Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Experimental Design
2.3. Electrocardiogram (ECG) Recordings
2.4. Sample Collection
2.5. Infarct Size Determination
2.6. Estimation of Antioxidant Markers, ATP, and Ca2+ in Heart Mitochondrial Fraction
2.7. Gene Expression Analysis
2.8. Western Blotting
2.9. Assessment of Inflammatory and Apoptotic Markers
2.10. Histopathological and Immunohistochemical Assays
2.11. Statistical Analysis
3. Results
3.1. Geraniol Reversed Electrocardiographic Alterations
3.2. Geraniol Effect on Infarct Area Size and Heart-to-Body Ratio
3.3. Geraniol Effect on Cardiac Enzymes
3.4. Geraniol Effect on Mitochondrial Antioxidant Activities
3.5. Effects of Geraniol on the Keap1/Nrf2/HO-1 Pathway
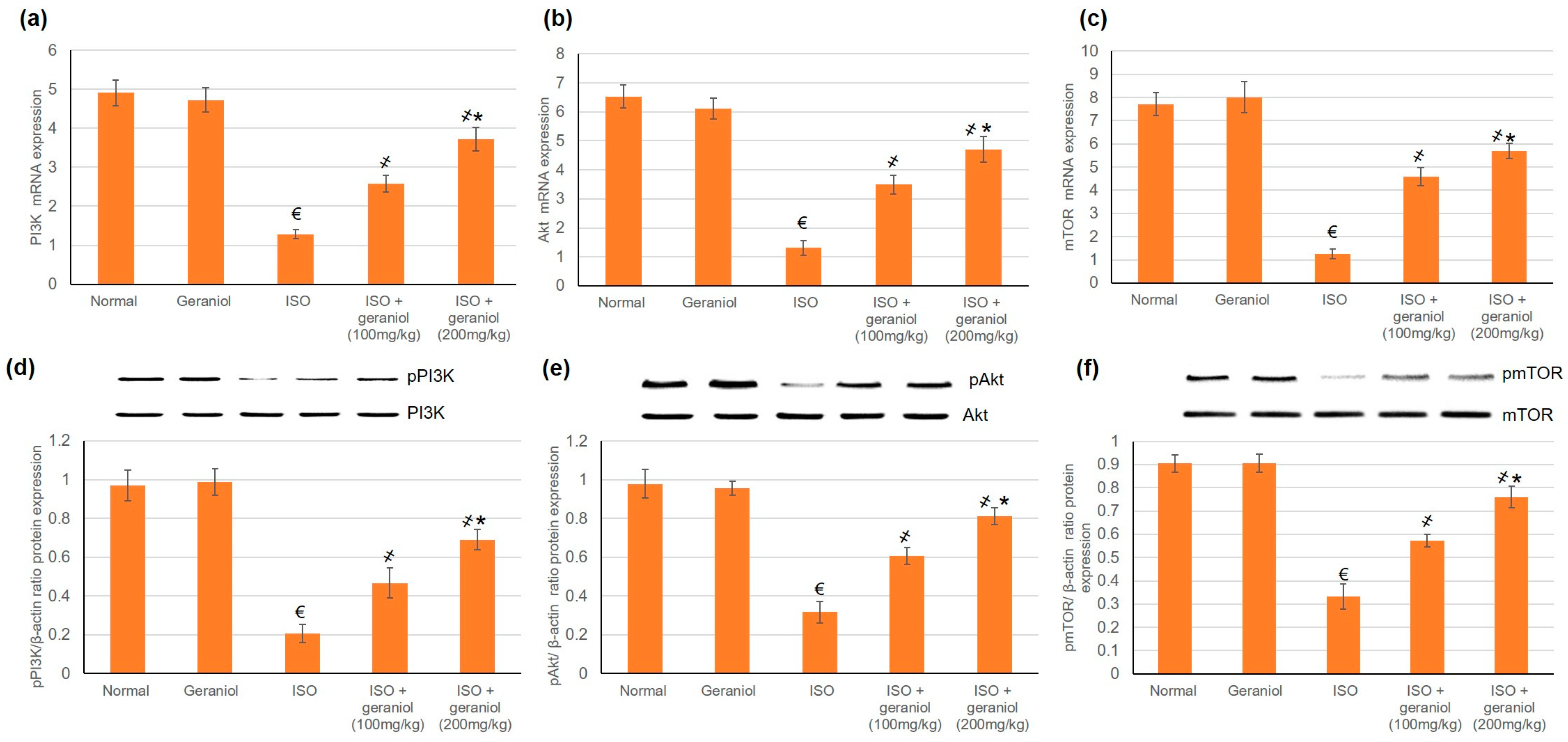
3.6. Effects of Geraniol on the PI3k/Akt/mTOR Pathway
3.7. Geraniol Pretreatement Inhibited Inflammatory Markers
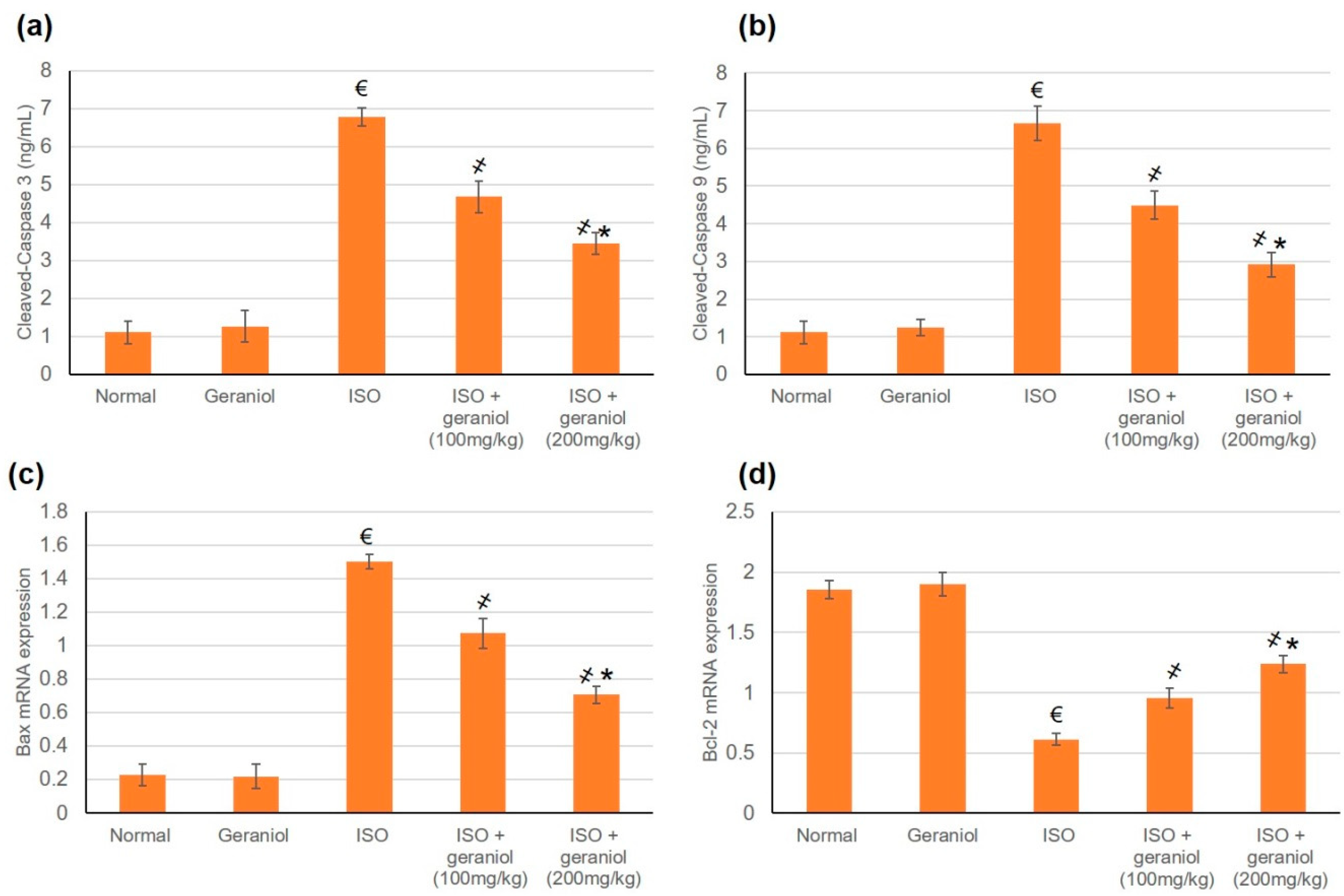
3.8. Effects of Geraniol on the Apoptotic Status
3.9. Histopathological and Immunohistochemical Assays
3.10. Effects of Geraniol on Heart Mitochondrial Ca2+ and ATP Content
4. Discussion
4.1. Cardioprotective Effects of Geraniol
4.2. Geraniol and Oxidative Stress: The Keap1/Nrf2 Pathway
4.3. Geraniol and Autophagy: The PI3K, Akt, mTOR Pathway
4.4. Geraniol and Inflammation
4.5. Geraniol and Apoptosis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crespo, R.; Wei, K.; Rodenak-Kladniew, B.; Mercola, M.; Ruiz-Lozano, P.; Hurtado, C. Effect of geraniol on rat cardiomyocytes and its potential use as a cardioprotective natural compound. Life Sci. 2017, 172, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Yang, H.; Li, M.; Ding, M. A possible underlying mechanism behind the cardioprotective efficacy of tangeretin on isoproterenol triggered cardiotoxicity via modulating PI3K/Akt signaling pathway in a rat model. J. Food Biochem. 2020, 44, e13368. [Google Scholar] [CrossRef]
- Sun, G.Z.; Meng, F.J.; Cai, H.Q.; Diao, X.B.; Zhang, B.; Bai, X.P. Ginsenoside Rg3 protects heart against isoproterenol-induced myocardial infarction by activating AMPK mediated autophagy. Cardiovasc. Diagn. Ther. 2020, 10, 153–160. [Google Scholar] [CrossRef]
- Kocak, C.; Kocak, F.E.; Akcilar, R.; Isiklar, O.O.; Kocak, H.; Bayat, Z.; Simsek, H.; Taser, F.; Altuntas, I. Molecular and biochemical evidence on the protective effects of embelin and carnosic acid in isoproterenol-induced acute myocardial injury in rats. Life Sci. 2016, 147, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Song, F.; Duan, L.R.; Sheng, J.J.; Xie, Y.H.; Yang, Q.; Chen, Y.; Dong, Q.Q.; Zhang, B.L.; Wang, S.W. Paeonol and danshensu combination attenuates apoptosis in myocardial infarcted rats by inhibiting oxidative stress: Roles of Nrf2/HO-1 and PI3K/Akt pathway. Sci. Rep. 2016, 6, 23693. [Google Scholar] [CrossRef]
- Li, J.; Ichikawa, T.; Villacorta, L.; Janicki, J.S.; Brower, G.L.; Yamamoto, M.; Cui, T. Nrf2 protects against maladaptive cardiac responses to hemodynamic stress. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1843–1850. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, K.; Hu, P. The Role of Autophagy in Acute Myocardial Infarction. Front. Pharmacol. 2019, 10, 551. [Google Scholar] [CrossRef] [Green Version]
- Shu, L.; Zhang, W.; Huang, C.; Huang, G.; Su, G. Troxerutin Protects Against Myocardial Ischemia/Reperfusion Injury Via Pi3k/Akt Pathway in Rats. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 44, 1939–1948. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Z.; Fu, X.; Lin, Z.; Yu, K. Geraniol-mediated osteoarthritis improvement by down-regulating PI3K/Akt/NF-κB and MAPK signals: In vivo and in vitro studies. Int. Immunopharmacol. 2020, 86, 106713. [Google Scholar] [CrossRef]
- El-Bassossy, H.M.; Ghaleb, H.; Elberry, A.A.; Balamash, K.S.; Ghareib, S.A.; Azhar, A.; Banjar, Z. Geraniol alleviates diabetic cardiac complications: Effect on cardiac ischemia and oxidative stress. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 88, 1025–1030. [Google Scholar] [CrossRef]
- Lei, Y.; Fu, P.; Jun, X.; Cheng, P. Pharmacological Properties of Geraniol—A Review. Planta Med. 2019, 85, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Menezes-Filho, J.E.; Gondim, A.N.; Cruz, J.S.; de Souza, A.A.; Santos, J.N.; Conde-Garcia, E.A.; de Sousa, D.P.; Santos, M.S.; de Oliveira, E.D.; de Vasconcelos, C.M. Geraniol blocks calcium and potassium channels in the mammalian myocardium: Useful effects to treat arrhythmias. Basic Clin. Pharmacol. Toxicol. 2014, 115, 534–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekha, K.R.; Inmozhi Sivakamasundari, R. Geraniol Protects Against the Protein and Oxidative Stress Induced by Rotenone in an In Vitro Model of Parkinson’s Disease. Neurochem. Res. 2018, 43, 1947–1962. [Google Scholar] [CrossRef]
- La Rocca, V.; da Fonsêca, D.V.; Silva-Alves, K.S.; Ferreira-da-Silva, F.W.; de Sousa, D.P.; Santos, P.L.; Quintans-Júnior, L.J.; Leal-Cardoso, J.H.; de Almeida, R.N. Geraniol Induces Antinociceptive Effect in Mice Evaluated in Behavioural and Electrophysiological Models. Basic Clin. Pharmacol. Toxicol. 2017, 120, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, M.C.; de Brito Bezerra, A.P.; de Sousa, J.P.; de Oliveira Lima, E. Investigating the antifungal activity and mechanism(s) of geraniol against Candida albicans strains. Med. Mycol. 2015, 53, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; So, I.; Chun, J.N.; Jeon, J.H. The antitumor effects of geraniol: Modulation of cancer hallmark pathways (Review). Int. J. Oncol. 2016, 48, 1772–1782. [Google Scholar] [CrossRef]
- Soubh, A.A.; Abdallah, D.M.; El-Abhar, H.S. Geraniol ameliorates TNBS-induced colitis: Involvement of Wnt/β-catenin, p38MAPK, NFκB, and PPARγ signaling pathways. Life Sci. 2015, 136, 142–150. [Google Scholar] [CrossRef]
- Chen, J.; Fan, X.; Zhou, L.; Gao, X. Treatment with geraniol ameliorates methionine-choline-deficient diet-induced non-alcoholic steatohepatitis in rats. J. Gastroenterol. Hepatol. 2016, 31, 1357–1365. [Google Scholar] [CrossRef]
- Galle, M.; Kladniew, B.R.; Castro, M.A.; Villegas, S.M.; Lacunza, E.; Polo, M.; de Bravo, M.G.; Crespo, R. Modulation by geraniol of gene expression involved in lipid metabolism leading to a reduction of serum-cholesterol and triglyceride levels. Phytomed. Int. J. Phytother. Phytopharm. 2015, 22, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Zhang, L.; Li, N.; Mai, N.; Zhang, Y.; Pan, S. Geraniol promotes functional recovery and attenuates neuropathic pain in rats with spinal cord injury. Can. J. Physiol. Pharmacol. 2017, 95, 1389–1395. [Google Scholar] [CrossRef]
- Rizzello, F.; Ricci, C.; Scandella, M.; Cavazza, E.; Giovanardi, E.; Valerii, M.C.; Campieri, M.; Comparone, A.; De Fazio, L.; Candela, M.; et al. Dietary geraniol ameliorates intestinal dysbiosis and relieves symptoms in irritable bowel syndrome patients: A pilot study. BMC Complementary Altern. Med. 2018, 18, 338. [Google Scholar] [CrossRef]
- Younis, N.S.; Mohamed, M.E. β-Caryophyllene as a Potential Protective Agent Against Myocardial Injury: The Role of Toll-Like Receptors. Molecules 2019, 24, 1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saranya, S.; Baskaran, R.; Poornima, P.; Vijaya Padma, V. Berbamine ameliorates isoproterenol-induced myocardial infarction by inhibiting mitochondrial dysfunction and apoptosis in rats. J. Cell. Biochem. 2019, 120, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Ziaee, M.; Khorrami, A.; Nourafcan, H.; Ebrahimi, M.; Amiraslanzadeh, M.; Garjani, M.; Rameshrad, M.; Garjani, A. Cardioprotective Effects of Essential Oil of Lavandula angustifolia on Isoproterenol-induced Acute Myocardial Infarction in Rat. Iran. J. Pharm. Res. (IJPR) 2015, 14, 279–289. [Google Scholar] [PubMed]
- Xue, R.Q.; Sun, L.; Yu, X.J.; Li, D.L.; Zang, W.J. Vagal nerve stimulation improves mitochondrial dynamics via an M(3) receptor/CaMKKβ/AMPK pathway in isoproterenol-induced myocardial ischaemia. J. Cell. Mol. Med. 2017, 21, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Zhang, L.; Ding, M.; Li, M. Protective Effect of Vasicine Against Myocardial Infarction In Rats Via Modulation Of Oxidative Stress, Inflammation, and The PI3K/Akt Pathway. Drug Des. Dev. Ther. 2019, 13, 3773–3784. [Google Scholar] [CrossRef] [Green Version]
- El-Bassossy, H.M.; Elberry, A.A.; Ghareib, S.A. Geraniol improves the impaired vascular reactivity in diabetes and metabolic syndrome through calcium channel blocking effect. J. Diabetes Its Complicat. 2016, 30, 1008–1016. [Google Scholar] [CrossRef]
- Fan, C.; Tang, X.; Ye, M.; Zhu, G.; Dai, Y.; Yao, Z.; Yao, X. Qi-Li-Qiang-Xin Alleviates Isoproterenol-Induced Myocardial Injury by Inhibiting Excessive Autophagy via Activating AKT/mTOR Pathway. Front. Pharmacol. 2019, 10, 1329. [Google Scholar] [CrossRef] [Green Version]
- Tawfik, M.K.; El-Kherbetawy, M.K.; Makary, S. Cardioprotective and Anti-Aggregatory Effects of Levosimendan on Isoproterenol-Induced Myocardial Injury in High-Fat-Fed Rats Involves Modulation of PI3K/Akt/mTOR Signaling Pathway and Inhibition of Apoptosis: Comparison to Cilostazol. J. Cardiovasc. Pharmacol. Ther. 2018, 23, 456–471. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TLR Pathway Mediators | Accession Number | Primer Sequence (5′ to 3′) | |
|---|---|---|---|
| Forward | Reverse | ||
| Keap1 | NM_057152.2 | CTTCGGGGAGGA GGAGTTCT | CGTTCAGATCATCGCGGCTG |
| Nrf2 | NM_031789.2 | CATTTGTAGATGACCATGAG TCGC | ATCAGGGGTGGTGAAGACTG |
| HO-1 | NM_012580.2 | GTGCACATCGTGCAGAGAA | GTGCACATCCGTGCAGAGAA |
| PI3K | NM_001371300.1 | GCCCAGGCTTACTACAGAC | AAGTAGGGAGGCATCTCG |
| Akt | NM_033230.2 | CGCCTGCCCTTCTACAACC | TCATACACATCTTGCCACACGA |
| mTOR | NM_019906.1 | CTGTAATTACATCCTCGACTG | CGTGTCGTGGTTAGTCG |
| Bax | NM_017059.2 | GTGGTTGCCCTCTTCTACTTTG | CAAAAGATGGTCACTGTCTGC |
| Bcl-2 | NM_016993.1 | CCGGGAGATCGTGATGAAGT | ATCCCAGCC TCCGTTATCCT |
| β-actin | NM_031144.3 | CACGATGGAGGGGCCGGACTCATC | TAAAGACCTCTATGCCAACACAGT |
| ECG Component | Control | Geraniol | ISO | ISO + Geraniol (100 mg/kg) | ISO + Geraniol (200 mg/kg) |
|---|---|---|---|---|---|
| ST elevation (mV) | 0.0267 ± 0.0016 | 0.0252 ± 0.0018 | 0.1840 ± 0.0126 € | 0.1210 ± 0.0086 ҂ | 0.0810 ± 0.0066 ҂* |
| QT interval (sec) | 0.0453 ± 0.0037 | 0.0460 ± 0.0006 | 0.0933 ± 0.0034 € | 0.0743 ± 0.0028 ҂ | 0.0643 ± 0.0028 ҂* |
| P Wave (sec) | 0.0192 ± 0.0030 | 0.0190 ± 0.0019 | 0.0047 ± 0.0005 € | 0.0133 ± 0.0008 ҂ | 0.0163 ± 0.0008 ҂* |
| QRS complex (sec) | 0.0416 ± 0.0008 | 0.0417 ± 0.0005 | 0.0231 ± 0.0009 € | 0.0305 ± 0.0009 ҂ | 0.0338 ± 0.0009 ҂* |
| P-R interval (sec) | 0.2450 ± 0.0105 | 0.2600 ± 0.0167 | 0.1398 ± 0.0026 € | 0.1817 ± 0.0194 ҂ | 0.2117 ± 0.0194 ҂* |
| R-R interval (sec) | 0.2300 ± 0.0310 | 0.2237 ± 0.0117 | 0.1453 ± 0.0008 € | 0.1798 ± 0.0023 ҂ | 0.1983 ± 0.0023 ҂* |
| Antioxidants | Control | Geraniol | ISO | ISO + Geraniol (100 mg/kg) | ISO + Geraniol (200 mg/kg) |
|---|---|---|---|---|---|
| GSH (nmol/mg protein) | 7.28 ± 0.24 | 7.45 ± 0.12 | 3.66 ± 0.19 € | 5.49 ± 0.19 ҂ | 6.24 ± 0.26 ҂* |
| SOD (micromol H2O2/min/mg protein) | 124.14 ± 6.25 | 128.16 ± 5.46 | 65.43 ± 8.4 € | 89.49 ± 4.60 ҂ | 95.19 ± 6.10 ҂* |
| CAT (micromol H2O2/min/mg protein) | 1.49 ± 0.06 | 1.46 ± 0.08 | 0.64 ± 0.06 € | 1.02 ± 0.05 ҂ | 1.17 ± 0.03 ҂ |
| GPx (µg of GSH utilized/min/mg protein) | 19.63 ± 1.4 | 20.25 ± 2.1 | 7.46 ± 1.51 € | 12.10 ± 1.24 ҂ | 15.54 ± 2.43 ҂* |
| GST (nmol/mg protein) | 524.2 ± 15.86 | 532.41 ± 6.71 | 382.5 ± 12.93 € | 457.11 ± 4.24 ҂ | 483 ± 16.48 ҂* |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younis, N.S.; Abduldaium, M.S.; Mohamed, M.E. Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways. Antioxidants 2020, 9, 977. https://doi.org/10.3390/antiox9100977
Younis NS, Abduldaium MS, Mohamed ME. Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways. Antioxidants. 2020; 9(10):977. https://doi.org/10.3390/antiox9100977
Chicago/Turabian StyleYounis, Nancy S., Mohamed S. Abduldaium, and Maged E. Mohamed. 2020. "Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways" Antioxidants 9, no. 10: 977. https://doi.org/10.3390/antiox9100977
APA StyleYounis, N. S., Abduldaium, M. S., & Mohamed, M. E. (2020). Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways. Antioxidants, 9(10), 977. https://doi.org/10.3390/antiox9100977