Inter-Relationship between Platelet-Derived Microparticles and Oxidative Stress in Patients with Venous Thromboembolism

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Study Population
2.1. Laboratory Tests
2.1.1. Platelet-Derived Microparticles (MPs)
2.1.2. Pro-Thrombinase-Induced Clotting Time Assay (PiCT)
2.1.3. Phospholipids (PLPs)
2.1.4. Oxidative Stress Measures (MDA, 4-HNE, and TBARSs)
2.1.5. Galectin-3 (Gal-3)
2.1.6. Superoxide Dismutase Analysis
2.2. Statistical Analysis
3. Results
4. Discussion
5. Strengths and Limitations
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Konstantinides, S.V.; Meyer, G.; Becattini, C.; Bueno, H.; Geersing, G.-J.; Harjola, V.-P.; Huisman, M.V.; Humbert, M.; Jennings, C.S.; Jiménez, D.; et al. 2019 ESC Guidelines for the diagnosis and management of acute pulmonary embolism developed in collaboration with the European Respiratory Society (ERS). Eur. Hear. J. 2020, 41, 543–603. [Google Scholar] [CrossRef] [PubMed]
- Patrono, C.; Baigent, C. Role of aspirin in primary prevention of cardiovascular disease. Nat. Rev. Cardiol. 2019, 16, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Mezouar, S.; Frère, C.; Darbousset, R.; Mege, D.; Crescence, L.; Dignat-George, F.; Panicot-Dubois, L.; Dubois, C. Role of platelets in cancer and cancer-associated thrombosis: Experimental and clinical evidences. Thromb. Res. 2016, 139, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.; Gerdsen, F.; Gutensohn, K.; Schoder, V.; Eifrig, B.; Hossfeld, D.K. Enhanced platelet aggregation with TRAP-6 and collagen in platelet aggregometry in patients with venous thromboembolism. Thromb. Res. 2002, 107, 325–328. [Google Scholar] [CrossRef]
- Ferreiro, J.L.; Gómez-Hospital, J.A.; Angiolillo, D.J. Review article: Platelet abnormalities in diabetes mellitus. Diabetes Vasc. Dis. Res. 2010, 7, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Vieira-De-Abreu, A.; Campbell, R.A.; Weyrich, A.S.; Zimmerman, G.A. Platelets: Versatile effector cells in hemostasis, inflammation, and the immune continuum. Semin. Immunopathol. 2012, 34, 5–30. [Google Scholar] [CrossRef] [Green Version]
- Becattini, C.; Agnelli, G.; Schenone, A.; Eichinger, S.; Bucherini, E.; Silingardi, M.; Bianchi, M.; Moia, M.; Ageno, W.; Vandelli, M.R.; et al. Aspirin for Preventing the Recurrence of Venous Thromboembolism. N. Engl. J. Med. 2012, 366, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Brighton, T.A.; Eikelboom, J.W.; Mann, K.; Mister, R.; Gallus, A.; Ockelford, P.; Gibbs, H.; Hague, W.; Xavier, D.; Diaz, R.; et al. Low-Dose Aspirin for Preventing Recurrent Venous Thromboembolism. N. Engl. J. Med. 2012, 367, 1979–1987. [Google Scholar] [CrossRef] [Green Version]
- Simes, J.; Becattini, C.; Agnelli, G.; Eikelboom, J.W.; Kirby, A.C.; Mister, R.; Prandoni, P.; Brighton, T.A. INSPIRE Study Investigators (International Collaboration of Aspirin Trials for Recurrent Venous Thromboembolism). Aspirin for the prevention of recurrent venous thromboembolism: The INSPIRE collaboration. Circulation 2014, 130, 1062–1071. [Google Scholar] [CrossRef]
- Piccin, A.; Murphy, W.G.; Smith, O.P. Circulating microparticles: Pathophysiology and clinical implications. Blood Rev. 2007, 21, 157–171. [Google Scholar] [CrossRef]
- Herault, J.P.; Perrin, B.; Jongbloet, C.; Pflieger, A.M.; Bernat, A.; Herbert, J.-M. Effect of Factor Xa Inhibitors on the Platelet-derived Microparticles Procoagulant Activity In Vitro> and In Vivo in Rats. Thromb. Haemost. 2000, 84, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Re, G.; Lanzarini, C.; Vaona, I.; Pazzaglia, M.; Palareti, G.; Bassein, L.; Guarnieri, C. Systemically circulating oxidative species in human deep venous thrombosis. Eur. J. Emerg. Med. 1998, 5, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Golomb, B.A.; Chan, V.T.; O’Denenberg, J.Q.; Koperski, S.; Criqui, M.H. Risk marker associations with venous thrombotic events: A cross-sectional analysis. BMJ Open 2014, 4, e003208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrante, M.; Fiore, M.; Conti, G.O.; Fiore, V.; Grasso, A.; Copat, C.; Signorelli, S.S. Transition and heavy metals compared to oxidative parameter balance in patients with deep vein thrombosis: A case-control study. Mol. Med. Rep. 2017, 15, 3438–3444. [Google Scholar] [CrossRef] [Green Version]
- Mühl, D.; Füredi, R.; Cristofari, J.; Ghosh, S.; Bogar, L.; Borsiczki, B.; Gasz, B.; Rőth, E.; Lantos, J. Evaluation of oxidative stress in the thrombolysis of pulmonary embolism. J. Thromb. Thrombolysis 2006, 22, 221–228. [Google Scholar] [CrossRef]
- Fashanu, O.E.; Heckbert, S.R.; Aguilar, D.; Jensen, P.N.; Ballantyne, C.M.; Basu, S.; Hoogeveen, R.C.; Defilippi, C.; Cushman, M.; Folsom, A.R. Galectin-3 and venous thromboembolism incidence: The Atherosclerosis Risk in Communities (ARIC) Study. Res. Pr. Thromb. Haemost. 2017, 1, 223–230. [Google Scholar] [CrossRef] [Green Version]
- DeRoo, E.P.; Wrobleski, S.K.; Shea, E.M.; Al-Khalil, R.K.; Hawley, A.E.; Henke, P.K.; Myers, J.D.D.; Wakefield, T.W.; Diaz, J.A. The role of galectin-3 and galectin-3–binding protein in venous thrombosis. Blood 2015, 125, 1813–18221. [Google Scholar] [CrossRef] [Green Version]
- Riva, N.; Branchford, B.R.; Carpenter, S.L. The Role of Inflammation in Venous Thromboembolism. Front. Pediatr. 2018, 6, 142. [Google Scholar] [CrossRef]
- Prandoni, P. Links between arterial and venous disease. J. Intern. Med. 2007, 262, 341–350. [Google Scholar] [CrossRef]
- Fox, E.A.; Kahn, S.R. The relationship between inflammation and venous thrombosis. A systematic review of clinical studies. Thromb. Haemost. 2005, 94, 362–365. [Google Scholar] [CrossRef]
- Piazza, G.; Goldhaber, S.Z. Venous Thromboembolism and Atherothrombosis An Integrated Approach. Circulation 2010, 121, 2146–2150. [Google Scholar] [CrossRef] [PubMed]
- Pigault, C.; Follenius-Wund, A.; Schmutz, M.; Freyssinet, J.M.; Brisson, A. Formation of two-dimensional arrays of annexin V on phosphatidylserine-containing liposomes. J. Mol. Biol. 1994, 236, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, A.-L.; Sartz, L.; Karpman, D. Complement activation on platelet-leukocyte complexes and microparticles in enterohemorrhagic Escherichia coli–induced hemolytic uremic syndrome. Blood 2011, 117, 5503–5513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, S.; Nakamura, T.; Cone, J.; Tandon, N.N.; Kambayashi, J. Cytometric analysis of high shear-induced platelet microparticles and effect of cytokines on microparticle generation. Cytometry 2000, 40, 173–181. [Google Scholar] [CrossRef]
- Morel, O.; Toti, F.; Hugel, B.; Bakouboula, B.; Camoin-Jau, L.; Dignat-George, F.; Freyssinet, J.-M. Procoagulant Microparticles: Disrupting the vascular equation? Arter. Thromb. Vasc. Biol. 2006, 26, 2594–2604. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.T.; Hsu, D.K.; Zuberi, R.I.; Hill, P.N.; Shenhav, A.; Kuwabara, I.; Chen, S.S. Modulation of functional properties of galectin-3 by monoclonal antibodies binding to the non-lectin domains. Biochemistry 1996, 35, 6073–6079. [Google Scholar] [CrossRef]
- Dumic, J.; Dabelic, S.; Flögel, M. Galectin-3: An open-ended story. Biochim. Biophys. Acta (BBA) Gen. Subj. 2006, 1760, 616–635. [Google Scholar] [CrossRef]
- Yang, R.-Y.; Rabinovich, G.A.; Liu, F.-T. Galectins: Structure, function and therapeutic potential. Expert Rev. Mol. Med. 2008, 10, e17. [Google Scholar] [CrossRef]
- Ramacciotti, E.; Hawley, A.E.; Wrobleski, S.K.; Myers, D.D., Jr.; Strahler, J.R.; Andrews, P.C.; Guire, K.E.; Henke, P.K.; Wakefield, T.W. Proteomics of microparticles after deep venous thrombosis. Thromb. Res. 2010, 125, e269-74. [Google Scholar] [CrossRef] [Green Version]
- Rupa-Matysek, J.; Gil, L.; Wojtasińska, E.; Ciepłuch, K.; Lewandowska, M.; Komarnicki, M. The relationship between mean platelet volume and thrombosis recurrence in patients diagnosed with antiphospholipid syndrome. Rheumatol. Int. 2014, 34, 1599–1605. [Google Scholar] [CrossRef] [Green Version]
- Brækkan, S.K.; Mathiesen, E.B.; Njølstad, I.; Wilsgaard, T.; Størmer, J.; Hansen, J.B. Mean platelet volume is a risk factor for venous thromboembolism: The Tromsø study. J. Thromb. Haemost. 2010, 8, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Kostrubiec, M.; Łabyk, A.; Pedowska-Włoszek, J.; Hrynkiewicz-Szymańska, A.; Pacho, S.; Jankowski, K.; Lichodziejewska, B.; Pruszczyk, P. Mean platelet volume predicts early death in acute pulmonary embolism. Heart 2009, 96, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Kaider, A.; Schwarzinger, I.; Riedl, J.; Reitter, E.-M.; Marosi, C.; Jäger, U.; Zielinski, C.; Pabinger, I.; Ay, C. Association of mean platelet volume with risk of venous thromboembolism and mortality in patients with cancer. Results from the Vienna Cancer and Thrombosis Study (CATS). Thromb. Haemost. 2014, 111, 670–678. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
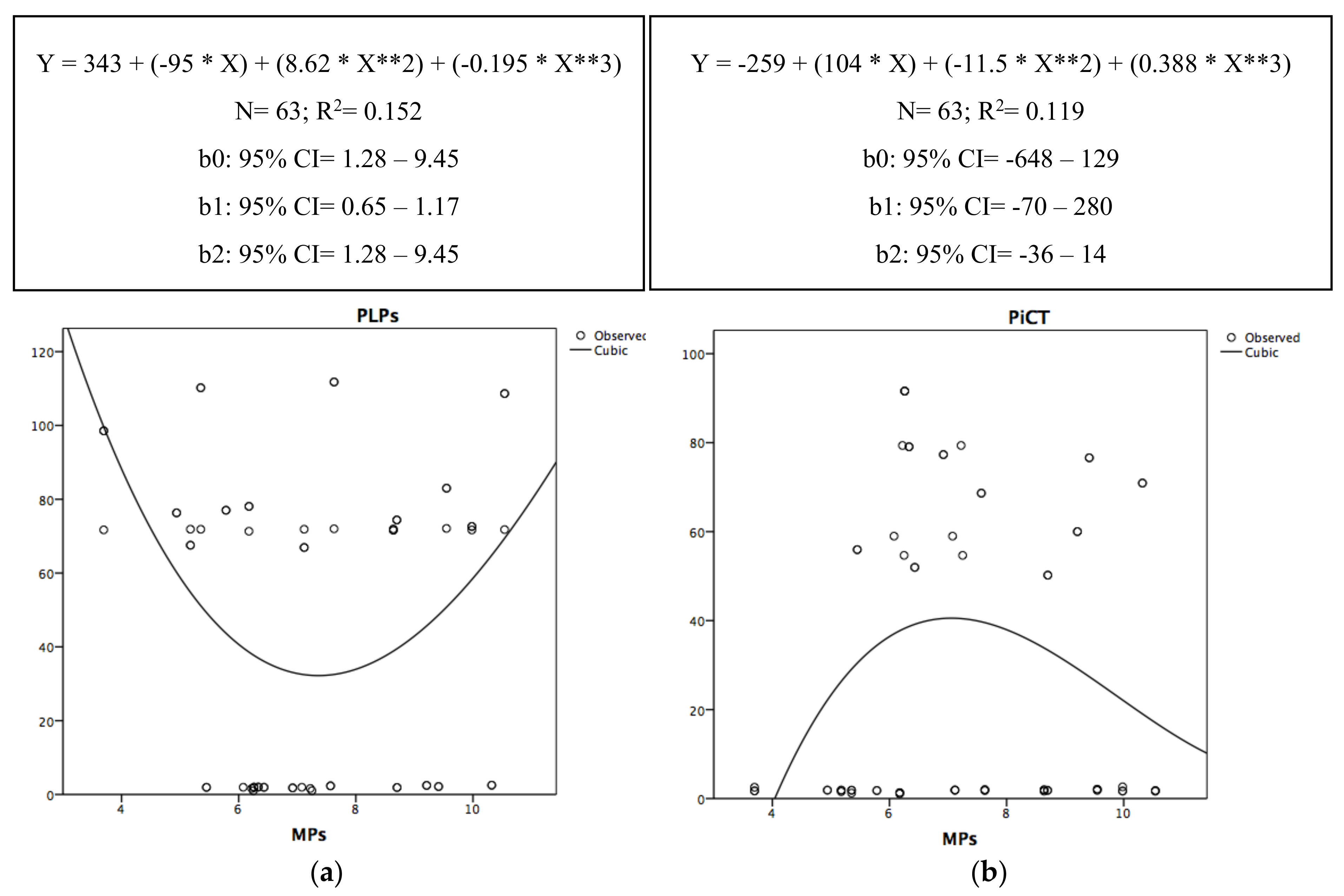{kind=link}
| Cases | Controls | |
|---|---|---|
| N | 36 | 36 |
| Males, n (%) | 24 (66.7) | 14 (53.8) |
| Median age (years) (IQR) * | 64 (50–72) | 55 (52–58) |
| Pulmonary embolism, n (%) | 11 (30.6) | 0 |
| Deep vein thrombosis, n (%) | 25 (69.4) | 0 |
| Previous VTE, n (%) | 10 (27.7) | 0 |
| No previous VTE, n (%) | 20 (55.5) | 0 |
| Heart disease, n (%) | 10 (27.7) | 0 |
| Thrombophilia, n (%) | 12 (33.3) | 0 |
| Type 2 diabetes, n (%) | 7 (19.4) | 0 |
| Overweight, n (%) | 10 (27.7) | 0 |
| Cases Median (IQR) * | Controls Median (IQR) * | DVT Median (IQR) * | PE Median (IQR) * | p-Value a | p-Value b | |
|---|---|---|---|---|---|---|
| MDA (µM/L) | 17.7 (15.1–22.4) | 3.8 (3.0–4.1) | 17.8 (15.3–22.4) | 14.7 (4.5–23.1) | 0.00 | 0.2 |
| 4-HNE (µM/L) | 11.5 (9.4–13.1) | 0.3 (0.2–0.4) | 11.5 (11.2–13.1) | 12.4 (7.7–14.7) | 0.00 | 0.49 |
| TBARs (µM/L) | 16.5 (14.2–19.3) | 2.7 (2.0–3.8) | 16.5 (13.8–19.3) | 16.9 (16.0–21.7) | 0.00 | 0.15 |
| Gal-3 (ng/mL) | 18.3 (14.9–19.1) | 12.7 (10.7–17.3) | 18.2 (14.9–19.6) | 18.3 (13.7–18.7) | 0.00 | 0.45 |
| SOD (U/mL) | 0.03 (0.01–0.07) | 17.05 (14.63–19.02) | 0.07 (0.03–0.08) | 0.01 (0.01–0.02) | 0.00 | 0.00 |
| MPs (nM PHS equivalent) | 7.1 (5.3–8.9) | 7.0 (6.3–8.8) | 7.1 (5.2–8.7) | 7.1 (5.4–10.5) | 0.54 | 0.64 |
| PLPs (µg/mL) | 74.4 (71.8–86.9) | 2.0 (1.9–2.2) | 74.4 (71.9–83.0) | 76.3 (71.6–108.6) | 0.00 | 0.92 |
| PiCT (0.00 s) | 1.9 (1.7–1.9) | 68.7 (55.6–77.7) | 1.9 (1.7–2.0) | 1.9 (1.8–2.0) | 0.00 | 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Signorelli, S.S.; Conti, G.O.; Fiore, M.; Elfio, M.G.; Cristaldi, A.; Nicolosi, I.; Zuccarello, P.; Zanoli, L.; Gaudio, A.; Di Raimondo, D.; et al. Inter-Relationship between Platelet-Derived Microparticles and Oxidative Stress in Patients with Venous Thromboembolism. Antioxidants 2020, 9, 1217. https://doi.org/10.3390/antiox9121217
Signorelli SS, Conti GO, Fiore M, Elfio MG, Cristaldi A, Nicolosi I, Zuccarello P, Zanoli L, Gaudio A, Di Raimondo D, et al. Inter-Relationship between Platelet-Derived Microparticles and Oxidative Stress in Patients with Venous Thromboembolism. Antioxidants. 2020; 9(12):1217. https://doi.org/10.3390/antiox9121217
Chicago/Turabian StyleSignorelli, Salvatore Santo, Gea Oliveri Conti, Maria Fiore, Maria Grazia Elfio, Antonio Cristaldi, Ilenia Nicolosi, Pietro Zuccarello, Luca Zanoli, Agostino Gaudio, Domenico Di Raimondo, and et al. 2020. "Inter-Relationship between Platelet-Derived Microparticles and Oxidative Stress in Patients with Venous Thromboembolism" Antioxidants 9, no. 12: 1217. https://doi.org/10.3390/antiox9121217
APA StyleSignorelli, S. S., Conti, G. O., Fiore, M., Elfio, M. G., Cristaldi, A., Nicolosi, I., Zuccarello, P., Zanoli, L., Gaudio, A., Di Raimondo, D., & Ferrante, M. (2020). Inter-Relationship between Platelet-Derived Microparticles and Oxidative Stress in Patients with Venous Thromboembolism. Antioxidants, 9(12), 1217. https://doi.org/10.3390/antiox9121217