Pechiche (Vitex cymosa Berteo ex Speng), a Nontraditional Fruit from Ecuador, is a Dietary Source of Phenolic Acids and Nutrient Minerals, in Addition to Efficiently Counteracting the Oxidative-Induced Damage in Human Dermal Fibroblasts

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pechiche Fruits
2.2. Phenolic Acids Analysis
2.2.1. Extraction of Phenolic Acids
2.2.2. HPLC-DAD Analysis of Phenolic Acids Profile
2.3. Vitamin C and Carotenoid Content
2.4. Inductively Coupled Plasma Optical Emission Spectrometry (ICP-OES) for Mineral Nutrient Composition
2.5. Protective Effect of Crude Extract of Pechiche on Human Dermal Fibroblasts against Oxidative Damage
2.5.1. Cell Line and Treatments
2.5.2. Determination of Oxidative Damage Markers
2.6. Statistical Analyses
3. Results and Discussion
3.1. Bioactive Compounds
3.2. Nutrient Minerals Composition
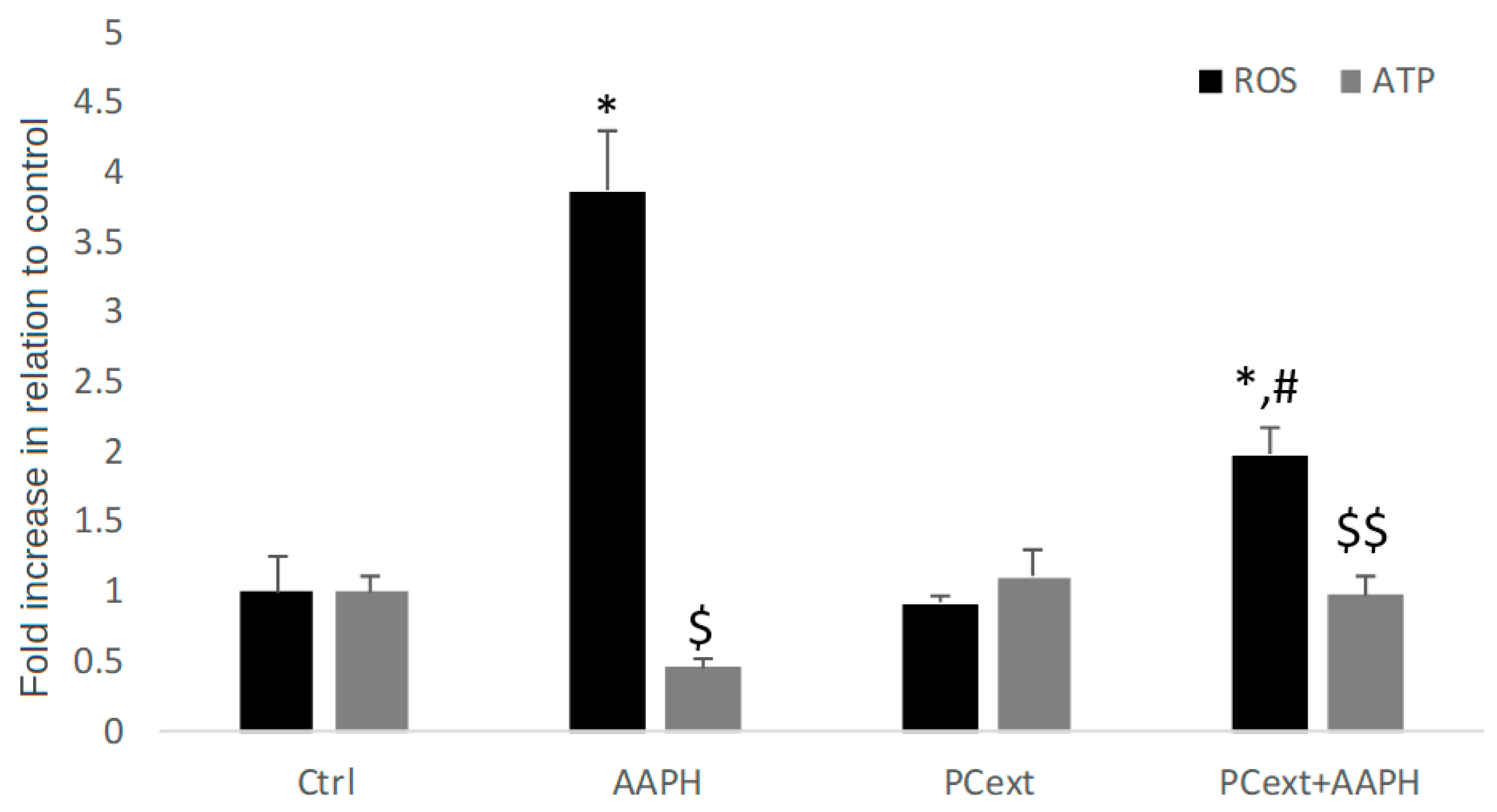
3.3. Protective Effect of Pechiche Extract against Oxidative Cytotoxic Damage in HDFa
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Tosti, V.; Bertozzi, B.; Fontana, L. Health Benefits of the Mediterranean Diet: Metabolic and Molecular Mechanisms. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Müller, M.J.; Oberritter, H.; Schulze, M.B.; et al. Critical review: vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [Green Version]
- Alarcón-Barrera, K.S.; Armijos-Montesinos, D.S.; García-Tenesaca, M.; Iturralde, G.; Jaramilo-Vivanco, T.; Granda-Albuja, M.G.; Giampieri, F.; Alvarez-Suarez, J.M. Wild Andean blackberry (Rubus glaucus Benth) and Andean blueberry (Vaccinium floribundum Kunth) from the Highlands of Ecuador: Nutritional composition and protective effect on human dermal fibroblasts against cytotoxic oxidative damage. J. Berry Res. 2018, 8, 223–236. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Carrillo-Perdomo, E.; Aller, A.; Giampieri, F.; Gasparrini, M.; González-Pérez, L.; Beltrán-Ayala, P.; Battino, M. Anti-inflammatory effect of Capuli cherry against LPS-induced cytotoxic damage in RAW 264.7 macrophages. Food Chem. Toxicol. 2017, 102, 46–52. [Google Scholar] [CrossRef]
- Pérez-Balladares, D.; Castañeda-Terán, M.; Granda-Albuja, M.G.; Tejera, E.; Iturralde, G.; Granda-Albuja, S.; Jaramillo-Vivanco, T.; Giampieri, F.; Battino, M.; Alvarez-Suarez, J.M. Chemical Composition and Antioxidant Activity of the Main Fruits, Tubers and Legumes Traditionally Consumed in the Andean Regions of Ecuador as a Source of Health-Promoting Compounds. Plant Foods Hum. Nutr. 2019, 74, 350–357. [Google Scholar] [CrossRef]
- Carrillo-Perdomo, E.; Aller, A.; Cruz-Quintana, S.M.; Giampieri, F.; Alvarez-Suarez, J.M. Andean berries from Ecuador: A review on Botany, Agronomy, Chemistry and Health Potential. J. Berry Res. 2015, 5, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Guevara, M.; Tejera, E.; Granda-Albuja, M.G.; Iturralde, G.; Chisaguano-Tonato, M.; Granda-Albuja, S.; Jaramillo-Vivanco, T.; Giampieri, F.; Battino, M.; Alvarez-Suarez, J.M. Chemical Composition and Antioxidant Activity of the Main Fruits Consumed in the Western Coastal Region of Ecuador as a Source of Health-Promoting Compounds. Antioxidants 2019, 8, 387. [Google Scholar] [CrossRef] [Green Version]
- Vasco, C.; Riihinen, K.; Ruales, J.; Kamal-Eldin, A. Phenolic Compounds in Rosaceae Fruits from Ecuador. J. Agric. Food Chem. 2009, 57, 1204–1212. [Google Scholar] [CrossRef]
- Vasco, C.; Riihinen, K.; Ruales, J.; Kamal-Eldin, A. Chemical composition and phenolic compound profile of mortiño (vaccinium floribundum kunth). J. Agric. Food Chem. 2009, 57, 8274–8281. [Google Scholar] [CrossRef]
- Vasco, C.; Ruales, J.; Kamal-Eldin, A. Total phenolic compounds and antioxidant capacities of major fruits from Ecuador. Food Chem. 2008, 111, 816–823. [Google Scholar] [CrossRef]
- Aguirre, Z. Especies forestales de los bosques secos del Ecuador. Guia dendrológica 393 para su identificación y caracterización. Proyecto Manejo Forestal Sostenible ante el Cambio; Quito, Ecuador. 2012. Available online: https://coin.fao.org/coin-static/cms/media/21/14042335632720/especies_forestales_bosques_secos_del_ecuador.pdf (accessed on 26 January 2020).
- Alicia, C.; Rodríguez-Sánchez, J.L. Caracterización nutricional y capacidad antioxidante del fruto del pechiche (Vitex gigantea Kunth). Cienc. Tecnol. Aliment. 2018, 27, 19–22. (in Spanish). [Google Scholar]
- De la Torre, L.; Muriel, M.; Navarrete, H.; Macía, M.J.; Balslev, H. Enciclopedia de las Plantas Útiles del Ecuador; 2008. Available online: https://scholarspace.manoa.hawaii.edu/bitstream/10125/47330/de%20la%20Torre%20et%20al.%202008%20Encyclopedia%20of%20useful%20plants%20of%20Ecuador.pdf (accessed on 26 January 2020).
- Gawlik-Dziki, U.; Świeca, M.; Dziki, D. Comparison of phenolic acids profile and antioxidant potential of six varieties of spelt (Triticum spelta L.). J. Agric. Food Chem. 2012, 60, 4603–4612. [Google Scholar] [CrossRef]
- Maines, M.D. Current Protocols in Toxicology; John Wiley & Sons, Inc.: New York, NY, USA, 1998. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar]
- Giampieri, F.; Alvarez-Suarez, J.M.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Afrin, S.; Bompadre, S.; Rubini, C.; Zizzi, A.; Astolfi, P.; Santos-Buelga, C.; et al. Strawberry consumption alleviates doxorubicin-induced toxicity by suppressing oxidative stress. Food Chem. Toxicol. 2016, 94, 128–137. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Jiang, Z.-Y.; Hunt, J.V.; Wolff, S.P. Ferrous ion oxidation in the presence of xylenol orange for detection of lipid hydroperoxide in low density lipoprotein. Anal. Biochem. 1992, 202, 384–389. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.-G.; Ahn, B.-W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [Green Version]
- Bianco, A.; Uccella, N. Biophenolic components of olives. Food Res. Int. 2000, 33, 475–485. [Google Scholar] [CrossRef]
- Papadopoulos, G.; Boskou, D. Antioxidant effect of natural phenols on olive oil. J. Am. Oil Chem. Soc. 1991, 68, 669–671. [Google Scholar] [CrossRef]
- Nardini, M.; Ghiselli, A. Determination of free and bound phenolic acids in beer. Food Chem. 2004, 84, 137–143. [Google Scholar] [CrossRef]
- Ito, K.; Nakazato, T.; Yamato, K.; Miyakawa, Y.; Yamada, T.; Hozumi, N.; Segawa, K.; Ikeda, Y.; Kizaki, M. Induction of Apoptosis in Leukemic Cells by Homovanillic Acid Derivative, Capsaicin, through Oxidative Stress. Cancer Res. 2004, 64, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Anton, D.; Bender, I.; Kaart, T.; Roasto, M.; Heinonen, M.; Luik, A.; Püssa, T. Changes in Polyphenols Contents and Antioxidant Capacities of Organically and Conventionally Cultivated Tomato (Solanum lycopersicum L.) Fruits during Ripening. Int. J. Anal. Chem. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Owen, R.W.; Haubner, R.; Mier, W.; Giacosa, A.; Hull, W.E.; Spiegelhalder, B.; Bartsch, H. Isolation, structure elucidation and antioxidant potential of the major phenolic and flavonoid compounds in brined olive drupes. Food Chem. Toxicol. 2003, 41, 703–717. [Google Scholar] [CrossRef]
- Besnard, G.; Rubio de Casas, R.; Christin, P.A.; Vargas, P. Phylogenetics of Olea (Oleaceae) based on plastid and nuclear ribosomal DNA sequences: Tertiary climatic shifts and lineage differentiation times. Ann. Bot. 2009, 104, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, H. Brazilian Trees: A Guide to the Identification and Cultivation of Brazilian Native Trees; Instituto Plantarum de Estudos da Flora: Nova Odessa, SP, Brazil, 2002; ISBN 9788586714153. [Google Scholar]
- Almeida, M.M.B.; De Sousa, P.H.M.; Arriaga, Â.M.C.; Prado, G.M.D.; Magalhães, C.E.D.C.; Maia, G.A.; De Lemos, T.L.G. Bioactive compounds and antioxidant activity of fresh exotic fruits from northeastern Brazil. Food Res. Int. 2011, 44, 2155–2159. [Google Scholar] [CrossRef] [Green Version]
- USDA. United States Standards for Grades of Extracted Honey. In Agricultural Marketing Service Fruit and Vegetable Division Processed Products Branch; US Department of Agriculture: Washington, DC, USA, 1985. [Google Scholar]
- Food and Nutrition Board Staff; Panel on Dietary Antioxidants; Institute of Medicine Staff. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium and Carotenoids: A Report of the Panel on Dietary Antioxidants and Related Compounds, Subcommitties on Upper Reference Levels of Nutrients and of the Interpretation and Use of Dietary Reference Intakes, and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes, Food and Nutrition Board, Institute of Medicine; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Ramful, D.; Tarnus, E.; Aruoma, O.I.; Bourdon, E.; Bahorun, T. Polyphenol composition, vitamin C content and antioxidant capacity of Mauritian citrus fruit pulps. Food Res. Int. 2011, 44, 2088–2099. [Google Scholar] [CrossRef]
- Khayyat, M.; Barati, Z.; Aminifard, M.H.; Samadzadeh, A. Changes in fruit maturity indices and growth pattern along the harvest season in seedless barberry under different altitude conditions. J. Berry Res. 2018, 8, 25–40. [Google Scholar] [CrossRef]
- FAO; WHO. Vitamin and mineral requirements in human nutrition, Second edition; World Health Organization: Geneva, Switzerland, 2004; ISBN 1014-9228. [Google Scholar]
- Weaver, C.M. Potassium and Health. Adv. Nutr. 2013, 4, 368S–377S. [Google Scholar] [CrossRef]
- Chakraborti, S.; Chakraborti, T.; Mandal, M.; Mandal, A.; Das, S.; Ghosh, S. Protective role of magnesium in cardiovascular diseases: A review. Mol. Cell. Biochem. 2002, 238, 163–179. [Google Scholar] [CrossRef]
- Pu, F.; Chen, N.; Xue, S. Calcium intake, calcium homeostasis and health. Food Sci. Hum. Wellness 2016, 5, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Lieu, P.T.; Heiskala, M.; Peterson, P.A.; Yang, Y. The roles of iron in health and disease. Mol. Asp. Med. 2001, 22, 1–87. [Google Scholar] [CrossRef]
- Cherasse, Y.; Urade, Y. Dietary Zinc Acts as a Sleep Modulator. Int. J. Mol. Sci. 2017, 18, E2334. [Google Scholar] [CrossRef] [Green Version]
- Santhakumar, A.B.; Battino, M.; Alvarez-Suarez, J.M. Dietary polyphenols: Structures, bioavailability and protective effects against atherosclerosis. Food Chem. Toxicol. 2018, 113, 49–65. [Google Scholar] [CrossRef]
- Werber, J.; Wang, Y.J.; Milligan, M.; Li, X.; Ji, J.A. Analysis of 2,2’-Azobis (2-Amidinopropane) Dihydrochloride Degradation and Hydrolysis in Aqueous Solutions. J. Pharm. Sci. 2011, 100, 3307–3315. [Google Scholar] [CrossRef]
- Terry, L.A. Health-Promoting Properties of Fruits and Vegetables; CABI: Wallingford, UK, 2011; ISBN 1845935284. [Google Scholar]
- Alvarez-Suarez, J.M.; Giampieri, F.; Gasparrini, M.; Mazzoni, L.; Santos-Buelga, C.; González-Paramás, A.M.; Forbes-Hernández, T.Y.; Afrin, S.; Páez-Watson, T.; Quiles, J.L.; et al. The protective effect of acerola (Malpighia emarginata) against oxidative damage in human dermal fibroblasts through the improvement of antioxidant enzyme activity and mitochondrial functionality. Food Funct. 2017, 8, 3250–3258. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Giampieri, F.; Gasparrini, M.; Mazzoni, L.; Forbes-Hernandez, T.Y.; Afrin, S.; Battino, M. Guava (Psidium guajava L. cv. Red Suprema) Crude Extract Protect Human Dermal Fibroblasts against Cytotoxic Damage Mediated by Oxidative Stress. Plant Foods Hum. Nutr. 2018, 73, 18–24. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Mazzoni, L.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Gonzàlez-Paramàs, A.M.; Santos-Buelga, C.; Quiles, J.L.; Bompadre, S.; Mezzetti, B.; et al. Polyphenol-Rich Strawberry Extract Protects Human Dermal Fibroblasts against Hydrogen Peroxide Oxidative Damage and Improves Mitochondrial Functionality. Molecules 2014, 19, 7798–7816. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Analysis | Values |
|---|---|
| Phenolic acids (mg/100 g of FW) | |
| Gallic acid | 22.60 ± 12.74 |
| 3-(4-Hydroxyphenyl)propanoic acid | 24.11 ± 2.62 |
| p-Coumaric acid | 4.87 ± 0.01 |
| Homovanillic acid | 42.92 ± 1.34 |
| 4-Hydroxyphenylacetic acid | 12.52 ± 1.55 |
| Carotenoids (µg/g of FW) | |
| β-carotene | 0.78 ± 0.02 |
| Lutein | 22.7 ± 3.84 |
| Vitamin C (mg/100 g of FW) | 25.56 ± 1.14 |
| Macrominerals (mg/Kg of FW) | |
| Na | 197.13 ± 14.15 |
| K | 852.28 ± 21.05 |
| Ca | 546.56 ± 26.18 |
| Mg | 653.65 ± 32.25 |
| P | 1550.05 ± 102.36 |
| Microminerals (mg/Kg of FW) | |
| Fe | 28.87 ± 3.16 |
| Mn | 6.25 ± 1.16 |
| Zn | 9.94 ± 1.28 |
| Markers | Ctrl | AAPH | PCext | PCext + AAPH |
|---|---|---|---|---|
| CAT (U/mg prot/min) | 46.85 ± 1.64 | 18.74 ± 2.41 ** | 42.06 ± 3.11 | 32.11 ± 2.66 # |
| SOD (U/mg prot/min) | 78.22 ± 3.44 | 32.98 ± 2.87 ** | 81.74 ± 5.84 | 58.51 ± 3.21 ## |
| GPx (nmol mg prot/min) | 271.82 ± 22.47 | 83.41 ± 4.20 ** | 282.44 ± 36.21 | 176.52 ± 10.84 ## |
| GR (nmol mg prot/min) | 282.65 ± 18.41 | 72.68 ± 9.41 ** | 286.84 ± 21.46 | 188.32 ± 9.23 ## |
| GST (nmol /mg protein/min) | 564.51 ± 36.87 | 226.12 ± 14.94 ** | 571.68 ± 26.13 | 396.44 ± 21.03 ## |
| TBARS (µM) | 5.22 ± 0.84 | 9.69 ± 0.54 ** | 5.86 ± 0.64 | 6.44 ± 0.31 # |
| Lipid hydroperoxides (µM) | 66.54 ± 7.12 | 167.51 ± 10.41 ** | 68.71 ± 6.91 | 71.84 ± 4.52 # |
| Protein carbonyl (nmol/mg prot) | 0.82 ± 0.08 | 2.01 ± 0.21 ** | 0.76 ± 0.04 | 1.12 ± 0.04 ## |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guevara, M.; Valdés-Silverio, L.A.; Granda-Albuja, M.G.; Iturralde, G.; Jaramillo-Vivanco, T.; Giampieri, F.; Santos-Buelga, C.; González-Paramás, A.M.; Battino, M.; Álvarez-Suarez, J.M. Pechiche (Vitex cymosa Berteo ex Speng), a Nontraditional Fruit from Ecuador, is a Dietary Source of Phenolic Acids and Nutrient Minerals, in Addition to Efficiently Counteracting the Oxidative-Induced Damage in Human Dermal Fibroblasts. Antioxidants 2020, 9, 109. https://doi.org/10.3390/antiox9020109
Guevara M, Valdés-Silverio LA, Granda-Albuja MG, Iturralde G, Jaramillo-Vivanco T, Giampieri F, Santos-Buelga C, González-Paramás AM, Battino M, Álvarez-Suarez JM. Pechiche (Vitex cymosa Berteo ex Speng), a Nontraditional Fruit from Ecuador, is a Dietary Source of Phenolic Acids and Nutrient Minerals, in Addition to Efficiently Counteracting the Oxidative-Induced Damage in Human Dermal Fibroblasts. Antioxidants. 2020; 9(2):109. https://doi.org/10.3390/antiox9020109
Chicago/Turabian StyleGuevara, Mabel, Luis A. Valdés-Silverio, María G. Granda-Albuja, Gabriel Iturralde, Tatiana Jaramillo-Vivanco, Francesca Giampieri, Celestino Santos-Buelga, Ana M. González-Paramás, Maurizio Battino, and José M. Álvarez-Suarez. 2020. "Pechiche (Vitex cymosa Berteo ex Speng), a Nontraditional Fruit from Ecuador, is a Dietary Source of Phenolic Acids and Nutrient Minerals, in Addition to Efficiently Counteracting the Oxidative-Induced Damage in Human Dermal Fibroblasts" Antioxidants 9, no. 2: 109. https://doi.org/10.3390/antiox9020109
APA StyleGuevara, M., Valdés-Silverio, L. A., Granda-Albuja, M. G., Iturralde, G., Jaramillo-Vivanco, T., Giampieri, F., Santos-Buelga, C., González-Paramás, A. M., Battino, M., & Álvarez-Suarez, J. M. (2020). Pechiche (Vitex cymosa Berteo ex Speng), a Nontraditional Fruit from Ecuador, is a Dietary Source of Phenolic Acids and Nutrient Minerals, in Addition to Efficiently Counteracting the Oxidative-Induced Damage in Human Dermal Fibroblasts. Antioxidants, 9(2), 109. https://doi.org/10.3390/antiox9020109