Antimicrobial and Antibiofilm Activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. Cladode Polyphenolic Extracts

 ,
, 
 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extraction
2.3. Identification and Quantification of Phenolic Compounds
2.4. In Vitro Antioxidant Activity
2.5. Ex Vivo Antioxidant Activity
2.5.1. Cellular Antioxidant Activity (CAA-RBC) Assay in Red Blood Cells
2.5.2. Erythrocytes Oxidative Hemolysis
2.6. Antimicrobial Activity
2.6.1. Bacterial Media
2.6.2. Bacterial Strains and Growth Conditions
2.6.3. Inhibition Assay–MINIMUM Inhibitory Concentration (MIC)
2.7. Biofilm Production and Inhibition Assay
2.8. Statistical Analysis
3. Results and Discussion
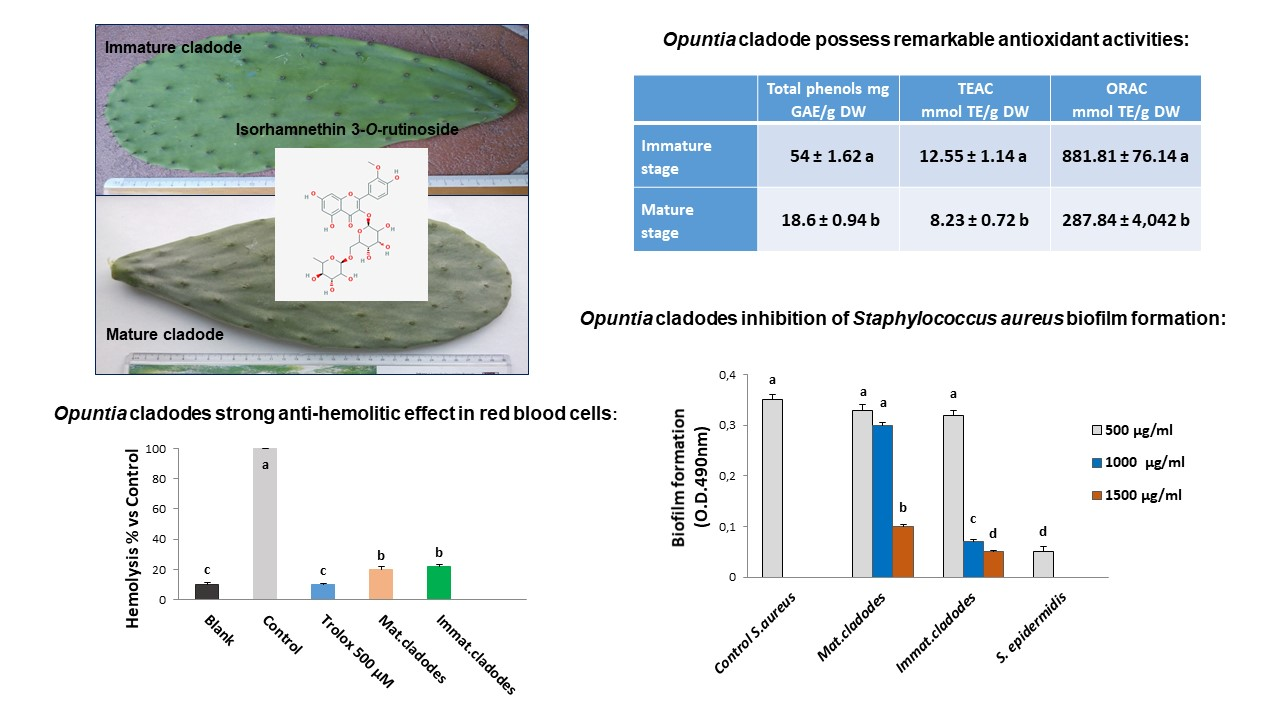
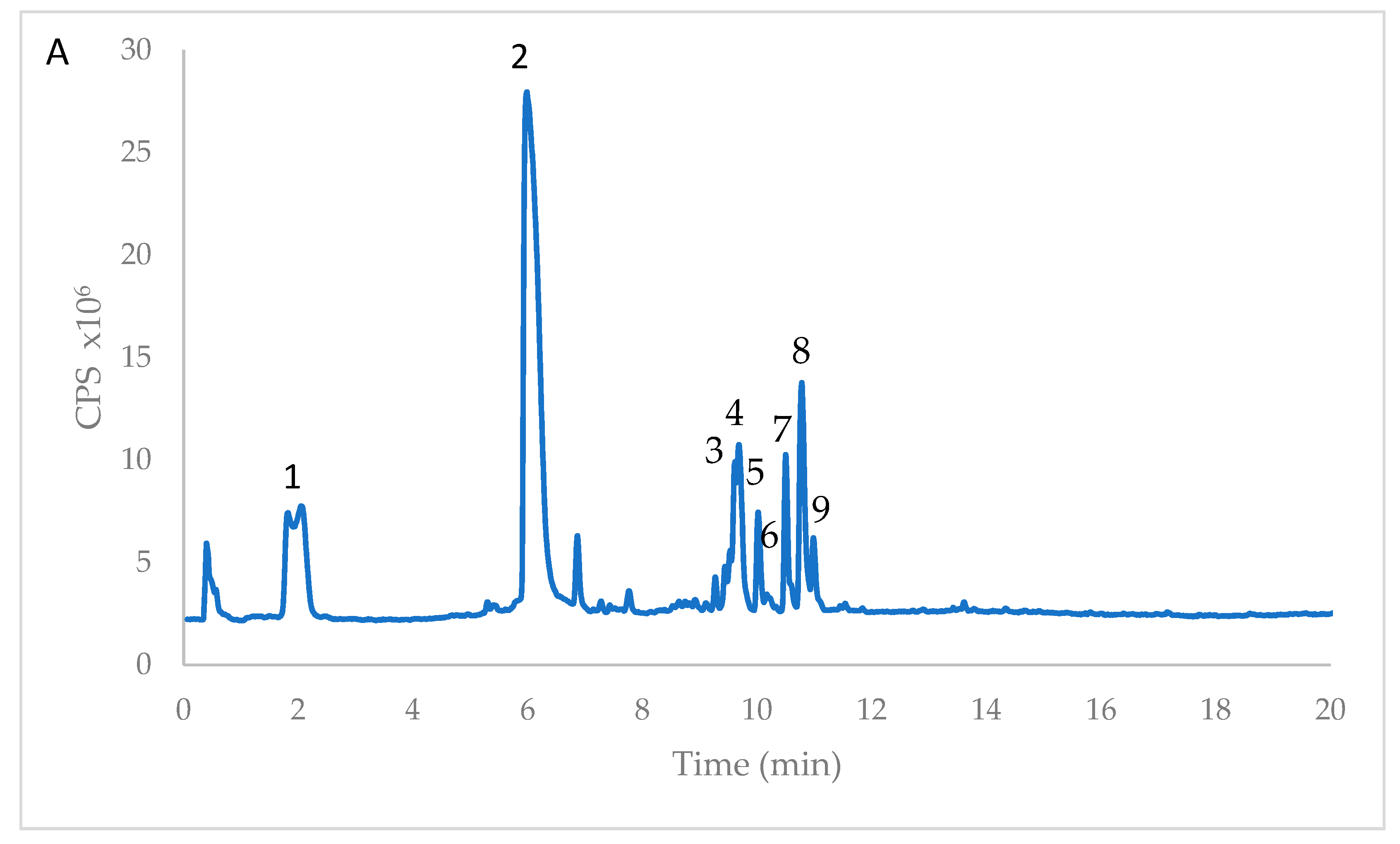
3.1. Polyphenolic Composition of Opuntia ficus-indica Cladode Extracts
3.2. In Vitro Antioxidant Activity
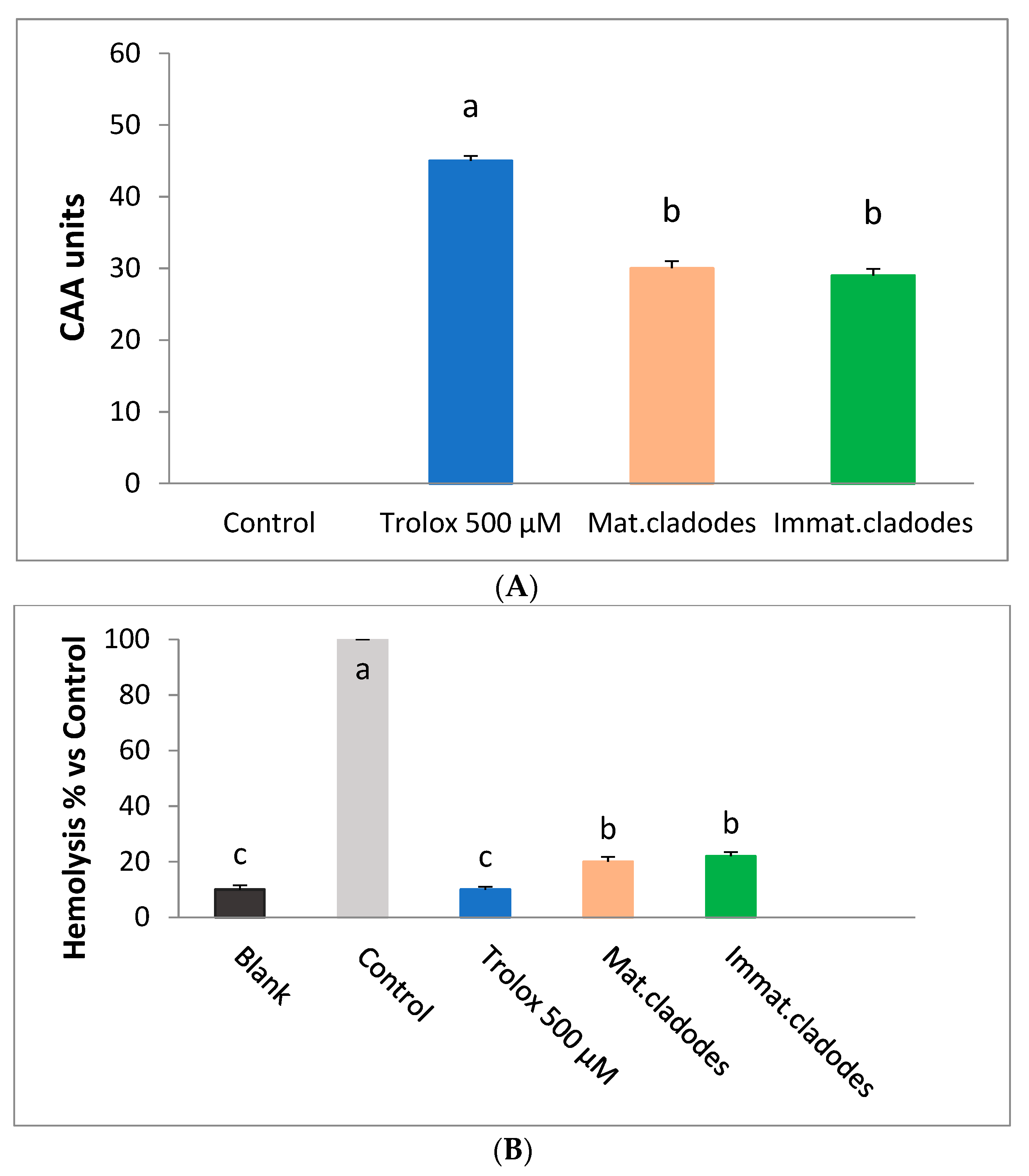
3.3. Ex Vivo Antioxidant Activity
3.3.1. Cellular Antioxidant Activity Assay in Red Blood Cells (CAA-RBC)
3.3.2. Haemolysis Assay
3.4. Antimicrobial Activity
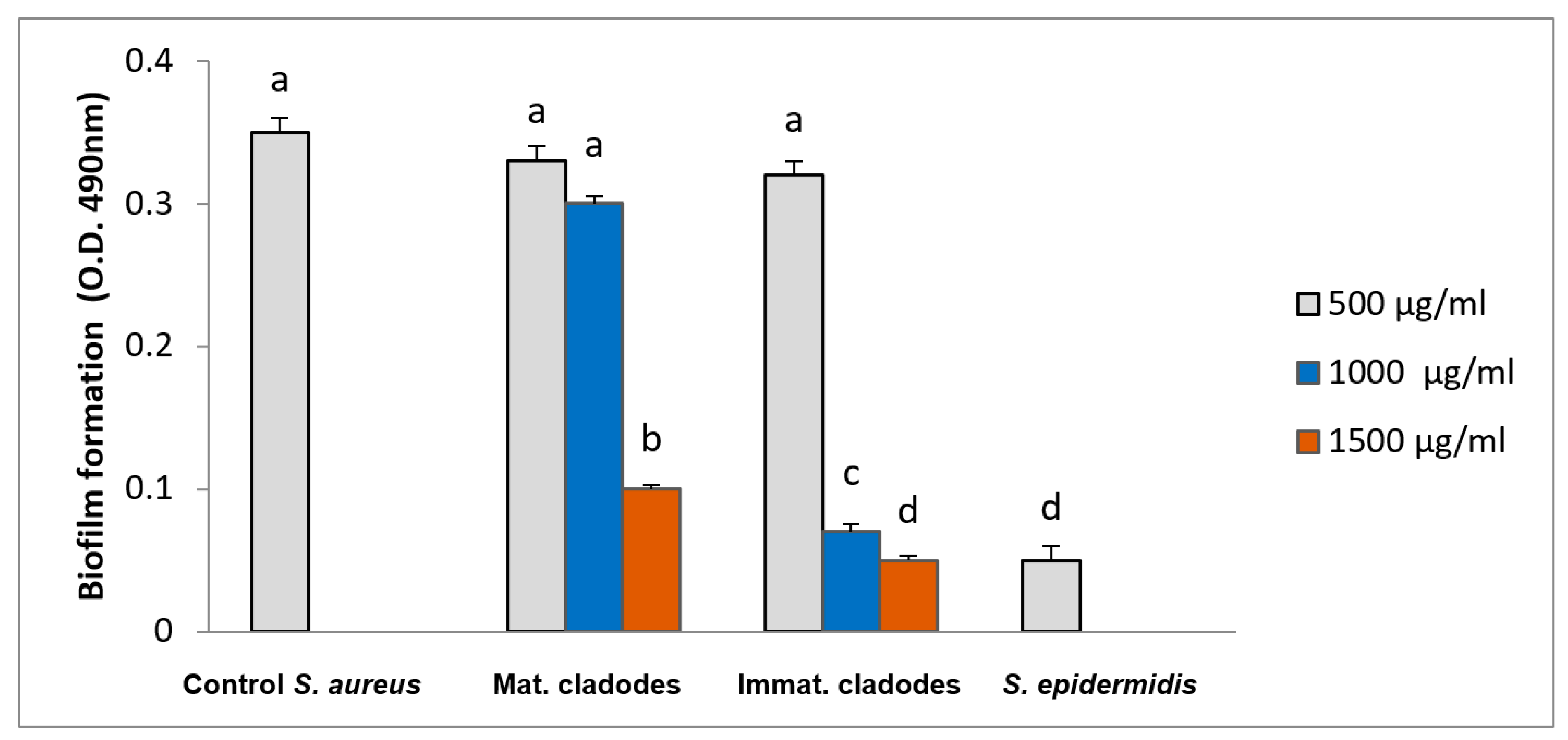
3.5. Biofilm Inhibition
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gyawali, R.; Hayek, S.A.; Ibrahim, S.A. Plant extracts as antimicrobials. In Handbook of Natural Antimicrobials for Food Safety and Quality; Taylor, T.M., Ed.; Elsevier: Cambridge, UK, 2015; pp. 31–47. [Google Scholar]
- Di Ciccio, P.; Vergara, A.; Festino, A.R.; Paludi, D.; Zanardi, E.; Ghidini, S.; Ianieri, A. Biofilm formation by Staphilococcus aureus on food contact surfaces: Relationship with temperature and cell surface hydrophobicity. Food Control 2015, 50, 930–936. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Satpathy, S.; Sen, S.K.; Pattanaik, S.; Raut, S. Review on bacterial biofilm: An universal cause of contamination. Biocatal. Agric. Biotechnol. 2016, 7, 56–66. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Di Ciccio, P.; Ianieri, A.; Nychas, G.E. Methicillin-resistant food-related Staphylococcus aureus: A review of current knowledge and biofilm formation for future studies and applications. Res. Microbiol. 2017, 168, 1–15. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Delgado Adámez, J.; Gamero Samino, E.; Valdés Sánchez, E.; González-Gómez, D. In vitro estimation of the antibacterial activity and antioxidant capacity of aqueous extracts from grape seeds (Vitis vinifera L.). Food Control 2012, 24, 136–141. [Google Scholar] [CrossRef]
- Chibane, L.B.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric. 2018, 99, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Nostro, A.; Guerrini, A.; Marino, A.; Tacchini, M.; Di Giulio, M.; Grandini, A.; Saraçoğlu, H.T. In vitro activity of plant extracts against biofilm-producing food-related bacteria. Int. J. Food Microbiol. 2016, 238, 33–39. [Google Scholar] [CrossRef]
- Brambilla, L.Z.S.; Endo, E.H.; Cortez, D.A.G.; Dias Filho, B.P. Anti-biofilm activity against Staphylococcus aureus MRSA and MSSA of neolignans and extract of Piper regnellii. Rev. Bras. Farmacognosia 2017, 27, 112–117. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Schieber, A.; Carle, R. Phytochemical and nutritional significance of cactus pear. Eur. Food Res. Technol. 2001, 212, 396–407. [Google Scholar] [CrossRef]
- Hernández-Urbiola, M.I.; Pérez-Torrero, E.; Rodríguez-Garcia, M.E. Chemical analysis of nutritional content of prickly pads (Opuntia ficus-indica L.) at varied ages in an organic harvest. Int. J. Environ. Res. Public Health 2011, 8, 1287–1295. [Google Scholar]
- Sabella, E.; Luvisi, A.; Aprile, A.; Negro, C.; Vergine, M.; Nicolì, F.; Miceli, A.; De Bellis, L. Xylella fastidiosa induces differential expression of lignification related-genes and lignin accumulation in tolerant olive trees cv. Leccino. J. Plant Physiol. 2018, 220, 60–68. [Google Scholar] [CrossRef]
- Albano, C.; Negro, C.; Tommasi, N.; Gerardi, C.; Mita, G.; Miceli, A.; De Bellis, L.; Blando, F. Betalains, phenols and antioxidant capacity in cactus pear [Opuntia ficus-indica (L.) Mill.] fruits from Apulia (South Italy) genotypes. Antioxidants 2015, 4, 269–280. [Google Scholar] [CrossRef]
- Frassinetti, S.; Gabriele, M.; Caltavuturo, L.; Longo, V.; Pucci, L. Antimutagenic and antioxidant activity of a selected lectin-free common bean (Phaseolus vulgaris L.) in two cell-based models. Plant Foods Hum. Nutr. 2015, 70, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47. [Google Scholar] [CrossRef]
- Ginestra, G.; Parker, M.L.; Bennet, R.N.; Robertson, J.; Mandalari, G.; Narbad, A.; Lo Curto, R.B.; Bisignano, G.; Faulds, C.B.; Waldron, K.W. Anatomical, Chemical, and Biochemical Characterization of Cladodes from Prickly Pear [Opuntia ficus-indica (L.) Mill.]. J. Agric. Food Chem. 2009, 57, 10323–10330. [Google Scholar] [CrossRef]
- Santos-Zea, L.; Guitiérrez-Uribe, J.A.; Serna-Saldivar, O. Comparative analysis of total phenols, antioxidant activity, and flavonol glycoside profile of cladode flours from different varieties of Opuntia spp. J. Agric. Food Chem. 2011, 59, 7054–7061. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, F.; Silipo, A.; Molinaro, A.; Parrilli, M.; Schiraldi, C.; D’Agostino, A.; Izzo, E.; Rizza, L.; Bonina, A.; Bonina, F.; Lanzetta, R. The polysaccharide and low molecular weight components of Opuntia ficus indica cladodes: Structure and skin repairing properties. Carbohydr. Polym. 2017, 157, 128–136. [Google Scholar] [CrossRef]
- Guevara-Figueroa, T.; Jiménez-Islas, H.; Reyes-Escogido, M.L.; Mortensen, A.G.; Laursen, B.B.; Lin, L.-W.; Barba de la Rosa, A.P. Proximate composition, phenolic acids, and flavonoids characterization of commercial and wild nopal (Opuntia spp.). J. Food Compos. Anal. 2010, 23, 525–532. [Google Scholar] [CrossRef]
- Ressaissi, A.; Attia, N.; Falé, P.L.; Pacheco, R.; Teixeira, V.H.; Machuqueiro, M.; Borges, C.; Serralheiro, M.L.M. Aqueous extracts from nopal (Opuntia ficus-indica): Antiacetylcholinesterase and antioxidant activity from phenolic bioactive compounds. Int. J. Green Herb. Chem. 2016, 5, 337–348. [Google Scholar]
- Medina-Torres, L.; Vernon-Carter, E.J.; Gallegos-Infante, J.A.; Rocha-Guzman, N.E.; Herrera-Valencia, E.E.; Calderas, F.; Jiménez-Alvarado, R. Study of the antioxidant properties of extracts obtained from nopal cactus (Opuntia ficus-indica L.) cladodes after convective drying. J. Sci. Food Agric. 2011, 91, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Ressaissi, A.; Attia, N.; Falé, P.L.; Pacheco, R.; Victor, B.L.; Machuqueiro, M.; Borges, C.; Serralheiro, M.L.M. Isorhamnetin derivatives and piscidic acid for hypercholesterolemia: Cholesterol permeability, HMG-CoA reductase inhibition, and docking studies. Arch. Pharmacal Res. 2017, 40, 1278–1286. [Google Scholar] [CrossRef]
- Petruk, G.; Di Lorenzo, F.; Imbimbo, P.; Silipo, A.; Bonina, A.; Rizza, L.; Piccoli, R.; Monti, D.M.; Lanzetta, R. Protective effect of Opuntia ficus-indica L. cladodes against UVA-induced oxidative stress in normal human keratinocytes. Bioorg. Med. Chem. Lett. 2017, 27, 5485–5489. [Google Scholar] [CrossRef]
- Rocchetti, G.; Pellizzoni, M.; Montesano, D.; Lucini, L. Italian Opuntia ficus-indica Cladodes as Rich Source of Bioactive Compounds with Health-Promoting Properties. Foods 2018, 7, 24. [Google Scholar] [CrossRef]
- Pérez Méndez, L.; Tejera Flores, F.; Darias Martín, J.; Rodríguez Rodríguez, E.M.; Díaz Romero, C. Physicochemical characterization of cactus pads from Opuntia dillenii and Opuntia ficus indica. Food Chem. 2015, 188, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Corral-Aguayo, R.D.; Yahia, E.M.; Carrillo-Lopez, A.; González-Aguilar, G. Correlation between some nutritional components and the total antioxidant capacity measured with six different assays in eight horticultural crops. J. Agric. Food Chem. 2008, 56, 10498–10504. [Google Scholar] [CrossRef]
- Avila-Nava, A.; Calderón-Oliver, M.; Medina-Campos, O.N.; Zou, T.; Gu, L.; Torres, N.; Pedraza-Chaverri, J. Extract of cactus (Opuntia ficus indica) cladodes scavenge reactive oxygen species in vitro and enhances plasma antioxidant capacity in humans. J. Funct. Foods 2014, 10, 13–24. [Google Scholar] [CrossRef]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. J. Agric. Food Chem. 2004, 52, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, C.G.; Garbois, G.D.; Amaral, L.M.; Diniz, C.C.; Le Hyaric, M. Relationship between structure and antibacterial activity of lipophilic N-acyldiamines. Biomed. Pharmacother. 2010, 64, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Gabriele, M.; Frassinetti, S.; Caltavuturo, L.; Montero, L.; Dinelli, G.; Longo, V.; Pucci, L. Citrus bergamia powder: Antioxidant, antimicrobial and anti-inflammatory properties. J. Funct. Foods 2017, 31, 255–265. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef]
- El-Mostafa, K.; El Karrassi, Y.; Badreddine, A.; Andreoletti, P.; Vamecq, J.; El Kebbai, M.S.; Latruffe, N.; Lizard, G.; Nasser, B.; Cherkaoui-Malki, M. Nopal cactus (Opuntia ficus-indica) as a source of bioactive compounds for nutrition, health and disease. Molecules 2014, 19, 14879–14901. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Rivas Morales, C.; Castillo, S.; Leos-Rivas, C.; García-Becerra, L.; Ortiz Martínez, D.M. Antibacterial and Antibiofilm Activity of Methanolic Plant Extracts against Nosocomial Microorganisms. Evid. Based Complement. Altern. Med. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Koley, H. Antibacterial activity of polyphenolic fraction of kombucha against enteric bacterial pathogens. Curr. Microbiol. 2016, 73, 885–896. [Google Scholar] [CrossRef]
- Bazargani, M.; Rohloff, J. Antibiofilm activity of essential oils and plant extracts against Staphylococcus aureus and Escherichia coli biofilms. Food Control 2016, 61, 156–164. [Google Scholar] [CrossRef]
- Slobodníková, L.; Fialová, S.; Rendeková, K.; Kováč, J.; & Mučaji, P. Antibiofilm activity of plant polyphenols. Molecules 2016, 21, 1717. [Google Scholar] [CrossRef]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Khan, M.S.; Khan, R.A.; Khan, J.M. Rutin inhibits mono and multi-species biofilm formation by foodborne drug resistant Escherichia coli and Staphylococcus aureus. Food Control 2017, 79, 325–332. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT | Formula [M − H]− | m/z Exp | m/z Calc | Δ (ppm) | Compound Name | Reference |
|---|---|---|---|---|---|---|---|
| 1 | 1.988 | C11H11O7 | 255.0512 | 255.0510 | −0.67 | Piscidic acid | Ginestra et al., 2009 [18] |
| 2 | 6.170 | C11H11O6 | 239.0567 | 239.0561 | −2.52 | Eucomic acid | Ginestra et al., 2009 [18] |
| 3 | 9.628 | C34H41O20 | 769.6770 | 769.6773 | −0.39 | Isorhamnetin rhamnosyl rutinoside | Santos-Zea et al., 2011 [19] |
| 4 | 9.695 | C33H39O20 | 755.6512 | 755.6508 | 0.53 | Isorhamnetin glucosyl rhamnosyl pentoside | Santos-Zea et al., 2011 [19] |
| 5 | 10.022 | C27H29O16 | 609.1459 | 609.1461 | 0.34 | Rutin | Ginestra et al., 2009 [18] |
| 6 | 10.034 | C27H29O16 | 609,1470 | 609,1461 | −1.43 | Isorhamnetin sambubioside | Santos-Zea et al., 2011 [19] |
| 7 | 10.503 | C27H29O15 | 593.1517 | 593.1512 | −0.88 | Kampherol glucosyl rhamnoside | Santos-Zea et al., 2011 [19] |
| 8 | 10.785 | C28H31O16 | 623.1645 | 623.1618 | −4.37 | Narcissin (isorhamnetin rutinoside) | Santos-Zea et al., 2011 [19] |
| 9 | 10.987 | C22H21O12 | 477.1045 | 477.1038 | −1.42 | Isorhamnetin glucoside | Ginestra et al., 2009 [18] |
| Immature Stage | mg/g DW | mg/100g FW |
| (1) Piscidic acid | 1.984 ± 0.019 c | 14.681 ± 0.146 c |
| (2) Eucomic acid | 13.506 ± 0.143 a | 99.944 ± 1.062 a |
| (4) Isorhamnetin rhamnosyl rutinoside | 0.411 ± 0.003 d | 3.041 ± 0.027 e |
| (5) Isorhamnetin glucosyl rhamnosyl pentoside | 0.296 ± 0.004 e | 2.192 ± 0.033 d |
| (6) Rutin | 2.030 ± 0.023 c | 15.022 ± 0.171 c |
| (8) Narcissin (isorhamnetin rutinoside) | 3.188 ± 0.042 b | 23.598 ± 0.302 b |
| (9) Isorhamnetin glucoside | 0.465 ± 0.021 d | 3.444 ± 0.161 e |
| Mature Stage | mg/g DW | mg/100g FW |
| (1) Piscidic acid | 3.281 ± 0.032 b | 28.257 ± 0.279 b |
| (2) Eucomic acid | 1.616 ± 0.02 f | 13.917 ± 0.174 f |
| (4) Isorhamnetin rhamnosyl rutinoside | 0.187 ± 0.012 e | 1.618 ± 0.104 d |
| (5) Isorhamnetin glucosyl rhamnosyl pentoside | 0.266 ± 0.007 e | 2.293 ± 0.067 d |
| (6) Rutin | 0.752 ± 0.004 g | 6.474 ± 0.034 g |
| (8) Narcissin (isorhamnetin rutinoside) | 1.160 ± 0.029 f | 9.985 ± 0.25 h |
| (9) Isorhamnetin glucoside | ND | ND |
| Total Phenols | TEAC | ORAC | ||||
|---|---|---|---|---|---|---|
| mg GAE/g DW | mg GAE/100 g FW | μmol TE/g DW | μmol TE/100 g FW | mmol TE/g DW | mmol TE/100 g FW | |
| Immature stage | 54.02 ± 1.62 a | 399.61 ± 11.98 a | 12.55 ± 1.14 a | 92.87 ± 8.51 a | 0.88 ± 0.08 a | 6.52 ± 0.56 a |
| Mature stage | 18.63 ± 0.94 b | 160.14 ± 8.09 b | 8.23 ± 0.72 b | 70.85 ± 6.22 b | 0.29 ± 0.04 b | 2.47 ± 0.36 b |
| Strain | MIC (m) µg/mL | MIC (imm) µg/mL | |
|---|---|---|---|
| Escherichia coli | ATCC 25922 | 2000 | 1500 |
| Salmonella typhimurium | ATCC 14028 | 2000 | 1500 |
| Enterobacter aerogenes | ATCC 13048 | 2000 | 1500 |
| Enterococcus faecalis | ATCC 29212 | 1500 | 1000 |
| Staphylococcus aureus | ATCC 25923 | 1500 | 1000 |
| Staphylococcus aureus | ATCC 35556 | 1000 | 700 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blando, F.; Russo, R.; Negro, C.; De Bellis, L.; Frassinetti, S. Antimicrobial and Antibiofilm Activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. Cladode Polyphenolic Extracts. Antioxidants 2019, 8, 117. https://doi.org/10.3390/antiox8050117
Blando F, Russo R, Negro C, De Bellis L, Frassinetti S. Antimicrobial and Antibiofilm Activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. Cladode Polyphenolic Extracts. Antioxidants. 2019; 8(5):117. https://doi.org/10.3390/antiox8050117
Chicago/Turabian StyleBlando, Federica, Rossella Russo, Carmine Negro, Luigi De Bellis, and Stefania Frassinetti. 2019. "Antimicrobial and Antibiofilm Activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. Cladode Polyphenolic Extracts" Antioxidants 8, no. 5: 117. https://doi.org/10.3390/antiox8050117
APA StyleBlando, F., Russo, R., Negro, C., De Bellis, L., & Frassinetti, S. (2019). Antimicrobial and Antibiofilm Activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. Cladode Polyphenolic Extracts. Antioxidants, 8(5), 117. https://doi.org/10.3390/antiox8050117