Thunbergia laurifolia Leaf Extract Increased Levels of Antioxidant Enzymes and Protected Human Cell-Lines In Vitro Against Cadmium

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Raw Material
2.3. Plant Preparation and Extraction
2.4. Proximate Compositions and Mineral Contents
2.5. Cell Culture
2.6. Determination of the CDE Effects on Antioxidant Enzyme Activity and Malondialdehyde Value
2.6.1. Preparation of Endogenous Cellular Extract
2.6.2. Catalase (CAT) Activity
2.6.3. Glutathione Peroxidase (GPx) Activity
2.6.4. Malondialdehyde (MDA) Value
2.7. Determination of CDE Cytotoxicity
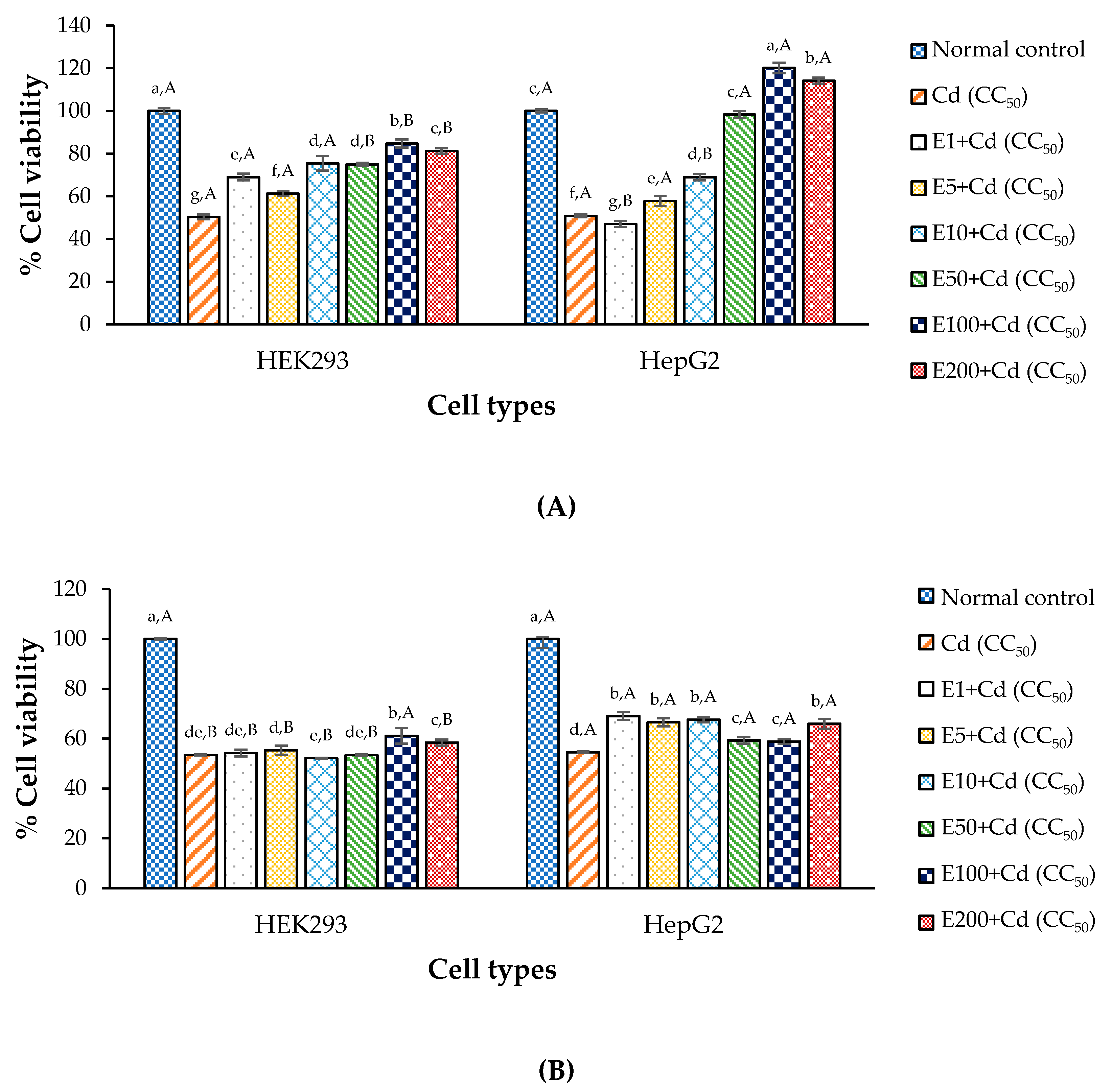
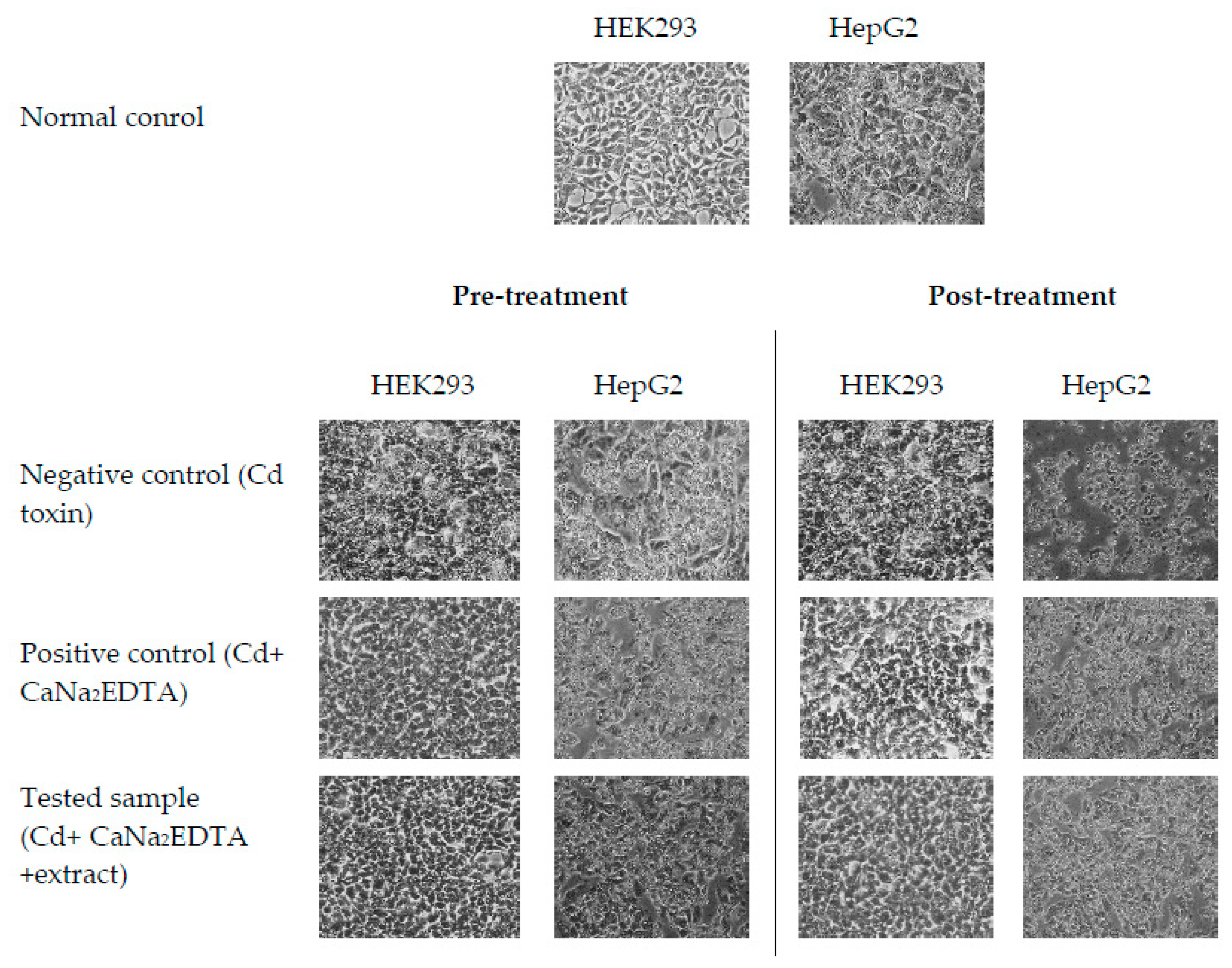
2.8. Determination of CDE’s Anti-Cd Property
| Normal control | : Media (24 h) |
| CC50 of Cd (negative control) | : Media and Cd at CC50 (24 h) |
| Tested sample | : CDE/CaNa2EDTA and Cd at CC50 (24 h) |
| Normal control | : Media (24 h) + Media (24 h) |
| CC50 of Cd (negative control) | : Media (24 h) + Cd at CC50 (24 h) |
| Tested sample | : CDE (24 h)/CaNa2EDTA + Cd at CC50 (24 h) |
| Normal control | : Media (24 h) + Media (24 h) |
| CC50 of Cd (negative control) | : Cd at CC50 (24 h) + Media (24 h) |
| Tested sample | : Cd at CC50 (24 h) + CDE/CaNa2EDTA (24 h) |
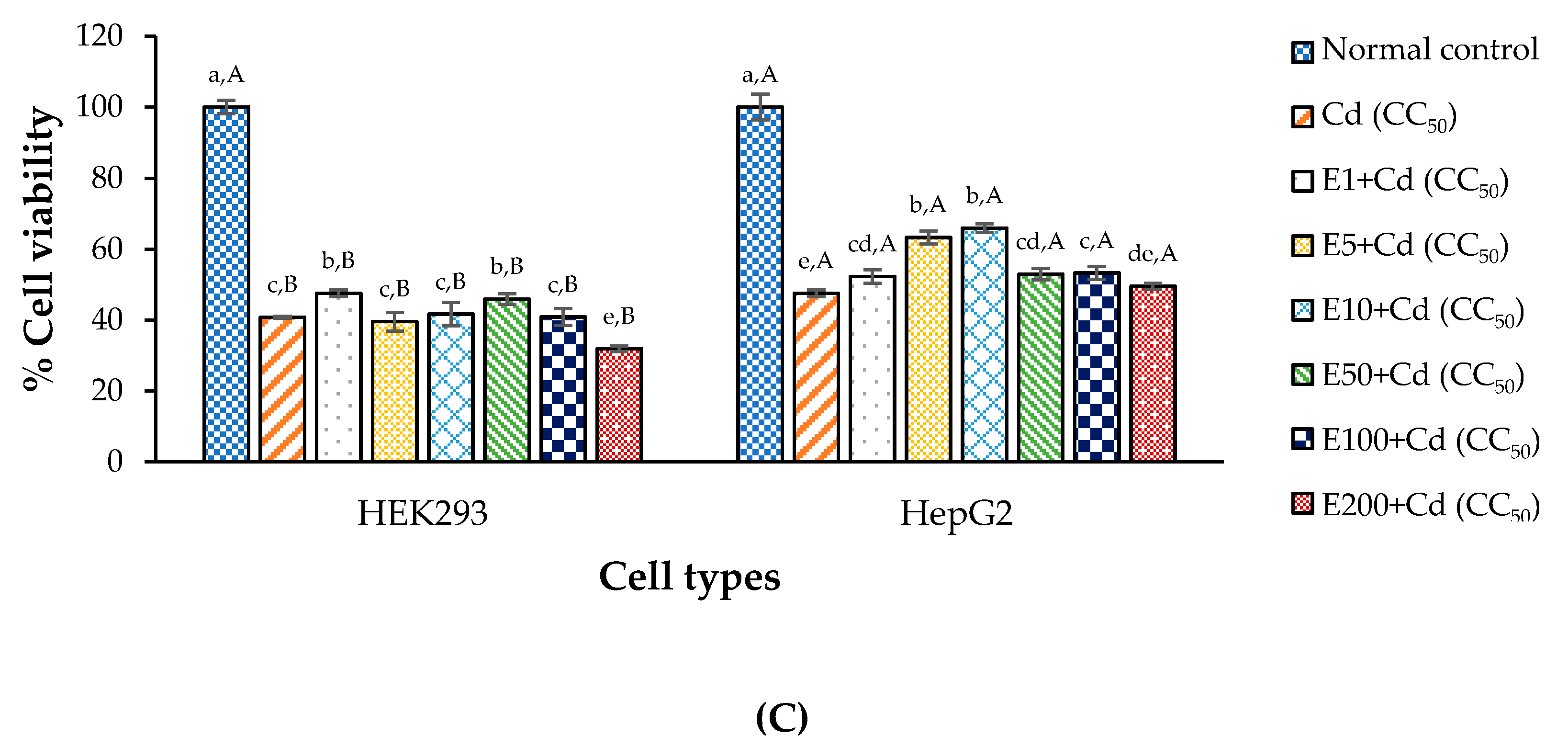
2.9. CDE Coupled with a Chelating Agent (CaNa2EDTA) as Anti-Cd Treatment
| Normal control | : Media (24 h) + Media (24 h) |
| CC50 of Cd (negative control) | : Media (24 h) + Cd at CC50 (24 h) |
| CC50 of Cd + EDTA (positive control) | : Media (24 h) + CaNa2EDTA mixed with Cd at CC50 (24 h) |
| Tested sample | : CDE (24 h) + CaNa2EDTA mixed with Cd at CC50 (24 h) |
| Normal control | : Media (24 h) + Media (24 h) |
| CC50 of Cd (negative control) | : Cd at CC50 (24 h) + Media (24 h) |
| CC50 of Cd + EDTA (positive control) | : CaNa2EDTA mixed with Cd at CC50 (24 h) + Media (24 h) |
| Tested sample | : CaNa2EDTA mixed with Cd at CC50 (24 h) + CDE (24 h) |
2.10. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition and Mineral Contents in T. laurifolia Leaves
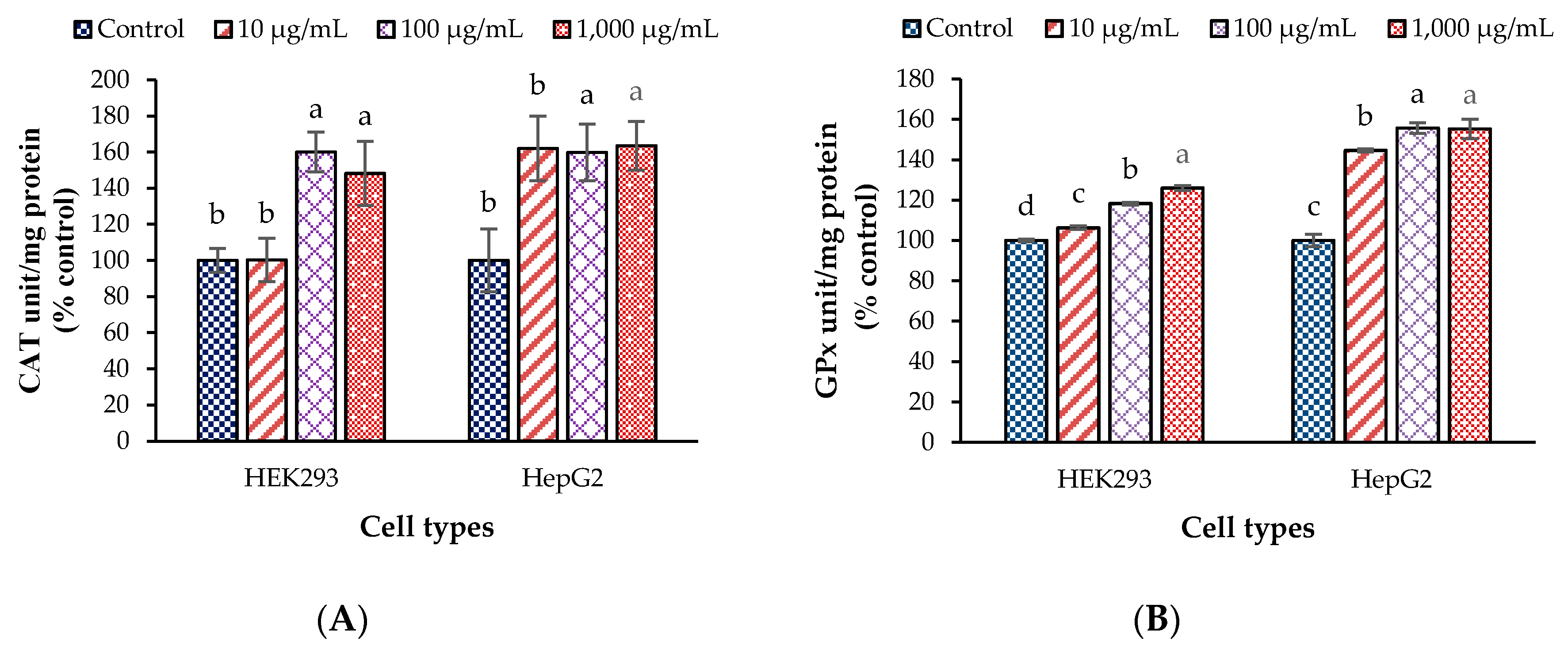
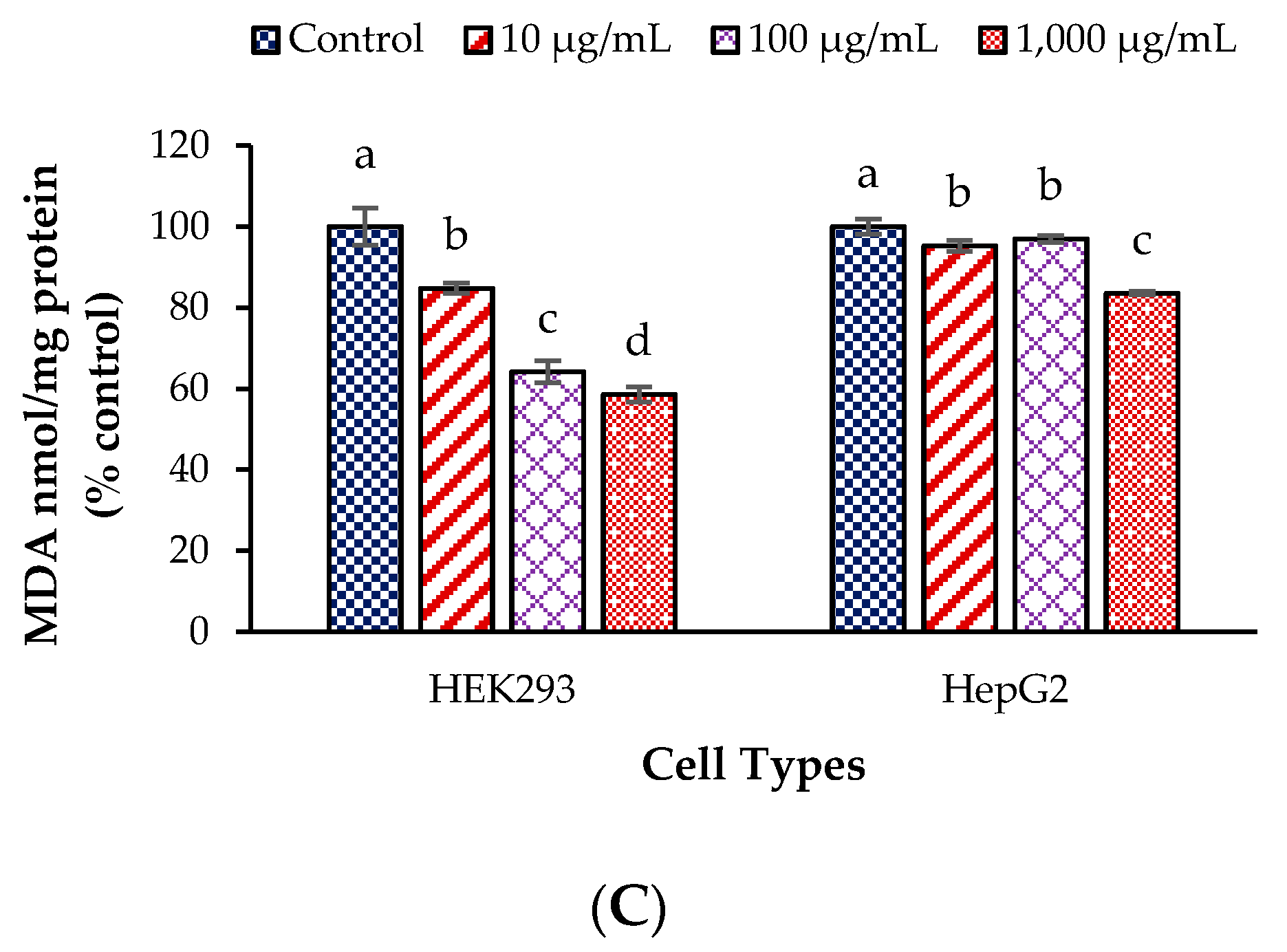
3.2. Antioxidant Enzyme Activities and Malondialdehyde Values in the Test Cells
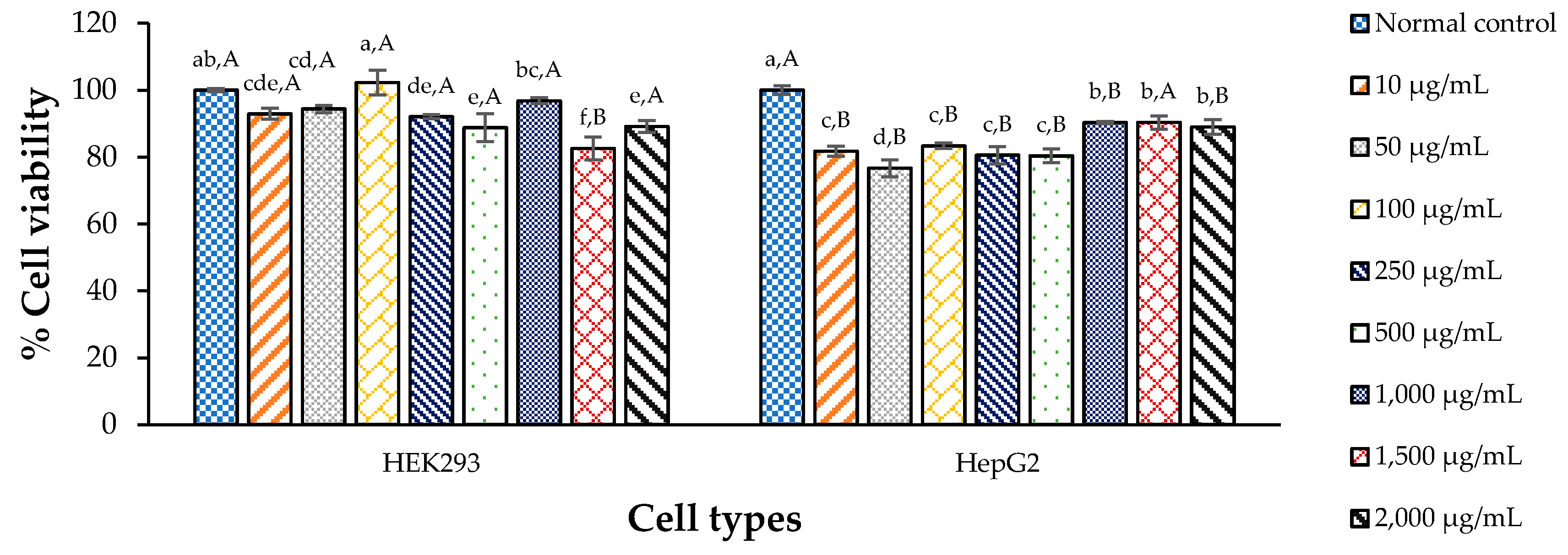
3.3. Cytotoxicity of CDE against the Test Cells
3.4. Cytotoxicity of Cd against the Test Cells
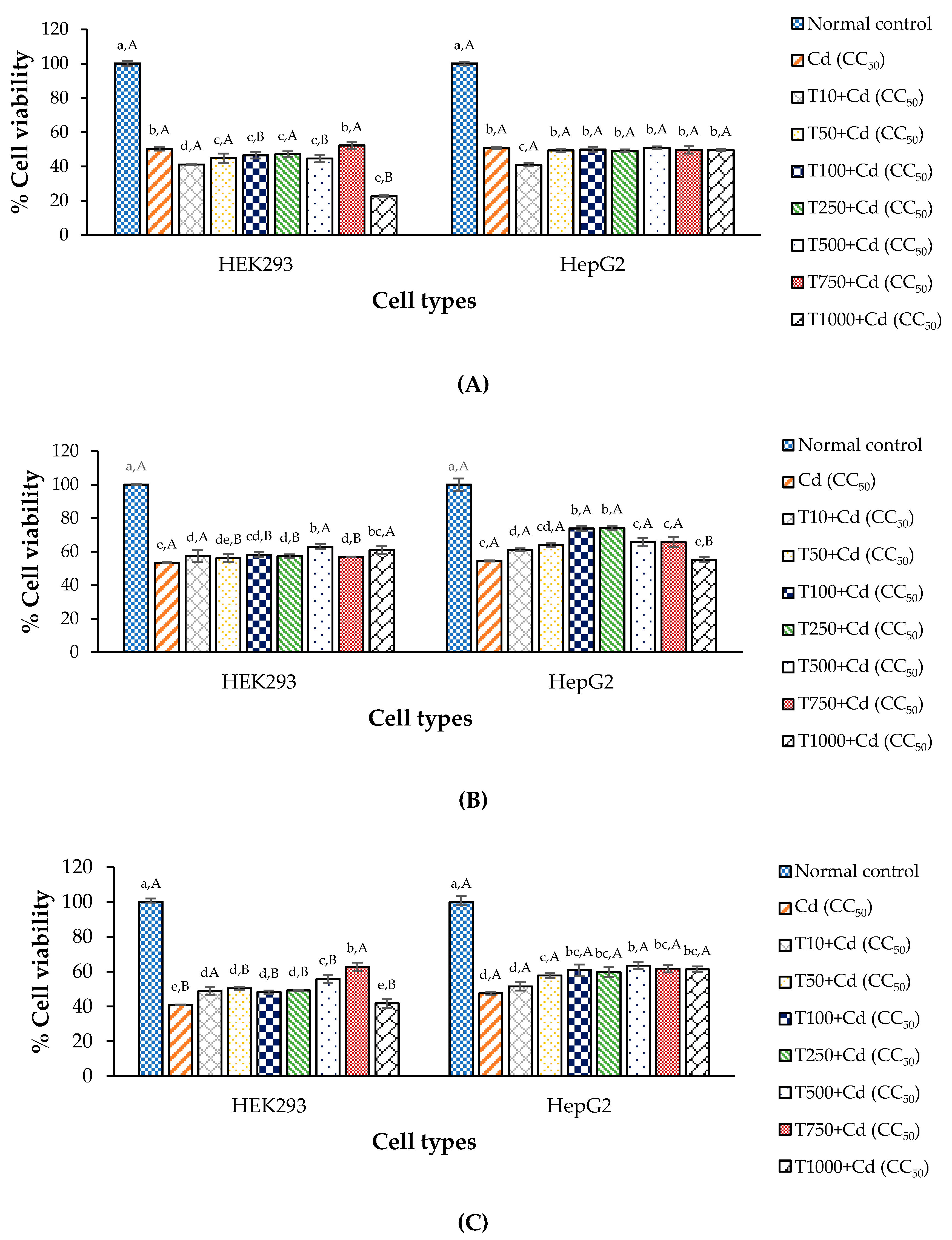
3.5. Anti-Cd Toxicity of CDE in Cell Lines
3.6. Anti-Cd Toxicity Property of Chelating Agent in the Test Cells
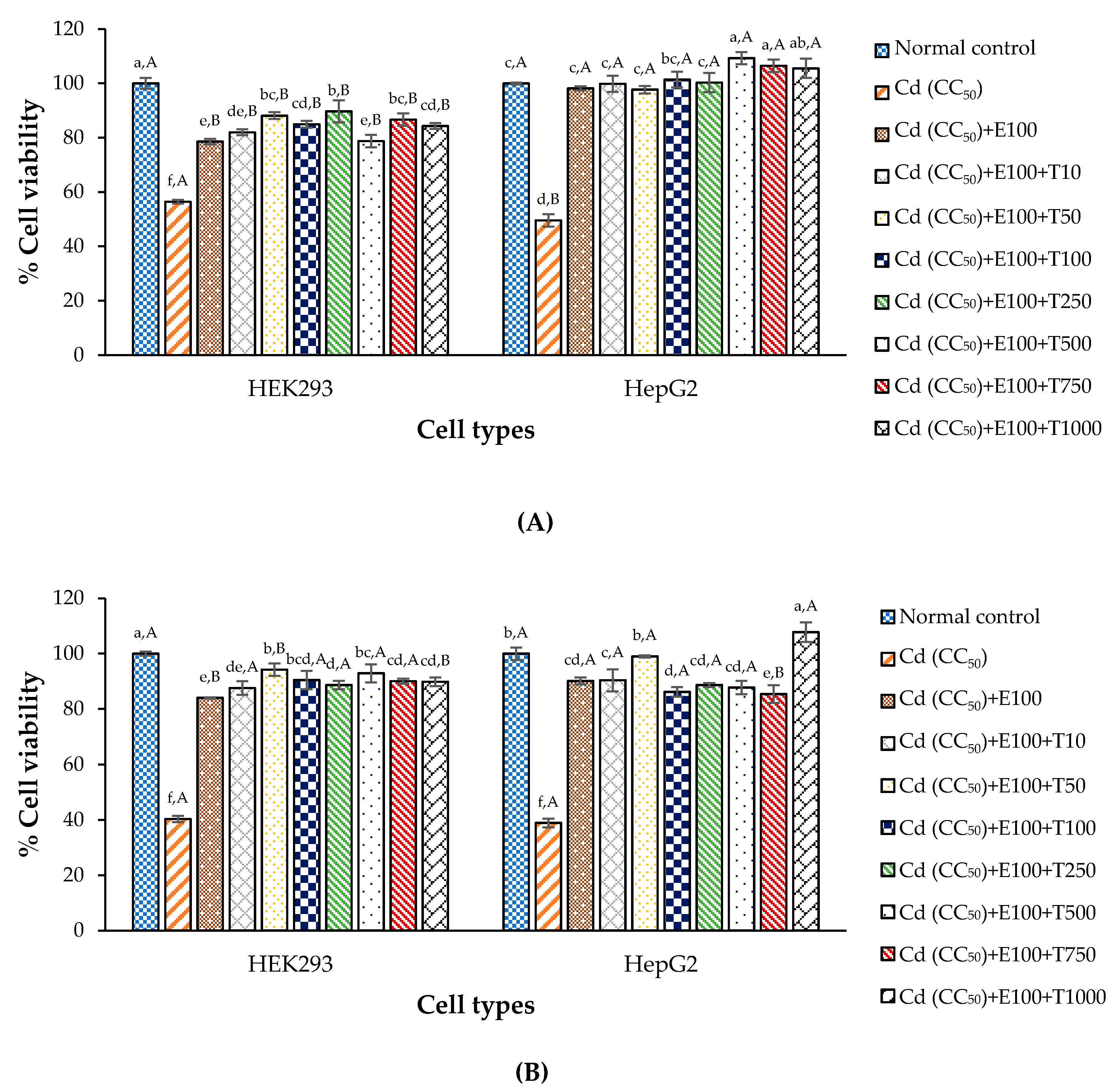
3.7. Anti-Cd Toxicity Property of CDE and Chelating Agent on the Test Cells
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roopha, P.D.; Padmalatha, C. Effect of herbal preparation on heavy metal (cadmium) induced antioxidant system in female wistar rats. J. Med. Toxicol. 2012, 8, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Järup, L. Hazards of heavy metal contamination. Brit. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Han, X.Y.; Xu, Z.R.; Wang, Y.Z.; Huang, Q.C. Effect of cadmium on lipid peroxidation and activities of antioxidant enzymes in growing pigs. Biol. Trace Elem. Res. 2006, 110, 251–263. [Google Scholar] [CrossRef]
- Gonick, H.C. Nephrotoxicity of cadmium and lead. Indian J. Med. Res. 2008, 128, 335–352. [Google Scholar]
- Ercal, N.; Gurer-Orhan, H.; Aykin-Burns, N. Toxic metals and oxidative stress part I: Mechanisms involved in metal induced oxidative damage. Curr. Top. Med. Chem. 2001, 1, 529–539. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Lee, J.Y.; Tokumoto, M.; Satoh, M. Cadmium renal toxicity via apoptotic pathways. Biol. Pharm. Bull. 2012, 35, 1892–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.Y.; Chan, K.M. Mechanism of cadmium-induced cytotoxicity on the ZFL zebrafish liver cell line. Metallomics 2012, 4, 1064–1076. [Google Scholar] [CrossRef] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Rutin, a bioflavonoid antioxidant protects rat pheochromocytoma (PC-12) cells against 6-hydroxydopamine (6-OHDA)-induced neurotoxicity. Int. J. Mol. Med. 2013, 32, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trchounian, A.; Petrosyan, M.; Sahakyan, N. Plant Cell Redox Homeostasis and Reactive Oxygen Species. In Redox State as a Central Regulator of Plant-Cell Stress Responses; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer Nature: Basel, Switzerland, 2016; pp. 25–50. [Google Scholar]
- Thongsaard, W.; Marsden, C.A. A herbal medicine used in the treatment of addiction mimics the action of amphetamine on in vitro rat striatal dopamine release. Neurosci. Lett. 2002, 329, 129–132. [Google Scholar] [CrossRef]
- Chuthaputti, A. Rang Jerd: Laurel clock vine (Thunbergia laurifolia Lindl.): A detoxifying herb. J. Thai Tradit. Altern. Med. 2010, 8, 211–220. [Google Scholar]
- Oonsivilai, R. Functional and Nutraceutical Properties of Rang Chuet (Thunbergia laurifolia Lindl.) Extracts. Ph.D. Thesis, Suranaree University of Technology, Nakhon Ratchasima, Thailand, 2006. [Google Scholar]
- Chan, E.W.C.; Lye, P.Y.; Eng, S.Y.; Tan, Y.P. Antioxidant properties of herbs with enhancement effects of drying treatments: A synopsis. Free Radic. Antioxid. 2013, 3, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Junsi, M.; Siripongvutikorn, S.; Takahashi Yupanqui, C.; Usawakesmanee, W. Effcacy of Thunbergia laurifolia (Rang Jued) aqueous leaf extract for specific biological activities using RAW 264.7 macrophage cells as test model. Int. Food Res. J. 2017, 24, 2317–2329. [Google Scholar]
- Sabolić, I.; Breljak, D.; Škarica, M.; Herak-Kramberger, C.M. Role of metallothionein in cadmium traffic and toxicity in kidneys and other mammalian organs. Biometals 2010, 23, 897–926. [Google Scholar] [CrossRef] [PubMed]
- Helali, I.; Ferchichi, S.; Maaouia, A.; Aouni, M.; Harizi, H. Modulation of macrophage functionality induced in vitro by chlorpyrifos and carbendazim pesticides. J. Immunotoxicol. 2016, 13, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Ruangyuttikarn, W.; Chattaviriya, P.; Morkmek, N.; Chuncharunee, S.; Lertprasertsuke, N. Thunbergia laurifolia leaf extract mitigates cadmium toxicity in rats. Sci. Asia 2013, 39, 19–25. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- American Type Culture Collection. ATCC Animal Cell Culture Guide: American Type Culture Collection; University Blvd: Manassas, VA, USA, 2014; pp. 2–7. [Google Scholar]
- Du, Y.; Esfandi, R.; Willmore, W.G.; Tsopmo, A. Antioxidant activity of oat proteins derived peptides in stressed hepatic HepG2 cells. Antioxidants 2016, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Flohe, L.; Gunzler, W. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxic assays. J. Immunol. Met. 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Jaiboon, V.; Boonyanupahap, J.; Suwansri, S.; Ratanatraiwong, P.; Hansawasdi, C. Alpha amylase inhibition and roasting time of local vegetables and herbs prepared for diabetes risk reduction chili paste. As. J. Food Ag-Ind. 2010, 3, 1–12. [Google Scholar]
- Adnan, M.; Ahmad, A.; Ahmed, A.; Khalid, N.; Hayat, I.; Ahmed, I. Chemical composition and sensory evaluation of tea (Camellia sinensis) commercialized in Pakistan. Pak. J. Bot. 2013, 45, 901–907. [Google Scholar]
- Pyankov, V.I.; Ivanov, L.A.; Lambers, H. Chemical composition of the leaves of plants with different ecological strategies from the boreal zone. Russ. J. Ecol. 2001, 32, 221–229. [Google Scholar] [CrossRef]
- Stein, R.J.; Horeth, S.; Melo, J.R.F.; Syllwasschy, L.; Lee, G.; Garbin, M.L.; Clemens, S.; Krämer, U. Relationships between soil and leaf mineral composition are element-specific, environment-dependent and geographically structured in the emerging model Arabidopsis halleri. New Phytol. 2017, 213, 1274–1286. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, K.; Kocot, J.; Horecka, A. Biochemistry of magnesium. J. Elementol. 2010, 15, 601–616. [Google Scholar] [CrossRef]
- Page, M.J.; Cera, E.D. Role of Na+ and K+ in Enzyme Function. Physiol. Rev. 2006, 86, 1049–1092. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.H.; Huang, J.Q.; Li, X.Q.; Zheng, B.S.; Wu, J.S.; Wang, Z.J.; Liu, G.H.; Chen, M. Effects of potassium supply on limitations of photosynthesis by mesophyll diffusion conductance in Carya cathayensis. Tree Physiol. 2011, 31, 1142–1151. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, S.A.; Krämer, U. The zinc homeostasis network of land plants. Biochim. Biophys. Acta. 2012, 1823, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Marreiro, D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; Oliveira, A.R.S. Zinc and oxidative stress: Current mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Brinate, S.V.B.; Martins, L.D.; Rosa, G.N.G.P.; Cunha, V.V.; Sotero, A.J.; Amaral, J.F.T.; Junior, W.C.J.; Tomaz, M.A. Copper can influences growth, disease control and production in arabica coffee trees. Aus. J. Crop Sci. 2015, 9, 678–683. [Google Scholar]
- Copper for Crop Production. Available online: https://www.extension.umn.edu/agriculture/nutrientmanagement/micronutrients/copper-forcrop-production/ (accessed on 20 March 2018).
- Vural, H.; Demirin, H.; Kara, Y.; Eren, I.; Delibas, N. Alterations of plasma magnesium, copper, zinc, iron and selenium concentrations and some related erythrocyte antioxidant enzyme activities in patients with Alzheimer’s disease. J. Trace Elem. Med. Biol. 2010, 24, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Schriner, S.E.; Linford, N. Extension of mouse lifespan by overexpression of catalase. Age 2006, 28, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Flora, S.J.S.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501–523. [Google Scholar]
- Bak, M.J.; Jun, M.; Jeong, W.S. Antioxidant and hepatoprotective effects of the red ginseng essential oil in H2O2-treated HepG2 cells and CCl4-treated mice. Int. J. Mol. Sci. 2012, 13, 2314–2330. [Google Scholar] [CrossRef] [Green Version]
- Ognjanović, B.I.; Pavlović, S.Z.; Maletić, S.D.; Žikić, R.V.; Štajn, A.Š.; Radojičić, R.M.; Saičić, Z.S.; Petrović, V.M. Protective influence of vitamin E on antioxidant defense system in the blood of rats treated with cadmium. Physiol. Res. 2003, 52, 563–570. [Google Scholar]
- Junsi, M.; Siripongvutikorn, S.; Takahashi Yupanqui, C.; Usawakesmanee, W. Phenolic and flavonoid compounds in aqueous extracts of Thunbergia laurifolia leaves and their effect on the toxicity of the carbamate insecticide methomyl to murine macrophage cells. Funct. Food Health Dis. 2017, 7, 529–544. [Google Scholar] [CrossRef]
- Olayinka, E.T.; Ola, O.S.; Ore, A.; Adeyemo, O.A. Ameliorative effect of caffeic acid on capecitabine-induced hepatic and renal dysfunction: Involvement of the antioxidant defence system. Medicines 2017, 4, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayanthi, R.; Subash, P. Antioxidant effect of caffeic acid on oxytetracycline induced lipid peroxidation in albino rats. Ind. J. Clin. Biochem. 2010, 25, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almaghrabi, O.A. Molecular and biochemical investigations on the effect of quercetin on oxidative stress induced by cisplatin in rat kidney. Saudi J. Biol. Sci. 2015, 22, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.W. Protective Effect of apigenin against oxidative stress-induced damage in osteoblastic cells. Int. J. Mol. Med. 2014, 33, 1327–1334. [Google Scholar] [CrossRef] [Green Version]
- Haron, D. The aging: Major risk factor for disease and death. Proc. Natl. Acad. Sci. USA 1991, 88, 5360–5364. [Google Scholar]
- Jung, U.J.; Lee, M.K.; Park, Y.B.; Jeon, S.M.; Choi, M.S. Antihyperglycemic and antioxidant properties of caffeic acid in db/db mice. J. Pharmacol. Exp. Ther. 2006, 318, 476–483. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, F.B.; Fetoui, H.; Fakhfakh, F.; Keskes, L. Caffeic acid and quercetin protect erythrocytes against the oxidative stress and the genotoxic effects of lambda-cyhalothrin in vitro. Hum. Exp. Toxicol. 2011, 31, 92–100. [Google Scholar] [CrossRef]
- Ioset, J.; Brun, R.; Wenzler, T.; Kaiser, M.; Yardley, V. Drug Screening for Kinetoplastids Diseases: A Training Manual for Screening in Neglected Diseases; DNDi and Pan-Asian Screening Network: Genève, Switzerland, 2009; pp. 29–31. [Google Scholar]
- Oonsivilai, R.; Ferruzzi, M.G.; Ningsanond, S. Antioxidant activity and cytotoxicity of Rang Chuet (Thunbergia laurifolia Lindl.) extracts. As. J. Food Ag-Ind. 2008, 1, 116–128. [Google Scholar]
- Chivapat, S.; Chavalittumrong, P.; Attawish, A.; Bansiddhi, J.; Padungpat, S. Chronic toxicity of Thunbergia laurifolia Lindl. extract. J. Thai Tradit. Altern. Med. 2009, 7, 17–24. [Google Scholar]
- Brooks, A.N.; Turkarslan, S.; Beer, K.D.; Lo, F.Y.; Baliga, N.S. Adaptation of cells to new environments. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 544–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bara, M.; Guiet-Bara, A.; Durlach, J. Regulation of sodium and potassium pathways by magnesium in cell membranes. Magnes. Res. 1993, 6, 167–177. [Google Scholar] [PubMed]
- Rose, A.M.; Valdes, R. Understanding the sodium pump and its relevance to disease. Clin. Chem. 1994, 40, 1674–1685. [Google Scholar] [PubMed]
- Blaine, J.; Chonchol, M.; Levi, M. Renal control of calcium, phosphate, and magnesium homeostasis. Clin. J. Am. Soc. Nephrol. 2015, 10, 1257–1272. [Google Scholar] [CrossRef] [PubMed]
- Navas-Acien, A.; Selvin, E.; Sharrett, A.R.; Calderon-Aranda, E.; Silbergeld, E.; Guallar, E. Lead, cadmium, smoking, and increased risk of peripheral arterial disease. Circulation 2004, 109, 3196–3201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, S.K.; Gowder, S.J.T. Members of Antioxidant Machinery and Their Functions. In Basic Principles and Clinical Significance of Oxidative Stress; Gowder, S.J.T., Ed.; InTech: Rijeka, Croatia, 2015; pp. 59–85. [Google Scholar]
- Chattaviriya, P.; Morkmek, N.; Lertprasertsuke, N.; Ruangyuttikarn, W. Drinking Thunbergia laurifolia Lindl. leaf extract helps prevent renal toxicity induced by cadmium in rats. Thai J. Toxicology. 2010, 25, 124–132. [Google Scholar]
- Morkmek, N.; Chattaviriya, P.; Lertprasertsuke, N.; Chuncharunee, S.; Ruangyuttikarn, W. Detoxification of cadmium induced renal and hepatic injuries in rats by Thunbergia laurifolia Lindl. leaf extract. Thai J. Toxicology. 2010, 25, 115–123. [Google Scholar]
- Lachman, J.; Dudjak, J.; Miholová, D.; Kolihová, D.; Pivec, V. Effect of cadmium on flavonoid content in young barley (Hordeum sativum L.) plants. Plant Soil Environ. 2005, 51, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Tarahovsky, Y.S.; Kim, Y.A.; Yagolnik, E.A.; Muzafarov, E.N. Flavonoid–membrane interactions: Involvement of flavonoid–metal complexes in raft signaling. Biochim. Biophys. Acta 2014, 1838, 1235–1246. [Google Scholar] [CrossRef] [Green Version]
- Gonzales, G.B.; Camp, J.V.; Vissenaekens, H.; Raes, K.; Smagghe, G.; Grootaert, C. Review on the use of cell cultures to study metabolism, transport, and accumulation of flavonoids: From mono-cultures to co-culture systems. Compr. Rev. Food Sci. F. 2015, 14, 741–754. [Google Scholar] [CrossRef]
- Flora, S.J.S.; Pachauri, V. Chelation in metal intoxication. Int. J. Environ. Res. Pu. 2010, 7, 2745–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaurock-Busch, E.; Busch, Y.M. Comparison of chelating agents DMPS, DMSA and EDTA for the diagnosis and treatment of chronic metal exposure. Br. J. Med. Med. Res. 2014, 4, 1821–1835. [Google Scholar] [CrossRef]
- Smith, S.W. The role of chelation in the treatment of other metal poisonings. J. Med. Toxicol. 2013, 9, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudtson, M.L.; Wyse, K.G.; Galbraith, P.D. Chelation therapy for ischemic heart disease. A randomized controlled trail. JAMA 2002, 287, 481–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballal, N.V.; Kundabala, M.; Bhat, S.; Rao, N.; Rao, B.S. A comparative in vitro evaluation of cytotoxic effects of EDTA and maleic acid: Root canal irrigants. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2009, 108, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Zaccaro Scelza, M.F.; da Silva Pierro, V.S.; Chagas, M.A.; da Silva, L.E.; Scelza, P. Evaluation of inflammatory response of EDTA, EDTA-T, and citric acid in animal model. J. Endod. 2010, 36, 515–519. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Dried-Leaf T. laurifolia |
|---|---|
| Proximate compositions (% dry weight basis) | |
| Crude protein | 13.98 ± 1.90 |
| Crude fat | 1.83 ± 0.12 |
| Ash | 19.93 ± 1.90 |
| Crude fiber | 11.16 ± 0.44 |
| Carbohydrate | 53.10 ± 2.23 |
| Mineral contents (mg/kg dry leaf) | |
| Potassium (K) | 20,600.43 |
| Phosphorus (P) | 2500.75 |
| Magnesium (Mg) | 4548.30 |
| Iron (Fe) | 84.14 |
| Zinc (Zn) | 35.51 |
| Copper (Cu) | 16.99 |
| Selenium (Se) | 0.73 |
| Cell Types | CC50 of Cd (μmol/L) |
|---|---|
| HEK293 | 64.09 |
| HepG2 | 75.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Junsi, M.; Takahashi Yupanqui, C.; Usawakesmanee, W.; Slusarenko, A.; Siripongvutikorn, S. Thunbergia laurifolia Leaf Extract Increased Levels of Antioxidant Enzymes and Protected Human Cell-Lines In Vitro Against Cadmium. Antioxidants 2020, 9, 47. https://doi.org/10.3390/antiox9010047
Junsi M, Takahashi Yupanqui C, Usawakesmanee W, Slusarenko A, Siripongvutikorn S. Thunbergia laurifolia Leaf Extract Increased Levels of Antioxidant Enzymes and Protected Human Cell-Lines In Vitro Against Cadmium. Antioxidants. 2020; 9(1):47. https://doi.org/10.3390/antiox9010047
Chicago/Turabian StyleJunsi, Marasri, Chutha Takahashi Yupanqui, Worapong Usawakesmanee, Alan Slusarenko, and Sunisa Siripongvutikorn. 2020. "Thunbergia laurifolia Leaf Extract Increased Levels of Antioxidant Enzymes and Protected Human Cell-Lines In Vitro Against Cadmium" Antioxidants 9, no. 1: 47. https://doi.org/10.3390/antiox9010047
APA StyleJunsi, M., Takahashi Yupanqui, C., Usawakesmanee, W., Slusarenko, A., & Siripongvutikorn, S. (2020). Thunbergia laurifolia Leaf Extract Increased Levels of Antioxidant Enzymes and Protected Human Cell-Lines In Vitro Against Cadmium. Antioxidants, 9(1), 47. https://doi.org/10.3390/antiox9010047