Effects of Lespedeza Bicolor Extract on Regulation of AMPK Associated Hepatic Lipid Metabolism in Type 2 Diabetic Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Lespedeza Bicolor Extracts (LBE)
2.2. Animals and Experimental Design
2.3. Hemoglobin A1c (HbA1c)
2.4. Plasma Glutamate Oxaloacetate Transaminase (GOT) and Glutamate Pyruvate Transaminase (GPT)
2.5. Lipid Profile Analysis
2.6. Histological Analysis
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Effects of LBE Supplementation on Body Weight, Food Intake, and FBG Level in T2DM
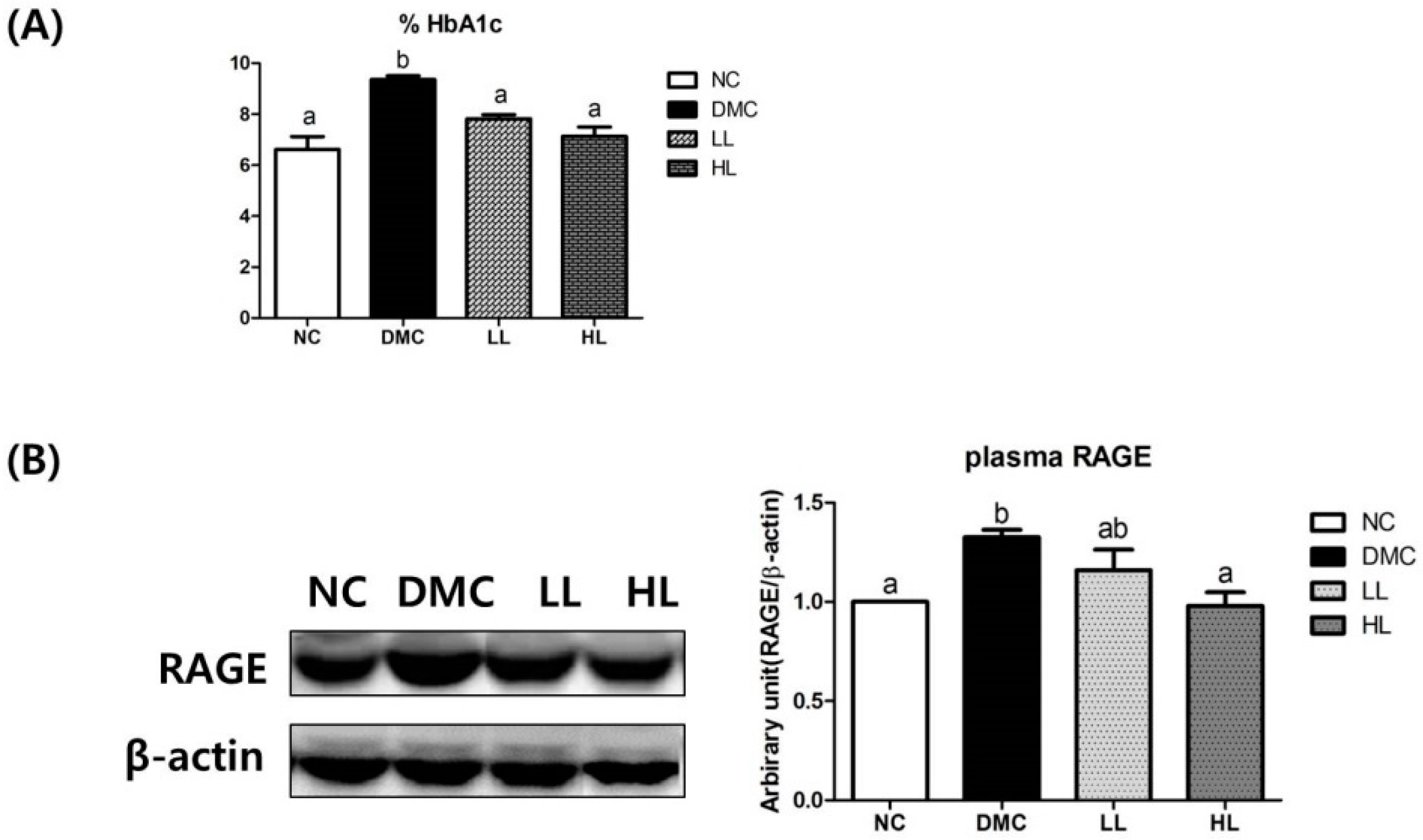
3.2. Effects of LBE Supplementation on Glycation Products in T2DM Mice
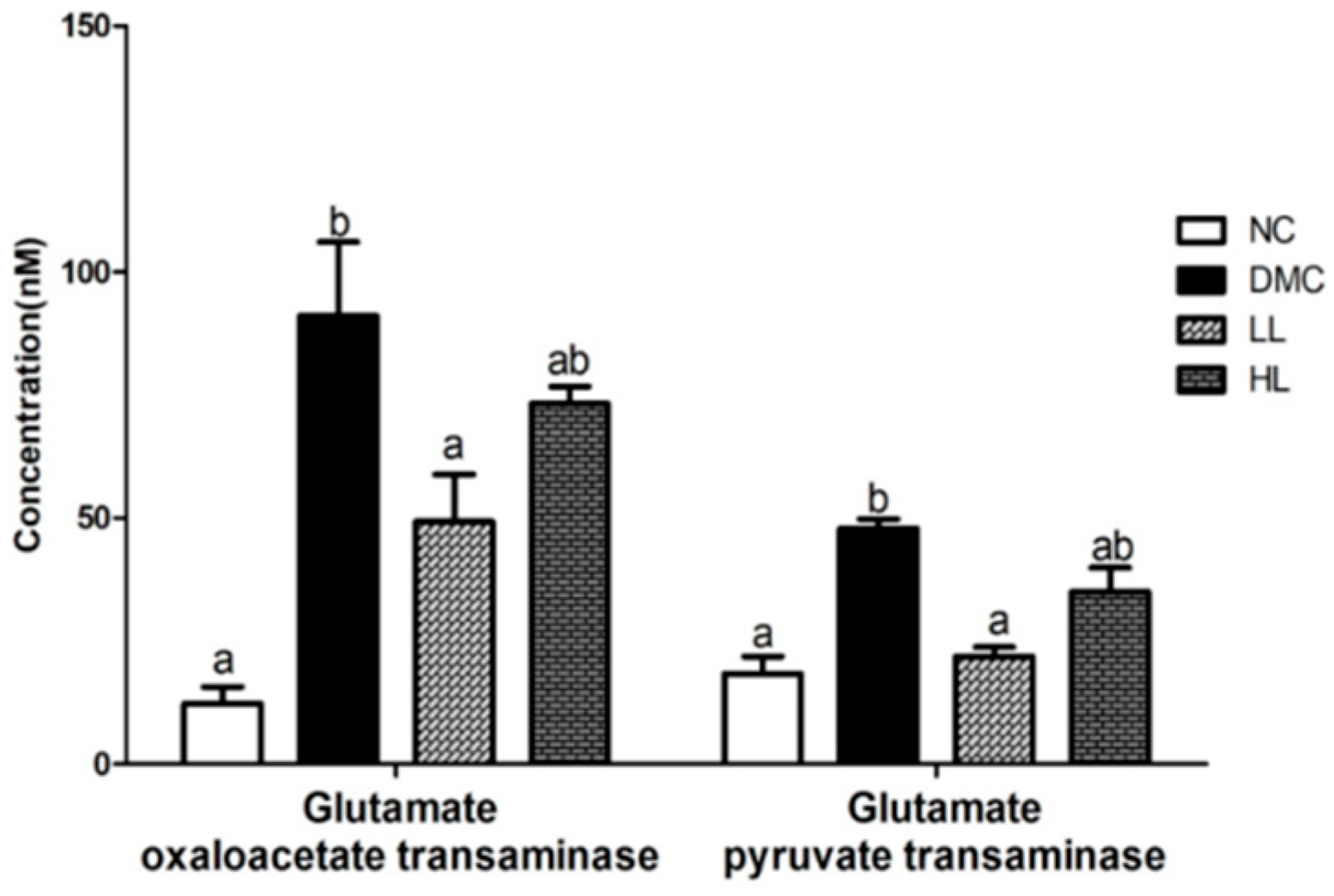
3.3. Effects of LBE Supplementation on Plasma GOT and GPT in T2DM Mice
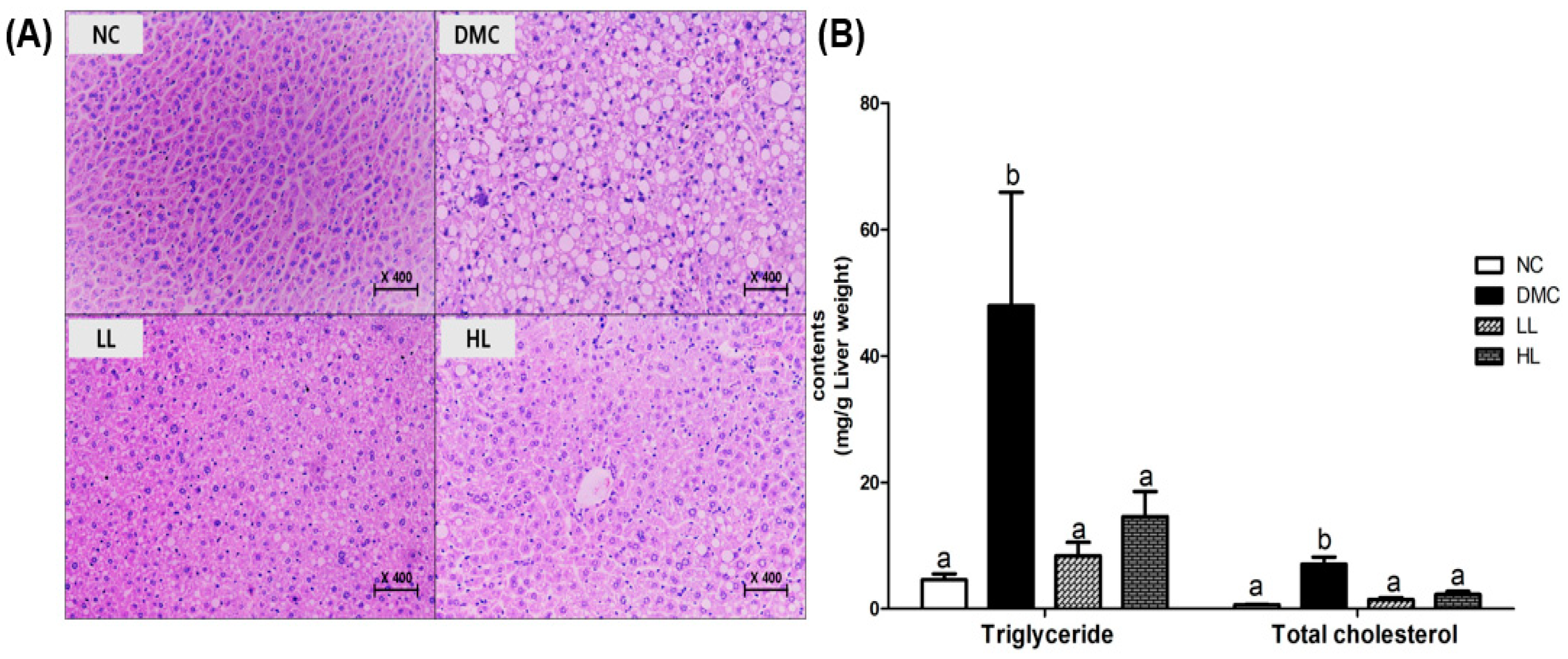
3.4. Effects of LBE on Hepatic Morphology and Lipid Profiles in T2DM Mice
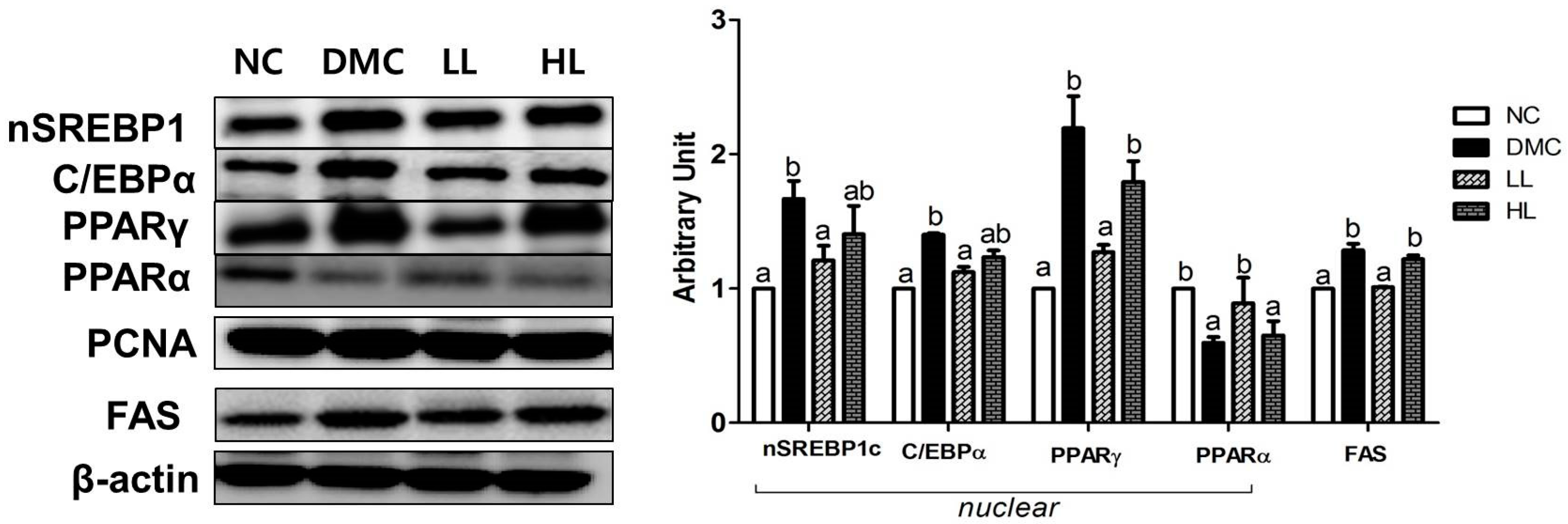
3.5. Effects of LBE Supplementation on Hepatic Protein Levels of Lipid Metabolism Related Markers in T2DM Mice
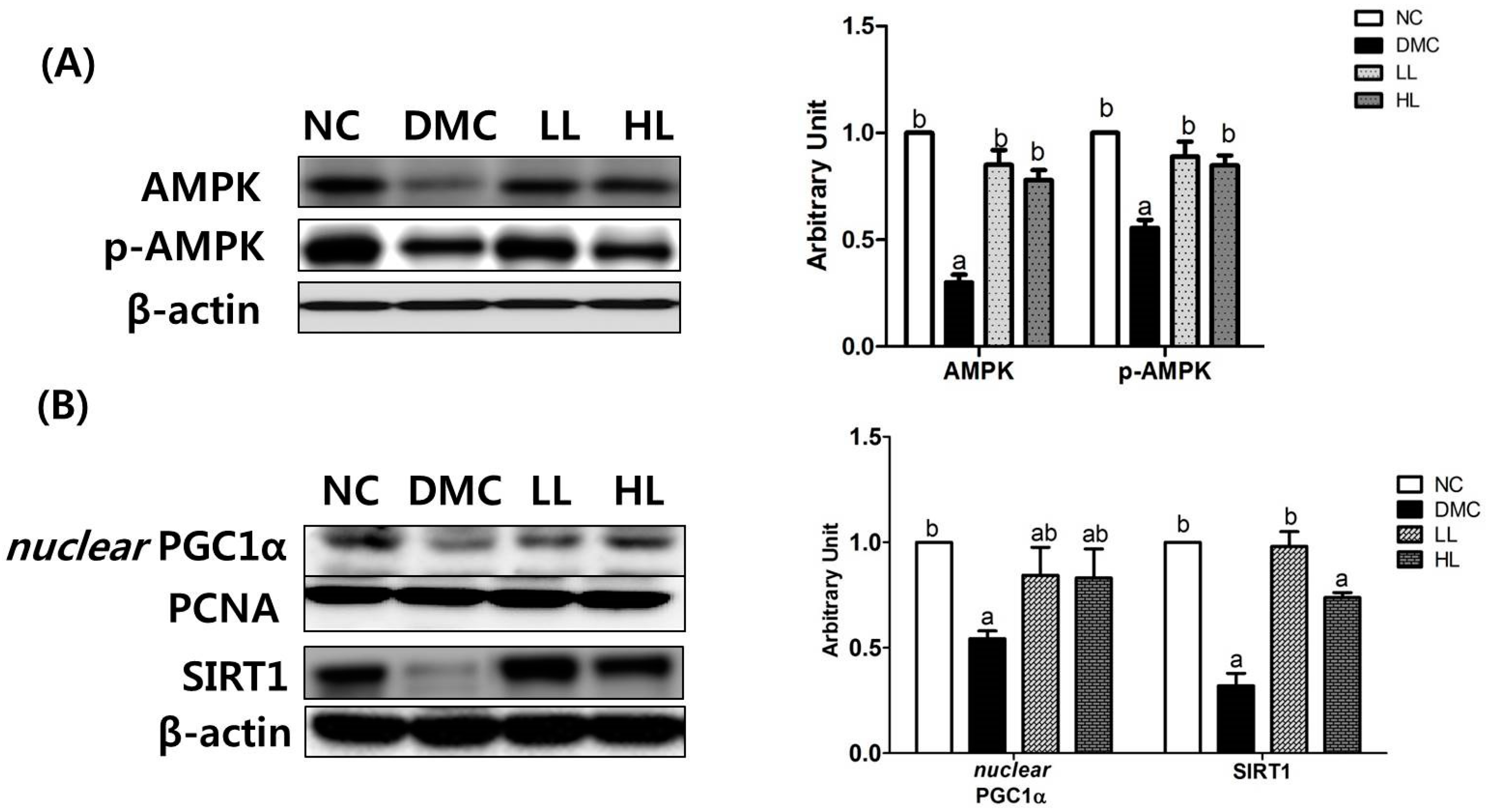
3.6. Effects of LBE Supplementation on Hepatic Protein Levels of Energy Metabolism Related Markers in T2DM Mice
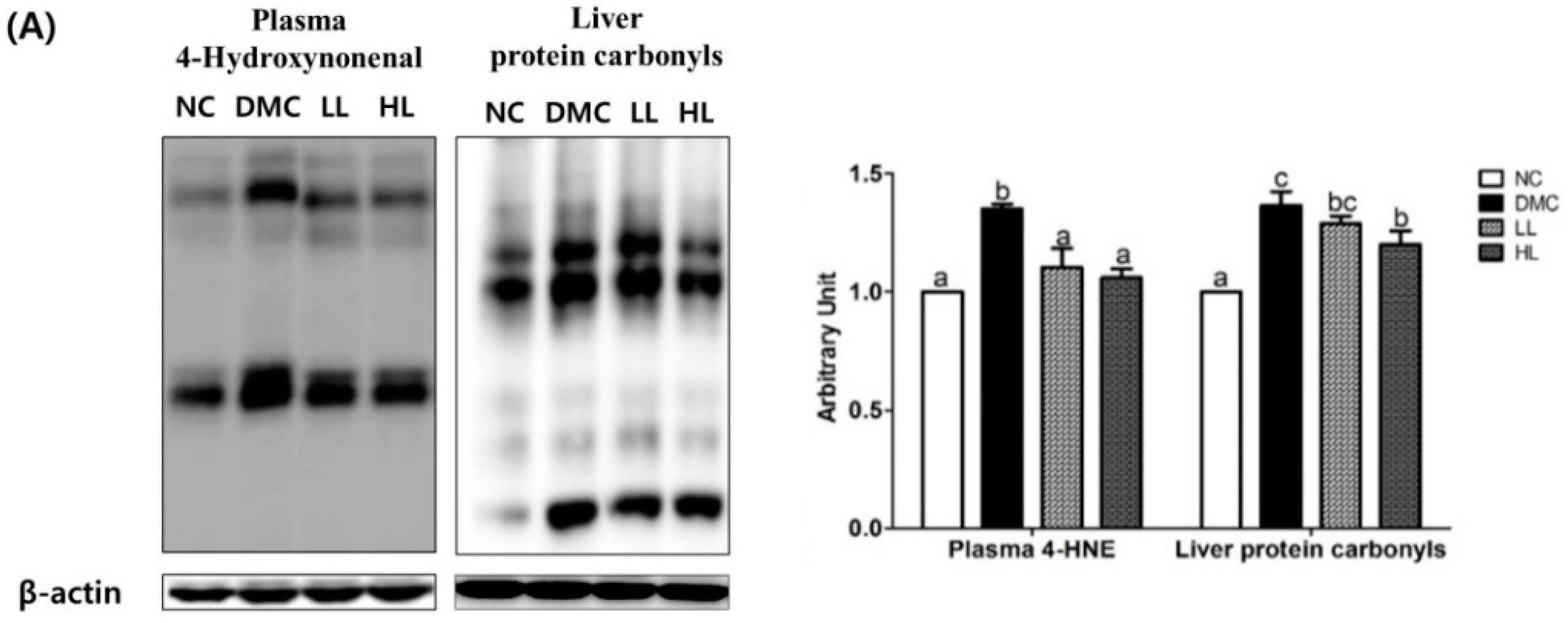
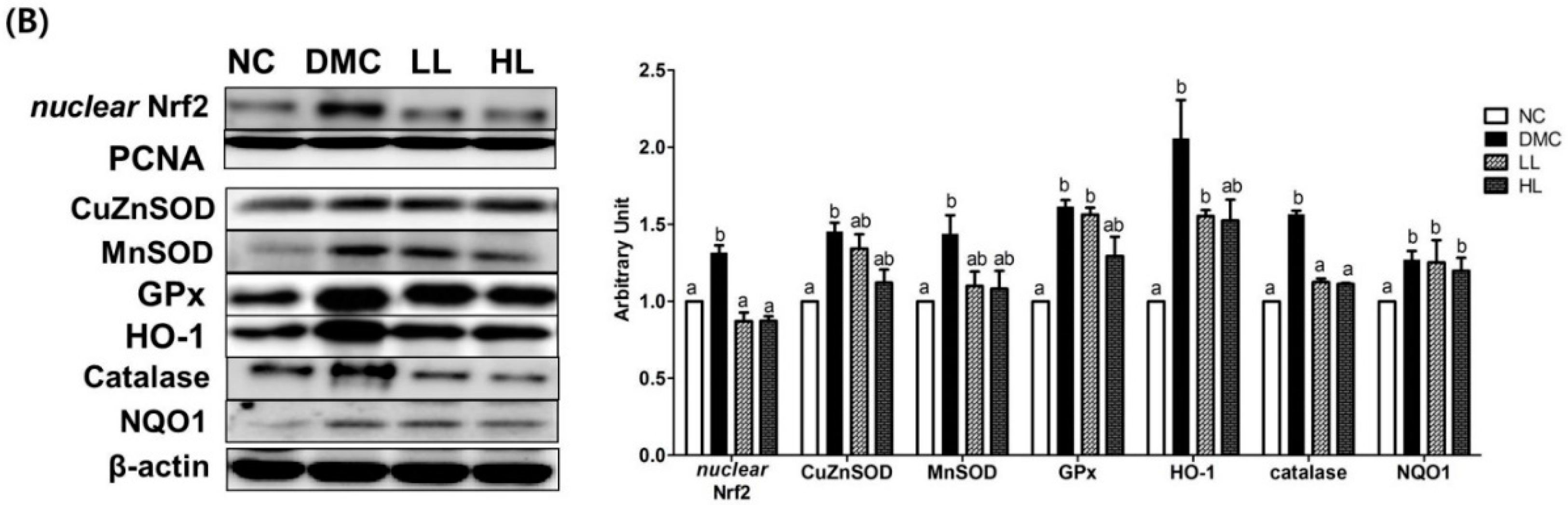
3.7. Effects of LBE Supplementation on Plasma and Hepatic Protein Levels of Oxidative Stress Markers T2DM Mice
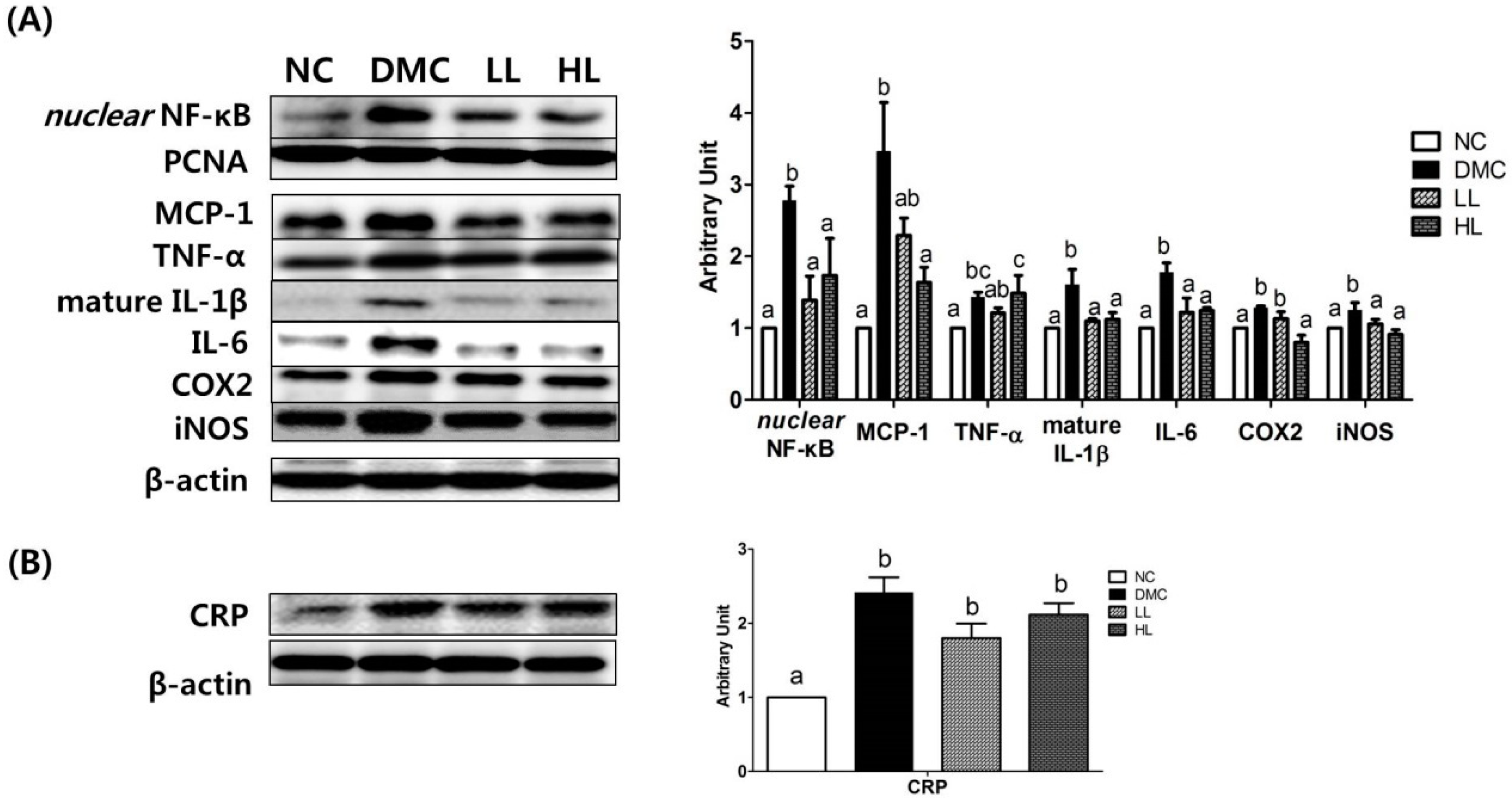
3.8. Effects of LBE Supplementation on Hepatic Protein Levels of Inflammatory Response Related Markers in T2DM Mice
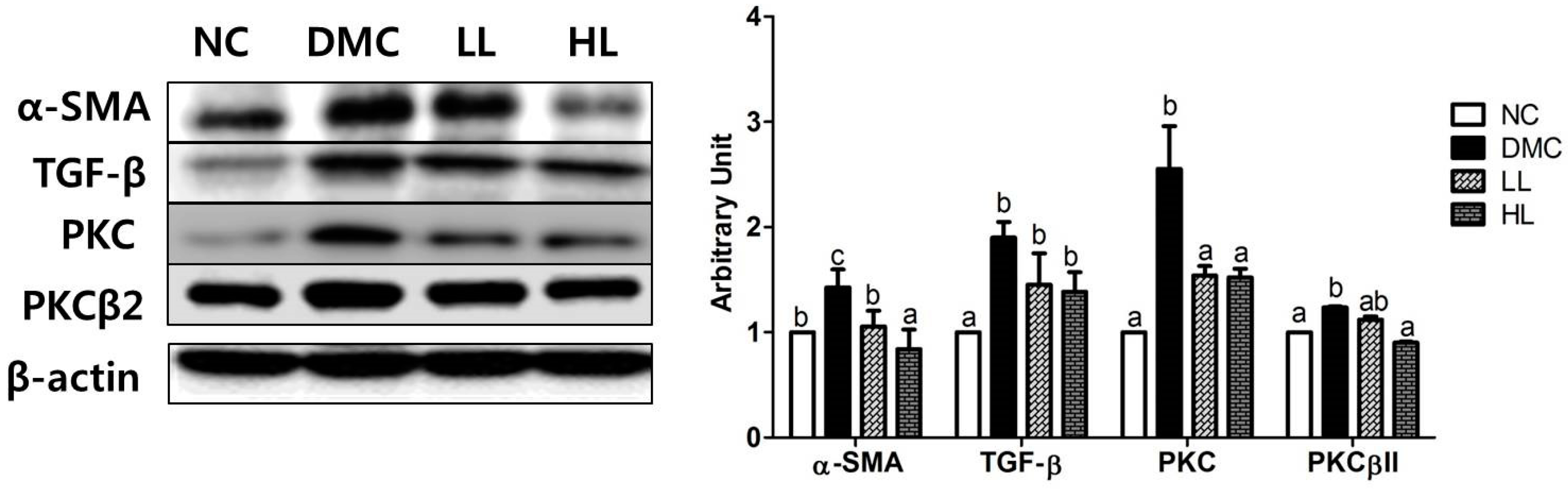
3.9. Effects of LBE Supplementation on Hepatic Fibrosis in T2DM Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AGE | Advanced glycation end products |
| AMPK | adenosine monophosphate activation kinase |
| AUC | Area under the curve |
| BSA | Bovine serum albumin |
| BW | Body weight |
| C/EBPα | CAAT box/enhancer binding protein alpha |
| DM | Diabetes mellitus |
| FAS | Fatty acid synthase |
| FBG | Fasting blood glucose |
| 4-HNE | Four-hydroxynonenal |
| GOT | Glutamate oxaloacetate transaminase |
| GPT | Glutamate pyruvate transaminase |
| GPx | Glutathione peroxidase |
| H&E | Hematoxylin and eosin |
| HSCs | Hepatic stellate cells |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin-6 |
| LB | Lespedeza Bicolor |
| MGO | Methylglyoxal |
| NC | Normal control |
| NF-κB | nuclear factor kappa B |
| NQO1 | NAD(P)H degydrogenase quinine 1 |
| Nrf2 | Nuclear factor erythroid2-related factor 2 |
| OGTT | Oral glucose tolerance test |
| PVDF | Polyvinylidene fluoride |
| SIRT1 | Sirtuin1 |
| SOD | Superoxide dismutase |
| STZ | Streptozotocin |
| RAGE | Advanced glycation end products receptor |
| ROS | Reactive oxygen species |
| RNS | Reactive nitrogen species |
| SREBP1 | Sterol regulatory element-binding protein 1 |
| T2DM | Type 2 diabetes mellitus |
| TC | Total cholesterol |
| TG | Triglyceride |
| TNFα | Tumor necrosis factor α |
References
- Blair, M. Diabetes Mellitus Review. Urol. Nurs. 2016, 36, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, O.B. Diabetes mellitus and malfunctions of insulin secretion. Ugeskr. Laeger. 1997, 159, 7118–7119. [Google Scholar] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef] [PubMed]
- Leite, N.C.; Villela-Nogueira, C.A.; Cardoso, C.R.; Salles, G.F. Non-alcoholic fatty liver disease and diabetes: From physiopathological interplay to diagnosis and treatment. World J. Gastroenterol. 2014, 20, 8377–8392. [Google Scholar] [CrossRef]
- Stefan, N.; Haring, H.U. The metabolically benign and malignant fatty liver. Diabetes 2011, 60, 2011–2017. [Google Scholar] [CrossRef]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef]
- Williams, K.H.; Shackel, N.A.; Gorrell, M.D.; McLennan, S.V.; Twigg, S.M. Diabetes and nonalcoholic Fatty liver disease: A pathogenic duo. Endocr. Rev. 2013, 34, 84–129. [Google Scholar] [CrossRef]
- Marchesini, G.; Marzocchi, R. Metabolic syndrome and NASH. Clin. Liv. Dis. 2007, 11, 105–117. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Zhang, H.; Davies, K.J.A.; Forman, H.J. Oxidative stress response and Nrf2 signaling in aging. Free Radic. Biol. Med. 2015, 88, 314–336. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, M.; Yu, H.; Wang, W.; Han, L.; Chen, Q.; Ruan, J.; Wen, S.; Zhang, Y.; Wang, T. Mangiferin Improves Hepatic Lipid Metabolism Mainly Through Its Metabolite-Norathyriol by Modulating SIRT-1/AMPK/SREBP-1c Signaling. Front. Pharmacol. 2018, 9, 201. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ramiro, I.; Vauzour, D.; Minihane, A.M. Polyphenols and non-alcoholic fatty liver disease: Impact and mechanisms. Proc. Nutr. Soc. 2016, 75, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florencio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Hossaine, M.D.; Park, S.C. A potential anti-inflammation activity and depigmentation effect of Lespedeza bicolor extract and its fractions. Saudi J. Boil. Sci. 2016, 23, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Maximov, O.B.; Kulesh, N.I.; Stepanenko, L.S.; Dmitrenok, P.S. New prenylated isoflavanones and other constituents of Lespedeza bicolor. Fitoterapia 2004, 75, 96–98. [Google Scholar] [CrossRef]
- Miyase, T.; Sano, M.; Yoshino, K.; Nonaka, K. Antioxidants from Lespedeza homoloba (II). Phytochemistry 1999, 52, 311–319. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, J.H.; Wahedi, H.M.; Pak, C.; Lee, C.H.; Yeo, E.J.; Lim, Y.; Ha, S.K.; Choi, I.; Kim, S.Y. Lespedeza bicolor ameliorates endothelial dysfunction induced by methylglyoxal glucotoxicity. Phytomed. Int. J. Phytother. Phytopharm. 2017, 36, 26–36. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, J.H. Therapeutic Potential of Lespedeza bicolor to Prevent Methylglyoxal-Induced Glucotoxicity in Familiar Diabetic Nephropathy. J. Clin. Med. 2019, 8, 1138. [Google Scholar] [CrossRef]
- Zhang, M.; Lv, X.Y.; Li, J.; Xu, Z.G.; Chen, L. The characterization of high-fat diet and multiple low-dose streptozotocin induced type 2 diabetes rat model. Exp. Diabetes Res. 2008, 2008, 704045. [Google Scholar] [CrossRef]
- Lee, C.W.; Seo, J.Y.; Lee, J.; Choi, J.W.; Cho, S.; Bae, J.Y.; Sohng, J.K.; Kim, S.O.; Kim, J.; Park, Y.I. 3-O-Glucosylation of quercetin enhances inhibitory effects on the adipocyte differentiation and lipogenesis. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 95, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jiang, H. Therapeutic Effects of Quercetin on Inflammation, Obesity, and Type 2 Diabetes. Mediat. Inflamm. 2016, 2016, 9340637. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.L.; Germolec, D.R.; Zheng, J.F.; Kooistra, L.; Auttachoat, W.; Smith, M.J.; White, K.L.; Elmore, S.A. Genistein protects female nonobese diabetic mice from developing type 1 diabetes when fed a soy- and alfalfa-free diet. Toxicol. Pathol. 2015, 43, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Jung, J.H. Non-alcoholic fatty liver disease and flavonoids: Current perspectives. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 17–24. [Google Scholar] [CrossRef]
- Al-Dosari, D.I.; Ahmed, M.M.; Al-Rejaie, S.S.; Alhomida, A.S.; Ola, M.S. Flavonoid Naringenin Attenuates Oxidative Stress, Apoptosis and Improves Neurotrophic Effects in the Diabetic Rat Retina. Nutrients 2017, 9, 1161. [Google Scholar] [CrossRef]
- Do, M.H.; Hur, J.; Choi, J.; Kim, Y.; Park, H.Y. Spatholobus suberectus Ameliorates Diabetes-Induced Renal Damage by Suppressing Advanced Glycation End Products in db/db Mice. Int. J. Mol. Sci. 2018, 19, 2774. [Google Scholar] [CrossRef]
- Ann, J.Y.; Eo, H.; Lim, Y. Mulberry leaves (Morus alba L.) ameliorate obesity-induced hepatic lipogenesis, fibrosis, and oxidative stress in high-fat diet-fed mice. Genes. Nutr. 2015, 10, 46. [Google Scholar] [CrossRef]
- He, L.; Zhou, X.; Huang, N.; Li, H.; Tian, J.; Li, T.; Yao, K.; Nyachoti, C.M.; Kim, S.W.; Yin, Y. AMPK Regulation of Glucose, Lipid and Protein Metabolism: Mechanisms and Nutritional Significance. Curr. Protein Pept. Sci. 2017, 18, 562–570. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef]
- Thirupathi, A.; Souza, C.T. Multi-regulatory network of ROS: The interconnection of ROS, PGC-1 alpha, and AMPK-SIRT1 during exercise. J. Physiol. Biochem. 2017, 73, 487–494. [Google Scholar] [CrossRef]
- Shang, J.; Chen, L.L.; Xiao, F.X.; Sun, H.; Ding, H.C.; Xiao, H. Resveratrol improves non-alcoholic fatty liver disease by activating AMP-activated protein kinase. Acta Pharmacol. Sin. 2008, 29, 698–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joven, J.; Espinel, E.; Rull, A.; Aragones, G.; Rodriguez-Gallego, E.; Camps, J.; Micol, V.; Herranz-Lopez, M.; Menendez, J.A.; Borras, I.; et al. Plant-derived polyphenols regulate expression of miRNA paralogs miR-103/107 and miR-122 and prevent diet-induced fatty liver disease in hyperlipidemic mice. Biochim. Biophys. Acta 2012, 1820, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Qiao, X.; Dong, B. Neonatal exposure to genistein ameliorates high-fat diet-induced non-alcoholic steatohepatitis in rats. Br. J. Nutr. 2011, 106, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.M.; Wang, Y.J.; Cui, M.; Luo, W.J.; Wang, X.J.; Barber, P.A.; Chen, Z.Y. A dietary polyphenol resveratrol acts to provide neuroprotection in recurrent stroke models by regulating AMPK and SIRT1 signaling, thereby reducing energy requirements during ischemia. Eur. J. Neurosci. 2013, 37, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Salil, G.; Nithya, R.; Nevin, K.G.; Rajamohan, T. Dietary coconut kernel protein beneficially modulates NFkappaB and RAGE expression in streptozotocin induced diabetes in rats. J. Food Sci. Technol. 2014, 51, 2141–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, D.G. Review: HbA1c has low accuracy for prediabetes; lifestyle programs and metformin reduce progression to T2DM. Ann. Int. Med. 2017, 166, Jc41. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Chang, Z.; Park, S.C.; Lim, N.R.; Kim, N.W. Antioxidanat activity and lrritation response of Lespedeza bicolor. J. Toxicol. 2005, 21, 115–119. [Google Scholar]
- Dong, X.G.; An, Z.M.; Guo, Y.; Zhou, J.L.; Qin, T. Effect of triptolide on expression of oxidative carbonyl protein in renal cortex of rats with diabetic nephropathy. J. Huazhong Univ. Sci. Technol. Med. Sci. Hua Zhong Ke Ji Da Xue Xue Bao. YI Xue Ying De Wen Ban Huazhong Keji Daxue Xuebao. Yixue Yingdewen Ban. 2017, 37, 25–29, (Chinese). [Google Scholar] [CrossRef] [PubMed]
- Pieper, C.M.; Roza, A.M.; Henderson, J.D., Jr.; Zhu, Y.R.; Lai, C.S. Spatial distribution and temporal onset of NF-kB activation and inducible nitric oxide synthase within pancreatic islets in the pre-diabetic stage of genetic, diabetic-prone BB rats: Attenuation by drug intervention decreases inflammatory cell infiltration and incidence of diabetes. Inflammation 2004, 53, 22–30. [Google Scholar]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Med. Inflamm. 2007, 2007, 45673. [Google Scholar]
- Zhang, X.; Li, H.; Feng, H.; Xiong, H.; Zhang, L.; Song, Y.; Yu, L.; Deng, X. Valnemulin downregulates nitric oxide, prostaglandin E2, and cytokine production via inhibition of NF-kappaB and MAPK activity. Int. Immunopharmacol. 2009, 9, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Skaba, D.; Czuba, Z.P.; Krol, W. Inhibition of inflammatory mediators by neobavaisoflavone in activated RAW264.7 macrophages. Molecules 2011, 16, 3701–3712. [Google Scholar] [CrossRef]
- Guo, Z.; Xu, H.Y.; Xu, L.; Wang, S.S.; Zhang, X.M. In vivo and in vitro immunomodulatory and anti-inflammatory effects of total flavonoids of astragalus. Afr. J. Tradit. Complement. Altern. Med. AJTCAM 2016, 13, 60–73. [Google Scholar] [CrossRef] [Green Version]
- Richter, K.; Kietzmann, T. Reactive oxygen species and fibrosis: Further evidence of a significant liaison. Cell Tissue Res. 2016, 365, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Zhang, Y.; Liu, Y.; Yuan, Q.; Song, L.; Liu, M.; Liu, Z.; Yang, Y.; Li, J.; Li, D.; et al. Fibroblast growth factor 21 attenuates hepatic fibrogenesis through TGF-beta/smad2/3 and NF-kappaB signaling pathways. Toxicol. Appl. Pharmacol. 2016, 290, 43–53. [Google Scholar] [CrossRef]
- Fan, H.N.; Wang, H.J.; Yang-Dan, C.R.; Ren, L.; Wang, C.; Li, Y.F. Protective effects of hydrogen sulfide on oxidative stress and fibrosis in hepatic stellate cells. Mol. Med. 2013, 7, 247–253. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | NC | DMC | LL | HL |
|---|---|---|---|---|
| Body weight (g) | ||||
| Before treatment | 26.58 ± 1.35 a | 32.04 ± 3.29 b | 30.94 ± 2.40 b | 32.51 ± 4.13 b |
| After treatment | 30.42 ± 1.83 a | 43.93 ± 4.97 b | 39.02 ± 5.15 b | 40.34 ± 6.35 b |
| Gain | 3.85 ± 0.33 a | 8.89 ± 0.70 b | 8.09 ± 1.34 b | 7.82 ± 1.72 b |
| Food intake (g/day) | 2.40 ± 0.731 a | 3.36 ± 0.89 a,b | 3.56 ± 1.70 b | 4.16 ± 1.27 b |
| Fasting blood glucose (FBG) (mg/dL) | 122.2 ± 16.78 a | 173.8 ± 31.97 b | 135.6 ± 13.28 a,b | 170.8 ± 28.31 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Lee, H.; Kim, S.Y.; Lim, Y. Effects of Lespedeza Bicolor Extract on Regulation of AMPK Associated Hepatic Lipid Metabolism in Type 2 Diabetic Mice. Antioxidants 2019, 8, 599. https://doi.org/10.3390/antiox8120599
Kim Y, Lee H, Kim SY, Lim Y. Effects of Lespedeza Bicolor Extract on Regulation of AMPK Associated Hepatic Lipid Metabolism in Type 2 Diabetic Mice. Antioxidants. 2019; 8(12):599. https://doi.org/10.3390/antiox8120599
Chicago/Turabian StyleKim, Younmi, Heaji Lee, Sun Yeou Kim, and Yunsook Lim. 2019. "Effects of Lespedeza Bicolor Extract on Regulation of AMPK Associated Hepatic Lipid Metabolism in Type 2 Diabetic Mice" Antioxidants 8, no. 12: 599. https://doi.org/10.3390/antiox8120599
APA StyleKim, Y., Lee, H., Kim, S. Y., & Lim, Y. (2019). Effects of Lespedeza Bicolor Extract on Regulation of AMPK Associated Hepatic Lipid Metabolism in Type 2 Diabetic Mice. Antioxidants, 8(12), 599. https://doi.org/10.3390/antiox8120599