Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress


 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Diet and Experimental Design
2.3. Slaughter and Meat Quality Measures
2.4. High Performance Liqiud Chromatography Analysis of Betaine and Derivatives
2.5. Plasma Thyroid Hormone Analysis
2.6. Statistical Analyses
3. Results
3.1. Growth Performance
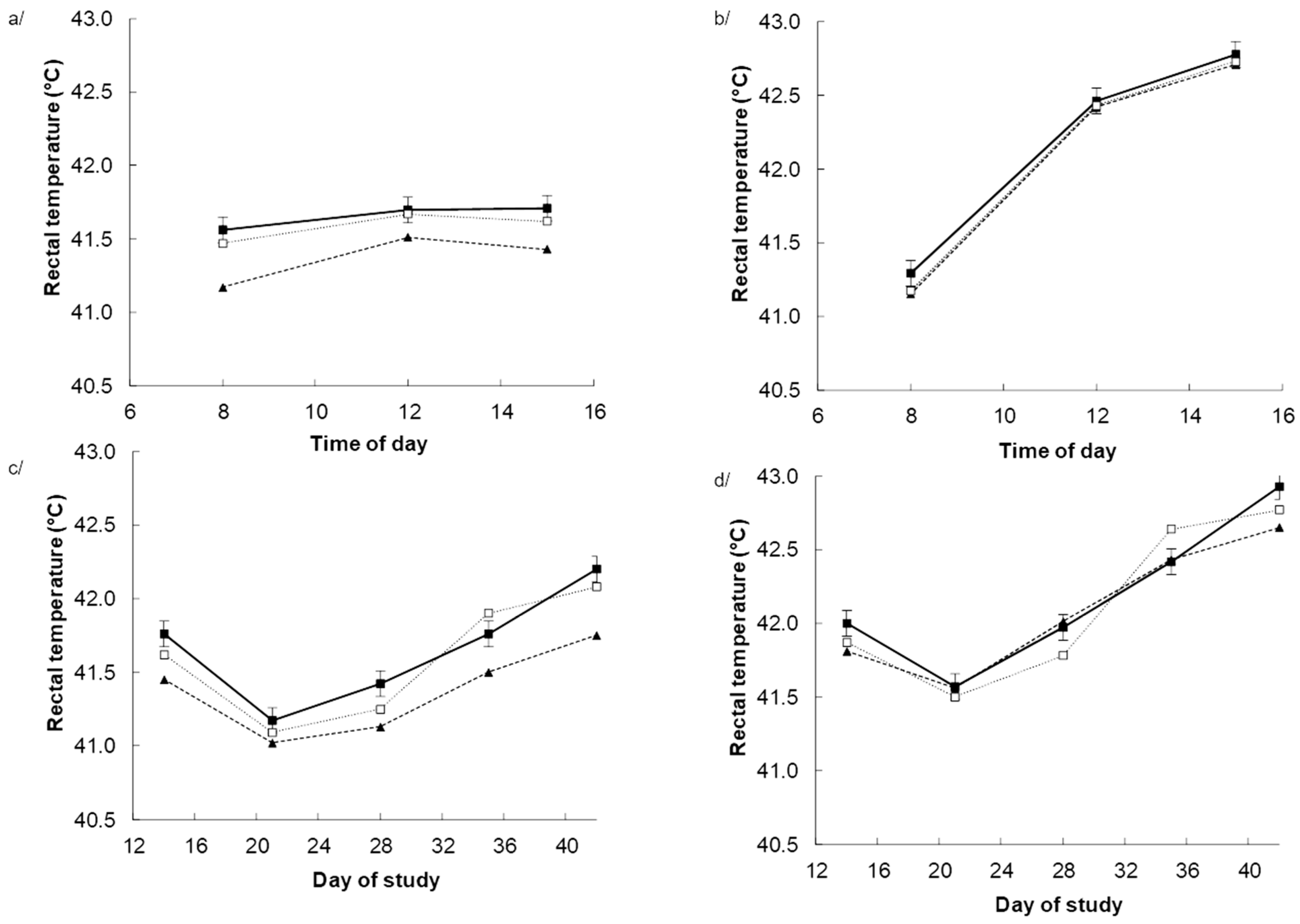
3.2. Physiological Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henry, B.; Charmley, E.; Eckard, R.; Gaughan, J.B.; Hegarty, R. Livestock production in a changing climate: Adaptation and mitigation research in Australia. Crop Pasture Sci. 2012, 63, 191–202. [Google Scholar] [CrossRef]
- Renaudeau, D.; Collin, A.; Yahav, S.; De Basilio, V.; Gourdine, J.; Collier, R.J. Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal 2012, 6, 707–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraert, P.; Padilha, J.; Guillaumin, S. Metabolic and endocrine changes induced by chronic heatexposure in broiler chickens: Growth performance, body composition and energy retention. Br. J. Nutr. 1996, 75, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Temim, S.; Chagneau, A.-M.; Peresson, R.; Tesseraud, S. Chronic heat exposure alters protein turnover of three different skeletal muscles in finishing broiler chickens fed 20 or 25% protein diets. J. Nutr. 2000, 130, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Ezzine, S.B.-O.; Everaert, N.; Metayer-Coustard, S.; Rideau, N.; Berri, C.; Joubert, R.; Temim, S.; Collin, A.; Tesseraud, S. Effects of heat exposure on Akt/S6K1 signaling and expression of genes related to protein and energy metabolism in chicken (Gallus gallus) pectoralis major muscle. Comp. Biochem. Physiol. B 2010, 157, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Akşit, M.; Yalcin, S.; Özkan, S.; Metin, K.; Özdemir, D. Effects of temperature during rearing and crating on stress parameters and meat quality of broilers. Poult. Sci. 2006, 85, 1867–1874. [Google Scholar] [PubMed]
- Love, J.D.; Pearson, A.M. Lipid oxidation in meat and meat products—A review. J. Am. Oil Chem. Soc. 1971, 48, 547–549. [Google Scholar] [CrossRef]
- Rajagopal, K.; Oommen, G.T. Myofibril fragmentation index as an immediate postmortem predictor of Buffalo meat tenderness. J. Food Process. Preserv. 2015, 39, 1166–1171. [Google Scholar] [CrossRef]
- Huang, C.; Jiao, H.; Song, Z.; Zhao, J.; Wang, X.; Lin, H. Heat stress impairs mitochondria functions and induces oxidative injury in broiler chickens. J. Anim. Sci. 2015, 93, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- DiGiacomo, K.; Leury, B.J.; Dunshea, F.R. Potential nutritional strategies for the amelioration or prevention of high rigor temperature in cattle–A review. Anim. Prod. Sci. 2014, 54, 430–443. [Google Scholar] [CrossRef]
- Cottrell, J.J.; Ponnampalam, E.N.; Dunshea, F.R.; Warner, R.D. Effects of infusing nitric oxide donors and inhibitors on plasma metabolites, muscle lactate production and meat quality in lambs fed a high quality roughage-based diet. Meat Sci. 2015, 105, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A.S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alirezaei, M.; Gheisari, H.R.; Ranjbar, V.R.; Hajibemani, A. Betaine: A promising antioxidant agent for enhancement of broiler meat quality. Br. Poult. Sci. 2012, 53, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Alirezaei, M.; Khoshdel, Z.; Dezfoulian, O.; Rashidipour, M.; Taghadosi, V. Beneficial antioxidant properties of betaine against oxidative stress mediated by levodopa/benserazide in the brain of rats. J. Physiol. Sci. 2015, 65, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Ringuet, M.; Furness, J.; Dunshea, F.R. Betaine and Antioxidants Improve Growth Performance, Breast Muscle Development and Ameliorate Thermoregulatory Responses to Cyclic Heat Exposure in Broiler Chickens. Animals 2018, 8, 162. [Google Scholar] [CrossRef]
- Tsai, M.T.; Chen, C.Y.; Pan, Y.H.; Wang, S.H.; Mersmann, H.J.; Ding, S.T. Alleviation of carbon-tetrachloride-induced liver injury and fibrosis by betaine supplementation in chickens. Evid. Based Complement. Alternat. Med. 2015, 2015, 725379. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Leury, B.J.; Fahri, F.; DiGiacomo, K.; Hung, A.; Chauhan, S.; Clarke, I.J.; Collier, R.; Little, S.; Baumgard, L. Amelioration of thermal stress impacts in dairy cows. Anim. Prod. Sci. 2013, 53, 965–975. [Google Scholar] [CrossRef]
- Suster, D.; Leury, B.J.; King, R.H.; Mottram, M.; Dunshea, F.R. Interrelationships between porcine somatotropin (pST), betaine, and energy level on body composition and tissue distribution of finisher boars. Aust. J. Agric. Res. 2004, 55, 983–990. [Google Scholar] [CrossRef]
- Sun, H.; Yang, W.R.; Yang, Z.B.; Wang, Y.; Jiang, S.Z.; Zhang, G.G. Effects of betaine supplementation to methionine deficient diet on growth performance and carcass characteristics of broilers. Am. J. Anim. Vet. Sci. 2008, 3, 78–84. [Google Scholar] [CrossRef]
- Attia, Y.A.; Hassan, R.A.; Qota, E.M.A. Recovery from adverse effects of heat stress on slow-growing chicks in the tropics 1: Effect of ascorbic acid and different levels of betaine. Trop. Anim. Health Prod. 2009, 41, 807–818. [Google Scholar] [CrossRef]
- Oliveira, T.F.B.; Rivera, D.F.R.; Mesquita, F.R.; Braga, H.; Ramos, E.M.; Bertechini, A.G. Effect of different sources and levels of selenium on performance, meat quality, and tissue characteristics of broilers. J. Appl. Poult. Res. 2014, 23, 15–22. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y. Influence of dietary nano elemental selenium on growth performance, tissue selenium distribution, meat quality, and glutathione peroxidase activity in Guangxi Yellow chicken. Poult. Sci. 2011, 90, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Perić, L.; Milošević, N.; Žikić, D.; Kanački, Z.; Džinić, N.; Nollet, L.; Spring, P. Effect of selenium sources on performance and meat characteristics of broiler chickens. J. Appl. Poult. Res. 2009, 18, 403–409. [Google Scholar] [CrossRef]
- Choct, M.; Naylor, A.J.; Reinke, N. Selenium supplementation affects broiler growth performance, meat yield and feather coverage. Br. Poult. Sci. 2004, 45, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Lin, H.; Wang, X.J.; Song, Z.G.; Jiao, H.C. Vitamin E supplementation alleviates the oxidative stress induced by dexamethasone treatment and improves meat quality in broiler chickens. Poult. Sci. 2010, 89, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Kucuk, O. Effects of vitamin C and vitamin E on performance, digestion of nutrients and carcass characteristics of Japanese quails reared under chronic heat stress (34C). J. Anim. Physiol. Anim. Nutr. (Berl.) 2001, 85, 335–341. [Google Scholar] [CrossRef]
- Hashizawa, Y.; Kubota, M.; Kadowaki, M.; Fujimura, S. Effect of dietary vitamin E on broiler meat qualities, color, water-holding capacity and shear force value, under heat stress conditions. Anim. Sci. J. 2013, 84, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Aviagen Broiler Breeders. Ross Broiler Management Manual; Aviagen: Midlothian, Scotland, UK, 2009; pp. 1–114. [Google Scholar]
- NRC. Nutrient Requirements of Poultry; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Culler, R.D.; Smith, G.C.; Cross, H.R. Relationship of myofibril fragmentation index to certain chemical, physical and sensory characteristics of bovine longissimus muscle. J. Food Sci. 1978, 43, 1177–1180. [Google Scholar] [CrossRef]
- Koohmaraie, M.; Seidemann, S.C.; Schollmeyer, J.E.; Dutson, T.R.; Crouse, J.D. Effect of post-mortem storage on Ca++-dependent proteases, their inhibitor and myofibril fragmentation. Meat Sci. 1987, 19, 187–196. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Laryea, M.D.; Steinhagen, F.; Pawliczek, S.; Wendel, U. Simple method for the routine determination of betaine and N, N-dimethylglycine in blood and urine. Clin. Chem. 1998, 44, 1937–1941. [Google Scholar] [PubMed]
- Dunshea, F.R.; Ostrowska, E.; Luxford, B.; Smits, R.J.; Campbell, R.G.; DSouza, D.N.; Mullan, B.P. Dietary conjugated linoleic acid can decrease backfat in pigs housed under commercial conditions. Asian-Australas. J. Anim. Sci. 2002, 15, 1011–1017. [Google Scholar] [CrossRef]
- He, S.; Zhao, S.; Dai, S.; Liu, D.; Bokhari, S.G. Effects of dietary betaine on growth performance, fat deposition and serum lipids in broilers subjected to chronic heat stress. Anim. Sci. J. 2015, 86, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Nofal, M.E.; Magda, A.G.; Mousa, S.M.M.; Doaa, M.M.; Bealsh, A.M.A. Effect of dietary betaine supplementation on productive, physiological and immunological performance and carcass characteristic of growing developed chicks uinder the condition of heat stress. Egypt. Poult. Sci. J. 2015, 35, 237–259. [Google Scholar]
- Mello, J.; Boiago, M.; Giampietro-Ganeco, A.; Berton, M.; Vieira, L.; Souza, R.; Ferrari, F.; Borba, H. Periods of heat stress during the growing affects negatively the performance and carcass yield of broilers. Arch. Zootec. 2015, 64, 339–345. [Google Scholar] [CrossRef]
- Kettunen, H.; Tiihonen, K.; Peuranen, S.; Saarinen, M.; Remus, J. Dietary betaine accumulates in the liver and intestinal tissue and stabilizes the intestinal epithelial structure in healthy and coccidia-infected broiler chicks. Comp. Biochem. Physiol. A 2001, 130, 759–769. [Google Scholar] [CrossRef]
- Moeckel, G.W.; Shadman, R.; Fogel, J.M.; Sadrzadeh, S.M. Organic osmolytes betaine, sorbitol and inositol are potent inhibitors of erythrocyte membrane ATPases. Life Sci. 2002, 71, 2413–2424. [Google Scholar] [CrossRef]
- Cronje, P. Heat stress in livestock—The role of the gut in its aetiology and a potential role for betaine in its alleviation. Rec. Adv. Anim. Nutr. Aust. 2005, 15, 107–122. [Google Scholar]
- Raghebian, M.; Sadeghi, A.A.; Aminafshar, M. Energy sources and levels influenced on performance parameters, thyroid hormones, and HSP70 gene expression of broiler chickens under heat stress. Trop. Anim. Health Prod. 2016, 48, 1697–1702. [Google Scholar] [CrossRef] [PubMed]
- Schwahn, B.C.; Hafner, D.; Hohlfeld, T.; Balkenhol, N.; Laryea, M.D.; Wendel, U. Pharmacokinetics of oral betaine in healthy subjects and patients with homocystinuria. Br. J. Clin. Pharmacol. 2003, 55, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eklund, M.; Bauer, E.; Wamatu, J.; Mosenthin, R. Potential nutritional and physiological functions of betaine in livestock. Nutr. Res. Rev. 2005, 18, 31–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, M.; Downing, J. The effects of water replacement by oral rehydration fluids with or without betaine supplementation on performance, acid-base balance, and water retention of heat-stressed broiler chickens. Poult. Sci. 2011, 90, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, H.; Li, H.; Lai, F.; Li, X.; Tang, Y.; Min, T.; Wu, H. Antioxidant mechanism of betaine without free radical scavenging ability. J. Agric. Food Chem. 2016, 64, 7921–7930. [Google Scholar] [CrossRef] [PubMed]
- Akhavan-Salamat, H.; Ghasemi, H.A. Alleviation of chronic heat stress in broilers by dietary supplementation of betaine and turmeric rhizome powder: Dynamics of performance, leukocyte profile, humoral immunity, and antioxidant status. Trop. Anim. Health Prod. 2016, 48, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Hassan, R.A.; Shehatta, M.H.; Abd-El-Hady, S.B. Growth, carcass quality and serum constituents of slow growing chicks as affected by betaine addition to diets containing 2. Different levels of methionine. Int. J. Poult. Sci. 2005, 4, 856–865. [Google Scholar]
- Remus, J. Betaine for increased breast meat yield in turkeys. World Poult. 2001, 17, 14–15. [Google Scholar]
- Swain, B.K.; Johri, T.S.; Majumdar, S. Effect of supplementation of vitamin E, selenium and their different combinations on the performance and immune response of broilers. Br. Poult. Sci. 2000, 41, 287–292. [Google Scholar] [CrossRef]
- Habibian, M.; Ghazi, S.; Moeini, M.M. Effects of dietary selenium and vitamin E on growth performance, meat yield, and selenium content and lipid oxidation of breast meat of broilers reared under heat stress. Biol. Trace Elem. Res. 2016, 169, 142–152. [Google Scholar] [CrossRef]
- Zduńczyk, Z.; Gruzauskas, R.; Semaskaite, A.; Juskiewicz, J.; Raceviciute-Stupeliene, A.; Wroblewska, M. Fatty acid profile of breast muscle of broiler chickens fed diets with different levels of selenium and vitamin E. Arch. Geflugelkd. 2011, 75, 264–267. [Google Scholar]
- Kuttappan, V.A.; Goodgame, S.D.; Bradley, C.D.; Mauromoustakos, A.; Hargis, B.M.; Waldroup, P.W.; Owens, C.M. Effect of different levels of dietary vitamin E (DL-α-tocopherol acetate) on the occurrence of various degrees of white striping on broiler breast fillets. Poult. Sci. 2012, 91, 3230–3235. [Google Scholar] [CrossRef]
- Borges, S.A.; Fischer da Silva, A.V.; Majorka, A.; Hooge, D.M.; Cummings, K.R. Physiological responses of broiler chickens to heat stress and dietary electrolyte balance (sodium plus potassium minus chloride, milliequivalents per kilogram). Poult. Sci. 2004, 83, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Ait-Boulahsen, A.; Garlich, J.D.; Edens, F.W. Potassium chloride improves the thermotolerance of chickens exposed to acute heat stress. Poult. Sci. 1995, 74, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.O.; Teeter, R.G. Potassium balance of the 5 to 8-week-old broiler exposed to constant heat or cycling high temperature stress and the effects of supplemental potassium chloride on body weight gain and feed efficiency. Poult. Sci. 1987, 66, 487–492. [Google Scholar] [CrossRef]
- Teeter, R.G.; Smith, M.O.; Owens, F.N.; Arp, S.C.; Sangiah, S.; Breazile, J.E. Chronic heat stress and respiratory alkalosis: Occurrence and treatment in broiler chicks. Poult. Sci. 1985, 64, 1060–1064. [Google Scholar] [CrossRef] [PubMed]
- Altan, Ö.; Altan, A.; Çabuk, M.; Bayraktar, H. Effects of heat stress on some blood parameters in broilers. Turk. J. Vet. Anim. Sci. 2000, 24, 145–148. [Google Scholar]
- Bodnár, D.; Ruzsnavszky, O.; Oláh, T.; Dienes, B.; Balatoni, I.; Ungvári, É.; Benkő, I.; Babka, B.; Prokisch, J.; Csernoch, L. Dietary selenium augments sarcoplasmic calcium release and mechanical performance in mice. Nutr. Metab. (Lond.) 2016, 13, 76. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Gautam, A.K.; Sinha, M.K. Histopathological alterations of selenium toxicity induced in broiler (Birds). Indian J. Anim. Res. 2018, 52, 599–604. [Google Scholar]
- Maina, J.N.; King, A.; Settle, G. An allometric study of pulmonary morphometric parameters in birds, with mammalian comparisons. Philos. Trans. R Soc. Lond. B Biol. Sci. 1989, 326, 1–57. [Google Scholar] [CrossRef]
- Zhou, W.T.; Fujita, M.; Yamamoto, S. Effects of ambient temperatures on blood viscosity and plasma protein concentration of broiler chickens (Gallus domesticus). J. Therm. Biol. 1999, 24, 105–112. [Google Scholar] [CrossRef]
- Zhou, W.T. Heat-induced the shift of body fluid in broiler chickens. In Proceedings of the 6th Asian Pacific Poultry Congress, Nagoya, Japan, 4–7 June 1998. [Google Scholar]
- Billett, H.H. Hemoglobin and hematocrit. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Butterworths: London, UK, 1990. [Google Scholar]
- Zhang, L.; Yue, H.Y.; Zhang, H.J.; Xu, L.; Wu, S.G.; Yan, H.J.; Gong, Y.S.; Qi, G.H. Transport stress in broilers: I. Blood metabolism, glycolytic potential, and meat quality. Poult. Sci. 2009, 88, 2033–2041. [Google Scholar]
- Tao, X.; Dong, H.; Zhang, Z.; Xin, H. Daily variation of thyroid hormones in broiler under high-temperature conditions. In Proceedings of the Livestock Environment VII, Beijing, China, 18–20 May 2005; pp. 18–20. [Google Scholar]
- Hosseini, E.; Cheraghi, J.; Taheri, S.S.; Taherpour, K.; Kaviani, K.Z.; Rezazadeh, L. Thyroid hormones investigation under heat stress in broilers administered with probiotic (BIO-SAF) and prebiotic (BIO-MOS). Eur. J. Exp. Biol. 2013, 3, 562–567. [Google Scholar]
- Ayo, J.O.; Egbuniwe, I.; Kawu, M.U.; Sinkalu, V.O. Ameliorative effects of betaine and ascorbic acid administration to broiler chickens during the hot-dry season in Zaria: A review. Afr. J. Biotechnol. 2014, 13, 2295–2306. [Google Scholar]
- Mack, L.A.; Felver-Gant, J.N.; Dennis, R.L.; Cheng, H.W. Genetic variations alter production and behavioral responses following heat stress in 2 strains of laying hens. Poult. Sci. 2013, 92, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Elnagar, S.A.; Scheideler, S.E.; Beck, M.M. Reproductive hormones, hepatic deiodinase messenger ribonucleic acid, and vasoactive intestinal polypeptide-immunoreactive cells in hypothalamus in the heat stress-induced or chemically induced hypothyroid laying hen. Poult. Sci. 2010, 89, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Star, L.; Decuypere, E.; Parmentier, H.K.; Kemp, B. Effect of single or combined climatic and hygienic stress in four layer lines: 2. Endocrine and oxidative stress responses. Poult. Sci. 2008, 87, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Bobek, S.; Niezgoda, J.; Pietras, M.; Kacińska, M.; Ewy, Z. The effect of acute cold and warm ambient temperatures on the thyroid hormone concentration in blood plasma, blood supply, and oxygen consumption in Japanese quail. Gen. Comp. Endocrinol. 1980, 40, 201–210. [Google Scholar] [CrossRef]
- Mitchell, M.A.; Carlisle, A.J. The effects of chronic exposure to elevated environmental temperature on intestinal morphology and nutrient absorption in the domestic fowl (Gallus domesticus). Comp. Bochem. Physiol. A 1992, 101, 137–142. [Google Scholar] [CrossRef]
- Priolo, A.; Micol, D.; Agabriel, J. Effects of grass feeding systems on ruminant meat colour and flavour. A review. Anim. Res. 2001, 50, 185–200. [Google Scholar] [CrossRef]
- Cramer, T.A.; Kim, H.W.; Chao, Y.; Wang, W.; Cheng, H.W.; Kim, Y.H.B. Effects of probiotic (Bacillus subtilis) supplementation on meat quality characteristics of breast muscle from broilers exposed to chronic heat stress. Poult. Sci. 2018, 97, 3358–3368. [Google Scholar] [CrossRef]
- Sandercock, D.A.; Hunter, R.R.; Nute, G.R.; Mitchell, M.A.; Hocking, P.M. Acute heat stress-induced alterations in blood acid-base status and skeletal muscle membrane integrity in broiler chickens at two ages: Implications for meat quality. Poult. Sci. 2001, 80, 418–425. [Google Scholar] [CrossRef]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Chronic heat stress impairs the quality of breast-muscle meat in broilers by affecting redox status and energy-substance metabolism. J. Agric. Food Chem. 2017, 65, 11251–11258. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.; Joo, S.T. Poultry meat quality in relation to muscle growth and muscle fiber characteristics. Korean J. Food Sci. Anim. Resour. 2017, 37, 873. [Google Scholar] [PubMed]
- Warner, R.D.; Ferguson, D.M.; Cottrell, J.J.; Knee, B. Acute stress induced by the preslaughter use of electric prodders causes tougher beef meat. Aust. J. Exp. Agric. 2007, 47, 782–788. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Warner, R.D.; Leury, B.J.; Gaughan, J.B.; Dunshea, F.R. Dietary betaine supplementation has energy-sparing effects in feedlot cattle during summer, particularly in those without access to shade. Anim. Prod. Sci. 2014, 54, 450–458. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Zhang, M.; Zheng, S.; Xie, P.; Ma, A. Effects of high temperature on multiple parameters of broilers in vitro and in vivo. Poult. Sci. 2008, 87, 2133–2139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Jia, G.Q.; Zuo, J.J.; Zhang, Y.; Lei, J.; Ren, L.; Feng, D.Y. Effects of constant and cyclic heat stress on muscle metabolism and meat quality of broiler breast fillet and thigh meat. Poult. Sci. 2012, 91, 2931–2937. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.M.; Chen, W.; Ruan, D.; Wang, S.; Xia, W.G.; Zheng, C.T. Impact of heat stress on meat, egg quality, immunity and fertility in poultry and nutritional factors that overcome these effects: A review. Int. J. Poult. Sci. 2016, 15, 81. [Google Scholar]
- Wang, R.H.; Liang, R.R.; Lin, H.; Zhu, L.X.; Zhang, Y.M.; Mao, Y.W.; Dong, P.C.; Niu, L.B.; Zhang, M.H.; Luo, X. Effect of acute heat stress and slaughter processing on poultry meat quality and postmortem carbohydrate metabolism. Poult. Sci. 2017, 96, 738–746. [Google Scholar] [CrossRef]
- Park, S.O.; Kim, W.K. Effects of betaine on biological functions in meat-type ducks exposed to heat stress. Poult. Sci. 2016, 96, 1212–1218. [Google Scholar]
- Zhu, Y.W.; Li, W.X.; Lu, L.; Zhang, L.Y.; Ji, C.; Lin, X.; Liu, H.C.; Odle, J.; Luo, X.G. Impact of maternal heat stress in conjunction with dietary zinc supplementation on hatchability, embryonic development, and growth performance in offspring broilers. Poult. Sci. 2017, 96, 2351–2359. [Google Scholar] [CrossRef]
- Gu, X.H.; Li, S.S.; Lin, H. Effects of hot environment and dietary protein level on growth performance and meat quality of broiler chickens. Asian-Australas. J. Anim. Sci. 2008, 21, 1616–1623. [Google Scholar] [CrossRef]
- McDonagh, M.; Herd, R.; Richardson, E.; Oddy, V.; Archer, J.; Arthur, P. Meat quality and the calpain system of feedlot steers following a single generation of divergent selection for residual feed intake. Aust. J. Exp. Agric. 2001, 41, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.C.; Geesink, G.; Alvarenga, T.I.R.C.; Polkinghorne, R.; Stark, J.; Lee, M.; Warner, R.D. Impact of high oxygen and vacuum retail ready packaging formats on lamb loin and topside eating quality. Meat Sci. 2017, 123, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Azad, K.M.A.; Kikusato, M.; Hoque, A.M.; Toyomizu, M. Effect of chronic heat stress on performance and oxidative damage in different strains of chickens. J. Poult. Sci 2010, 47, 333–337. [Google Scholar] [CrossRef]
- Mujahid, A.; Pumford, N.R.; Bottje, W.; Nakagawa, K.; Miyazawa, T.; Akiba, Y.; Toyomizu, M. Mitochondrial oxidative damage in chicken skeletal muscle induced by acute heat stress. J. Poult. Sci 2007, 44, 439–445. [Google Scholar] [CrossRef]
- Skrivan, M.; Dlouha, G.; Mašata, O.; Ševčíková, S. Effect of dietary selenium on lipid oxidation, selenium and vitamin E content in the meat of broiler chickens. Czech. J. Anim. Sci. 2008, 53, 306–311. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Temperature (T) | Thermoneutral | Heat Stress | s.e.d. 2 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON 1 | BET | BET + AOX | CON 1 | BET | BET + AOX | T | BET | Within BET | |
| Day 0–21 | ||||||||||
| ADG, g/d | 45.5 | 47.0 | 49.5 | 43.4 | 44.1 | 46.8 | 1.85 | 0.11 | 0.15 | 0.17 |
| ADFI, g/d | 70.8 | 64.3 | 67.3 | 59.7 | 67.9 | 66.3 | 4.35 | 0.50 | 0.88 | 0.87 |
| FCR, g/g | 1.56 | 1.37 | 1.36 | 1.38 | 1.55 | 1.42 | 0.098 | 0.70 | 0.46 | 0.49 |
| Day 21–42 | ||||||||||
| ADG, g/d | 93.3 | 98.9 | 98.0 | 83.3 | 85.2 | 89.3 | 3.09 | <0.001 | 0.10 | 0.61 |
| ADFI, g/d | 182 | 184 | 197 | 164 | 171 | 180 | 7.4 | 0.017 | 0.13 | 0.15 |
| FCR, g/g | 1.96 | 1.86 | 2.01 | 1.97 | 2.02 | 2.02 | 0.075 | 0.32 | 0.86 | 0.34 |
| Day 0–42 | ||||||||||
| ADG, g/d | 69.4 | 72.9 | 73.8 | 63.3 | 64.7 | 68.1 | 1.99 | <0.001 | 0.056 | 0.30 |
| ADFI, g/d | 126 | 124 | 132 | 112 | 120 | 123 | 4.8 | 0.033 | 0.21 | 0.24 |
| FCR, g/g | 1.82 | 1.70 | 1.79 | 1.76 | 1.85 | 1.81 | 0.055 | 0.35 | 0.82 | 0.69 |
| Final weight, g | 2953 | 3103 | 3138 | 2697 | 2755 | 2898 | 83.6 | <0.001 | 0.052 | 0.30 |
| Breast weight, g | 672 | 753 | 785 | 596 | 640 | 662 | 24.2 | <0.001 | 0.002 | 0.24 |
| Temperature (T) | Thermoneutral | Heat Stress | s.e.d. 2 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON 1 | BET | BET + AOX | CON 1 | BET | BET + AOX | T | BET | Within BET | |
| pH | 7.31 | 7.31 | 7.32 | 7.41 | 7.37 | 7.38 | 0.037 | 0.021 | 0.64 | 0.84 |
| pCO2, mm Hg | 54.1 | 51.9 | 54.1 | 40.5 | 42.7 | 41.4 | 4.13 | 0.002 | 0.96 | 0.92 |
| Total CO2, mM | 27.1 | 27.7 | 32.1 | 26.0 | 27.2 | 28.4 | 1.39 | 0.14 | 0.067 | 0.058 |
| pO2, mm Hg | 31.3 | 34.0 | 29.6 | 35.1 | 31.9 | 33.9 | 1.61 | 0.15 | 0.54 | 0.47 |
| O2 saturation, % | 49.2 | 58.8 | 56.8 | 62.0 | 64.0 | 67.1 | 4.88 | 0.029 | 0.17 | 0.91 |
| HCO3, mM | 27.3 | 26.5 | 30.4 | 24.7 | 25.9 | 26.8 | 1.14 | 0.024 | 0.17 | 0.050 |
| Anion gap, mM | 19.3 | 16.0 | 15.8 | 16.3 | 15.5 | 13.3 | 1.36 | 0.089 | 0.039 | 0.37 |
| Base excess, mM | −1.38 | −0.23 | 1.90 | −0.10 | 0.93 | 1.78 | 0.994 | 0.36 | 0.047 | 0.15 |
| Temperature (T) | Thermoneutral | Heat Stress | s.e.d. 2 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON 1 | BET | BET + AOX | CON 1 | BET | BET + AOX | T | BET | Within BET | |
| Haematocrit, % | 21.5 | 21.8 | 22.0 | 18.8 | 18.0 | 19.8 | 1.04 | 0.003 | 0.78 | 0.35 |
| Hgb4, g/dL | 6.93 | 7.38 | 7.45 | 6.38 | 6.20 | 6.83 | 0.312 | 0.006 | 0.26 | 0.28 |
| Potassium, mM | 6.38 | 5.80 | 5.80 | 5.73 | 5.83 | 5.30 | 0.239 | 0.071 | 0.092 | 0.29 |
| Sodium, mM | 150 | 149 | 150 | 149 | 150 | 149 | 1.3 | 0.82 | 0.96 | 0.93 |
| Chloride, mM | 112 | 112 | 110 | 114 | 115 | 114 | 1.3 | 0.014 | 0.82 | 0.26 |
| Calcium, mM | 1.48 | 1.46 | 1.49 | 1.36 | 1.43 | 1.42 | 0.037 | 0.034 | 0.42 | 0.82 |
| Lactate, mM | 9.17 | 7.89 | 5.24 | 5.53 | 5.17 | 3.73 | 0.74 | <0.001 | 0.010 | 0.012 |
| Glucose, mM | 15.6 | 14.5 | 14.8 | 15.2 | 15.7 | 15.5 | 0.61 | 0.30 | 0.56 | 0.92 |
| T34,5, pg/mL | 4.76 | 6.47 | 6.08 | 5.16 | 3.76 | 3.43 | 0.977 | 0.008 | 0.97 | 0.61 |
| T44, pg/mL | 4.57 | 5.54 | 5.74 | 3.23 | 3.16 | 4.49 | 1.366 | 0.049 | 0.33 | 0.78 |
| Diet | s.e.d. 2 | p-Value 3 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Time/Diet | Hours | CON1 | BET | BET + AOX | Time | BET | Within BET | Timex BET | Time × within BET | |
| L* | 24 | 54.3 | 55.1 | 54.3 | 0.967 | 0.011 | 0.31 | 0.52 | 0.061 | 0.82 |
| 96 | 54.2 | 56.0 | 55.4 | |||||||
| a* | 24 | 2.20 | 2.50 | 2.30 | 0.283 | 0.91 | 0.68 | 0.57 | 0.26 | 0.82 |
| 96 | 2.22 | 2.31 | 2.40 | |||||||
| b* | 24 | 1.43 | 1.59 | 1.65 | 0.268 | 0.062 | 0.97 | 0.49 | 0.77 | 0.82 |
| 96 | 1.74 | 1.82 | 1.88 | |||||||
| pH | 0.25 | 6.68 | 6.68 | 6.78 | 0.053 | <0.001 | 0.27 | 0.094 | 0.67 | 0.10 |
| 1 | 6.48 | 6.49 | 6.58 | |||||||
| 24 | 5.92 | 5.93 | 5.93 | |||||||
| Temperature (T) | Thermoneutral | Heat Stress | s.e.d. 2 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON 1 | BET | BET + AOX | CON 1 | BET | BET + AOX | T | BET | Within BET | |
| Cooking loss4, % | 27.5 | 27.4 | 26.9 | 23.0 | 25.8 | 26.0 | 1.27 | 0.007 | 0.21 | 0.84 |
| Drip loss5, % | 2.11 | 1.74 | 1.78 | 3.43 | 1.15 | 2.22 | 0.51 | 0.33 | 0.007 | 0.20 |
| Water content, % | 74.8 | 76.5 | 75.9 | 74.6 | 75.2 | 75.3 | 0.79 | 0.079 | 0.13 | 0.56 |
| Shear force6, N | 18.4 | 20.4 | 23.9 | 23.0 | 22.8 | 21.3 | 1.95 | 0.30 | 0.24 | 0.34 |
| Breast yield, % | 22.8 | 24.4 | 24.9 | 22.7 | 22.9 | 23.6 | 0.92 | 0.057 | 0.066 | 0.37 |
| MFI7 | 86.3 | 79.7 | 87.7 | 58.1 | 62.8 | 81.6 | 6.23 | <0.001 | 0.14 | 0.003 |
| 0.5 h TBARS8 | 0.21 | 0.20 | 0.17 | 0.21 | 0.21 | 0.17 | 0.040 | 0.88 | 0.37 | 0.20 |
| 24 h TBARS8,9 | 5.32 | 3.48 | 2.00 | 6.12 | 5.51 | 5.32 | 0.545 | <0.001 | <0.001 | 0.029 |
| Temperature (T) | Thermoneutral | Heat Stress | s.e.d. 2 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diet | CON 1 | BET | BET + AOX | CON 1 | BET | BET + AOX | T | BET | Within BET | |
| Plasma 4, µmol/L | 85.0 | 140 | 145 | 102 | 109 | 125 | 10.50 | 0.043 | <0.001 | 0.14 |
| Muscle 5, µmol/g | 321 | 635 | 413 | 425 | 518 | 567 | 9.38 | 0.43 | 0.009 | 0.16 |
| Liver 6, µmol/g | 403 | 191 | 187 | 208 | 143 | 145 | 45.1 | <0.001 | <0.001 | 0.98 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress. Antioxidants 2019, 8, 336. https://doi.org/10.3390/antiox8090336
Shakeri M, Cottrell JJ, Wilkinson S, Le HH, Suleria HAR, Warner RD, Dunshea FR. Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress. Antioxidants. 2019; 8(9):336. https://doi.org/10.3390/antiox8090336
Chicago/Turabian StyleShakeri, Majid, Jeremy J. Cottrell, Stuart Wilkinson, Hieu H. Le, Hafiz A. R. Suleria, Robyn D. Warner, and Frank R. Dunshea. 2019. "Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress" Antioxidants 8, no. 9: 336. https://doi.org/10.3390/antiox8090336
APA StyleShakeri, M., Cottrell, J. J., Wilkinson, S., Le, H. H., Suleria, H. A. R., Warner, R. D., & Dunshea, F. R. (2019). Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress. Antioxidants, 8(9), 336. https://doi.org/10.3390/antiox8090336