Oxidation of Methionine 77 in Calmodulin Alters Mouse Growth and Behavior


Abstract
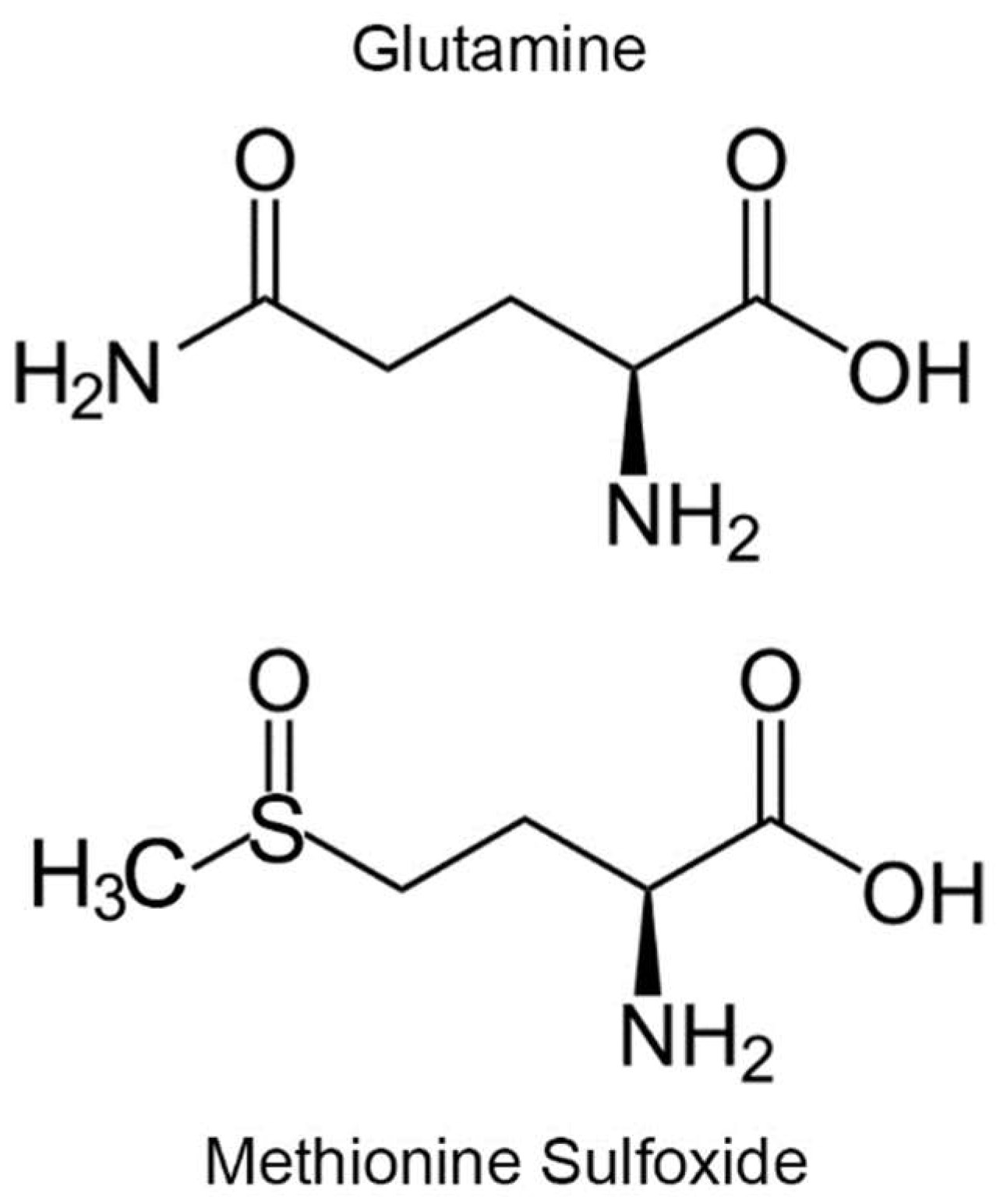
:1. Introduction
2. Materials and Methods
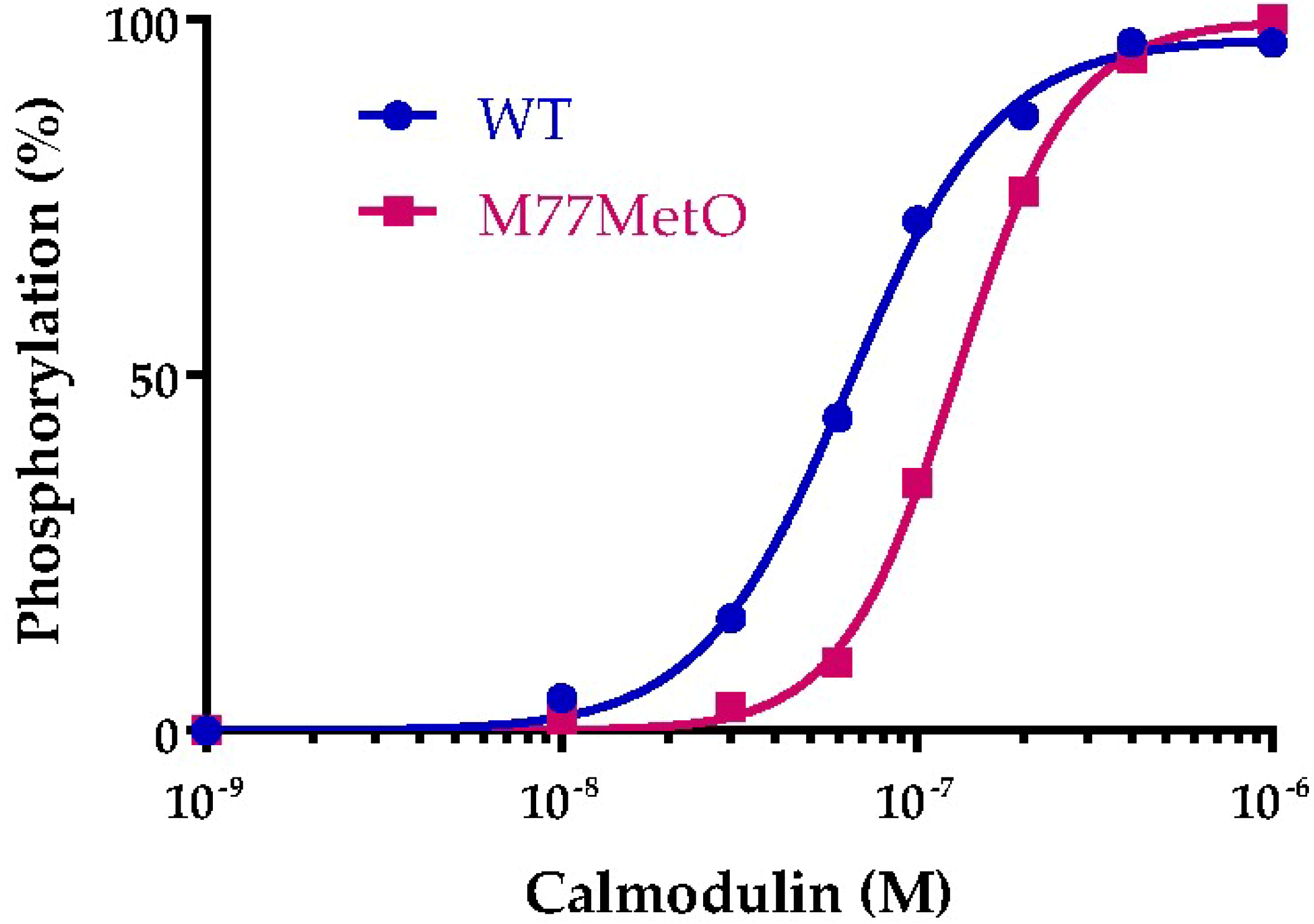
2.1. Recombinant Calmodulins and Assay of Calcium/Calmodulin Dependent Protein Kinase IIα (CaMKIIα)
2.2. Protein Array
2.3. Generation of the Calmodulin M77Q Mouse
2.4. Treadmill Test
2.5. Dobutamine Cardiac Stress Test
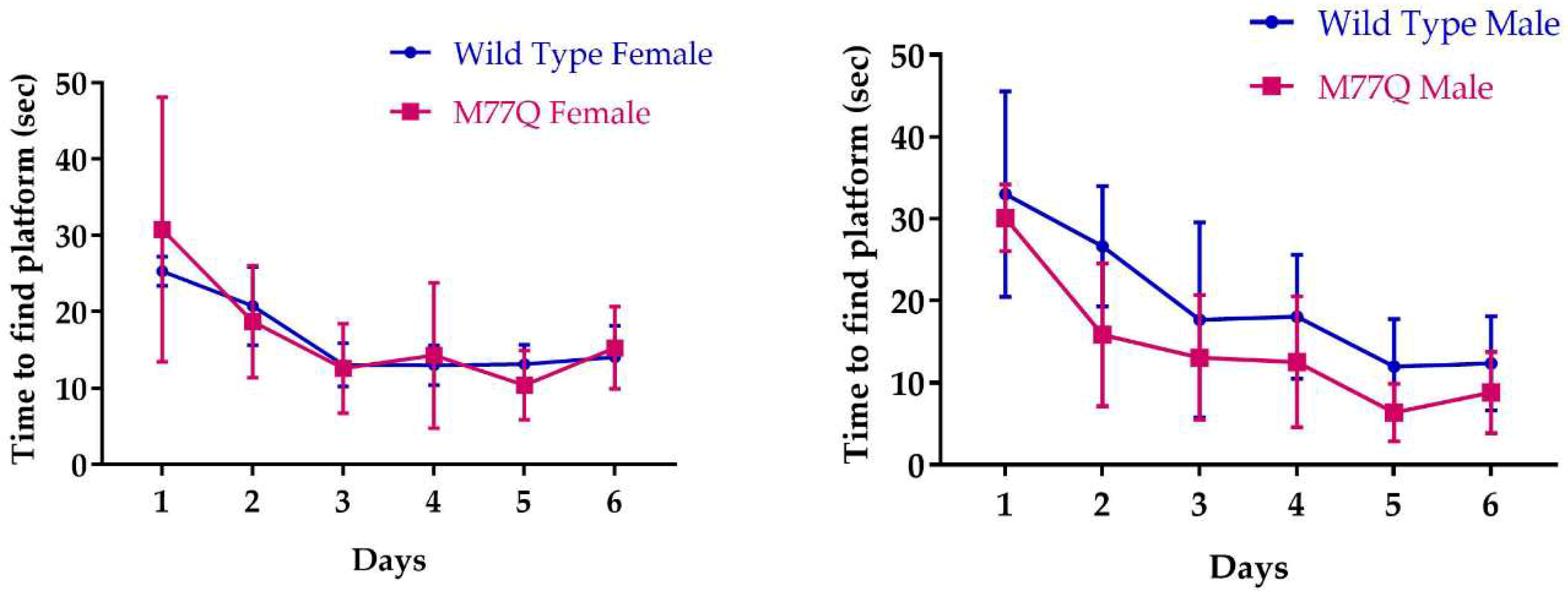
2.6. Spatial Learning Assessment with the Morris Water Maze
2.7. Open Field Test
2.8. Associative Learning Test
2.9. Body Composition
3. Results
3.1. Protein Array
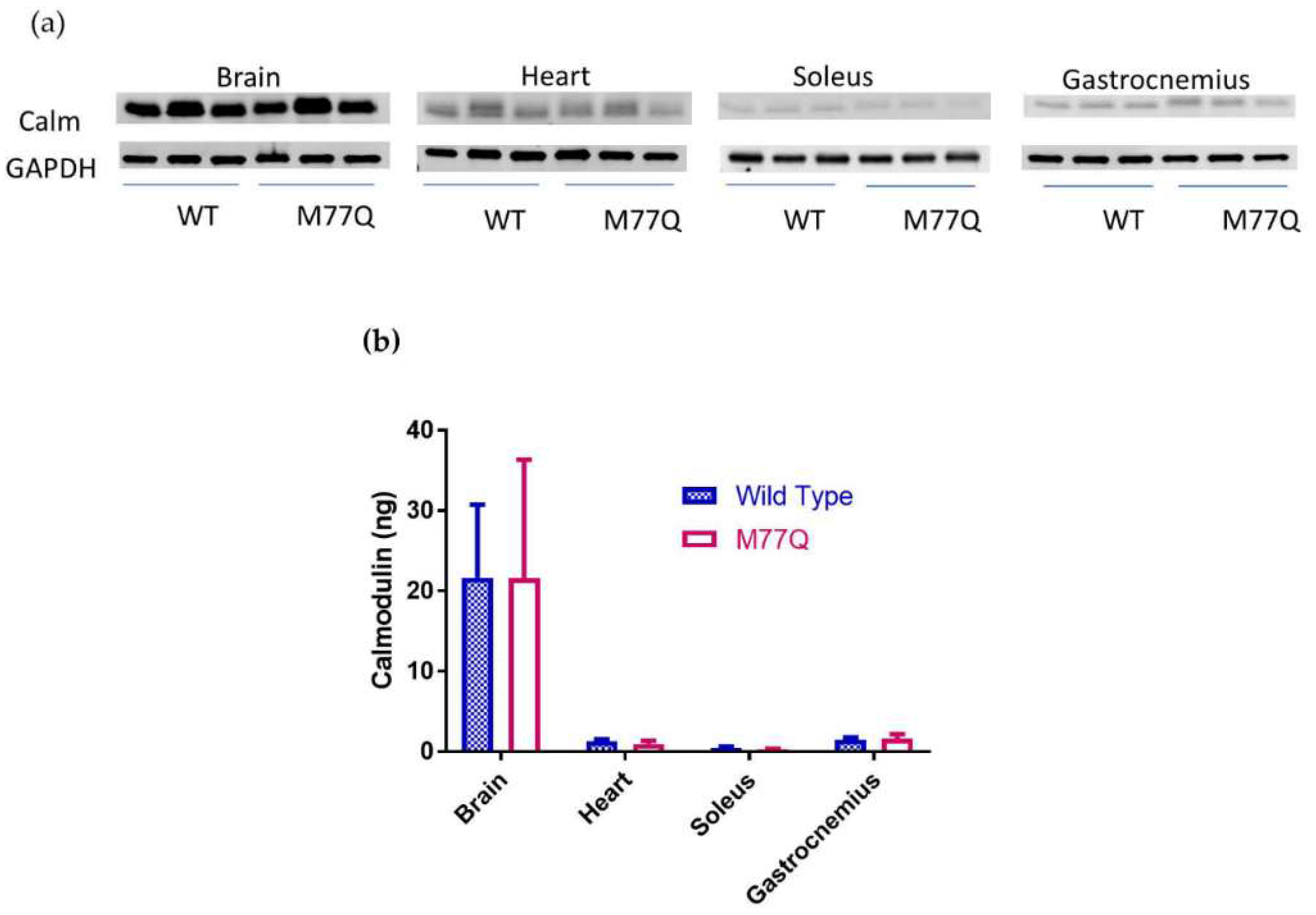
3.2. Calm1 was Mutated in M77Q Mice
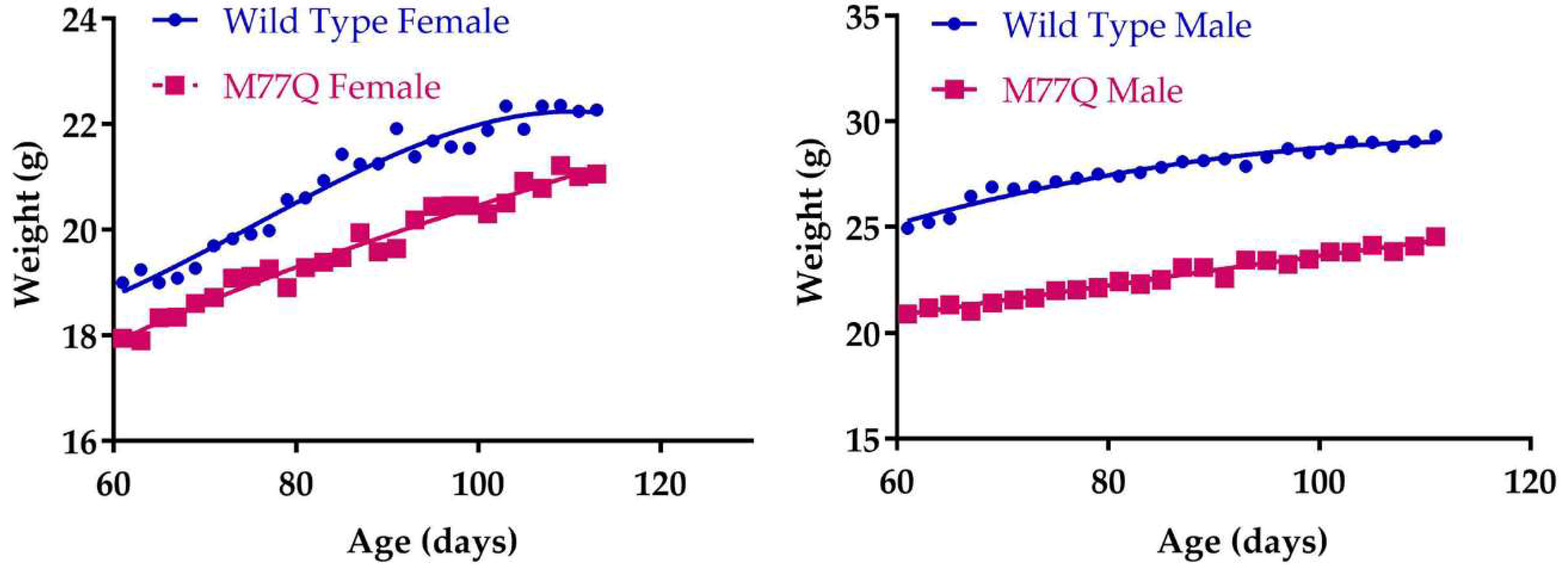
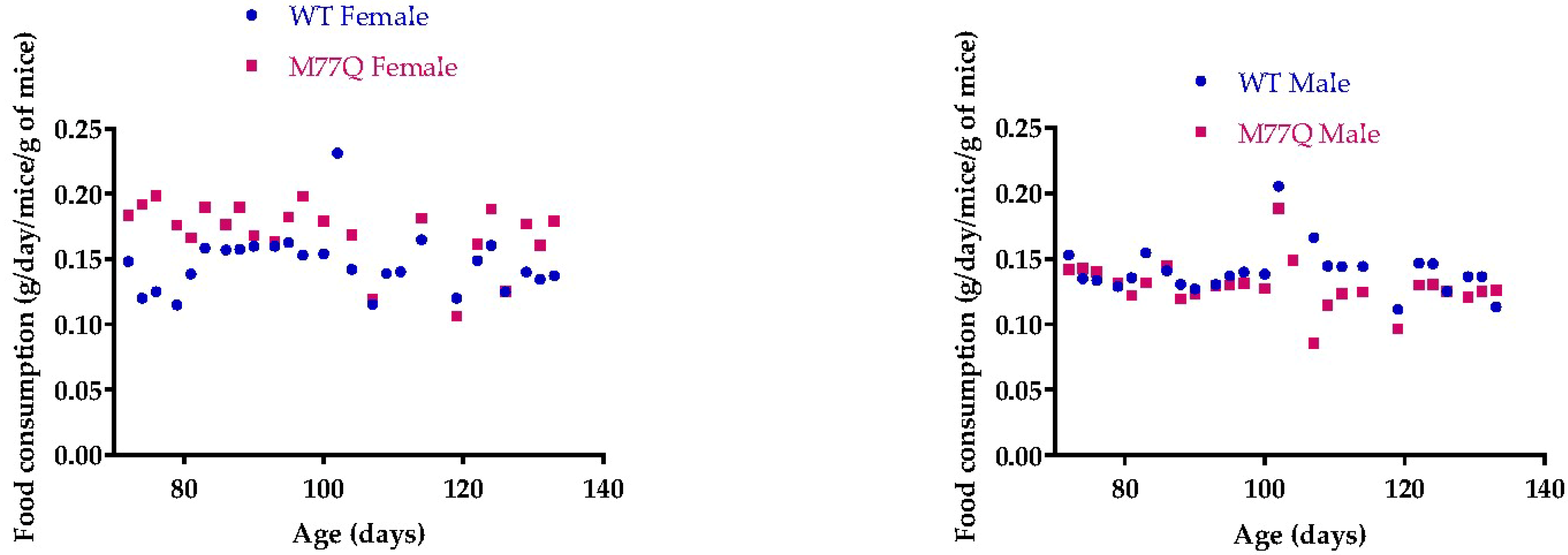
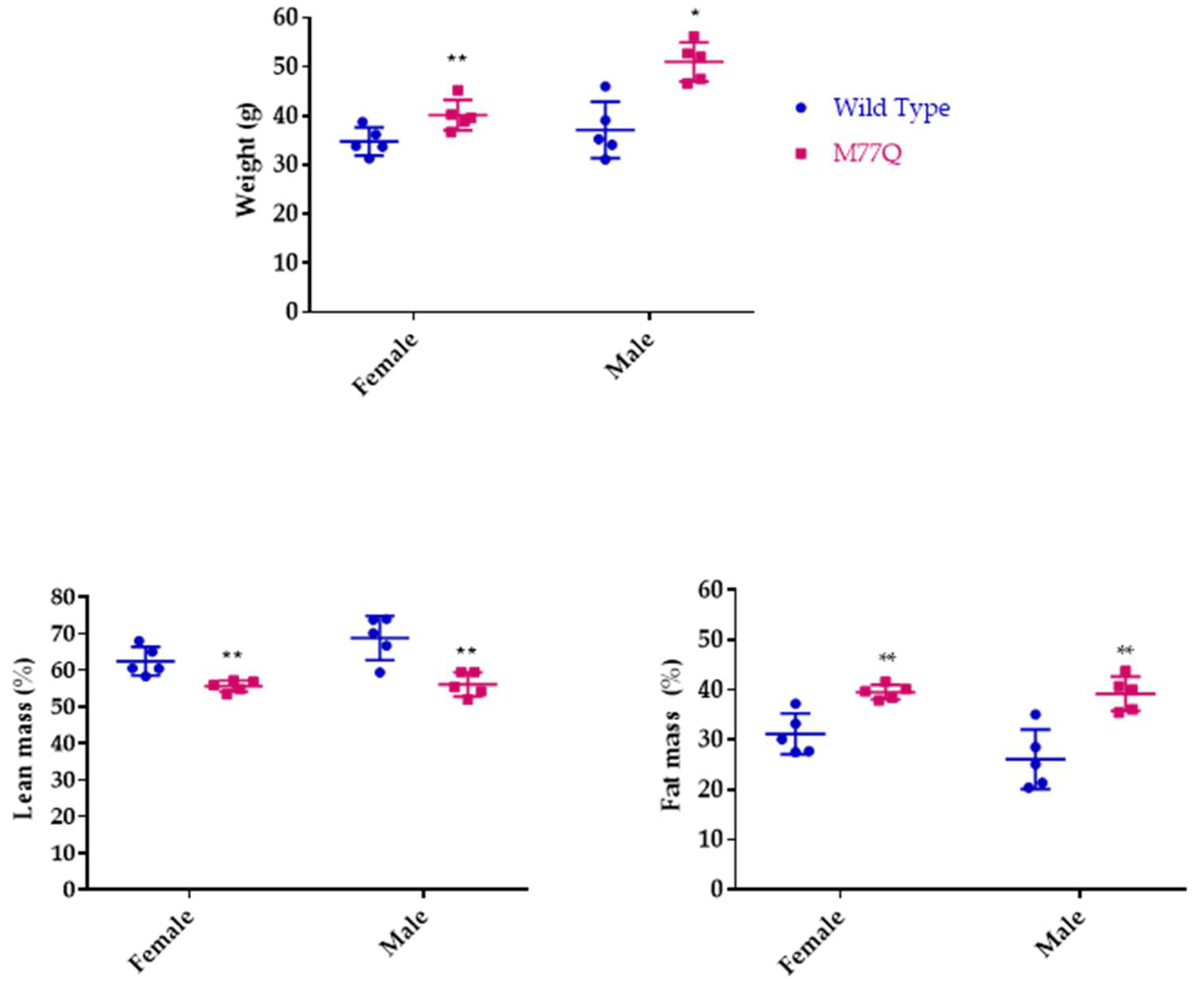
3.3. M77Q Mice are Growth Retarded When Younger and Become Fatter When Older
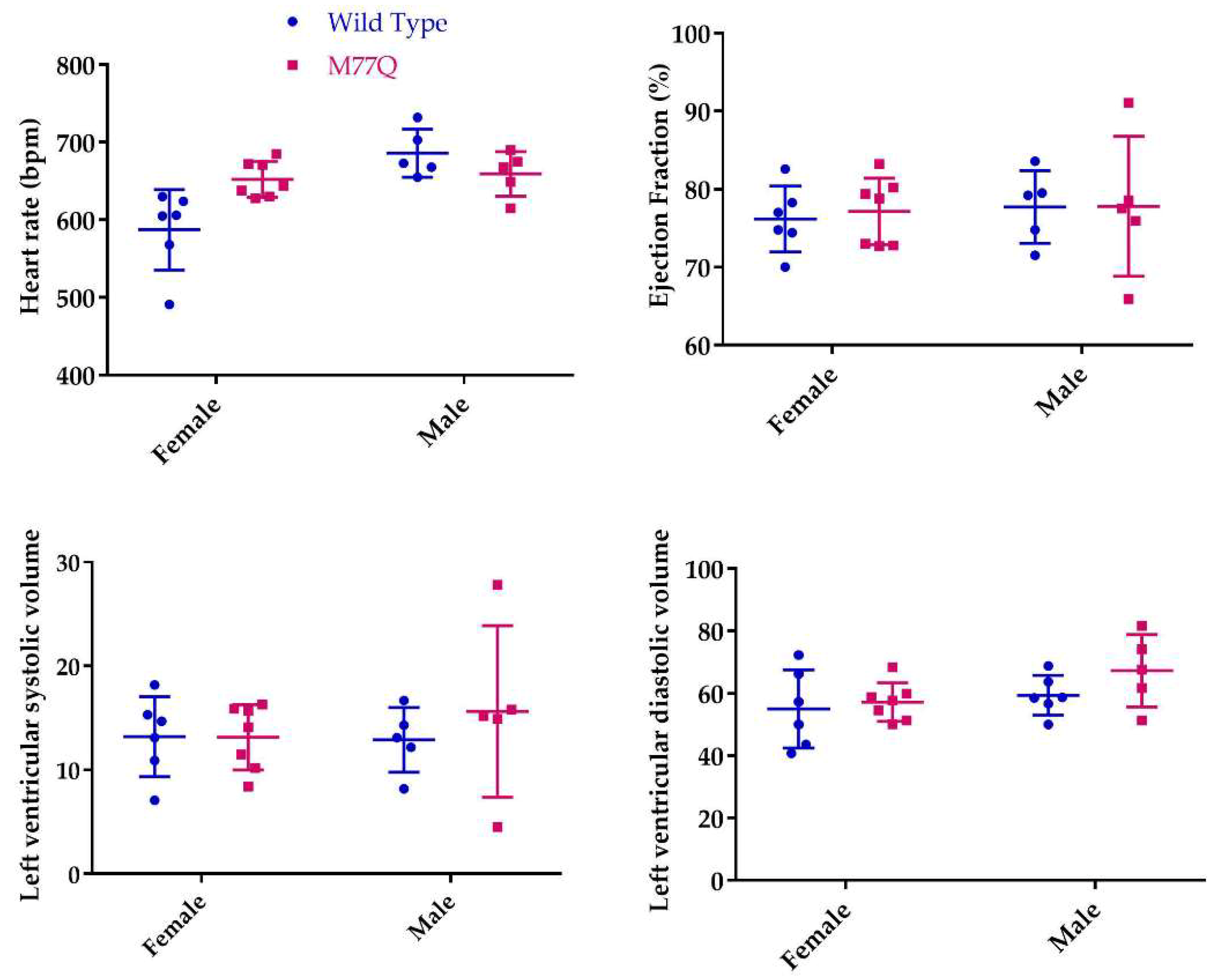
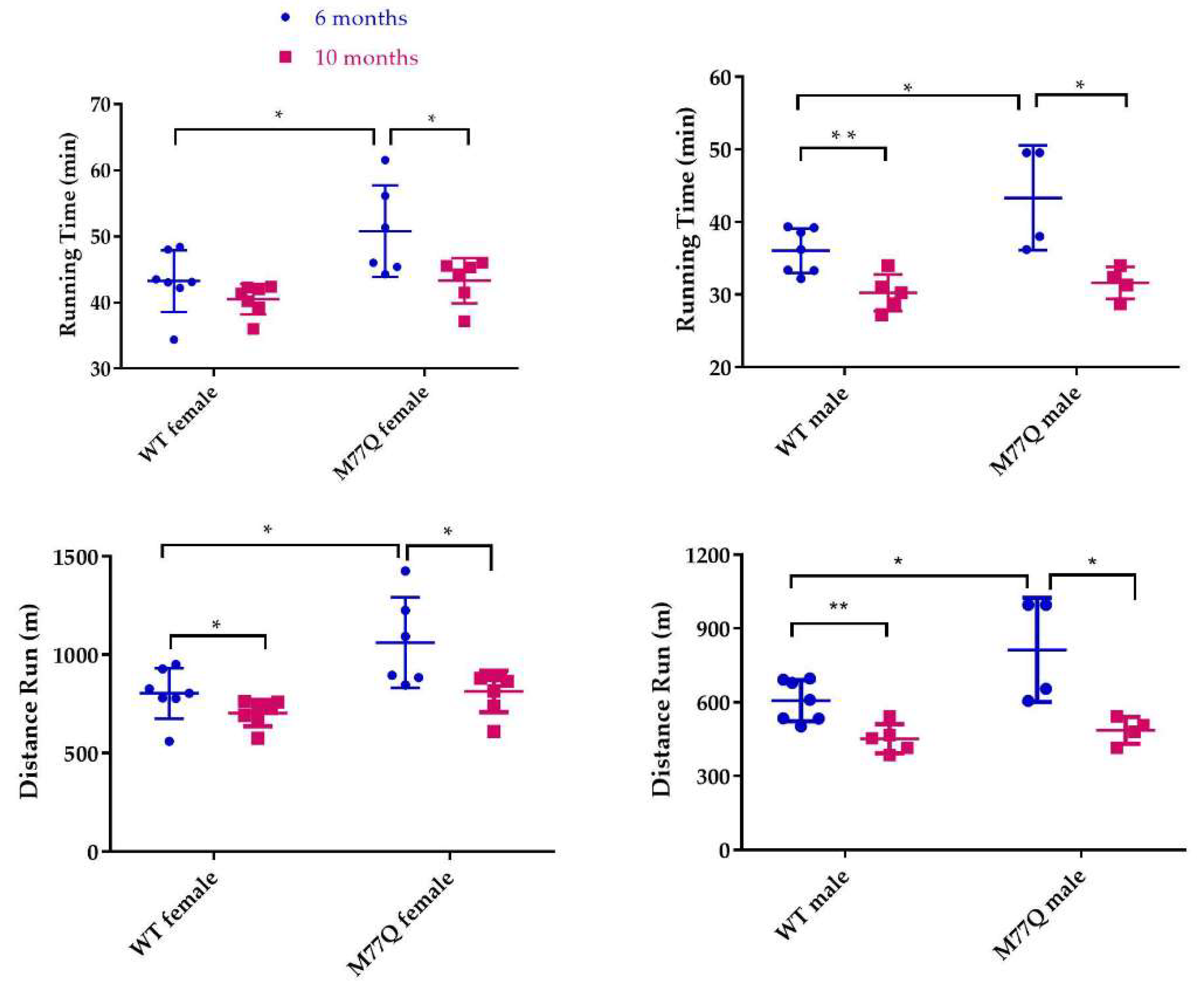
3.4. Cardiac Stress Test and Endurance Running
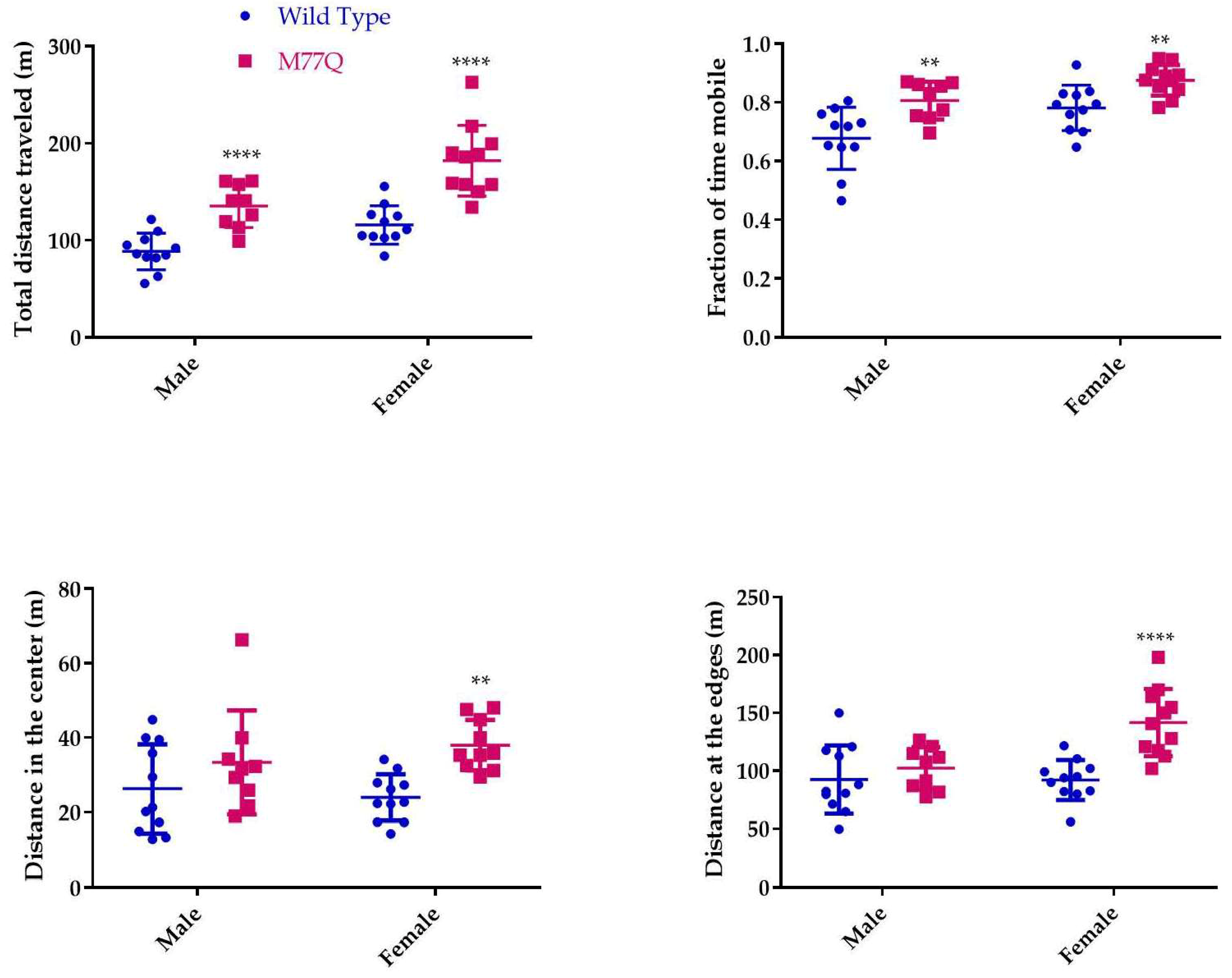
3.5. Neurobehavioral Phenotyping
3.6. Activation of Calcium/Calmodulin Dependent Protein Kinase II
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of Calmodulin M77Q Mice
References
- Levine, R.L.; Mosoni, L.; Berlett, B.S.; Stadtman, E.R. Methionine residues as endogenous antioxidants in proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 15036–15040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, A.; Hajieva, P.; Moosmann, B. Adaptive antioxidant methionine accumulation in respiratory chain complexes explains the use of a deviant genetic code in mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 16496–16501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valley, C.C.; Cembran, A.; Perlmutter, J.D.; Lewis, A.K.; Labello, N.P.; Gao, J.; Sachs, J.N. The methionine-aromatic motif plays a unique role in stabilizing protein structure. J. Biol. Chem. 2012, 287, 34979–34991. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Peterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Hung, R.J.; Spaeth, C.S.; Yesilyurt, H.G.; Terman, J.R. SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat. Cell Biol. 2013, 15, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, R.-J.; Pak, C.W.; Terman, J.R. Direct redox regulation of F-actin assembly and disassembly by Mical. Science 2011, 334, 1710–1713. [Google Scholar] [CrossRef] [PubMed]
- Netzer, N.; Goodenbour, J.M.; David, A.; Dittmar, K.A.; Jones, R.B.; Schneider, J.R.; Boone, D.; Eves, E.M.; Rosner, M.R.; Gibbs, J.S.; et al. Innate immune and chemically triggered oxidative stress modifies translational fidelity. Nature 2009, 462, 522–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kim, D.G.; Kim, B.G.; Yang, W.S.; Hong, J.; Kang, T.; Oh, Y.S.; Kim, K.R.; Han, B.W.; Hwang, B.J.; et al. Promiscuous methionyl-tRNA synthetase mediates adaptive mistranslation to protect cells against oxidative stress. J. Cell Sci. 2014, 127, 4234–4245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.C.; You, Z.; Kim, G.; Levine, R.L. Methionine sulfoxide reductase A is a stereospecific methionine oxidase. Proc. Natl. Acad. Sci. USA 2011, 108, 10472–10477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.C.; Kim, G.; Levine, R.L. Stereospecific oxidation of calmodulin by methionine sulfoxide reductase A. Free Radic. Biol. Med. 2013, 61, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, K.; Kim, J.; Truong, K.; Sherman, M.; Yuan, T.; Ikura, M. Calmodulin target database. J. Struct. Funct. Genomics 2000, 1, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Wu, F.; Li, Z.; Li, T.; Gupta, V.K.; Duan, X.; Jiang, Y. Sulfoxidation regulation of Musa acuminata calmodulin (MaCaM) Influences the functions of MaCaM-binding proteins. Plant Cell Physiol. 2018, 59, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, F.; Rhoads, A.R. Evolutionary aspects of calmodulin. IUBMB Life 2001, 51, 215–221. [Google Scholar] [PubMed]
- Chin, D.; Means, A.R. Methionine to glutamine substitutions in the C-terminal domain of calmodulin impair the activation of three protein kinases. J. Biol. Chem. 1996, 271, 30465–30471. [Google Scholar] [CrossRef] [PubMed]
- Drazic, A.; Miura, H.; Peschek, J.; Le, Y.; Bach, N.C.; Kriehuber, T.; Winter, J. Methionine oxidation activates a transcription factor in response to oxidative stress. Proc. Natl. Acad. Sci. USA 2013, 110, 9493–9498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veredas, F.J.; Cantón, F.R.; Aledo, J.C. Methionine residues around phosphorylation sites are preferentially oxidized in vivo under stress conditions. Sci. Rep. 2017, 7, 40403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigelow, D.J.; Squier, T.C. Thioredoxin-dependent redox regulation of cellular signaling and stress response through reversible oxidation of methionines. Mol Biosyst 2011, 7, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- CRISPR Design. Available online: http://crispr.mit.edu/ (accessed on 11 October 2018).
- Thermo Fisher Scientific. ProtoArray® Applications Guide. Available online: http://tools.thermofisher.com/content/sfs/manuals/protoarray_applicationsguide_man.pdf (accessed on 11 October 2018).
- Lim, J.M.; Lim, J.C.; Kim, G.; Levine, R.L. Myristoylated methionine sulfoxide reductase A is a late endosomal protein. J. Biol. Chem. 2018, 293, 7355–7366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kregel, K.C.; Allen, D.L.; Booth, F.W.; Fleshner, M.R.; Henriksen, E.J.; Musch, T.; O’Leary, D.; Parks, C.; Poole, D.; Ra’anan, A.; et al. Resource Book for the Design of Animal Exercise Protocols. Available online: http://www.the-aps.org/mm/SciencePolicy/AnimalResearch/Publications/Animal-Exercise-Protocols/book14824.pdf (accessed on 7 February 2006).
- Pritchett, K.; Mulder, G.B. Open-field assessment of spontaneous activity. J. Amer. Assoc. Lab. Animal Sci. 2003, 42, 57–58. [Google Scholar]
- Kim, J.J.; Jung, M.W. Neural circuits and mechanisms involved in Pavlovian fear conditioning: A critical review. Neurosci. Biobehav. Rev. 2006, 30, 188–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, G.B.; Pritchett, K. The Morris water maze. J. Amer. Assoc. Lab. Animal Sci. 2003, 42, 49–50. [Google Scholar]
- Mignogna, P.; Viggiano, D. Brain distribution of genes related to changes in locomotor activity. Physiol. Behav. 2010, 99, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Rainnie, D.G.; Greene, R.W.; Tonegawa, S. Abnormal fear response and aggressive behavior in mutant mice deficient for alpha-calcium-calmodulin kinase II. Science 1994, 266, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Rellos, P.; Pike, A.C.W.; Niesen, F.H.; Salah, E.; Lee, W.H.; von Delft, F.; Knapp, S. Structure of the CaMKIIδ/Calmodulin complex reveals the molecular mechanism of CaMKII kinase activation. PLOS Biol. 2010, 8, e1000426. [Google Scholar] [CrossRef] [PubMed]
- Persechini, A.; Cronk, B. The relationship between the free concentrations of Ca2+ and Ca2+-calmodulin in intact cells. J. Biol. Chem. 1999, 274, 6827–6830. [Google Scholar]
- Finn, B.E.; Forsén, S. The evolving model of calmodulin structure, function and activation. Structure 1995, 3, 7–11. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volume of LV in Systole (mm3) | Volume of LV in Diastole (mm3) | Ejection Fraction (%) | Diam LV Chamber in Systole (mm) | Diam LV Chamber in Diastole (mm) | Heart Rate (Beats Per Minute) | |
|---|---|---|---|---|---|---|
| Baseline | ||||||
| WT female | 29.2 ± 3.4 | 72.3 ± 7.4 | 60 ± 0.8 | 2.8 ± 0.1 | 4 ± 0.17 | 491.9 ± 45.8 |
| M77Q female | 30.5 ± 2.2 | 72.1 ± 5.1 | 58.9 ± 1.5 | 2.8 ± 0.1 | 4 ± 0.12 | 553.9 ± 34.4 |
| WT male | 31 ± 3.1 | 77 ± 4.7 | 59.8 ± 2.4 | 2.9 ± 0.1 | 4.2 ± 0.1 | 502.3 ± 33.8 |
| M77Q female | 30.78 ± 2.5 | 78.8 ± 5.5 | 60.9 ± 1.7 | 2.8 ± 0.08 | 4.2 ± 0.12 | 564.4 ± 18.3 |
| Low Dose | ||||||
| WT female | 16 ± 3.7 | 59 ± 9.1 | 73 ± 2.7 | 2.2 ± 0.2 | 3.7 ± 0.2 | 558 ± 40.4 |
| M77Q female | 18.1 ± 5.3 | 60.4 ± 7.4 | 70.7 ± 5.3 | 2.3 ± 0.3 | 3.8 ± 0.2 | 625.1 ± 18.4 |
| WT male | 18.8± 5.1 | 65.3 ± 7.6 | 71.7 ± 5 | 2.3 ± 0.2 | 3.9 ± 0.2 | 623.5 ± 37.5 |
| M77Q female | 19 ± 8.6 | 66.7 ± 11.9 | 73.1 ± 9 | 2.3 ± 0.5 | 3.9 ± 0.3 | 630.6 ± 23.6 |
| High dose | ||||||
| WT female | 13.2 ± 3.5 | 55 ± 11.5 | 76.2 ± 3.9 | 2.0 ± 0.2 | 3.6 ± 0.3 | 587.3 ± 47.4 |
| M77Q female | 13.2 ± 2.9 | 57.2 ± 5.7 | 77.2 ± 4.0 | 2.0 ± 0.2 | 3.7 ± 0.2 | 652.6 ± 21.3 |
| WT male | 13.4 ± 2.8 | 59.4 ± 5.8 | 77.6 ± 3.8 | 2.0 ± 0.2 | 3.7 ± 0.2 | 667.3 ± 49.2 |
| M77Q female | 15.6 ± 7.4 | 67.3 ± 10.3 | 77.8 ± 8 | 2.1 ± 0.4 | 3.9 ± 0.3 | 659.4 ± 25.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marimoutou, M.; Springer, D.A.; Liu, C.; Kim, G.; Levine, R.L. Oxidation of Methionine 77 in Calmodulin Alters Mouse Growth and Behavior. Antioxidants 2018, 7, 140. https://doi.org/10.3390/antiox7100140
Marimoutou M, Springer DA, Liu C, Kim G, Levine RL. Oxidation of Methionine 77 in Calmodulin Alters Mouse Growth and Behavior. Antioxidants. 2018; 7(10):140. https://doi.org/10.3390/antiox7100140
Chicago/Turabian StyleMarimoutou, Méry, Danielle A. Springer, Chengyu Liu, Geumsoo Kim, and Rodney L. Levine. 2018. "Oxidation of Methionine 77 in Calmodulin Alters Mouse Growth and Behavior" Antioxidants 7, no. 10: 140. https://doi.org/10.3390/antiox7100140
APA StyleMarimoutou, M., Springer, D. A., Liu, C., Kim, G., & Levine, R. L. (2018). Oxidation of Methionine 77 in Calmodulin Alters Mouse Growth and Behavior. Antioxidants, 7(10), 140. https://doi.org/10.3390/antiox7100140