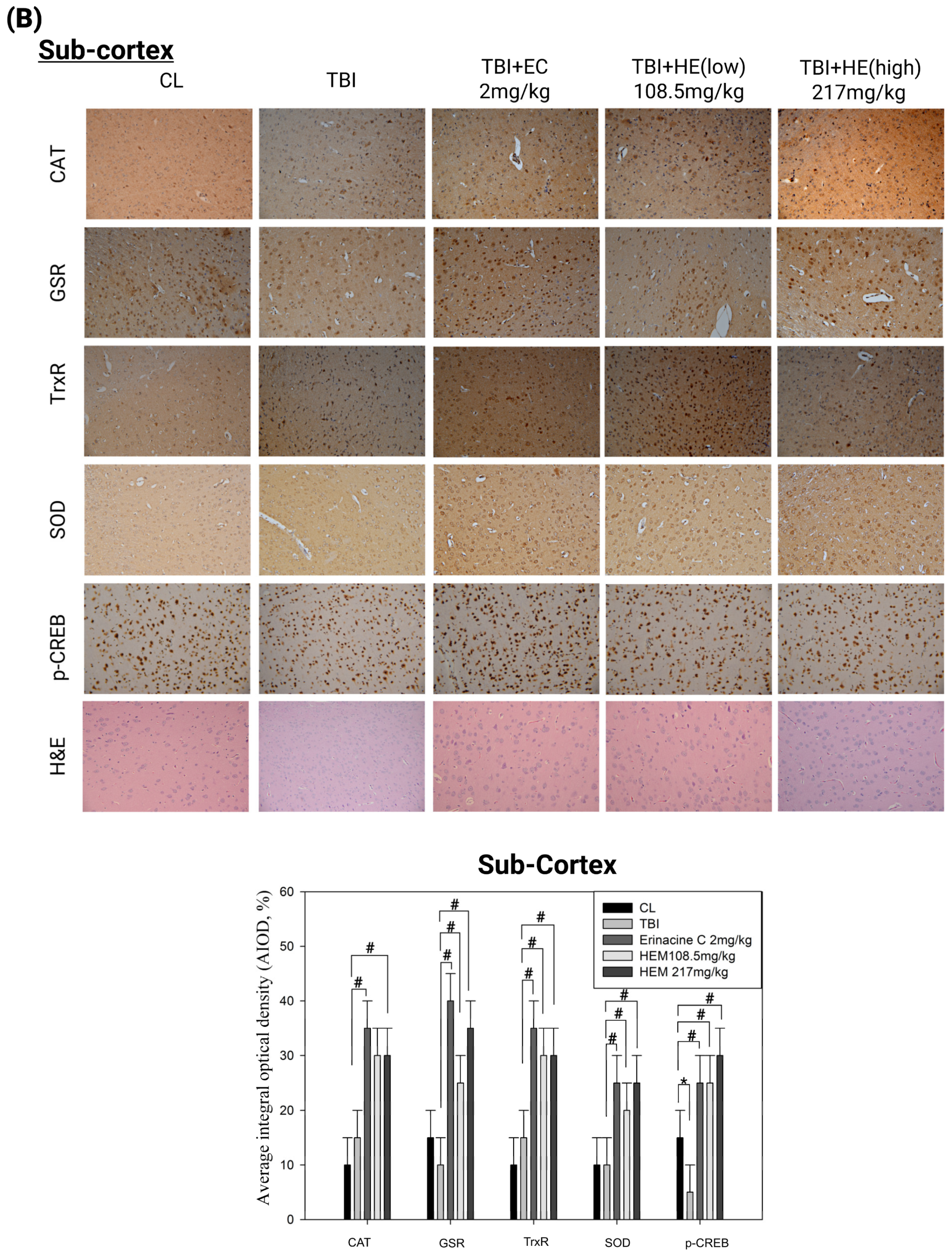
3.3. Post-Treatment with H. erinaceus Mycelium Reversed the Several Antioxidant Enzymes and Phospho-CAMP-Response Element-Binding Levels in a Rat Model of Mild Traumatic Brain Injury
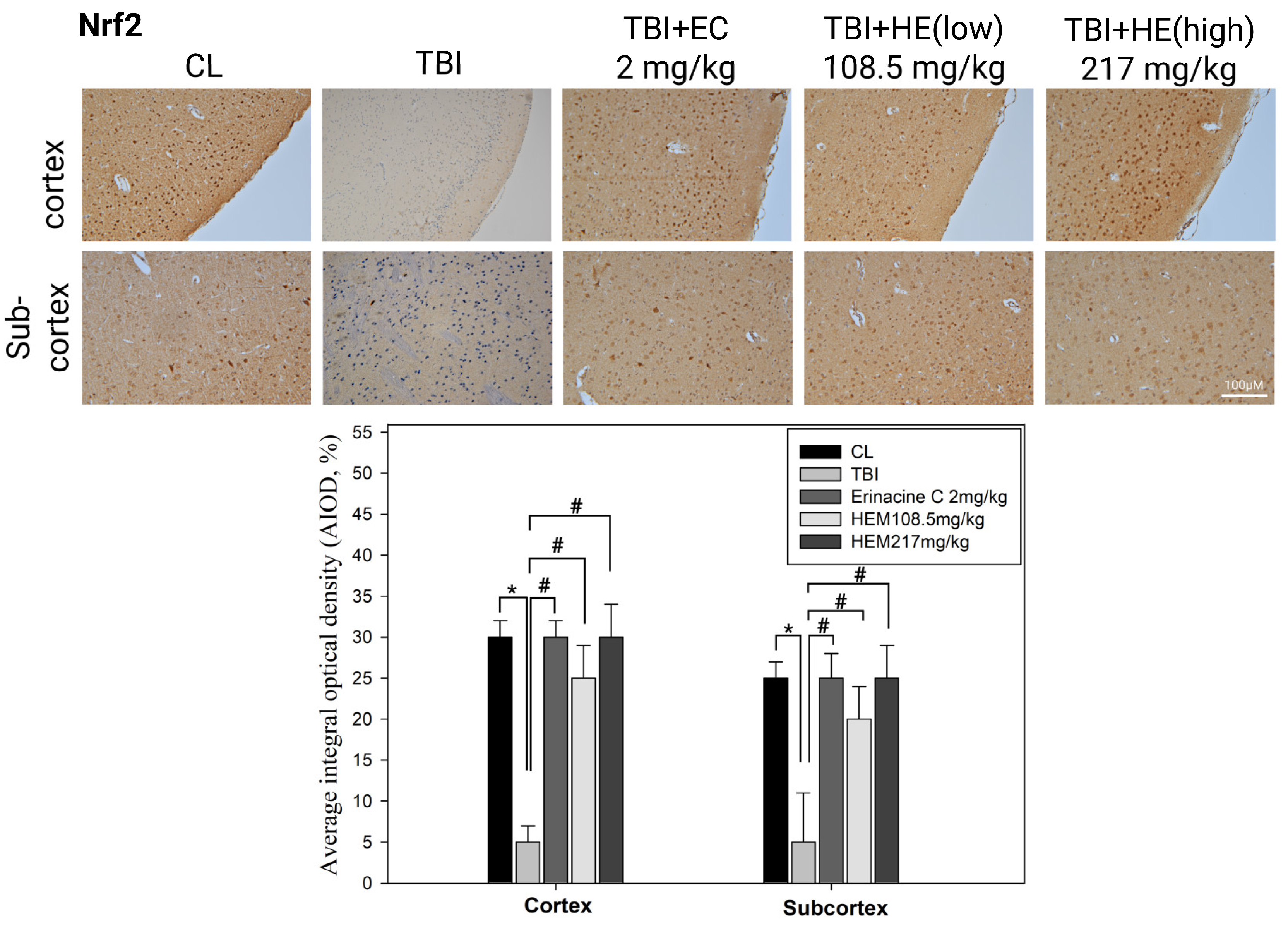
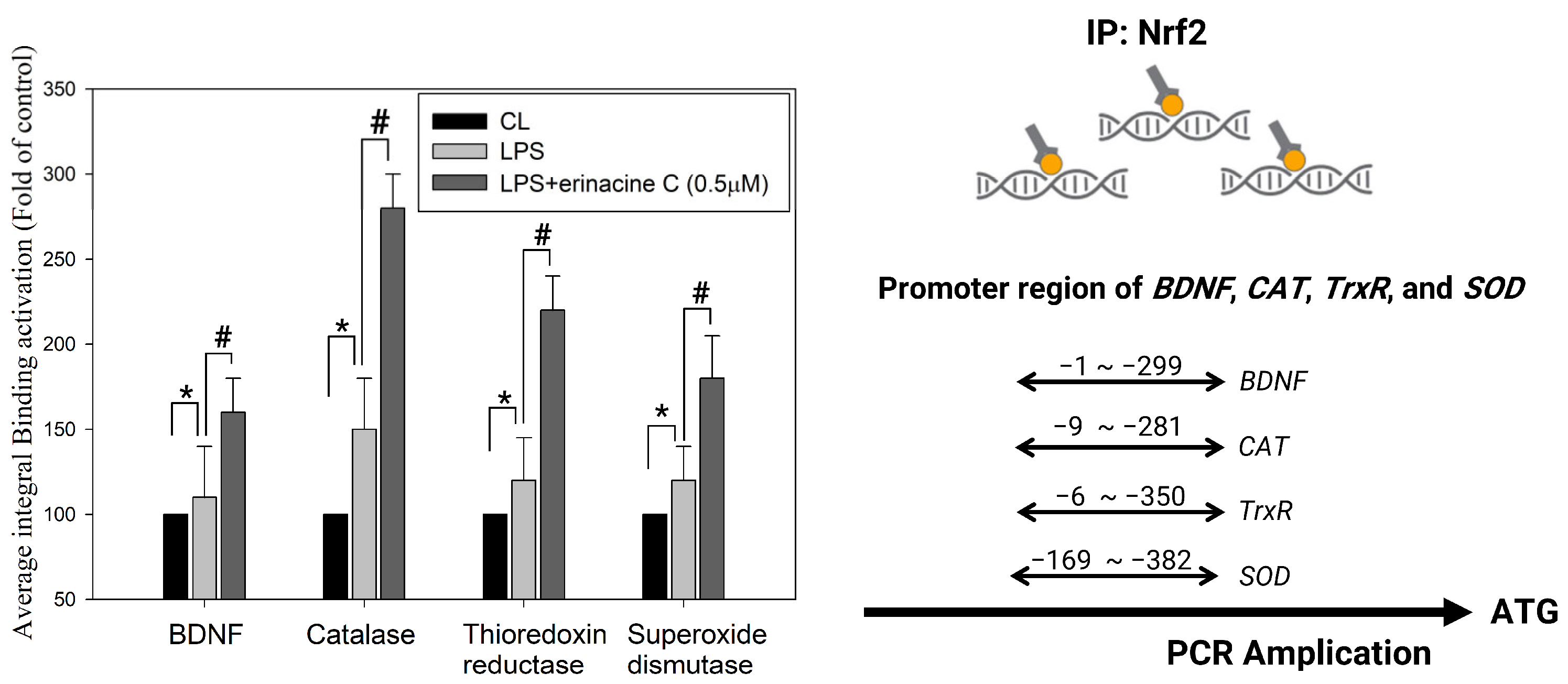
3.4. Immunohistochemistry Stain of NF-E2-Related Factor 2 Proteins on Brain Histopathology by H. erinaceus Mycelium and Erinacine C in Animal Model Rats with Mild Traumatic Brain Injury
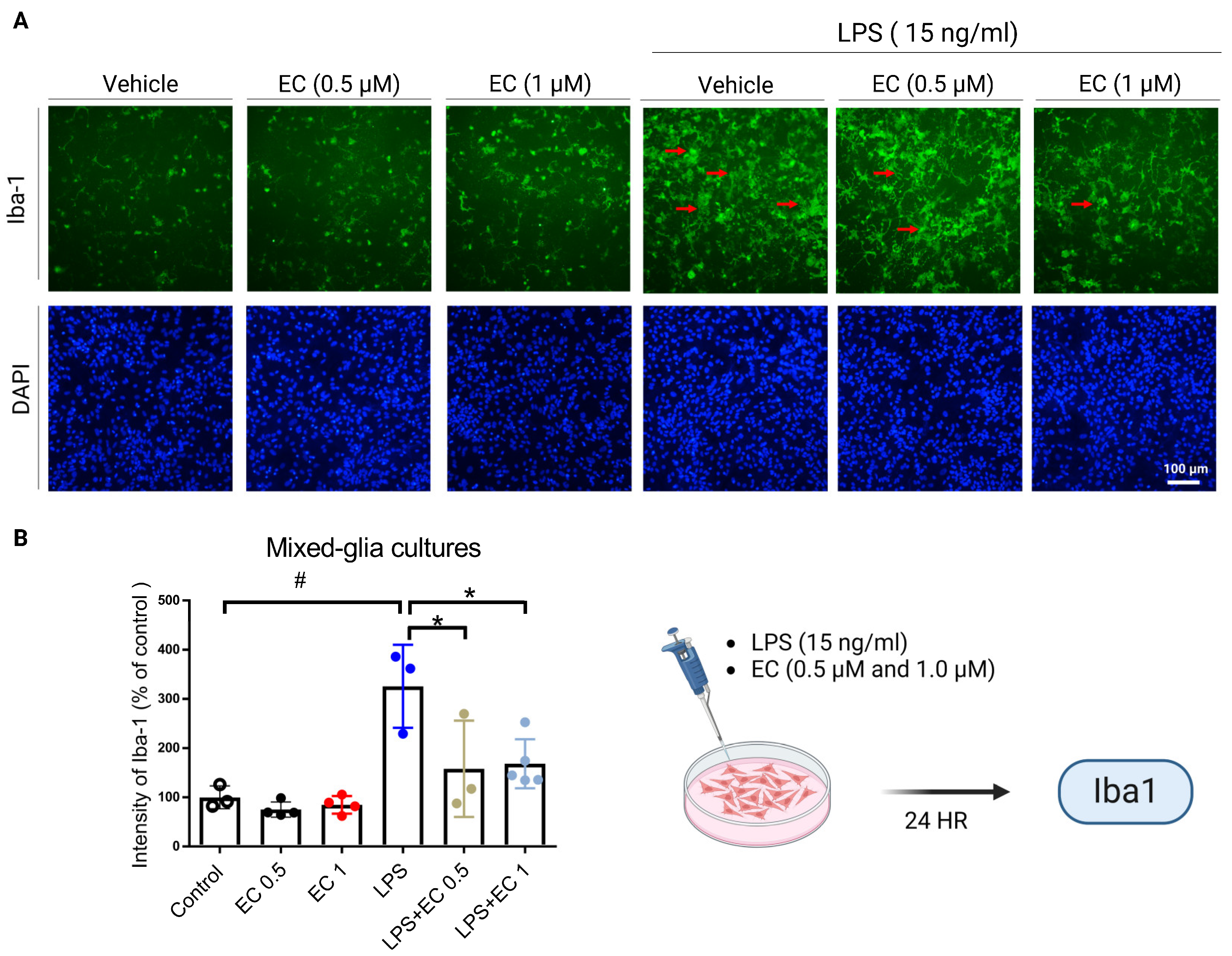
3.5. Erinacine C Reduction of Lipopolysaccharide (LPS)-Induced Microglia Cells Activation in Primary Mixed-Glia Cultures
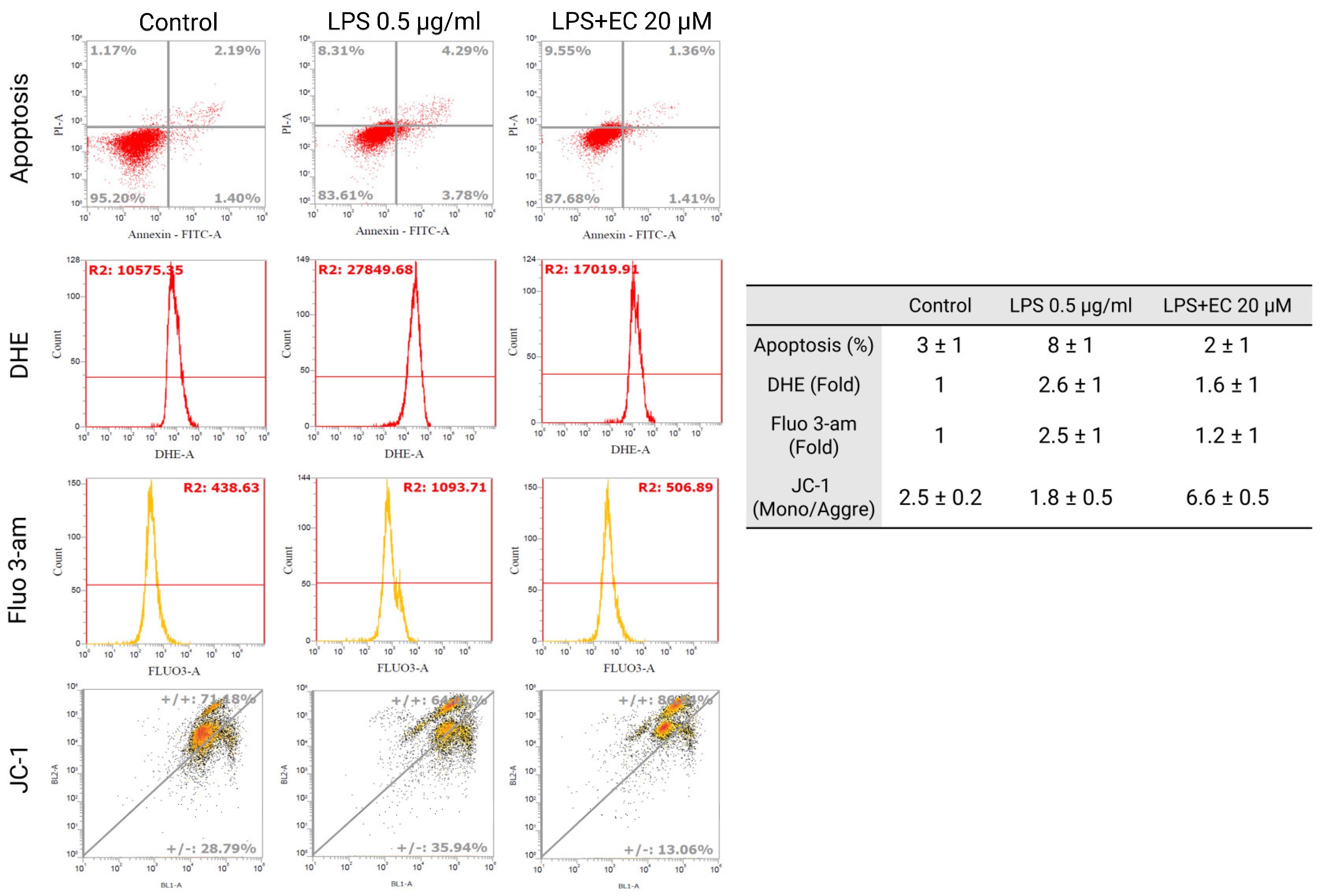
3.6. Erinacine C Decline of Lipopolysaccharide-Induced Mitochondrial Potential, Reactive Oxygen Species Production and Calcium Dysregulation in BV2 Microglia Cells
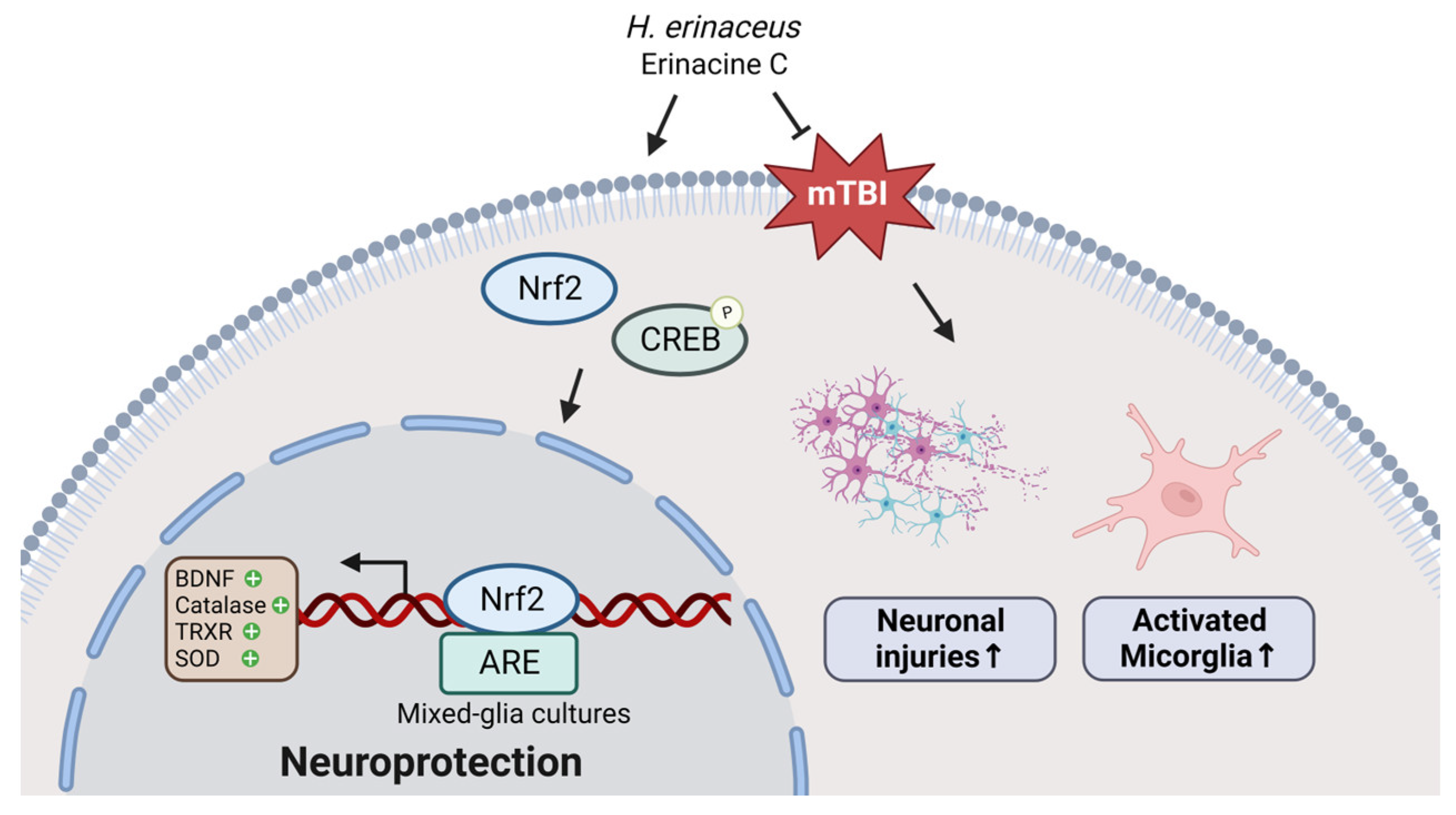
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeKosky, S.T.; Asken, B.M. Injury cascades in TBI-related neurodegeneration. Brain Inj. 2017, 31, 1177–1182. [Google Scholar] [CrossRef]
- Smith, D.H.; Johnson, V.E.; Stewart, W. Chronic neuropathologies of single and repetitive TBI: Substrates of dementia? Nat. Rev. Neurol. 2013, 9, 211–221. [Google Scholar] [CrossRef]
- Poloyac, S.M.; Bertz, R.J.; McDermott, L.A.; Marathe, P. Pharmacological Optimization for Successful Traumatic Brain Injury Drug Development. J. Neurotrauma 2020, 37, 2435–2444. [Google Scholar] [CrossRef]
- Ahmad, A.; Ali, T.; Rehman, S.U.; Kim, M.O. Phytomedicine-Based Potent Antioxidant, Fisetin Protects CNS-Insult LPS-Induced Oxidative Stress-Mediated Neurodegeneration and Memory Impairment. J. Clin. Med. 2019, 8, 850. [Google Scholar] [CrossRef]
- VanItallie, T.B. Traumatic brain injury (TBI) in collision sports: Possible mechanisms of transformation into chronic traumatic encephalopathy (CTE). Metabolism 2019, 100S, 153943. [Google Scholar] [CrossRef]
- Lipton, M.L.; Gellella, E.; Lo, C.; Gold, T.; Ardekani, B.A.; Shifteh, K.; Bello, J.A.; Branch, C.A. Multifocal white matter ultrastructural abnormalities in mild traumatic brain injury with cognitive disability: A voxel-wise analysis of diffusion tensor imaging. J. Neurotrauma 2008, 25, 1335–1342. [Google Scholar] [CrossRef]
- Niogi, S.N.; Mukherjee, P.; Ghajar, J.; Johnson, C.; Kolster, R.A.; Sarkar, R.; Lee, H.; Meeker, M.; Zimmerman, R.D.; Manley, G.T.; et al. Extent of microstructural white matter injury in postconcussive syndrome correlates with impaired cognitive reaction time: A 3T diffusion tensor imaging study of mild traumatic brain injury. AJNR Am. J. Neuroradiol. 2008, 29, 967–973. [Google Scholar] [CrossRef]
- Rubovitch, V.; Ten-Bosch, M.; Zohar, O.; Harrison, C.R.; Tempel-Brami, C.; Stein, E.; Hoffer, B.J.; Balaban, C.D.; Schreiber, S.; Chiu, W.T.; et al. A mouse model of blast-induced mild traumatic brain injury. Exp. Neurol. 2011, 232, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Shultz, S.R.; MacFabe, D.F.; Foley, K.A.; Taylor, R.; Cain, D.P. A single mild fluid percussion injury induces short-term behavioral and neuropathological changes in the Long-Evans rat: Support for an animal model of concussion. Behav. Brain Res. 2011, 224, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Dewitt, D.S.; Perez-Polo, R.; Hulsebosch, C.E.; Dash, P.K.; Robertson, C.S. Challenges in the development of rodent models of mild traumatic brain injury. J. Neurotrauma 2013, 30, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Mahmood, A.; Chopp, M. Animal models of traumatic brain injury. Nat. Rev. Neurosci. 2013, 14, 128–142. [Google Scholar] [CrossRef]
- Mychasiuk, R.; Farran, A.; Angoa-Perez, M.; Briggs, D.; Kuhn, D.; Esser, M.J. A novel model of mild traumatic brain injury for juvenile rats. J. Vis. Exp. 2014, 51820. [Google Scholar] [CrossRef]
- Naeser, M.A.; Zafonte, R.; Krengel, M.H.; Martin, P.I.; Frazier, J.; Hamblin, M.R.; Knight, J.A.; Meehan, W.P., 3rd; Baker, E.H. Significant improvements in cognitive performance post-transcranial, red/near-infrared light-emitting diode treatments in chronic, mild traumatic brain injury: Open-protocol study. J. Neurotrauma 2014, 31, 1008–1017. [Google Scholar] [CrossRef]
- Zhang, Z.; Larner, S.F.; Kobeissy, F.; Hayes, R.L.; Wang, K.K. Systems biology and theranostic approach to drug discovery and development to treat traumatic brain injury. Methods Mol. Biol. 2010, 662, 317–329. [Google Scholar] [CrossRef]
- Xiong, Y.; Gu, Q.; Peterson, P.L.; Muizelaar, J.P.; Lee, C.P. Mitochondrial dysfunction and calcium perturbation induced by traumatic brain injury. J. Neurotrauma 1997, 14, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Rehman, S.U.; Shah, F.A.; Kim, M.O. Acute dose of melatonin via Nrf2 dependently prevents acute ethanol-induced neurotoxicity in the developing rodent brain. J. Neuroinflammation 2018, 15, 119. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, J.; Hu, X.; Zhang, L.; Mao, L.; Jiang, X.; Liou, A.K.; Leak, R.K.; Gao, Y.; Chen, J. Microglia/macrophage polarization dynamics in white matter after traumatic brain injury. J. Cereb. Blood Flow. Metab. 2013, 33, 1864–1874. [Google Scholar] [CrossRef] [PubMed]
- Islekel, S.; Islekel, H.; Guner, G.; Ozdamar, N. Alterations in superoxide dismutase, glutathione peroxidase and catalase activities in experimental cerebral ischemia-reperfusion. Res. Exp. Med. 1999, 199, 167–176. [Google Scholar] [CrossRef]
- DeKosky, S.T.; Taffe, K.M.; Abrahamson, E.E.; Dixon, C.E.; Kochanek, P.M.; Ikonomovic, M.D. Time course analysis of hippocampal nerve growth factor and antioxidant enzyme activity following lateral controlled cortical impact brain injury in the rat. J. Neurotrauma 2004, 21, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Dringen, R.; Pawlowski, P.G.; Hirrlinger, J. Peroxide detoxification by brain cells. J. Neurosci. Res. 2005, 79, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Jassam, Y.N.; Izzy, S.; Whalen, M.; McGavern, D.B.; El Khoury, J. Neuroimmunology of Traumatic Brain Injury: Time for a Paradigm Shift. Neuron 2017, 95, 1246–1265. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Hu, Q.; Wu, X.; Xu, H.; Su, A.; Xie, M.; Yang, W. Bioactivities and molecular mechanisms of polysaccharides from Hericium erinaceus. J. Future Foods 2022, 2, 103–111. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Ulziijargal, E.; Mau, J.L. Nutrient compositions of culinary-medicinal mushroom fruiting bodies and mycelia. Int. J. Med. Mushrooms 2011, 13, 343–349. [Google Scholar] [CrossRef]
- Kawagishi, H.; Shimada, A.; Shirai, R.; Okamoto, K.; Ojima, F.; Sakamoto, H.; Ishiguro, Y.; Furukawa, S. Erinacine-a, Erinacine-B and Erinacine-C, Strong Stimulators of Nerve Growth-Factor (Ngf)-Synthesis, from the Mycelia of Hericium Erinaceum. Tetrahedron Lett. 1994, 35, 1569–1572. [Google Scholar] [CrossRef]
- Kuo, H.C.; Lu, C.C.; Shen, C.H.; Tung, S.Y.; Hsieh, M.C.; Lee, K.C.; Lee, L.Y.; Chen, C.C.; Teng, C.C.; Huang, W.S.; et al. Hericium erinaceus mycelium and its isolated erinacine A protection from MPTP-induced neurotoxicity through the ER stress, triggering an apoptosis cascade. J. Transl. Med. 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Huang, C.S.; Chen, Y.H.; Chen, C.C.; Chen, C.C.; Chuang, C.H. Anti-Inflammatory Effect of Erinacine C on NO Production Through Down-Regulation of NF-kappaB and Activation of Nrf2-Mediated HO-1 in BV2 Microglial Cells Treated with LPS. Molecules 2019, 24, 3317. [Google Scholar] [CrossRef] [PubMed]
- Kane, M.J.; Angoa-Perez, M.; Briggs, D.I.; Viano, D.C.; Kreipke, C.W.; Kuhn, D.M. A mouse model of human repetitive mild traumatic brain injury. J. Neurosci. Methods 2012, 203, 41–49. [Google Scholar] [CrossRef]
- Mychasiuk, R.; Hehar, H.; Farran, A.; Esser, M.J. Mean girls: Sex differences in the effects of mild traumatic brain injury on the social dynamics of juvenile rat play behaviour. Behav. Brain Res. 2014, 259, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wei, X.T.; Guan, J.H.; Yuan, J.W.; Peng, Y.T.; Song, L.; Liu, Y.H. Inhibition of c-Jun N-terminal kinase prevents blood-brain barrier disruption and normalizes the expression of tight junction proteins clautin-5 and ZO-1 in a rat model of subarachnoid hemorrhage. Acta Neurochir. 2012, 154, 1469–1476; discussion 1476. [Google Scholar] [CrossRef]
- Kuo, H.C.; Lee, K.F.; Chen, S.L.; Chiu, S.C.; Lee, L.Y.; Chen, W.P.; Chen, C.C.; Chu, C.H. Neuron-Microglia Contacts Govern the PGE(2) Tolerance through TLR4-Mediated de Novo Protein Synthesis. Biomedicines 2022, 10, 419. [Google Scholar] [CrossRef]
- Chi, V.; Chandy, K.G. Immunohistochemistry: Paraffin sections using the Vectastain ABC kit from vector labs. J. Vis. Exp. 2007, 308. [Google Scholar] [CrossRef]
- Li, J.J.; Chen, W.L.; Wang, J.Y.; Hu, Q.W.; Sun, Z.P.; Zhang, S.; Liu, S.; Han, X.H. Wenshen Zhuanggu formula effectively suppresses breast cancer bone metastases in a mouse Xenograft model. Acta Pharmacol. Sin. 2017, 38, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Pillai-Kastoori, L.; Schutz-Geschwender, A.R.; Harford, J.A. A systematic approach to quantitative Western blot analysis. Anal. Biochem. 2020, 593, 113608. [Google Scholar] [CrossRef]
- Lee, K.C.; Kuo, H.C.; Shen, C.H.; Lu, C.C.; Huang, W.S.; Hsieh, M.C.; Huang, C.Y.; Kuo, Y.H.; Hsieh, Y.Y.; Teng, C.C.; et al. A proteomics approach to identifying novel protein targets involved in erinacine A-mediated inhibition of colorectal cancer cells’ aggressiveness. J. Cell Mol. Med. 2017, 21, 588–599. [Google Scholar] [CrossRef]
- Kitagawa, K. CREB and cAMP response element-mediated gene expression in the ischemic brain. FEBS J. 2007, 274, 3210–3217. [Google Scholar] [CrossRef]
- Li, C.; Chen, T.; Zhou, H.; Feng, Y.; Hoi, M.P.M.; Ma, D.; Zhao, C.; Zheng, Y.; Lee, S.M.Y. BHDPC Is a Novel Neuroprotectant That Provides Anti-neuroinflammatory and Neuroprotective Effects by Inactivating NF-kappaB and Activating PKA/CREB. Front. Pharmacol. 2018, 9, 614. [Google Scholar] [CrossRef]
- Corrigan, F.; Mander, K.A.; Leonard, A.V.; Vink, R. Neurogenic inflammation after traumatic brain injury and its potentiation of classical inflammation. J. Neuroinflammation 2016, 13, 264. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wang, H.D.; Hu, Z.G.; Wang, Q.F.; Yin, H.X. Activation of Nrf2-ARE pathway in brain after traumatic brain injury. Neurosci. Lett. 2008, 431, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fields, J.; Zhao, C.; Langer, J.; Thimmulappa, R.K.; Kensler, T.W.; Yamamoto, M.; Biswal, S.; Dore, S. Role of Nrf2 in protection against intracerebral hemorrhage injury in mice. Free Radic. Biol. Med. 2007, 43, 408–414. [Google Scholar] [CrossRef]
- Miller, D.M.; Wang, J.A.; Buchanan, A.K.; Hall, E.D. Temporal and spatial dynamics of nrf2-antioxidant response elements mediated gene targets in cortex and hippocampus after controlled cortical impact traumatic brain injury in mice. J. Neurotrauma 2014, 31, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.M.; Singh, I.N.; Wang, J.A.; Hall, E.D. Nrf2-ARE activator carnosic acid decreases mitochondrial dysfunction, oxidative damage and neuronal cytoskeletal degradation following traumatic brain injury in mice. Exp. Neurol. 2015, 264, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.L.; Kobori, N.; Zhao, J.; Rozas, N.S.; Hylin, M.J.; Moore, A.N.; Dash, P.K. Traumatic brain injury decreases AMP-activated protein kinase activity and pharmacological enhancement of its activity improves cognitive outcome. J. Neurochem. 2016, 139, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.G.; Yong, Y.Y.; Pan, Y.R.; Zhang, L.; Wu, J.M.; Zhang, Y.; Tang, Y.; Wei, J.; Yu, L.; Law, B.Y.; et al. Targeting Nrf2-Mediated Oxidative Stress Response in Traumatic Brain Injury: Therapeutic Perspectives of Phytochemicals. Oxid. Med. Cell Longev. 2022, 2022, 1015791. [Google Scholar] [CrossRef] [PubMed]
- Lerouet, D.; Marchand-Leroux, C.; Besson, V.C. Neuropharmacology in traumatic brain injury: From preclinical to clinical neuroprotection? Fundam. Clin. Pharmacol. 2021, 35, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Wang, W.; Li, Q.; Han, X.; Xing, J.; Qi, C.; Lan, X.; Wan, J.; Potts, A.; Guan, F.; et al. Cerebroprotection of flavanol (−)-epicatechin after traumatic brain injury via Nrf2-dependent and -independent pathways. Free Radic. Biol. Med. 2016, 92, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, E.; Brouillard, F.; Molet, J.; Claverie, D.; Cabungcal, J.H.; Cresto, N.; Doligez, N.; Rivat, C.; Do, K.Q.; Bernard, C.; et al. Nrf2-dependent persistent oxidative stress results in stress-induced vulnerability to depression. Mol. Psychiatry 2017, 22, 1701–1713. [Google Scholar] [CrossRef]
- Rhee, S.G. Cell signaling. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- Bruna, B.; Lobos, P.; Herrera-Molina, R.; Hidalgo, C.; Paula-Lima, A.; Adasme, T. The signaling pathways underlying BDNF-induced Nrf2 hippocampal nuclear translocation involve ROS, RyR-Mediated Ca(2+) signals, ERK and PI3K. Biochem. Biophys. Res. Commun. 2018, 505, 201–207. [Google Scholar] [CrossRef]
- Rehman, S.U.; Ikram, M.; Ullah, N.; Alam, S.I.; Park, H.Y.; Badshah, H.; Choe, K.; Kim, M.O. Neurological Enhancement Effects of Melatonin against Brain Injury-Induced Oxidative Stress, Neuroinflammation, and Neurodegeneration via AMPK/CREB Signaling. Cells 2019, 8, 760. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Q.L.; Zhang, X.Y.; Lv, C.; Li, J.; Yuan, Y.; Yin, F.X. Pharmacokinetics and blood-brain barrier penetration of (+)-catechin and (−)-epicatechin in rats by microdialysis sampling coupled to high-performance liquid chromatography with chemiluminescence detection. J. Agric. Food Chem. 2012, 60, 9377–9383. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ferruzzi, M.G.; Ho, L.; Blount, J.; Janle, E.M.; Gong, B.; Pan, Y.; Gowda, G.A.; Raftery, D.; Arrieta-Cruz, I.; et al. Brain-targeted proanthocyanidin metabolites for Alzheimer’s disease treatment. J. Neurosci. 2012, 32, 5144–5150. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gao, X.; Zhao, S.; Hu, W.; Chen, J. The Small-Molecule TrkB Agonist 7, 8-Dihydroxyflavone Decreases Hippocampal Newborn Neuron Death After Traumatic Brain Injury. J. Neuropathol. Exp. Neurol. 2015, 74, 557–567. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BDNF −1 to −299 bp NCBI Reference Sequence: NM_001285421.1 Sequence ID: NC_000068.8 |
| 5′-TTGTTTTGTGCCGTTTACCA-3 |
| 5′-GGTAAGAGAGCCAGCCACTG-3 |
| Catalsase −9 to −281 bp NCBI Reference Sequence: NM_009804 Sequence ID: NC_000068.7 |
| 5′-ATTGGACCCTGAGCTGTGAC-3′ |
| 5′-GGAGAAGGCAATCTTGTTGG-3′ |
| Thioredoxin reductas −6 to −350 bp NCBI Reference Sequence: NM_015762 Sequence ID: NC_000076.7 |
| 5′-CGATTCTGGTTCCCAACATT-3′ |
| 5′-TAAAGAGCTGCGGGTTCCTA-3′ |
| Superoxidase dismutase −169 to −382 bp NCBI Reference Sequence: NM_011434 Sequence ID: NC_000082.7 |
| 5′-ATCTTGGCGCATCTCAACTT-3′ |
| 5′-GTGCGGACTGAGAAAGTTCC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-F.; Hsieh, Y.-Y.; Tung, S.-Y.; Teng, C.-C.; Cheng, K.-C.; Hsieh, M.-C.; Huang, C.-Y.; Lee, K.-C.; Lee, L.-Y.; Chen, W.-P.; et al. The Cerebral Protective Effect of Novel Erinacines from Hericium erinaceus Mycelium on In Vivo Mild Traumatic Brain Injury Animal Model and Primary Mixed Glial Cells via Nrf2-Dependent Pathways. Antioxidants 2024, 13, 371. https://doi.org/10.3390/antiox13030371
Lee K-F, Hsieh Y-Y, Tung S-Y, Teng C-C, Cheng K-C, Hsieh M-C, Huang C-Y, Lee K-C, Lee L-Y, Chen W-P, et al. The Cerebral Protective Effect of Novel Erinacines from Hericium erinaceus Mycelium on In Vivo Mild Traumatic Brain Injury Animal Model and Primary Mixed Glial Cells via Nrf2-Dependent Pathways. Antioxidants. 2024; 13(3):371. https://doi.org/10.3390/antiox13030371
Chicago/Turabian StyleLee, Kam-Fai, Yung-Yu Hsieh, Shui-Yi Tung, Chih-Chuan Teng, Kung-Chuan Cheng, Meng-Chiao Hsieh, Cheng-Yi Huang, Ko-Chao Lee, Li-Ya Lee, Wan-Ping Chen, and et al. 2024. "The Cerebral Protective Effect of Novel Erinacines from Hericium erinaceus Mycelium on In Vivo Mild Traumatic Brain Injury Animal Model and Primary Mixed Glial Cells via Nrf2-Dependent Pathways" Antioxidants 13, no. 3: 371. https://doi.org/10.3390/antiox13030371
APA StyleLee, K. -F., Hsieh, Y. -Y., Tung, S. -Y., Teng, C. -C., Cheng, K. -C., Hsieh, M. -C., Huang, C. -Y., Lee, K. -C., Lee, L. -Y., Chen, W. -P., Chen, C. -C., & Kuo, H. -C. (2024). The Cerebral Protective Effect of Novel Erinacines from Hericium erinaceus Mycelium on In Vivo Mild Traumatic Brain Injury Animal Model and Primary Mixed Glial Cells via Nrf2-Dependent Pathways. Antioxidants, 13(3), 371. https://doi.org/10.3390/antiox13030371