Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology



Abstract
:1. Introduction
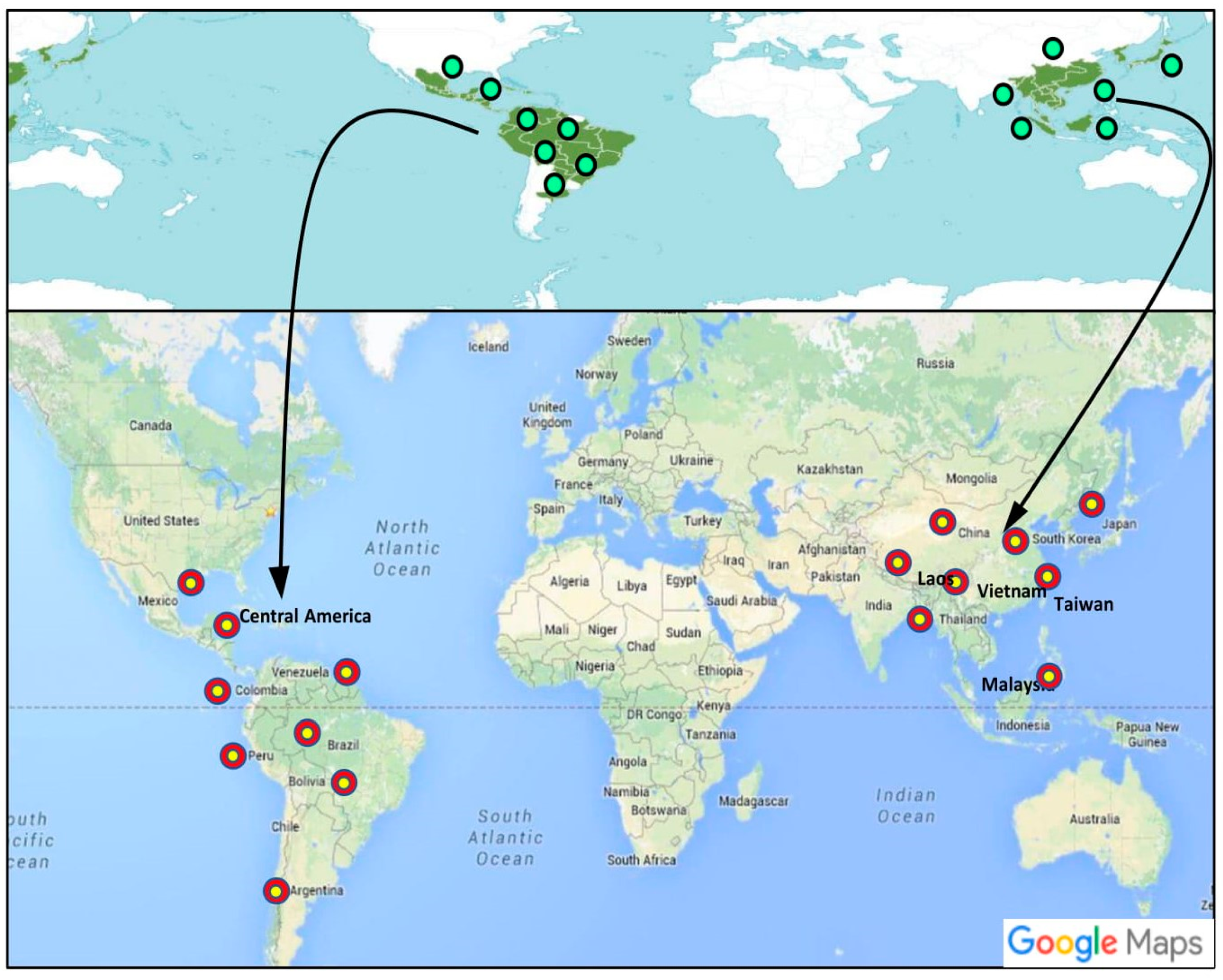
2. Taxonomy, Distribution, and Description of Dendropanax Species
3. Phytochemistry of Dendropanax Species
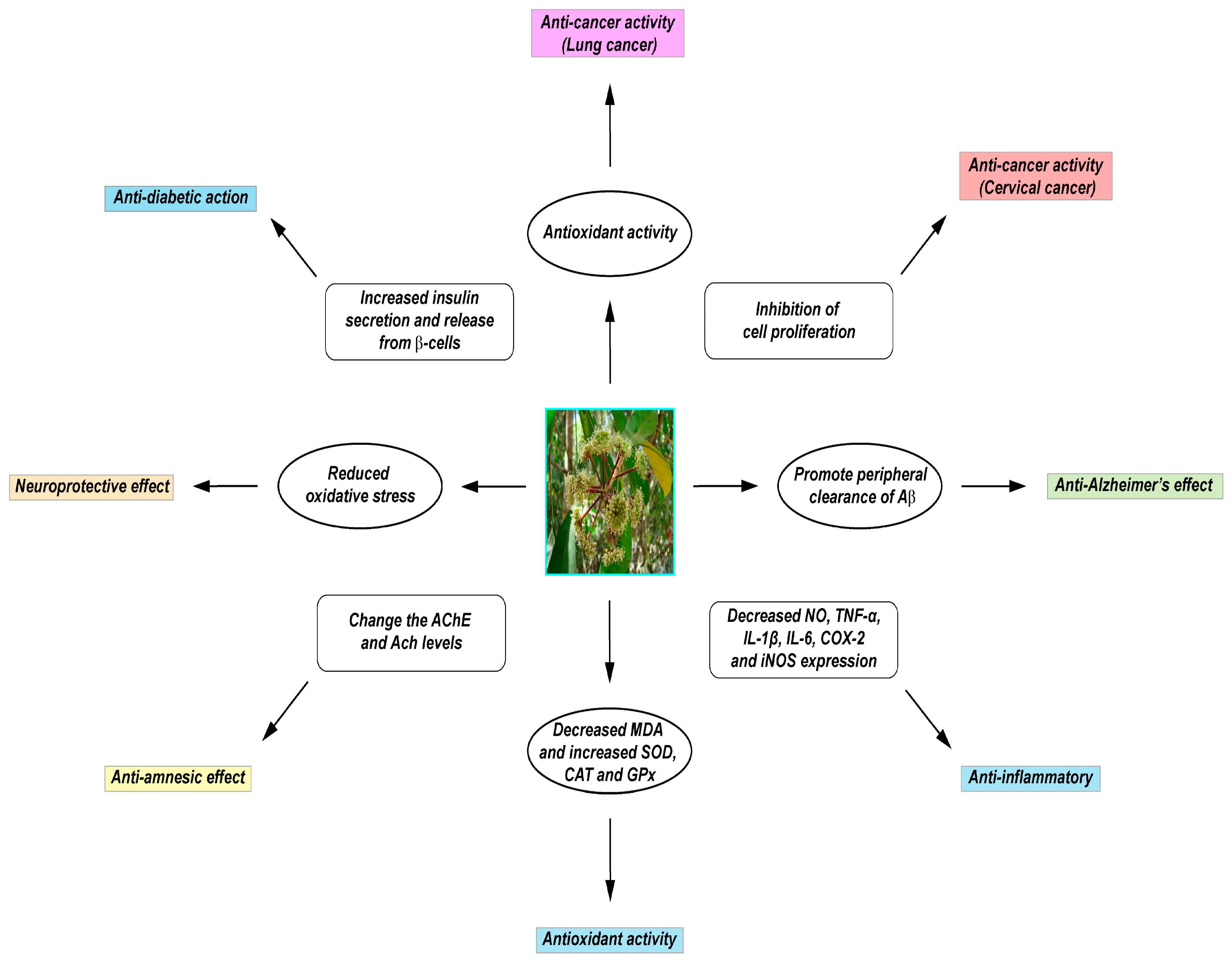
4. Medicinal Uses and Pharmacological Properties of D. morbifera
4.1. Antioxidant Properties of D. morbifera
4.1.1. In Vitro Studies
4.1.2. In Vivo Studies
4.2. Anti-Inflammatory Properties of D. morbifera
4.2.1. In Vitro Studies
4.2.2. In Vivo Studies
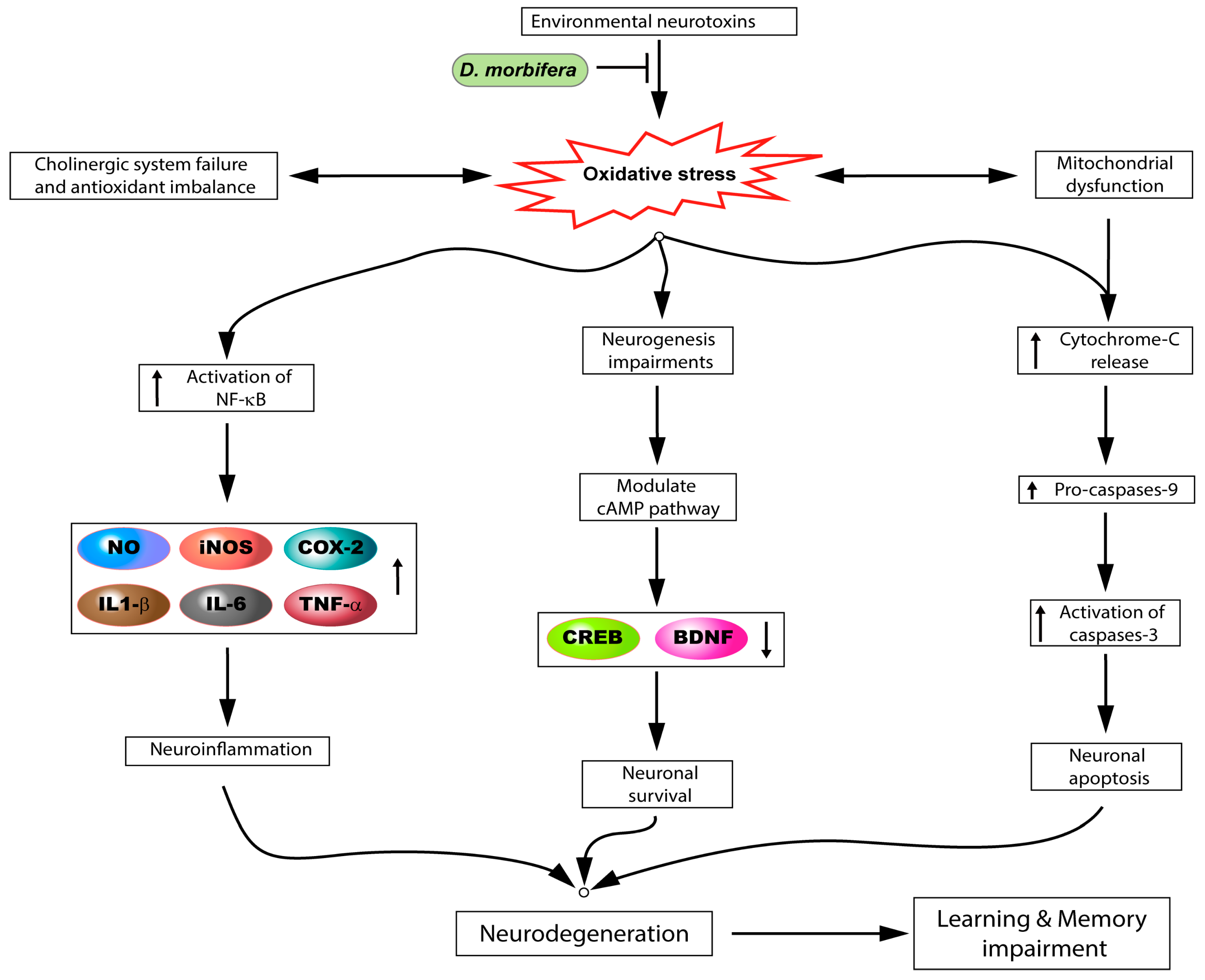
4.3. Action of D. morbifera on the Central Nervous System
4.3.1. In Vitro
4.3.2. In Vivo
4.4. Anti-cancer Activity of D. morbifera
In Vitro and In Vivo Studies
4.5. Anti-Diabetic Activity of D. morbifera
In Vitro and In Vivo Studies
4.6. Hepatoprotective and Immunomodulatory Effects of D. morbifera
4.7. Antimicrobial, Antiplasmodial, and Anticomplementary Activities of D. morbifera
4.8. Cytotoxicity and Toxicity Effects of D. morbifera
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Asadi-Samani, M.; Kafash-Farkhad, N.; Azimi, N.; Fasihi, A.; Alinia-Ahandani, E.; Rafieian-Kopaei, M. Medicinal plants with hepatoprotective activity in Iranian folk medicine. Asian Pac. J. Trop. Biomed. 2015, 5, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Samani, M.A.; Rafieian, M.K.; Azimi, N. Gundelia: A systematic review of medicinal and molecular perspective. Pakistan J. Biol. Sci. 2013, 16, 1238–1247. [Google Scholar]
- Kafash-Farkhad, N.; Asadi-Samani, M.; Rafieian-Kopaei, M. A review on phytochemistry and pharmacological effects of Prangos ferulacea (L.) Lindl. Life Sci. J. 2013, 10, 360–367. [Google Scholar]
- Asadi-Samani, M.; Bahmani, M.; Rafieian-Kopaei, M. The chemical composition, botanical characteristic and biological activities of Borago officinalis: A review. Asian Pac. J. Trop. Med. 2014, 7, S22–S28. [Google Scholar] [CrossRef] [Green Version]
- Kooti, W.; Moradi, M.; Ali-Akbari, S.; Sharafi-Ahvazi, N.; Asadi-Samani, M.; Ashtary-Larky, D. Therapeutic and pharmacological potential of Foeniculum vulgare Mill: A review. J. HerbMed Pharmacol. 2015, 4, 1–9. [Google Scholar]
- Popović, Z.; Matić, R.; Bojović, S.; Stefanović, M.; Vidaković, V. Ethnobotany and herbal medicine in modern complementary and alternative medicine: An overview of publications in the field of I&C medicine 2001-2013. J. Ethnopharmacol. 2016, 181, 182–192. [Google Scholar]
- Rashrash, M.; Schommer, J.C.; Brown, L.M. Prevalence and Predictors of Herbal Medicine Use Among Adults in the United States. J. Patient Exp. 2017, 4, 108–113. [Google Scholar] [CrossRef]
- Rates, S.M.K. Plants as source of drugs. Toxicon 2001, 39, 603–613. [Google Scholar] [CrossRef]
- Song, J.H.; Kang, H.B.; Kim, J.H.; Kwak, S.; Sung, G.J.; Park, S.H.; Jeong, J.H.; Kim, H.; Lee, J.; Jun, W.; et al. Antiobesity and cholesterol-lowering effects of Dendropanax morbifera water extracts in mouse 3T3-L1 Cells. J. Med. Food. 2018, 21, 793–800. [Google Scholar] [CrossRef]
- Sun, S.; Li, T.; Jin, L.; Piao, Z.H.; Liu, B.; Ryu, Y.; Choi, S.Y.; Kim, G.R.; Jeong, J.E.; Wi, A.J.; et al. Dendropanax morbifera prevents cardiomyocyte hypertrophy by inhibiting the Sp1/GATA4 pathway. Am. J. Chin. Med. 2018, 46, 1021–1044. [Google Scholar] [CrossRef]
- Setzer, W.N.; Green, T.J.; Whitaker, K.W.; Moriarity, D.M.; Yancey, C.A.; Lawton, R.O.; Bates, R.B. A cytotoxic diacetylene from Dendropanax arboreus. Planta Med. 1995, 61, 470–471. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wen, J. Phylogeny and Biogeography of Dendropanax (Araliaceae), an Amphi-Pacific Disjunct Genus Between Tropical/Subtropical Asia and the Neotropics. Syst. Bot. 2013, 38, 536–551. [Google Scholar] [CrossRef]
- Park, S.Y.; Karthivashan, G.; Ko, H.M.; Cho, D.Y.; Kim, J.; Cho, D.J.; Ganesan, P.; Su-Kim, I.; Choi, D.K. Aqueous extract of Dendropanax morbiferus leaves effectively alleviated neuroinflammation and behavioral impediments in MPTP-induced Parkinson’s mouse model. Oxid. Med. Cell. Longev. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Park, S.K.; Guo, T.J.; Kang, J.Y.; Ha, J.S.; Lee, D.S.; Lee, U.; Heo, H.J. Anti-amnesic effect of Dendropanax morbifera via JNK signaling pathway on cognitive dysfunction in high-fat diet-induced diabetic mice. Behav. Brain Res. 2016, 312, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, D.W.; Yoo, D.Y.; Jung, H.Y.; Nam, S.M.; Kim, J.W.; Hong, S.M.; Kim, D.W.; Choi, J.H.; Moon, S.M.; et al. Dendropanax morbifera Léveille extract facilitates cadmium excretion and prevents oxidative damage in the hippocampus by increasing antioxidant levels in cadmium-exposed rats. BMC Complement. Altern. Med. 2014, 14, 428. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Kim, S. Antioxidant and antithrombotic properties of Dendropanax morbifera Léveille (Araliaceae) and its ferments produced by fermentation processing. J. Food Biochem. 2019, 43, e13056. [Google Scholar] [CrossRef]
- Choo, G.S.; Lim, D.P.; Kim, S.M.; Yoo, E.S.; Kim, S.H.; Kim, C.H.; Woo, J.S.; Kim, H.J.; Jung, J.Y. Anti-inflammatory effects of Dendropanax morbifera in lipopolysaccharide-stimulated RAW264.7 macrophages and in an animal model of atopic dermatitis. Mol. Med. Rep. 2019, 19, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Chien, S.C.; Tseng, Y.H.; Hsu, W.N.; Chu, F.H.; Chang, S.T.; Kuo, Y.H.; Wang, S.Y. Anti-inflammatory and anti-oxidative activities of polyacetylene from Dendropanax dentiger. Nat. Prod. Commun. 2014, 9, 1589–1590. [Google Scholar] [CrossRef] [Green Version]
- Song, M.J.; Kim, H.; Heldenbrand, B.; Jeon, J.; Lee, S. Ethnopharmacological survey of medicinal plants in Jeju Island, Korea. J. Ethnobiol. Ethnomed. 2013, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Shim, H.J.; Park, S.; Lee, J.W.; Park, H.J.; Baek, S.H.; Kim, E.K.; Yu, S.W. Extracts from Dendropanax morbifera Leaves Have Modulatory Effects on Neuroinflammation in Microglia. Am. J. Chin. Med. 2016, 44, 119–132. [Google Scholar] [CrossRef]
- Hyun, T.K.; Kim, M.O.; Lee, H.; Kim, Y.; Kim, E.; Kim, J.S. Evaluation of anti-oxidant and anti-cancer properties of Dendropanax morbifera Léveille. Food Chem. 2013, 141, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.I. Antidiabetic effects of dendropanoxide from leaves of Dendropanax morbifera Leveille in normal and streptozotocin-induced diabetic rats. Hum. Exp. Toxicol. 2011, 30, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Lee, J.S.; Akram, M.; Kim, K.A.; Shin, Y.J.; Yu, J.H.; Bae, O.N. Protective activity of Dendropanax morbifera against cisplatin-induced acute kidney injury. Kidney Blood Press. Res. 2015, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, D.W.; Yoo, D.Y.; Jung, H.Y.; Kim, J.W.; Kim, D.W.; Choi, J.H.; Moon, S.M.; Yoon, Y.S.; Hwang, I.K. Antioxidant effects of Dendropanax morbifera Léveille extract in the hippocampus of mercury-exposed rats. BMC Complement. Altern. Med. 2015, 15, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youn, J.S.; Kim, Y.J.; Na, H.J.; Jung, H.R.; Song, C.K.; Kang, S.Y.; Kim, J.Y. Antioxidant activity and contents of leaf extracts obtained from Dendropanax morbifera LEV are dependent on the collecting season and extraction conditions. Food Sci. Biotechnol. 2019, 28, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.; Kim, K.A.; Kim, E.S.; Syed, A.S.; Kim, C.Y.; Lee, J.S.; Bae, O.N. Potent Anti-inflammatory and Analgesic Actions of the Chloroform Extract of Dendropanax morbifera Mediated by the Nrf2/HO-1 Pathway. Biol. Pharm. Bull. 2016, 39, 728–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birhanu, B.T.; Kim, J.Y.; Hossain, M.A.; Choi, J.W.; Lee, S.P.; Park, S.C. An in vivo immunomodulatory and anti-inflammatory study of fermented Dendropanax morbifera Léveille leaf extract. BMC Complement. Altern. Med. 2018, 18, 222. [Google Scholar] [CrossRef]
- Yoo, D.Y.; Jung, H.Y.; Kwon, H.J.; Kim, J.W.; Nam, S.M.; Chung, J.Y.; Choi, J.H.; Kim, D.W.; Yoon, Y.S.; Hwang, I.K. Effects of Dendropanax morbifera Léveille extract on hypothyroidism-induced oxidative stress in the rat hippocampus. Food Sci. Biotechnol. 2016, 25, 1761–1766. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, K.S.; An, H.K.; Kim, C.H.; Moon, H.I.; Lee, Y.C. Dendropanoxide induces autophagy through ERK1/2 activation in MG-63 human osteosarcoma cells and autophagy inhibition enhances dendropanoxide-induced apoptosis. PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef]
- Sachan, R.; Kundu, A.; Dey, P.; Son, J.Y.; Kim, K.S.; Lee, D.E.; Kim, H.R.; Park, J.H.; Lee, S.H.; Kim, J.H.; et al. Dendropanax morbifera protects against renal fibrosis in streptozotocin-induced diabetic rats. Antioxidants 2020, 9, 84. [Google Scholar] [CrossRef] [Green Version]
- An, N.Y.; Kim, J.E.; Hwang, D.; Ryu, H.K. Anti-diabetic effects of aqueous and ethanol extract of Dendropanax morbifera Leveille in streptozotocin-Induced diabetes model. J. Nutr. Heal. 2014, 47, 394–402. [Google Scholar] [CrossRef]
- Lee, C.; Yang, M.; Moon, J.O. Antioxidant and hepatoprotective effects of the ethanol extract of Dendropanax morbifera Leveille on the T-Butyl Hydroperoxide-Induced HepG2 cell damages. Korean J. Pharmacogn. 2019, 50, 32–36. [Google Scholar]
- Bae, D.; Kim, J.; Lee, S.Y.; Choi, E.J.; Jung, M.A.; Jeong, C.S.; Na, J.R.; Kim, J.J.; Kim, S. Hepatoprotective effects of aqueous extracts from leaves of Dendropanax morbifera leveille against alcohol-induced hepatotoxicity in rats and in vitro anti-oxidant effects. Food Sci. Biotechnol. 2015, 24, 1495–1503. [Google Scholar] [CrossRef]
- Song, J.H.; Kwak, S.; Kim, H.; Jun, W.; Lee, J.; Yoon, H.G.; Kim, Y.; Choi, K.C. Dendropanax morbifera Branch Water Extract Increases the Immunostimulatory Activity of RAW264.7 Macrophages and Primary Mouse Splenocytes. J. Med. Food 2019, 22, 1136–1145. [Google Scholar] [CrossRef]
- Kim, R.W.; Lee, S.Y.; Kim, S.G.; Heo, Y.R.; Son, M.K. Antimicrobial, antioxidant and cytotoxic activities of dendropanax morbifera léveille extract for mouthwash and denture cleaning solution. J. Adv. Prosthodont. 2016, 8, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.M.; Kim, M.Y.; Park, S.D.; Park, W.H.; Moon, H.I. In vitro evaluation of the antiplasmodial activity of Dendropanax morbifera against chloroquine-sensitive strains of Plasmodium falciparum. Phyther. Res. 2009, 23, 1634–1637. [Google Scholar] [CrossRef]
- Yun, J.W.; Kim, S.H.; Kim, Y.S.; Choi, E.J.; You, J.R.; Cho, E.Y.; Yoon, J.H.; Kwon, E.; Kim, H.C.; Jang, J.J.; et al. Preclinical study of safety of Dendropanax morbifera Leveille leaf extract: General and genetic toxicology. J. Ethnopharmacol. 2019, 238, 111874. [Google Scholar] [CrossRef]
- Wang, C.; Mathiyalagan, R.; Kim, Y.J.; Castro-Aceituno, V.; Singh, P.; Ahn, S.; Wang, D.; Yang, D.C. Rapid green synthesis of silver and gold nanoparticles using Dendropanax morbifera leaf extract and their anticancer activities. Int. J. Nanomed. 2016, 11, 3691. [Google Scholar]
- Chung, I.M.; Seo, S.H.; Kang, E.Y.; Park, S.D.; Park, W.H.; Moon, H.I. Chemical composition and larvicidal effects of essential oil of Dendropanax morbifera against Aedes aegypti L. Biochem. Syst. Ecol. 2009, 37, 470–473. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Lee, J.Y.; Kim, J.W.; An, Y.J. Effect of Leaf Extracts of Dendropanax morbifera on Selected Probiotics and Pathogenic Bacteria. FASEB J. 2017, 31, lb407. [Google Scholar]
- Kim, K.; Nguyen, V.B.; Dong, J.; Wang, Y.; Park, J.Y.; Lee, S.C.; Yang, T.J. Evolution of the Araliaceae family inferred from complete chloroplast genomes and 45S nrDNAs of 10 Panax-related species. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dilcher, D.L.; Dolph, G.E. Fossil Leaves of Dendropanax from Eocene Sediments of Southeastern North America. Am. J. Bot. 1970, 57, 153–160. [Google Scholar] [CrossRef]
- Ramos-Garza, J.; Rodríguez-Tovar, A.V.; Flores-Cotera, L.B.; Rivera-Orduña, F.N.; Vásquez-Murrieta, M.S.; Ponce-Mendoza, A.; Wang, E.T. Diversity of fungal endophytes from the medicinal plant Dendropanax arboreus in a protected area of Mexico. Ann. Microbiol. 2016, 66, 991–1002. [Google Scholar] [CrossRef]
- Fiaschi, P.; Jung-Mendaçolli, S.L. Three new species of Dendropanax Decne & Planch (Araliaceae) from São Paulo state, Brazil. Candollea 2006, 61, 457–466. [Google Scholar]
- Piedrahíta, Á.I.; II, P.P.L.; De Gracia Cruz, J.E. A New Species of Dendropanax (Araliaceae) of Restricted Range from the Caribbean Slope of Panama. Novon A J. Bot. Nomencl. 2015, 24, 165–169. [Google Scholar] [CrossRef]
- Oka, K.; Sai, O.F.; Yasuhara, T.A.; Sugimo, O.A.A. The major allergen of Dendropanax trifidus Makino. Contact Dermat. 1997, 36, 252–255. [Google Scholar] [CrossRef]
- Park, B.Y.; Min, B.S.; Oh, S.R.; Kim, J.H.; Kim, T.J.; Kim, D.H.; Bae, K.H.; Lee, H.K. Isolation and anticomplement activity of compounds from Dendropanax morbifera. J. Ethnopharmacol. 2004, 90, 403–408. [Google Scholar] [CrossRef]
- Kim, E.H.; Jo, C.S.; Ryu, S.Y.; Kim, S.H.; Lee, J.Y. Anti-osteoclastogenic diacetylenic components of Dendropanax morbifera. Arch. Pharm. Res. 2018, 41, 506–512. [Google Scholar] [CrossRef]
- Kim, J.Y.; Yoon, J.Y.; Sugiura, Y.; Lee, S.K.; Park, J.D.; Song, G.J.; Yang, H.J. Dendropanax morbiferus leaf extract facilitates oligodendrocyte development. R. Soc. Open Sci. 2019, 6, 190266. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.M.; Kim, S.H.; Kwon, C.; Kim, S.Y.; Yang, Y.J.; Kim, J.S.; Ali, M.; Ahmad, A. New chemical constituents from the bark of Dendropanax morbifera leveille and their evaluation of antioxidant activities. Molecules 2019, 24, 3967. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Kang, S.; Yang, S.Y.; Kim, Y.H.; Li, W. Chemical constituents from Dendropanax morbiferus H. Lév. Stems and leaves and their chemotaxonomic significance. Biochem. Syst. Ecol. 2019, 87, 103936. [Google Scholar] [CrossRef]
- Yang, H.Y.; Kim, K.S.; Lee, Y.H.; Park, J.H.; Kim, J.H.; Lee, S.Y.; Kim, Y.M.; Kim, I.S.; Kacew, S.; Lee, B.M.; et al. Dendropanax morbifera ameliorates thioacetamide-induced hepatic fibrosis via TGF-β1/smads pathways. Int. J. Biol. Sci. 2019, 15, 800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, I.M.; Song, H.K.; Kim, S.J.; Moon, H.I. Anticomplement activity of polyacetylenes from leaves of Dendropanax morbifera Leveille. Phyther. Res. 2011, 25, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.; Yun, J.J.; Jeong, J.E.; Wi, A.J.; Song, H. Inhibitory effects of nano-extract from Dendropanax morbifera on proliferation and migration of vascular smooth muscle cells. J. Nanosci. Nanotechnol. 2015, 15, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Lee, H.M.; Ha, J.H.; Jeon, S.H.; Park, S.N. Inhibitoiy effects of Dendropanax morbifera leaf extracts on melanogenesis through down-regulation of tyrosinase and TRP-2. Appl. Chem. Eng. 2014, 25, 468–473. [Google Scholar] [CrossRef]
- Shin, D.-C.; Kim, G.-C.; Song, S.-Y.; Kim, H.-J.; Yang, J.-C.; Kim, B.-A. Antioxidant and Antiaging Activities of Complex Supercritical Fluid Extracts from Dendropanax morbifera, Corni fructus and Lycii Fructus. Korea J. Herbol. 2013, 28, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Im, K.-J.; Jang, S.-B.; Yoo, D.-Y. Anti-cancer Effects of Dendropanax morbifera Extract in MCF-7 and MDA-MB-231 Cells. J. Orient. Obstet. Gynecol. 2015, 28, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.-G.; Kim, K.-O. Effect of Resina Dendropanacis morbiferus on Stress and Sleep Hormone in Chronic Mild Stress-Induced Rats. J Orient. Neuropsychiatry 2017, 28, 287–302. [Google Scholar]
- Research Report: Screening of Immune Activation Activities in the Leaves of Dendropanax morbifera Lev. Korean J. Med. Crop Sci. 2002, 10, 109–115.
- Jung, M.-A.; Oh, K.; Jin Choi, E.; Kim, Y.J.; Bae, D.; Oh, D.-R.; Man Kim, K.; Kim, D.-W.; Choi, C. Effect of Aqueous Extract of Dendropanax morbifera Leaf on Sexual Behavior in Male Rats. J. Food Nutr. Res. 2017, 5, 518–521. [Google Scholar] [CrossRef]
- Lee, S.Y.; Choi, E.-J.; Bae, D.-H.; Lee, D.-W.; Kim, S. Effects of 1-tetradecanol and β-sitosterol Isolated from Dendropanax morbifera Lev. on Skin Whitening, Moisturizing and Preventing Hair Loss. J. Soc. Cosmet. Sci. Korea. 2015, 41, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Kim, K.S.; Son, J.Y.; Kim, H.R.; Park, J.H.; Lee, S.H.; Lee, D.E.; Kim, I.S.; Lee, K.Y.; Lee, B.M.; et al. Protective effects of Dendropanax morbifera against cisplatin-induced nephrotoxicity without altering chemotherapeutic efficacy. Antioxidants 2019, 8, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.E.; Sapkota, K.; Choi, J.H.; Kim, M.K.; Kim, Y.H.; Kim, K.M.; Kim, K.J.; Oh, H.N.; Kim, S.J.; Kim, S. Rutin from Dendropanax morbifera leveille protects human dopaminergic cells against rotenone induced cell injury through inhibiting JNK and p38 MAPK signaling. Neurochem. Res. 2014, 39, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kwak, M.; Baek, S.H. Neuroprotective effects of Dendropanax morbifera leaves on glutamate-induced oxidative cell death in HT22 mouse hippocampal neuronal cells. J. Ethnopharmacol. 2020, 90, 403–408. [Google Scholar] [CrossRef]
- Lee, K.Y.; Jung, H.Y.; Yoo, D.Y.; Kim, W.; Kim, J.W.; Kwon, H.J.; Kim, D.W.; Yoon, Y.S.; Hwang, I.K.; Choi, J.H. Dendropanax morbifera Léveille extract ameliorates D-galactose-induced memory deficits by decreasing inflammatory responses in the hippocampus. Lab. Anim. Res. 2017, 33, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Park, S.Y.; Lee, S.H.; Kim, Y.; Kim, Y.J.; Jun, W.; Yoon, H.G. Ameliorative Effects of Dendropanax morbifera on Cognitive Impairment Via Enhancing Cholinergic Functions and Brain-Derived Neurotrophic Factor Expression in β-Amyloid-Induced Mice. J. Med. Food. 2019, 22, 587–593. [Google Scholar] [CrossRef] [Green Version]
- Aceituno, V.C.; Ahn, S.; Simu, S.Y.; Wang, C.; Mathiyalagan, R.; Yang, D.C. Silver nanoparticles from Dendropanax morbifera Léveille inhibit cell migration, induce apoptosis, and increase generation of reactive oxygen species in A549 lung cancer cells. Vitr. Cell. Dev. Biol. Anim. 2016, 52, 1012–1019. [Google Scholar] [CrossRef]
- Lee, J.W.; Park, C.; Han, M.H.; Hong, S.H.; Lee, T.K.; Lee, S.H.; Kim, G.Y.; Choi, Y.H. Induction of human leukemia U937 cell apoptosis by an ethanol extract of Dendropanax morbifera Lev. through the caspase-dependent pathway. Oncol. Rep. 2013, 30, 1231–1238. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS generation and antioxidant defense systems in normal and malignant cells. Oxid. Med. Cell. Longev. 2020, 2019, 1–17. [Google Scholar] [CrossRef]
- Balakrishnan, R.; Vijayraja, D.; Jo, S.H.; Ganesan, P.; Su-kim, I.; Choi, D.K. Medicinal profile, phytochemistry, and pharmacological activities of murraya koenigii and its primary bioactive compounds. Antioxidants 2020, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.S.; Yoo, D.Y.; Jung, H.Y.; Kim, D.W.; Hwang, I.K.; Lee, J.Y.; Moon, S.M. Effects of Dendropanax morbifera Léveille extracts on cadmium and mercury secretion as well as oxidative capacity: A randomized, double-blind, placebo-controlled trial. Biomed. Rep. 2016, 4, 623–627. [Google Scholar] [CrossRef] [Green Version]
- Ranjithkumar, R.; Alhadidi, Q.; Shah, Z.A.; Ramanathan, M. Tribulusterine Containing Tribulus terrestris Extract Exhibited Neuroprotection Through Attenuating Stress Kinases Mediated Inflammatory Mechanism: In Vitro and In Vivo Studies. Neurochem. Res. 2019, 44, 1228–1242. [Google Scholar] [CrossRef] [PubMed]
- Hyun, T.K.; Ko, Y.J.; Kim, E.H.; Chung, I.M.; Kim, J.S. Anti-inflammatory activity and phenolic composition of Dendropanax morbifera leaf extracts. Ind. Crops Prod. 2015, 74, 263–270. [Google Scholar] [CrossRef]
- Kim, G.; Kim, J.E.; Kang, M.J.; Jang, A.R.; Kim, Y.R.; Kim, S.; Chang, K.T.; Hong, J.J.; Park, J.H. Inhibitory effect of 1-tetradecanol on Helicobacter pylori-induced production of interleukin-8 and vascular endothelial growth factor in gastric epithelial cells. Mol. Med. Rep. 2017, 16, 9573–9578. [Google Scholar] [CrossRef]
- Yu, H.Y.; Kim, K.S.; Lee, Y.C.; Moon, H.I.; Lee, J.H. Oleifolioside A, a new active compound, attenuates LPS-stimulated iNOS and COX-2 expression through the downregulation of NF-κ B and MAPK Activities in RAW 264.7 Macrophages. Evidence-Based Complement. Altern. Med. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Park, S.K.; Guo, T.J.; Kang, J.Y.; Ha, J.S.; Lee, D.S.; Kwon, O.J.; Lee, U.; Heo, H.J. Neuronal cell protective effect of Dendropanax morbifera extract against high glucose-induced oxidative stress. J. Korean Soc. Food Sci. Nutr. 2016, 45, 938–947. [Google Scholar] [CrossRef]
- Kim, W.; Yim, H.S.; Yoo, D.Y.; Jung, H.Y.; Kim, J.W.; Choi, J.H.; Yoon, Y.S.; Kim, D.W.; Hwang, I.K. Dendropanax morbifera Léveille extract ameliorates cadmium-induced impairment in memory and hippocampal neurogenesis in rats. BMC Complement. Altern. Med. 2016, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Bae, D.H.; Lee, U.; Heo, H.J. Ameliorating effect of water extract from Dendropanax morbifera Lev. On memory dysfunction in streptozotocin-induced diabetic rats. Korean J. Food Sci. Technol. 2016, 48, 275–283. [Google Scholar] [CrossRef]
- Jung, H.Y.; Kwon, H.J.; Hahn, K.R.; Yoo, D.Y.; Kim, W.; Kim, J.W.; Kim, Y.J.; Yoon, Y.S.; Kim, D.W.; Hwang, I.K. Dendropanax morbifera Léveille extract ameliorates cesium-induced inflammation in the kidney and decreases antioxidant enzyme levels in the hippocampus. Mol. Cell. Toxicol. 2018, 14, 193–199. [Google Scholar] [CrossRef]
- Mehndiratta, S.; Kumar, S.; Meena, A.; Koul, S.; Suri, O.; Dhar, K. A Review on Plants a useful source of anti-cancer drugs. J Pharm Res. 2011, 4, 264–271. [Google Scholar]
- Ali, R.; Mirza, Z.; Ashraf, G.M.D.; Kamal, M.A.; Ansari, S.A.; Damanhouri, G.A.; Abuzenadah, A.M.; Chaudhary, A.G.; Sheikh, I.A. New anticancer agents: Recent developments in tumor therapy. Anticancer Res. 2012, 32, 2999–3005. [Google Scholar] [PubMed]
- Schneider-Stock, R.; Ghantous, A.; Bajbouj, K.; Saikali, M.; Darwiche, N. Epigenetic mechanisms of plant-derived anticancer drugs. Front. Biosci. 2012, 17, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.Y.; Yu, H.Y.; Park, C.; Han, M.H.; Hong, S.H.; Kim, K.S.; Lee, Y.C.; Chang, Y.C.; Cheong, J.; Moon, S.K.; et al. Oleifolioside B-mediated autophagy promotes apoptosis in A549 human non-small cell lung cancer cells. Int. J. Oncol. 2013, 43, 1943–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.Y.; Jin, C.Y.; Kim, K.S.; Lee, Y.C.; Park, S.H.; Kim, G.Y.; Kim, W.J.; Moon, H.I.; Choi, Y.H.; Lee, J.H. Oleifolioside A mediates caspase-independent human cervical carcinoma HeLa cell apoptosis involving nuclear relocation of mitochondrial apoptogenic factors AIF and EndoG. J. Agric. Food Chem. 2012, 60, 5400–5406. [Google Scholar] [CrossRef]
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes. 2015, 6, 598. [Google Scholar] [CrossRef]
- Zimmet, P.Z. Diabetes and its drivers: The largest epidemic in human history? Clin. Diabetes Endocrinol. 2017, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Taskinen, M.R. Diabetic dyslipidemia. Atheroscler. Suppl. 2002, 3, 47–51. [Google Scholar] [CrossRef]
- Ozsoy-Sacan, O.; Karabulut-Bulan, O.; Bolkent, S.; Yanardag, R.; Ozgey, Y. Effects of chard (Beta vulgaris L. var cicla) on the liver of the diabetic rats: A morphological and biochemical study. Biosci. Biotechnol. Biochem. 2004, 68, 1640–1648. [Google Scholar] [CrossRef] [Green Version]
- Heo, M.G.; Byun, J.H.; Kim, J.; Choung, S.Y. Treatment of Dendropanax morbifera leaves extract improves diabetic phenotype and inhibits diabetes induced retinal degeneration in db/db mice. J. Funct. Foods. 2018, 46, 136–146. [Google Scholar] [CrossRef]
- Manfo, F.P.T.; Nantia, E.A.; Kuete, V. Hepatotoxicity and Hepatoprotective Effects of African Medicinal Plants. In Toxicological Survey of African Medicinal Plants; Elsevier: Amsterdam, The Netherlands, 2014; pp. 323–355. [Google Scholar]
- Eom, T.; Kim, K.C.; Kim, J.S. Dendropanax morbifera leaf polyphenolic compounds: Optimal extraction using the response surface method and their protective effects against alcohol-induced liver damage. Antioxidants 2020, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Oberholzer, A.; Oberholzer, C.; Moldawer, L.L. Cytokine Signaling—Regulation of the immune response in normal and critically ill states. Crit. Care Med. 2000, 28, N3–N12. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Bajpai, V.K.; Kim, M. Plants as potential sources of natural immunomodulators. Rev. Environ. Sci. Biotechnol. 2014, 13, 17–33. [Google Scholar] [CrossRef]
- Patwardhan, B.; Gautam, M. Botanical immunodrugs: Scope and opportunities. Drug Discov. Today 2005, 10, 495–502. [Google Scholar] [CrossRef]
- Brindha, P.; Venkatalakshmi, P.; Vadivel, V. Role of phytochemicals as immunomodulatory agents: A review. Int. J. Green Pharm. 2016, 10, 1–18. [Google Scholar]
- Choi, J.M.; Kim, H.J.; Lee, K.Y.; Choi, H.J.; Lee, I.S.; Kang, B.Y. Increased IL-2 production in T cells by xanthohumol through enhanced NF-AT and AP-1 activity. Int. Immunopharmacol. 2009, 9, 103–107. [Google Scholar] [CrossRef]
- Knight, J.A. Review: Free radicals, antioxidants, and the immune system. Ann. Clin. Lab. Sci. 2000, 30, 145–158. [Google Scholar]
- Park, J.U.; Kang, B.Y.; Kim, Y.R. Ethyl Acetate Fraction from Dendropanax morbifera Leaves Increases T Cell Growth by Upregulating NF-AT-Mediated IL-2 Secretion. Am. J. Chin. Med. 2018, 46, 453–467. [Google Scholar] [CrossRef]
- Harris, C.L.; Pouw, R.B.; Kavanagh, D.; Sun, R.; Ricklin, D. Developments in anti-complement therapy; from disease to clinical trial. Mol. Immunol. 2018, 102, 89–119. [Google Scholar] [CrossRef]
- Seongmin, K.I.M.; Park, S.G.Y.U.; Song, Y.J.A.E.; Park, J.K.O.O.; Choi, C.H.E.E.; Sanghoon, L.E.E.; Hoffman, R.M. Analysis of anticancer activity and chemical sensitization effects of Dendropanax morbifera and commersonia bartramia extracts. Anticancer Res. 2018, 38, 3853–3861. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial. No | Compound | Molecular Formula | Plant | Plant Part | Extraction Method | Content in the Extract | References |
|---|---|---|---|---|---|---|---|
| D. morbifera | |||||||
| 1 | (9Z,16S)-16-hydroxy-9,17-octadecadiene-12,14-diynoic acid | C18H24O3 | D. morbifera | Leaves | Methanol | 1.1 g/1.1 kg | [47] |
| 2 | Neochlorogenic acid | C16H18O9 | D. morbifera | Leaf and stems | Water | 656 mg/100 g | [30] |
| 3 | Stigmasterol | C29H48O | D. morbifera | Aerial parts | Methanol | 490.7 mg/5 kg | [48] |
| 4 | β-Amyrin | C30H50O | D. morbifera | Aerial parts | Methanol | 150 mg/1.1 kg | [47] |
| 5 | Epifriedelanol | C30H52O | D. morbifera | Aerial parts | Methanol | 120 mg/5 kg | [48] |
| 6 | 1-Tetradecanol | C14H30O | D. morbifera | Stems and leaves | Methanol | 98.6 mg/2.5 kg | [51] |
| 7 | Quercetin 3β-gentiobioside | C27H30O17 | D. morbifera | Stems and leaves | Methanol | 80 mg/2.5 kg | [51] |
| 8 | α-Glutinol | C30H50O | D. morbifera | Leaves | Methanol | 49 mg/1.1 kg | [47] |
| 9 | 1-cyclohex-20,50-dienyl 1-cyclohexylethanol-O-β-d-xylopyranoside | C19H31O5 | D. morbifera | Dried barks | Acetone | 41 mg/558 g | [50] |
| 10 | Glyceryl-1, 3-dipalmito-2-olein | C53H100O6 | D. morbifera | Dried barks | Acetone | 41 mg/558 g | [50] |
| 11 | Oleyl-O-β-d-xyloside | C23H43O6 | D. morbifera | Dried barks | Acetone | 38 mg/558 g | [50] |
| 12 | Cis-6-oxogeran-4-enyl-10-oxy-O-β-arabinopyranosyl-4′-O-β-arabinopyranosyl-2”-octadec-9”’,12”’,15”’-trienoate | C38H63O11 | D. morbifera | Dried barks | Acetone | 32 mg/558 g | [50] |
| 13 | Linolenyl-O-β-D-arabinofuranoside | C23H38O6 | D. morbifera | Dried barks | Acetone | 31 mg/558 g | [50] |
| 14 | n-tetradecanyl oleate | C32H63O2 | D. morbifera | Dried barks | Acetone | 30 mg/558 g | [50] |
| 15 | Icariside D1 | C23H26O10 | D. morbifera | Stems and leaves | Methanol | 30 mg/2.5 kg | [51] |
| 16 | α-Amyrin | C30H50O | D. morbifera | Aerial parts | Methanol | 30 mg/1.1 kg | [47] |
| 17 | Geranilan-8-oxy-O-α-d-xylopyranosyl-20-n-octadec-9”,12”,15”-trienoate | C33H59O6 | D. morbifera | Dried barks | Acetone | 29 mg/558 g | [50] |
| 18 | Geran-3(10)-enyl-1-oxy-O-β-arabinopyranosyl-4′-O-β-arabinopyranosyl-2”-octadec-9”’, 12”’,15”’-trienoate | C38H65O10 | D. morbifera | Dried barks | Acetone | 28 mg/558 g | [50] |
| 19 | Guaiacol-O-β-D-arabinopyaranoside | C12H17O6 | D. morbifera | Dried barks | Acetone | 26 mg/558 g | [50] |
| 20 | β-Sitosterol | C29H50O | D. morbifera | Aerial parts | Methanol | 25.6 mg/1.1 kg | [47] |
| 21 | trans-Phytol | C20H40O | D. morbifera | Leaves | Methanol | 22 mg/1.1 kg | [47] |
| 22 | n-octadec-9,12-dienoyl-O-β-D-arabinopyranoside | C23H40O6 | D. morbifera | Dried barks | Acetone | 21 mg/558 g | [50] |
| 23 | Citroside A | C19H30O8 | D. morbifera | Stems and leaves | Methanol | 20 mg/2.5 kg | [51] |
| 24 | Chlorogenic acid | C16H18O9 | D. morbifera | Branch | Methanol | 19.5 mg/g | [10] |
| 25 | Friedelin | C30H50O | D. morbifera | Aerial parts | Methanol | 14.5 mg/5 kg | [48] |
| 26 | (3S,8S)-falcarindiol | C17H24O2 | D. morbifera | Stems and leaves | Methanol | 13 mg/2.5 kg | [51] |
| 27 | Uridine | C9H12N2O6 | D. morbifera | Stems and leaves | Methanol | 11.7 mg/2.5 kg | [51] |
| 28 | Lutexin | C21H20O11 | D. morbifera | Stems and leaves | Methanol | 10.3 mg/2.5 kg | [51] |
| 29 | Dendropanoxide | C30H50O | D. morbifera | Aerial parts | Methanol | 10 mg/5 kg | [48] |
| 30 | Tetradecanol | C14H30O | D. morbifera | Stems and leaves | Methanol | 10 mg/2.5 kg | [51] |
| 31 | Rutin | C27H30O16 | D. morbifera | Dried aerial parts | Water | 6.38 mg/g | [13] |
| 32 | Nikoenoside | C16H24O9 | D. morbifera | Stems and leaves | Methanol | 5.2 mg/2.5 kg | [51] |
| 33 | Syringaresinol β-D-glucoside | C28H36O13 | D. morbifera | Stems and leaves | Methanol | 4.8 mg/2.5 kg | [51] |
| 34 | Isotachioside | C13H18O8 | D. morbifera | Stems and leaves | Methanol | 4.0 mg/2.5 kg | [51] |
| 35 | Scorzonoside | C27H34O12 | D. morbifera | Stems and leaves | Methanol | 3.7 mg/2.5 kg | [51] |
| 36 | Oleifolioside-B | C45H76O17 | D. morbifera | Stems and leaf | Methanol | 3 mg/0.3 g | [36] |
| 37 | Olean-12-en-3,24 β-diol | C30H50O2 | D. morbifera | Stems and leaves | Methanol | 3 mg/2.5 kg | [51] |
| 38 | Methyl chlorogenate | C17H20O9 | D. morbifera | Stems and leaves | Methanol | 2.7 mg/2.5 kg | [51] |
| 39 | Oleifolioside-A | C45H76O17 | D. morbifera | Stems and leaf | Methanol | 2.5 mg/0.3 g | [36] |
| 40 | (1R,2S)-1-(4-hydroxy-3-methoxyphenyl)-2-[4-[(1E)-3-hydroxy-1-propen-1-yl]-2-methoxyphenoxy]-1,3-propanediol | C20H24O7 | D. morbifera | Stems and leaves | Methanol | 2.4 mg/2.5 kg | [51] |
| 41 | Hyuganoside IIIa | C26H34O12 | D. morbifera | Stems and leaves | Methanol | 2.0 mg/2.5 kg | [51] |
| 42 | Rel-(1R,2R)-1-(4-hydroxy-3-methoxyphenyl)-2-[4-[(1E) -3-hydroxy-1-propen-1-yl]-2-methoxyphenoxy]-1,3-propanediol | C19H24O7 | D. morbifera | Stems and leaves | Methanol | 2.0 mg/2.5 kg | [51] |
| 43 | Protocatechuic acid | C7H6O4 | D. morbifera | Stems and leaves | Methanol | 1.8 mg/2.5 kg | [51] |
| 44 | Gastrodin | C13H18O7 | D. morbifera | Stems and leaves | Methanol | 1.8 mg/2.5 kg | [51] |
| 45 | Syringylglycerol 2-O-β-d-glucopyranoside | C28H36O13 | D. morbifera | Stems and leaves | Methanol | 1.4 mg/2.5 kg | [51] |
| 46 | Citroside B | C19H30O8 | D. morbifera | Stems and leaves | Methanol | 1.2 mg/2.5 kg | [51] |
| 47 | Nicotinic acid | C6H5NO2 | D. morbifera | Stems and leaves | Methanol | 1.2 mg/2.5 kg | [51] |
| 48 | Adenosine | C10H13N5O4 | D. morbifera | Stems and leaves | Methanol | 1.0 mg/2.5 kg | [51] |
| 49 | Thymidine | C10H14N2O5 | D. morbifera | Stems and leaves | Methanol | 1.0 mg/2.5 kg | [51] |
| 50 | Hesperidin | C28H34O15 | D. morbifera | Branch | Methanol | 1576.65 µg/g | [10] |
| 51 | Syringin | C17H24O9 | D. morbifera | Dried aerial parts | Water | 450.5 µg/g | [52] |
| 52 | Catechin | C15H14O6 | D. morbifera | Branch | Methanol | 359.07 µg/g | [10] |
| 53 | 2,5-Dihydroxybenzoic acid | C7H6O4 | D. morbifera | Branch | Methanol | 259.05 µg/g | [10] |
| 54 | Salicylic acid | C7H6O3 | D. morbifera | Branch | Methanol | 248.91 µg/g | [10] |
| 55 | Vitexin | C21H20O10 | D. morbifera | Branch | Methanol | 200 µg/g | [13] |
| 56 | Quercetin | C15H10O7 | D. morbifera | Branch | Water | 100 µg/g | [13] |
| 57 | Myricetin | C15H10O8 | D. morbifera | Branch | Methanol | 97.35 µg/g | [10] |
| 58 | 4-Hydroxybenzoic acid | C7H6O3 | D. morbifera | Branch | Methanol | 70.57 µg/g | [10] |
| 59 | Resveratrol | C14H12O3 | D. morbifera | Branch | Methanol | 66.23 µg/g | [10] |
| 60 | Tricin | C17H14O7 | D. morbifera | Branch | Methanol | 60 µg/g | [13] |
| 61 | Naringin | C27H32O14 | D. morbifera | Branch | Methanol | 53.19 µg/g | [10] |
| 66 | Syringic acid | C9H10O5 | D. morbifera | Branch | Methanol | 42.13 µg/g | [10] |
| 63 | Trans-Ferulic acid | C10H10O4 | D. morbifera | Branch | Methanol | 34.87 µg/g | [10] |
| 64 | Caffeic acid | C9H8O4 | D. morbifera | Branch | Methanol | 31.96 µg/g | [10] |
| 65 | Luteolin | C15H10O6 | D. morbifera | Branch | Methanol | 20 µg/g | [13] |
| 66 | Kaempferol | C15H10O6 | D. morbifera | Branch | Methanol | 20 µg/g | [13] |
| 67 | Gallic acid | C7H6O5 | D. morbifera | Branch | Methanol | 17.37 µg/g | [10] |
| 68 | p-Coumaric acid | C9H8O3 | D. morbifera | Branch | Methanol | 2.11 µg/g | [10] |
| 69 | trans-Cinnamic acid | C9H8O2 | D. morbifera | Branch | Methanol | 0.61 µg/g | [10] |
| 70 | Luteolin-7-O-rutinoside | C27H30O15 | D. morbifera | Roots | Methanol | N/A | [14] |
| 71 | Isoorientin | C21H20O11 | D. morbifera | Roots | Methanol | N/A | [14] |
| 72 | Orientin | C21H20O11 | D. morbifera | Roots | Methanol | N/A | [14] |
| 73 | Carnosol | C20H26O4 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 74 | Dextromethorphan | C18 H25 N O | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 75 | Cannabidiol | C21 H30 O2 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 76 | (−)-Bremazocine | C20 H29 N O2 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 77 | Doxapram | C24 H30 N2 O2 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 78 | Resolvin D2 | C22 H32 O5 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 79 | Procyclidine | C19 H29 N O | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 80 | 2-arachidonoylglycerol | C23 H38 O4 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| 81 | Eplerenone | C24 H30 O6 | D. morbifera | Leaves | Ethanol | N/A | [49] |
| Compound | Molecular Structure | Plant Species | Activity Tested |
|---|---|---|---|
| Quercetin |  | D. morbifera | Antioxidant, anti-inflammatory and neuroprotective |
| Chlorogenic acid |  | D. morbifera | Antioxidant and anti-inflammatory |
| Rutin |  | D. morbifera | Antioxidant, anti-inflammatory and neuroprotective |
| Carnosol |  | D. morbifera | Anti-inflammatory |
| Dextromethorphan |  | D. morbifera | Antioxidant, anti-inflammatory and neuroprotective |
| Cannabidiol |  | D. morbifera | Antioxidant, anti-inflammatory and neuroprotective |
| (−)-Bremazocine |  | D. morbifera | κ-opioid agonist |
| Doxapram |  | D. morbifera | Respiratory stimulant |
| Resolvin D2 |  | D. morbifera | Anti-inflammatory |
| Procyclidine |  | D. morbifera | Anticholinergic agent |
| 2-arachidonoylglycerol |  | D. morbifera | Cannabimimetic activity |
| Eplerenone |  | D. morbifera | Prevention of heart failure and mild symptoms |
| Cis-6-oxogeran-4-enyl-10-oxy-O-β-arabinopyranosyl -4′-O-β-arabinopyranosyl-2”-octadec-9”’,12”’,15”’-trienoate |  | D. morbifera | Antioxidant |
| Geran-3(10)-enyl-1-oxy-O-β-arabinopyranosyl -4′-O-β-arabinopyranosyl-2”-octadec-9”’, 12”’,15”’-trienoate |  | D. morbifera | Antioxidant |
| Geranilan-8-oxy-O-α-d-xylopyranosyl -20-n-octadec-9”,12”,15”-trienoate |  | D. morbifera | Antioxidant |
| 1-cyclohex-20,50-dienyl 1-cyclohexylethanol -O-β-D-xylopyranoside |  | D. morbifera | Antioxidant |
| (9Z,16S)-16-Hydroxy-9,17-octadecadiene -12,14-diynoic acid |  | D. morbifera | Anti-complement activity |
| Dendropanoxide |  | D. morbifera | Anti-complement activity and anti-osteoclastogenic activity |
| α-Glutinol |  | D. morbifera | Anti-complement activity |
| β-Amyrin |  | D. morbifera | Anti-complement activity and anti-osteoclastogenic activity |
| α-Amyrin |  | D. morbifera | Anti-complement activity |
| Trans-Phytol |  | D. morbifera | Anti-complement activity |
| β-Sitosterol |  | D. morbifera | Anti-complement activity |
| Friedelin |  | D. morbifera | Anti-osteoclastogenic activity |
| Epifriedelanol |  | D. morbifera | Anti-osteoclastogenic activity |
| Gallic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| 2,5-Dihydroxybenzoic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Catechin |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| 4-Hydroxybenzoic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Caffeic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Syringic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| p-Coumaric acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Trans-Ferulic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Salicylic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Hesperidin |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Naringin |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Resveratrol |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Myricetin |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Trans-Cinnamic acid |  | D. morbifera | Antioxidant, anti-inflammatory and anti-cancer activity |
| Activity Tested | Extract | Plant Part | Model | Major Effects | Reference |
|---|---|---|---|---|---|
| Antioxidant | Ethanol | Boughs | DPPH-radical scavenging assays | -D. morbifera extracts showed similar antioxidant capacities (82.92 ± 0.49%) to that of the vitamin C (90.11 ± 0.13%) positive control at the same concentration of 500 µg/mL | [35] |
| Antioxidant | Fermented | Leaves | DPPH-radical scavenging assays | -Exhibited activities with IC50 value of 65.30 µg/mL | [16] |
| Antioxidant | Methanol | Leaves, debarked stems, and bark | DPPH-radical scavenging assays | -Exhibited activities with RC50 values of 16.7 µg/mL for debarked stem extracts, 31.6 µg/mL for yellow leaves extract, and 37.8 µg/mL green leaf extracts | [21] |
| Antioxidant | Hot water and ethanol | Leaves | ABTS-radical scavenging assays | -Exhibited activities with IC50 value of 3.79 mg/mL for hot water, 3.75 mg/mL for 30% ethanol, and 3.58 mg/mL for 60% ethanol | [25] |
| Antioxidant | Fermented | Leaves | Hydroxyl-radical scavenging activity | -Exhibited activities with IC50 value of 57.52 µg/mL | [50] |
| Antioxidant | Water | Stem and leaves | Sprague–Dawley rats | -D. morbifera extracts significant enhancements in GSH, SOD, and CAT activities and a reduction in the MDA level | [30] |
| Antioxidant | Water | Aerial part | Sprague–Dawley rats | -D. morbifera extracts significantly decreased the hepatic MDA content and ameliorated the SOD and GSH content. | [52] |
| Anti-inflammatory | Aqueous | Leaves | BV-2 cells | -D. morbifera extracts effectively attenuates NO production, cell viability, and proinflammatory mediators, and subsequently suppressed the phosphorylation of both the IκB-α and NF-κB p65 subunit and MAPK signaling | [13] |
| Anti-inflammatory | Ethanol | Leaves | RAW264.7 macrophages | -Doses of 200 and 400 µg/mL effectively inhibited the activity of inflammatory mediators NO, TNF-α, and IL-6 | [17] |
| Anti-inflammatory | Ethyl acetate | Leaves | BV-2 cells | -D. morbifera treatment significantly attenuated the activation of MAPKs and NF-κB, and subsequently upregulated M2 polarization of alternative anti-inflammatory markers | [20] |
| Anti-inflammatory | Methanol | Leaves | RAW264.7 macrophages | -D. morbifera extracts significantly and dose-dependently reduced the production of NO and PGE2 and significantly inhibited protein and mRNA expression in COX-2 and iNOS activities and could modulate NF-κB and MAPK signaling -D. morbifera extracts significantly induced Nrf2 nuclear translocation and thereby induced HO-1 expression | [26] |
| Anti-inflammatory | Methanol | Leaves | Ear edema mouse model | Treatment with-D. morbifera extracts triggered a protective effect against the increase in ear thickness induced by TPA Treatment with-D. morbifera reversed ear edema and epidermal hyperproliferation, and increased the number of neutrophils induced by TPA | [26] |
| Anti-inflammatory | Fermented | Leaves | BALB/C mice | -D. morbifera extracts at 125, 250, and 500 mg/kg reduced levels of TNF-α, IL-2, IL-4, IL-5, IL-6, IL-10, IL-12, IL-12p70, IL-13 and IFN-γ in immunized BALB/C mice | [27] |
| Anti-inflammatory | Water | Stem and leaves | Sprague–Dawley rats | -D. morbifera extracts administered at 25 mg/kg markedly inhibited inflammatory cytokines and TGF-β1 expression in diabetic rats | [30] |
| Anti-inflammatory | Water | Aerial parts | Sprague–Dawley rats | - D. morbifera extracts at 25 mg/kg showed significant anti-inflammatory effects by reducing the levels of inflammatory cytokines such as TNF-α, IL-1β, and IL-6, and reversed IL-10 levels in a cisplatin-induced rat model | [62] |
| Anti-amnesic | Ethanol | Leaves | PC12 and MC-ⅨC cells | -D. morbifera extracts treatment effectively inhibited the AChE as an ACh-hydrolyzing enzyme in high glucose-induced PC12 and MC-ⅨC cells | [14] |
| Neuroprotective | Ethanol | Leaves | SH-SY5Y cells | -Rutin, a bioflavonoid isolated from D. morbifera, protected the higher level of intracellular Ca2+ and depleted the level of MMP, as well as subsequently decreased rotenone-induced ROS generation -Rutin prevented a decrease the levels of Bax/Bcl-2 ratio, caspase-9, and caspase-3 | [63] |
| Neuroprotective | Ethyl acetate | Leaves | HT22 cells | -D. morbifera extracts significantly inhibited mitochondrial dysfunction and the elevation of Ca2+ levels and reversed subsequent AIF nuclear translocation in glutamate-induced HT22 mouse hippocampal neuronal cells | [64] |
| Neuroprotective | Ethanol | Stem | Sprague–Dawley rats | -D. morbifera administration ameliorates cognitive dysfunction via an increase in cell proliferation, neuroblast differentiation, and AChE activity in the hippocampus induced by cadmium-induced neurotoxicity | [14] |
| Neuroprotective | Water | Leaves | Sprague–Dawley rats | -D. morbifera extracts significantly reduced mercury levels in the hippocampus and ROS generation and reversed hippocampal activities in dimethylmercury-induced rats | [24] |
| Neuroprotective | Ethanol | Stems | Sprague–Dawley rats | -D. morbifera administration changes serum triiodothyronine (T3), thyroxine (T4), and thyroid-stimulating hormone levels in the hippocampus induced by hypothyroidism neurotoxicity | [28] |
| Neuroprotective | Ethanol | Leaves | C57BL/6 mice | -D. morbifera administration significantly improved D-galactose-induced reduction in microglial activation, escape latency, swimming speed, and spatial preference behavior | [65] |
| Neuroprotective | Aqueous | Leaves | C57BL/6 mice | -D. morbifera treatment effectively improved behavioral function, and protected dopaminergic neuronal loss by restoring TH levels in the brain tissue of MPTP-induced PD mice | [13] |
| Neuroprotective | Ethanol | Leaves | C57BL/6 mice | -Ethanol extracts of D. morbifera significantly improved glucose tolerance status, and behavioral impairments, and significantly protects the abnormal activity of mitochondria by inhibiting phosphorylated p-JNK, p-IRS, p-Akt, and p-tau in high-fat diet-induced mice | [66] |
| Anti-cancer | Methanol | Leaves and debarked stems | Huh-7 cells | -D. morbifera extracts showed strong induction of p53, and p16 inhibited the activation of ERK and reduced Akt levels and the suppression of Huh-7 cell proliferation | [21] |
| Anti-cancer | Silver nanoparticles | Leaves | A549 and HepG2 | -Silver nanoparticles synthesized from D. morbifera enhanced ROS production in both cell lines and the modification of EGFR/p38 MAPK signaling | [67] |
| Anti-cancer | Ethanol | Stem bark | U937 cells | -D. morbifera-induced apoptosis in U937 cells was associated with the activation of caspase-8, -9, and -3 and downregulation of anti-apoptotic IAP family proteins | [68] |
| Anti-cancer | Water | Aerial parts | Sprague–Dawley rats | -D. morbifera administration protects against kidney damage induced by CDDP in tumor models | [62] |
| Anti-diabetes | Water | Leaves | 3T3-L1 cells | -D. morbiferus treatment reduced intracellular triglyceride levels and glucose uptake by lowering protein and mRNA expression levels of adipogenesis-related genes | [9] |
| Anti-diabetes | Water | Leaves and stem | Sprague–Dawley rats | -D. morbifera administration protected body and organ weight loss, significantly increased BUN, and significantly reduced KIM-1, SBP1, and PKM2 levels in the urinary excretions of diabetic rats | [30] |
| Anti-diabetes | Methanol | Leaves | Sprague–Dawley rats | -D. morbifera administration showed significant hypoglycemic activity by decreasing the total cholesterol, serum glucose, urea, triglycerides, creatinine, uric acid, alanine aminotransferase (ALT), and aspartate aminotransferase (AST) levels in streptozotocin-induced diabetic rats | [22] |
| Anti-diabetes | Ethanol | Leaves | C57BL/6 | -D. morbifera treatment protected against high-fat diet-induced abnormal mitochondrial activity and improved p-JNK, p-IRS, p-Akt, and p-tau in high-fat diet-induced diabetic mice | [14] |
| Anti-diabetes | Water and ethanol | Leaves and stem | ICR mice | -D. morbifera administration maintained a high level of body weight and increased insulin secretion by reducing the glucose concentration in the blood in streptozotocin-induced diabetic mice | [31] |
| Hepatoprotective | Ethanol | Root, leaves and stem | HepG2 cells | -D. morbifera exhibited strong antioxidant activity, and showed hepatoprotective activity against t-butyl hydroperoxide-induced HepG2 cells | [32] |
| Hepatoprotective | Aqueous | Leaves | Sprague–Dawley rats | -D. morbifera administration prevented ethanol-induced hepatotoxicity due to reductions of serum aspartate aminotransferase and alanine aminotransferase levels, and maintained enzymatic oxidant status, and suppressed cytochrome P-450 2E1 expression | [33] |
| Immunomodulatory | Fermented | Leaves | BALB/C mice | -D. morbifera administration showed an increase in spleen cells and CD8a+, CD11b, and CD3+ T-cell expression, and downregulated the IgG super-family | [27] |
| Immunomodulatory | Ethanol | Leaves, branch, sapling, and mixed | BALB/c mice | -D. morbifera administration significantly increased splenocyte cytokines, NO production, and LDH, and enhances innate immunity by modulator NF-κB signaling | [34] |
| Antimicrobial | Ethanol | Leaves | Paper disc test | With-D. morbifera extract concentrations of 40, 80, and 100 µg/mL, a 3.0 mm suggesting higher antibiotic effects against S. mutans and C. albicans | [35] |
| Antiplasmodial | Methanol | Stem bark | Semi-automated micro-dilution assay | -Extracts exhibited activities with IC50 values of 6.2 µg/mL and 5.3 µg/mL, against D10 | [36] |
| Anticomplementary | Aqueous | Leaves | Classical and alternative pathway assay | -D. morbifera exhibited significant inhibitory activity against complementary system with IC50 values of 87.3 mM for (3S)-falcarinol, 15.2 mM for (3S,8S)-falcarindiol, and 39.8 mM for (3S)-diynene. | [53] |
| Anticomplementary | Methanol | Leaves | Classical pathway assay | -(9Z,16S)-16-hydroxy-9,17-octadecadiene-12,14-diynoic acid from D. morbifera exhibited activities with an IC50 value of 56.98 µM. | [47] |
| Cytotoxicity | Methanol | Leaves | MTT assay | -Extracts exhibited cytotoxic activities greater than 93% at doses of more than 100 μg/mL, and 6 to 11% at doses of less than 50 μg/mL | [35] |
| Cytotoxicity | Silver nanoparticles | Leaves | MTT assay | -D-AgNPs at 100 µg/mL showed potent cytotoxicity after 48 hours | [38] |
| Cytotoxicity | Methanol | Leaves | MTT assay | -Extracts exhibited low cytotoxicity with IC50 values exceeding 50 µg/mL, maintaining up to 80% cell viability induced by glutamate toxicity | [64] |
| Cytotoxicity | Aqueous | Leaves | MTT assay | -Extracts with a high concentration of D. morbifera of 500 μg/mL showed no toxic effects and maintained up to 100% cell viability induced by LPS toxicity | [13] |
| Toxicity | Water | Leaves | Sprague–Dawley rats | -No deaths were observed at the highest concentration tested, and the LD50 values for both extracts was above 2000 mg/kg body weight | [37] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balakrishnan, R.; Cho, D.-Y.; Su-Kim, I.; Choi, D.-K. Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology. Antioxidants 2020, 9, 962. https://doi.org/10.3390/antiox9100962
Balakrishnan R, Cho D-Y, Su-Kim I, Choi D-K. Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology. Antioxidants. 2020; 9(10):962. https://doi.org/10.3390/antiox9100962
Chicago/Turabian StyleBalakrishnan, Rengasamy, Duk-Yeon Cho, In Su-Kim, and Dong-Kug Choi. 2020. "Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology" Antioxidants 9, no. 10: 962. https://doi.org/10.3390/antiox9100962
APA StyleBalakrishnan, R., Cho, D. -Y., Su-Kim, I., & Choi, D. -K. (2020). Dendropanax Morbiferus and Other Species from the Genus Dendropanax: Therapeutic Potential of Its Traditional Uses, Phytochemistry, and Pharmacology. Antioxidants, 9(10), 962. https://doi.org/10.3390/antiox9100962