The Effect of Diet Supplementation with Pomegranate and Bitter Melon on Lipidomic Profile of Serum and Cancerous Tissues of Rats with Mammary Tumours

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bitter Melon Aqueous Extract (BME)
2.2. Pomegranate Seed Oil (PSO)
2.3. Animals
- -
- CON and CONplus—control groups without diet supplementation, fed a standard diet and water ad libitum,
- -
- M and Mplus—animals fed a standard diet supplemented with 1% aqueous extract of bitter melon dried fruits ad libitum,
- -
- G and Gplus—animals were fed the standard diet and water ad libitum and were given 0.15 mL/day pomegranate seed oil via gavage,
- -
- GM and GMplus—animals were fed the standard diet and were supplemented with both 0.15 mL/day pomegranate seed oil administered via gavage and 1% aqueous extract of bitter melon dried fruits ad libitum.
2.4. Preparation of Research Material
2.5. FA Analysis in Serum
2.6. FA Analysis in Mammary Tumours
2.7. CFA Analysis in Mammary Tumours
2.8. Malondialdehyde (MDA) Analysis in Mammary Tumours
2.9. Total Cholesterol, Squalene and Oxysterol Analysis in Mammary Tumours
2.10. LOX Metabolites Analysis in Serum and Mammary Tumours
2.11. Statistical Analysis
3. Results
3.1. FA in Serum
3.2. LOX Metabolites in Serum
3.3. FA in Mammary Tumours
3.4. CFA in Mammary Tumours
3.5. LOX Metabolites in Mammary Tumours
3.6. MDA Content in Mammary Tumours
3.7. Squalene, Cholesterol and Oxysterol Content in Mammary Tumours
3.8. Linear Discriminant Analysis (LDA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
Appendix A

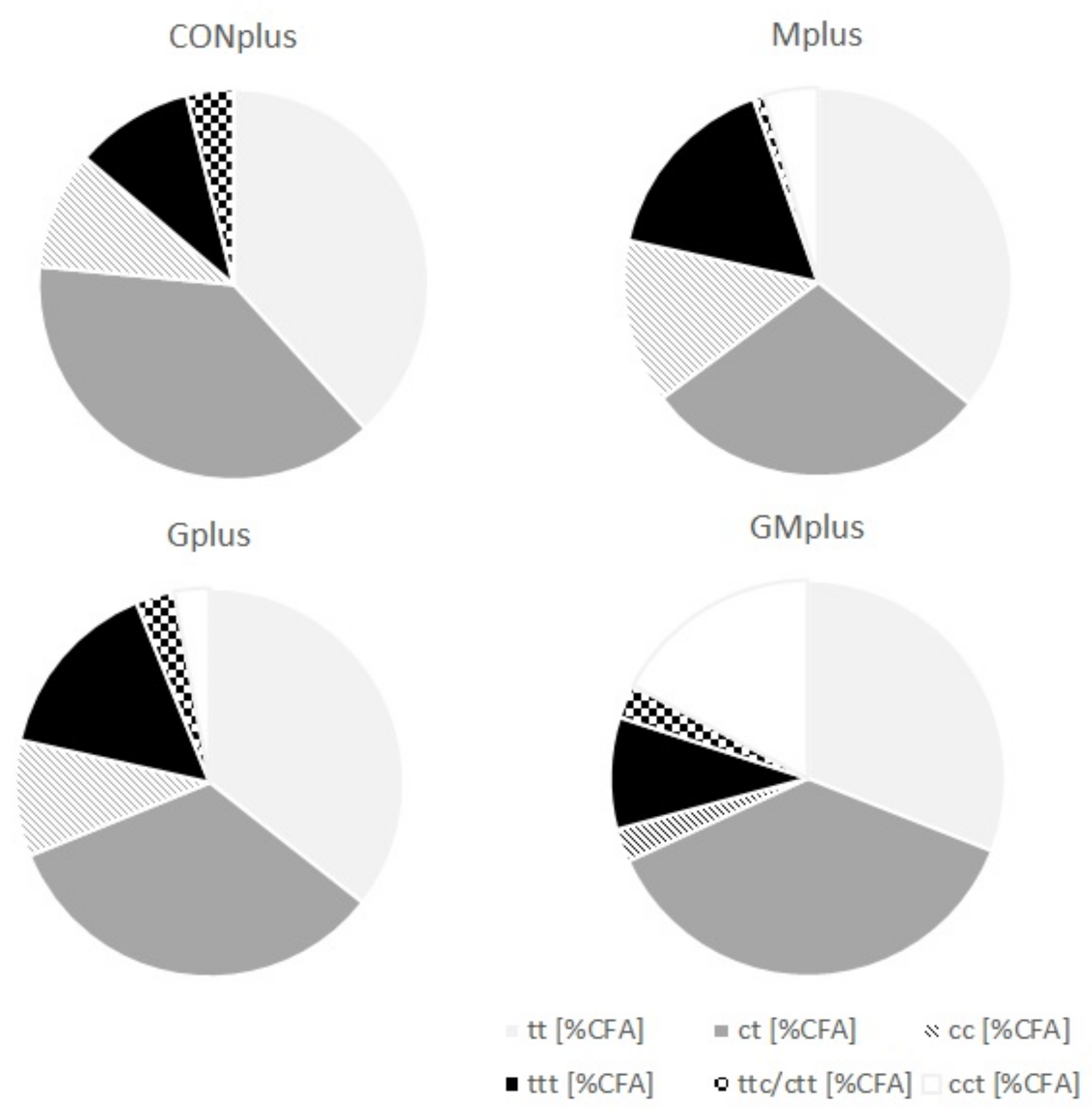{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | CONplus | Mplus | Gplus | GMplus | p Value |
|---|---|---|---|---|---|
| C14:0 | 2.79 ± 1.27 | 3.04 ± 1.08 | 5.64 ± 3.89 | 5.05 ± 2.48 | n.s. |
| C15:0 | 4.53 ± 0.92 | 4.84 ± 1.66 | 8.35 ± 3.88 | 7.34 ± 2.41 | n.s. |
| C16:0 | 481 ± 207 | 628 ± 205 | 606 ± 251 | 524 ± 137 | n.s. |
| C17:0 | 15.3 ± 6.9 | 14.7 ± 6.0 | 16.7 ± 7.7 | 14.9 ± 4.9 | n.s. |
| C18:0 | 441 ± 181 | 386 ± 50 | 292 ± 123 | 283 ± 104 | n.s. |
| C21:0 | 0.99 ± 0.20 | 1.82 ± 2.90 | 0.78 ± 0.56 | 1.45 ± 0.85 | n.s. |
| C24:0 | 8.86 ± 4.03 | 18.8 ± 11.7 | 7.06 ± 2.21 | 9.32 ± 2.87 | n.s. |
| SFA | 951 ± 390 | 1057 ± 215 | 937 ± 298 | 845 ± 234 | n.s. |
| C16:1 | 23.3 ± 14.7 | 22.4 ± 0.6 | 45.7 ± 40.5 | 30.4 ± 13.5 | n.s. |
| C17:1, c-Δ10 | 4.84 ± 1.75 | 2.56 ± 1.23 | 8.07 ± 5.82 | 4.79 ± 2.66 | n.s. |
| C18:1, c-Δ9 (OL) | 246 ± 124 | 288 ± 160 | 339 ± 221 | 249 ± 119 | n.s. |
| C20:1, c-Δ11 | 1.02 ± 0.47 | 0.49 (0.09-2.78) * | 1.44 ± 1.12 | 1.23 ± 0.80 | n.s. |
| MUFA | 274 ± 138 | 314 ± 163 | 387 ± 262 | 285 ± 133 | n.s. |
| C18:2, c,c-Δ9,12 (LA) | 351 ± 166 | 561 ± 299 | 566 ± 174 | 527 ± 131 | n.s. |
| C18:3, c,c,c-Δ6,9,12 (GLA) | 5.32 ± 2.82 | 6.98 ± 0.53 | 7.85 ± 7.42 | 4.73 ± 2.60 | n.s. |
| C18:3, c,c,c-Δ9,12,15 (ALA) | 5.31 ± 2.10 | 6.87 (2.44–9.32) * | 14.9 ± 10.6 | 11.9 ± 6.96 | n.s. |
| C18:2, c,t-Δ9,11 (RA) | - | - | 0.46 ± 0.01 | 0.52 ± 0.01 | n.s. |
| C20:2, c,c-Δ11,14 | 0.42 ± 0.17 | 0.97 (0.35–2.66) * | 1.82 (0.57–5.81) * | 1.55 ± 1.14 | n.s. |
| C20:3, c,c,c-Δ8,11,14 | 6.57 ± 2.52 | 6.28 ± 2.77 | 8.73 ± 3.41 | 7.06 ± 3.00 | n.s. |
| C20:4, c,c,c,c-Δ5,8,11,14 (AA) | 914 ± 451 | 784 ± 172 | 564 ± 233 | 572 ± 187 | n.s. |
| C20:3, c,c,c-Δ11,14,17 | 5.19 ± 2.28 | 4.69 ± 0.10 | 2.55 (0.67–9.67) * | 8.68 (1.49–50.7) * | n.s. |
| C20:5, c,c,c,c,c-Δ5,8,11,14,17 (EPA) | 5.90 ± 3.68 | 4.88 ± 1.24 | 11.3 ± 4.5 | 10.3 ± 8.1 | n.s. |
| C22:2, c,c-Δ13,16 | 1.07 ± 0.44 | 2.01 ± 0.96 | 2.19 ± 0.82 | 1.02 ± 0.23 | 0.0100 |
| C22:5, c,c,c,c,c-Δ7,10,13,16,19 | 12.2 ± 6.3 | 20.8 ± 5.2 | 18.4 ± 3.5 | 20.1 ± 5.3 | n.s. |
| C22:6, c,c,c,c,c,c-Δ4,7,10,13,16,19 (DHA) | 194 ± 101 | 178 ± 20 | 115 ± 49 | 122 ± 45 | n.s. |
| PUFA | 1499 ± 707 | 1576 ± 180 | 1315 ± 333 | 1314 ± 325 | n.s. |
| n3PUFA | 208 ± 104 | 194 ± 32 | 146 ± 57 | 180 ± 91 | n.s. |
| n6PUFA | 1290 ± 606 | 1379 ± 148 | 1165 ± 281 | 1131 ± 281 | n.s. |
| n6/n3 | 6.23 ± 0.71 | 7.14 ± 0.43 | 8.49 ± 1.89 | 7.18 ± 2.17 | n.s. |
| Coefficients of Canonical Variables | |||
|---|---|---|---|
| Variable (Discriminatory Power) | DF1 (74.3%) | DF2 (19.0%) | DF3 (6.7%) |
| tt | −2.87486 | −0.23823 | 0.84560 |
| cc | 0.41445 | −0.88030 | −0.61785 |
| tumour mass | 0.46920 | −0.81715 | −0.51740 |
| ct | 3.27553 | 0.21526 | 0.24013 |
| ttt | −0.15933 | −0.65019 | −0.23386 |
| 7α-OH-Ch | −0.53719 | −0.62480 | 0.20176 |
| 5-HETE/15-HETE | −0.38686 | 2.92710 | −0.12543 |
| HODE | −1.59109 | 0.16220 | 0.67288 |
| 12-HETE/15-HETE | 1.21217 | −2.15447 | −0.25894 |
| MDA | 1.03007 | −0.39936 | −0.05724 |
| squalene | 0.85398 | −0.66452 | 0.73374 |
| 7ß-OH-Ch | 0.03831 | 0.24962 | −1.68921 |
| RA | −2.05800 | 0.10286 | −0.65535 |
| 5α,6α-epoxy-Ch | −0.44120 | 0.35613 | 1.36723 |
| 15-HETE | 1.44316 | −0.55230 | −0.08770 |
| 12-HETE | −0.58887 | 1.98339 | −0.45563 |
| 5-HETE | −0.02018 | −2.08316 | 0.29005 |
| 25-OH-Ch | 0.40469 | −0.10663 | −0.07427 |
| cholesterol/squalene | −0.34923 | 0.20157 | −0.12313 |
| Average value of canonical variables | |||
| CONplus | 6.63668 | −0.19222 | 2.16305 |
| Mplus | 4.64837 | 0.27063 | −1.58810 |
| Gplus | −1.31511 | -2.42687 | −0.04219 |
| GMplus | −1.49936 | 1.03765 | 0.11535 |

References
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumours in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Temkin, S.M.; Hawkridge, A.M.; Guo, C.; Wang, W.; Wang, X.Y.; Fang, X. Fatty acid oxidation: An emerging facet of metabolic transformation in cancer. Cancer Lett. 2018, 435, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Orešič, M. Regulation of lipid metabolism in breast cancer provides diagnostic and therapeutic opportunities. Clin. Lipidol. 2012, 7, 177–188. [Google Scholar] [CrossRef]
- Luo, X.; Zhao, X.; Cheng, C.; Li, N.; Liu, Y.; Cao, Y. The implications of signaling lipids in cancer metastasis. Exp. Mol. Med. 2018, 50, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Du, G. Dysregulated lipid metabolism in cancer. World J. Biol. Chem. 2012, 3, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Geng, F.; Cheng, X.; Guo, D. Lipid metabolism reprogramming and its potential targets in cancer. Cancer Commun. 2018, 38, 27. [Google Scholar] [CrossRef]
- Cha, J.-Y.; Lee, H.-J. Targeting Lipid Metabolic Reprogramming as Anticancer Therapeutics. J. Cancer Prev. 2016, 21, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Kinlaw, W.B.; Baures, P.W.; Lupien, L.E.; Davis, W.L.; Kuemmerle, N.B. Fatty Acids and Breast Cancer: Make Them on Site or Have Them Delivered. J. Cell. Physiol. 2016, 231, 2128–2141. [Google Scholar] [CrossRef] [Green Version]
- Petan, T.; Jarc, E.; Jusović, M. Lipid droplets in cancer: Guardians of fat in a stressful world. Molecules 2018, 23, 1941. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, N.; Lupien, L.; Kuemmerle, N.B.; Kinlaw, W.B.; Swinnen, J.V.; Smans, K. Lipogenesis and lipolysis: The pathways exploited by the cancer cells to acquire fatty acids. Prog. Lipid Res. 2013, 52, 585–589. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Luo, Q.; Halim, A.; Song, G. Targeting lipid metabolism of cancer cells: A promising therapeutic strategy for cancer. Cancer Lett. 2017, 401, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Zhang, C.-J.; Zhu, N.; Du, K.; Yin, Y.-F.; Tan, X.; Liao, D.-F.; Qin, L. Lipid metabolism and carcinogenesis, cancer development. Am. J. Cancer Res. 2018, 8, 778–791. [Google Scholar] [PubMed]
- Kloudova, A.; Guengerich, F.P.; Soucek, P. The Role of Oxysterols in Human Cancer. Trends Endocrinol. Metab. 2017, 28, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Barriuso, B.; Ansorena, D.; Astiasarán, I. Oxysterols formation: A review of a multifactorial process. J. Steroid Biochem. Mol. Biol. 2017, 169, 39–45. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Wang, Y. Oxysterols as lipid mediators: Their biosynthetic genes, enzymes and metabolites. Prostaglandins Lipid Mediat. 2020, 147, 106381. [Google Scholar] [CrossRef]
- Barrera, G. Oxidative Stress and Lipid Peroxidation Products in Cancer Progression and Therapy. ISRN Oncol. 2012, 2012, 137289. [Google Scholar] [CrossRef] [Green Version]
- Gago-Dominguez, M.; Jiang, X.; Castelao, J.E. Lipid peroxidation, oxidative stress genes and dietary factors in breast cancer protection: A hypothesis. Breast Cancer Res. 2007, 9, 201. [Google Scholar] [CrossRef] [Green Version]
- Białek, A.; Jelińska, M.; Tokarz, A.; Pergół, A.; Pinkiewicz, K. Influence of pomegranate seed oil and bitter melon aqueous extract on polyunsaturated fatty acids and their lipoxygenase metabolites concentration in serum of rats. Prostaglandins Lipid Mediat. 2016, 126, 29–37. [Google Scholar] [CrossRef]
- Białek, A.; Tokarz, A. Conjugated linoleic acid as a potential protective factor in prevention of breast cancer. Postepy Hig. Med. Dosw. (Online) 2013, 67, 6–14. [Google Scholar] [CrossRef]
- Białek, A.; Teryks, M.; Tokarz, A. Sprzężone trieny kwasu linolenowego (conjugated linolenic acid—CLnA, super CLA)—źródła i działanie biologiczne. Conjugated linolenic acids (CLnA, super CLA)—Natural sources and biological activity. Postępy Higieny i Medycyny Doświadczalnej (Adv. Hyg. Exp. Med.) 2014, 68, 1238–1250. [Google Scholar]
- den Hartigh, L.J. Conjugated linoleic acid effects on cancer, obesity, and atherosclerosis: A review of pre-clinical and human trials with current perspectives. Nutrients 2018, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Białek, A.; Stawarska, A.; Bodecka, J.; Białek, M.; Tokarz, A. Pomegranate seed oil influences the fatty acids profile and reduces the activity of desaturases in livers of Sprague-Dawley rats. Prostaglandins Lipid Mediat. 2017, 131, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Białek, A.; Stawarska, A.; Tokarz, A.; Czuba, K.; Konarska, A.; Mazurkiewicz, M.; Stanimirova-Daszykowska, I. Enrichment of maternal diet with conjugated linoleic acids influences desaturases activity and fatty acids profile in livers and hepatic microsomes of the offspring with 7,12-dimethylbenz[A]anthracene-induced mammary tumors. Acta Pol. Pharm. - Drug Res. 2014, 71, 747–761. [Google Scholar]
- Stawarska, A.; Białek, A.; Stanimirova, I.; Stawarski, T.; Tokarz, A. The effect of Conjugated Linoleic Acids (CLA) supplementation on the activity of enzymes participating in the formation of arachidonic acid in liver microsomes of rats—Probable mechanism of CLA anticancer activity. Nutr. Cancer 2015, 67, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Jelińska, M.; Białek, A.; Gielecińska, I.; Mojska, H.; Tokarz, A. Impact of conjugated linoleic acid administered to rats prior and after carcinogenic agent on arachidonic and linoleic acid metabolites in serum and tumors. Prostaglandins Leukot. Essent. Fat. Acids 2017, 126, 1–8. [Google Scholar] [CrossRef]
- Białek, A.; Jelińska, M.; Tokarz, A. Influence of maternal diet enrichment with conjugated linoleic acids on lipoxygenase metabolites of polyunsaturated fatty acids in serum of their offspring with 7,12-dimethylbenz[a]anthracene induced mammary tumors. Prostaglandins Other Lipid Mediat. 2015, 116–117, 10–18. [Google Scholar] [CrossRef]
- Jelińska, M.; Białek, A.; Mojska, H.; Gielecińska, I.; Tokarz, A. Effect of conjugated linoleic acid mixture supplemented daily after carcinogen application on linoleic and arachidonic acid metabolites in rat serum and induced tumours. Biochim. Biophys. Acta - Mol. Basis Dis. 2014, 1842, 2230–2236. [Google Scholar] [CrossRef] [Green Version]
- Lepionka, T.; Białek, A.; Białek, M.; Czauderna, M.; Stawarska, A.; Wrzesień, R.; Bielecki, W.; Paśko, P.; Galanty, A.; Bobrowska-Korczak, B. Mammary cancer risk and serum lipid profile of rats supplemented with pomegranate seed oil and bitter melon extract. Prostaglandins Other Lipid Mediat. 2019, 142, 33–45. [Google Scholar] [CrossRef]
- Białek, M.; Białek, A.; Lepionka, T.; Paśko, P.; Galanty, A.; Tokarz, A.; Czauderna, M. Punica granatum (Pomegranate) Seed Oil and Momordica charantia (Bitter Melon) Extract Affect the Lipid’s Profile and Oxidative Stability of Femoral Muscles of Rats. Eur. J. Lipid Sci. Technol. 2019, 121, 1–10. [Google Scholar] [CrossRef]
- Białek, M.; Czauderna, M.; Białek, A. Partial replacement of rapeseed oil with fish oil, and dietary antioxidants supplementation affects concentrations of biohydrogenation products and conjugated fatty acids in rumen and selected lamb tissues. Anim. Feed Sci. Technol. 2018, 241, 63–74. [Google Scholar] [CrossRef]
- Czauderna, M.; Kowalczyk, J.; Korniluk, K.; Wasowska, I. Improved saponification then mild base and acid-catalyzed methylation is a useful method for quantifying fatty acids, with special emphasis on conjugated dienes. Acta Chromatogr. 2007, 18, 59–71. [Google Scholar]
- Czauderna, M.; Kowalczyk, J.; Marounek, M. The simple and sensitive measurement of malondialdehyde in selected specimens of biological origin and some feed by reversed phase high performance liquid chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 2251–2258. [Google Scholar] [CrossRef] [PubMed]
- Frohberg, P.; Drutkowski, G.; Wobst, I. Monitoring eicosanoid biosynthesis via lipoxygenase and cyclooxygenase pathways in human whole blood by single HPLC run. J. Pharm. Biomed. Anal. 2006, 41, 1317–1324. [Google Scholar] [CrossRef]
- Jelińska, M.; Skrajnowska, D.; Wrzosek, M.; Domanska, K.; Bielecki, W.; Zawistowska, M.; Bobrowska Korczak, B. Inflammation factors and element supplementation in cancer. J. Trace Elem. Med. Biol. 2020, 59, 126450. [Google Scholar] [CrossRef] [PubMed]
- Pakiet, A.; Kobiela, J.; Stepnowski, P.; Sledzinski, T.; Mika, A. Changes in lipids composition and metabolism in colorectal cancer: A review. Lipids Health Dis. 2019, 18, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceição, L.L.; Dias, M.D.M.; Pessoa, M.C.; Pena, G.D.G.; Mendes, M.C.S.; Neves, C.V.B.; Hermsdorf, H.H.M.; de Freitas, R.N.; do Peluzio, M.C.G. Difference in fatty acids composition of breast adipose tissue in women with breast cancer and benign breast disease. Nutr. Hosp. 2017, 33, 1354–1360. [Google Scholar] [CrossRef] [Green Version]
- Halczy-Kowalik, L.; Drozd, A.; Stachowska, E.; Drozd, R.; Żabski, T.; Domagała, W. Fatty acids distribution and content in oral squamous cell carcinoma tissue and its adjacent microenvironment. PLoS ONE 2019, 14, e0218246. [Google Scholar] [CrossRef]
- Amézaga, J.; Arranz, S.; Urruticoechea, A.; Ugartemendia, G.; Larraioz, A.; Louka, M.; Uriarte, M.; Ferreri, C.; Tueros, I. Altered red blood cell membrane fatty acid profile in cancer patients. Nutrients 2018, 10, 1853. [Google Scholar] [CrossRef] [Green Version]
- Lewin, A.A.; Storey, P.; Moccaldi, M.; Moy, L.; Gene Kim, S. Fatty acid composition in mammary adipose tissue measured by Gradient-echo Spectroscopic MRI and its association with breast cancers. Eur. J. Radiol. 2019, 116, 205–211. [Google Scholar] [CrossRef]
- Białek, A.; Tokarz, A.; Zagrodzki, P. Conjugated linoleic acids in diet of female rats inhibit the breast cancer formation in their offspring. J. Food Nutr. Res. 2014, 53, 39–50. [Google Scholar]
- Białek, A.; Zagrodzki, P.; Tokarz, A. Chemometric analysis of the interactions among different parameters describing health conditions, breast cancer risk and fatty acids profile in serum of rats supplemented with conjugated linoleic acids. Prostaglandins Leukot. Essent. Fat. Acids 2016, 106, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Białek, A.; Tokarz, A.; Zagrodzki, P. Conjugated Linoleic Acids (CLA) decrease the breast cancer risk in DMBA-treated rats. Acta Pol. Pharm. - Drug Res. 2015, 72, 1163–1176. [Google Scholar]
- Białek, A.; Tokarz, A.; Dudek, A.; Kazimierska, W.; Bielecki, W. Influence of diet enriched with conjugated linoleic acids on their distribution in tissues of rats with DMBA induced tumors. Lipids Health Dis. 2010, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avis, I.; Hong, S.H.; Martinez, A.; Moody, T.; Choi, Y.H.; Trepel, J.; Das, R.; Jett, M.; Mulshine, J.L. Five-lipoxygenase inhibitors can mediate apoptosis in human breast cancer cell lines through complex eicosanoid interactions. FASEB J. 2001, 15, 2007–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Blanco, G.; Burgers, P.C.; Dekker, L.J.M.; Ijzermans, J.J.N.; Wildhagen, M.F.; Schenk-Braat, E.A.M.; Bangma, C.H.; Jenster, G.; Luider, T.M. Serum levels of arachidonic acid metabolites change during prostate cancer progression. Prostate 2014, 74, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Vonach, C.; Viola, K.; Giessrigl, B.; Huttary, N.; Raab, I.; Kalt, R.; Krieger, S.; Vo, T.P.N.; Madlener, S.; Bauer, S.; et al. NF-κB mediates the 12(S)-HETE-induced endothelial to mesenchymal transition of lymphendothelial cells during the intravasation of breast carcinoma cells. Br. J. Cancer 2011, 105, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigby, D.A.; Ferguson, D.J.P.; Johnson, L.A.; Jackson, D.G. Neutrophils rapidly transit inflamed lymphatic vessel endothelium via integrin-dependent proteolysis and lipoxin-induced junctional retraction. J. Leukoc. Biol. 2015, 98, 897–912. [Google Scholar] [CrossRef]
- Romano, M.; Catalano, A.; Nutini, M.; D’Urbano, E.; Crescenzi, C.; Claria, J.; Libner, R.; Davi, G.; Procopio, A. 5-Lipoxygenase regulates malignant mesothelial cell survival: Involvement of vascular endothelial growth factor. FASEB J. 2001, 15, 2326–2336. [Google Scholar] [CrossRef]
- Rose, D.P.; Connolly, J.M.; Rayburn, J.; Coleman, M. Influence of diets containing eicosapentaenoic or docosahexaenoic acid on growth and metastasis of breast cancer cells in nude mice. J. Natl. Cancer Inst. 1995, 87, 587–592. [Google Scholar] [CrossRef]
- Zulyniak, M.A.; Perreault, M.; Gerling, C.; Spriet, L.L.; Mutch, D.M. Fish oil supplementation alters circulating eicosanoid concentrations in young healthy men. Metabolism 2013, 62, 1107–1113. [Google Scholar] [CrossRef]
- Te Chuang, L.; Thurmond, J.M.; Liu, J.W.; Kirchner, S.I.; Mukerji, P.; Bray, T.M.; Huang, Y.S. Effect of conjugated linoleic acid on fungal Δ6-desaturase activity in a transformed yeast system. Lipids 2001, 36, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, M.A.M.A.; Malpuech-Brugère, C.; Gregoire, S.; Chardigny, J.M.; Sébédio, J.L.; Mensink, R.P. Effects of specific CLA isomers on plasma fatty acid profile and expression of desaturases in humans. Lipids 2005, 40, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ren, X.L.; Fu, Y.Q.; Gao, J.L.; Li, D. Ratio of n-3/n-6 PUFAs and risk of breast cancer: A meta-analysis of 274135 adult females from 11 independent prospective studies. BMC Cancer 2014, 14, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nindrea, R.D.; Aryandono, T.; Lazuardi, L.; Dwiprahasto, I. Association of dietary intake ratio of n-3/n-6 polyunsaturated fatty acids with breast cancer risk in Western and Asian countries: A meta-analysis. Asian Pac. J. Cancer Prev. 2019, 20, 1321–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straka, S.; Lester, J.L.; Cole, R.M.; Andridge, R.R.; Puchala, S.; Rose, A.M.; Clinton, S.K.; Belury, M.A.; Yee, L.D. Incorporation of eicosapentaenioic and docosahexaenoic acids into breast adipose tissue of women at high risk of breast cancer: A randomized clinical trial of dietary fish and n-3 fatty acid capsules. Mol. Nutr. Food Res. 2015, 59, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Maillard, V.; Bougnoux, P.; Ferrari, P.; Jourdan, M.L.; Pinault, M.; Lavillonnìre, F.; Body, G.; Le Floch, O.; Chajès, V. n-3 and n-6 fatty acids in breast adipose tissue and relative risk of breast cancer in a case-control study in tours, France. Int. J. Cancer 2002, 98, 78–83. [Google Scholar] [CrossRef]
- Pizato, N.; Kiffer, L.F.M.V.; Luzete, B.C.; Assumpção, J.A.F.; Correa, L.H.; De Melo, H.A.B.; De Sant’Ana, L.P.; Ito, M.K.; Magalhães, K.G. Omega 3-DHA and delta-tocotrienol modulate lipid droplet biogenesis and lipophagy in breast cancer cells: The impact in cancer aggressiveness. Nutrients 2019, 11, 1199. [Google Scholar] [CrossRef] [Green Version]
- Chas, M.; Goupille, C.; Arbion, F.; Bougnoux, P.; Pinault, M.; Jourdan, M.L.; Chevalier, S.; Ouldamer, L. Low eicosapentaenoic acid and gamma-linolenic acid levels in breast adipose tissue are associated with inflammatory breast cancer. Breast 2019, 45, 113–117. [Google Scholar] [CrossRef]
- Serini, S.; Cassano, R.; Trombino, S.; Calviello, G. Nanomedicine-based formulations containing ω-3 polyunsaturated fatty acids: Potential application in cardiovascular and neoplastic diseases. Int. J. Nanomed. 2019, 14, 2809–2828. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Essential fatty acids enhance free radical generation and lipid peroxidation to induce apoptosis of tumor cells. Clin. Lipidol. 2011, 6, 463–489. [Google Scholar] [CrossRef]





| Fatty acids | CONplus | Mplus | Gplus | GMplus | p-Value |
|---|---|---|---|---|---|
| C14:0 | 0.24 ± 0.09 a,b | 0.30 ± 0.05 | 0.43 ± 0.10 a | 0.44 ± 0.06 b | 0.0052 |
| C15:0 | 0.25 ± 0.06 a,b | 0.28 ± 0.03 | 0.65 ± 0.36 a | 0.50 ± 0.13 b | 0.0016 |
| C16:0 | 16.0 ± 2.2 | 18.1 ± 2.9 | 19.6 ± 3.4 | 18.9 ± 2.4 | n.s. |
| C17:0 | 0.64 ± 0.05 | 0.57 ± 0.13 | 0.68 ± 0.17 | 0.68 ± 0.13 | n.s. |
| C18:0 | 17.2 ± 2.0 a,b | 13.7 ± 2.8 | 11.6 ± 3.1 a | 12.2 ± 1.6 b | 0.0125 |
| C21:0 | 0.09 ± 0.01 a | 0.08 ± 0.01 b | 0.13 ± 0.04 | 0.15 ± 0.04 a,b | 0.0072 |
| C24:0 | 0.41 ± 0.09 | 0.58 ± 0.03 | 0.41 ± 0.09 | 0.55 ± 0.09 | 0.0091 |
| SFA | 33.9 ± 1.4 | 33.8 ± 0.3 | 33.6 ± 1.2 | 33.6 ± 1.3 | n.s. |
| C16:1 | 1.27 ± 0.70 | 1.14 ± 0.22 | 2.23 ± 1.15 | 1.60 ± 0.21 | 0.0266 |
| C17:1, c-Δ10 | 0.24 ± 0.13 | 0.17 ± 0.04 | 0.37 ± 0.22 | 0.30 ± 0.10 | n.s. |
| C18:1, c-Δ9 | 8.46 ± 1.91 | 7.23 ± 0.34 | 11.0 ± 3.7 | 9.42 ± 1.89 | n.s. |
| C20:1, c-Δ11 | 0.07 ± 0.03 | 0.07 ± 0.03 | 0.09 ± 0.02 | 0.10 ± 0.03 | n.s. |
| MUFA | 10.1 ± 2.7 | 10.4 ± 2.9 | 13.7 ± 4.9 | 11.6 ± 2.2 | n.s. |
| C18:2, c,c-Δ9,12 (LA) | 11.1 ± 1.8 a,b | 11.9 ± 2.7 | 17.5 ± 2.8 a | 18.0 ± 2.4 b | 0.0022 |
| 6C18:3, c,c,c-Δ6,9,12 (GLA) | 0.31 ± 0.02 | 0.36 ± 0.07 | 0.33 ± 0.06 | 0.33 ± 0.13 | n.s. |
| C18:3, c,c,c-Δ9,12,15 (ALA) | 0.28 ± 0.14 a | 0.25 ± 0.05 | 0.73 ± 0.30 a | 0.62 ± 0.20 | 0.0068 |
| C18:2, c,t-Δ9,11 (RA) | 0.05 ± 0.06 | - | 0.08 ± 0.02 | 0.12 ± 0.07 | n.s. |
| C20:2, c,c-Δ11,14 | 0.07 ± 0.02 a | 0.10 ± 0.04 | 0.19 ± 0.10 a | 0.13 ± 0.06 | 0.0170 |
| C20:3, c,c,c-Δ8,11,14 | 0.37 ± 0.04 | 0.29 ± 0.07 | 0.41 ± 0.06 | 0.44 ± 0.18 | n.s. |
| C20:4, c,c,c-Δ5,8,11,14 (AA) | 29.8 ± 4.2 a | 29.3 ± 2.4 | 19.5 ± 6.2 a | 21.1 ± 4.0 | 0.0127 |
| C20:3, c,c,c-Δ11,14,17 | 0.27 ± 0.08 | 0.21 ± 0.00 | 0.39 ± 0.22 | 0.48 ± 0.38 | n.s. |
| C20:5, c,c,c,c,c-Δ5,8,11,14,17 (EPA) | 0.29 ± 0.13a | 0.23 ± 0.07b | 0.53 ± 0.15 a,b | 0.42 ± 0.08 | 0.0111 |
| C22:2, c,c-Δ13,16 | 0.06 ± 0.02 | 0.10 ± 0.05 | 0.11 ± 0.04 | 0.07 ± 0.03 | 0.0383 |
| C22:5, c,c,c,c,c-Δ7,10,13,16,19 | 0.35 ± 0.03 a | 0.44 ± 0.03 | 0.55 ± 0.19 | 0.63 ± 0.14 a | 0.0046 |
| C22:6, c,c,c,c,c,c-Δ4,7,10,13,16,19 (DHA) | 5.18 ± 0.97 a | 4.39 ± 0.32 | 3.21 ± 1.09 a | 3.65 ± 0.69 | 0.0123 |
| PUFA | 48.0 ± 3.2 | 46.1 ± 2.8 | 43.7 ± 5.4 | 45.8 ± 2.7 | n.s. |
| n3PUFA | 5.95 ± 0.66 | 5.10 ± 0.08 | 4.87 ± 1.03 | 5.35 ± 0.77 | n.s. |
| n6PUFA | 41.9 ± 2.7 | 40.8 ± 2.8 | 38.4 ± 4.8 | 40.5 ± 2.6 | n.s. |
| n6/n3 | 7.08 ± 0.55 | 8.00 ± 0.60 | 8.09 ± 1.31 | 7.63 ± 1.11 | n.s. |
| LOX Metabolites | CONplus | Mplus | Gplus | GMplus | p-Value |
|---|---|---|---|---|---|
| HODE | 42.9 ± 2.6 a | 258 ± 37 a,b | 88.0 ± 10.5 a | 58.9 ± 5.1 b | <0.0001 |
| 15-HETE | 8.9 ± 1.2 | 10.4 ± 1.9 | 7.5 ± 1.0 | 8.3 ± 1.3 | n.s. |
| 12-HETE | 316 ± 53 | 284 ± 41 | 391 ± 73 | 492 ± 89 | n.s. |
| 5-HETE | 8.1 ± 0.6 a | 20.1 ± 2.4b,c | 4.2 ± 0.4 a,b | 5.3 ± 0.9 c | <0.0001 |
| 12-HETE/15-HETE ratio | 38.5 ± 6.5 | 34.2 ± 5.8 | 55.2 ± 9.1 | 55.1 ± 10.3 | n.s. |
| 5-HETE/15-HETE ratio | 1.9 ± 0.6 | 2.4 ± 0.4 a,b | 1.1 ± 0.4 a | 0.7 ± 0.1 b | 0.0081 |
| Fatty Acids | CONplus | Mplus | Gplus | GMplus | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Median | Min–max | Median | Min–max | Median | Min–max | Median | Min–max | ||
| ΣFA | 7717 | (3198–31,692) | 10697 | (3404–25,781) | 4435 | (3181–29,080) | 7632 | (3049–51,091) | n.s. |
| C8:0 | 14.5 | (14.3–14.6) | 8.34 | (2.47–28.5) | 11.6 | (6.70–25.2) | 10.2 | (2.57–43.5) | n.s. |
| C10:0 | 29.7 | (26.8–32.6) | 6.83 | (2.50–234) | 13.8 | (3.97–37.1) | 12.6 | (2.89–158) | n.s. |
| C12:0 | 19.5 | (93.7–36.6) | 16.5 | (2.44–290) | 18.4 | (3.06–58.8) | 19.9 | (4.46–78.6) | n.s. |
| C14:0 | 85.8 | (293–360) | 139 | (39.3–429) | 67.1 | (26.2–370) | 122 | (31.3–660) | n.s. |
| C15:0 | 30.3 | (12.6–132) | 33.7 | (11.0–106) | 20.8 | (7.87–116) | 28.8 | (4.61–207) | n.s. |
| C16:0 | 1864 | (834–7223) | 2593 | (942–7147) | 1296 | (725–7248) | 2080 | (736–12,240) | n.s. |
| C17:0 | 57.2 | (26.0–264) | 49.3 | (23.8–191) | 29.6 | (11.8–177) | 42.6 | (13.6–363) | n.s. |
| C18:0 | 831 | (559–1937) | 855 | (632–1555) | 665 | (521–1521) | 786 | (553–2850) | n.s. |
| C20:0 | 12.8 | (9.15–27.4) | 13.9 | (6.81–30.1) | 17.4 a | (10.5–31.2) | 13.3a | (3.55–45.1) | 0.0427 |
| C21:0 | - | - | 9.40 | (1.39–16.8) | 6.87 | (1.87–12.5) | n.s. | ||
| C22:0 | 22.5 | (18.5–24.5) | 22.1 | (12.6–28.5) | 32.7 a | (19.5–48.3) | 8.03a | (1.35–57.6) | <0.0001 |
| C24:0 | 19.2 | (18.0–24.3) | 21.1 | (15.5–23.8) | 25.3 a | (16.8–42.5) | 7.75a | (1.74–60.1) | <0.0001 |
| SFA | 2921 | (1560–10,040) | 3859 | (1702–9096) | 2058 | (1528–9564) | 3084 | (1375–16,481) | n.s. |
| C16:1, c-Δ7 | 69.4 | (26.9–278) | 76.3 | (39.4–169) | 43.9 | (20.1–194) | 59.2 | (19.2–394) | n.s. |
| C16:1, c-Δ9 | 191 | (36.9–1353) | 501 | (49.7–2348) | 137 | (37.5–1665) | 356 | (47.6–3181) | n.s. |
| C17:1, c-Δ6 | - | - | 30.0 | (27.4–43.8) | - | n.s. | |||
| C17:1, c-Δ9 | - | - | 68.2 | (18.8–155) | 133.4 | (36.5–256) | n.s. | ||
| C18:1, t-Δ9 | 9.43 | (5.53–24.5) | 9.32 | (3.34–24.0) | 13.3 | (6.37–25.4) | 13.8 | (0.77–50.3) | n.s. |
| C18:1, c-Δ6 | 8.82 | (3.07–14.4) | 7.10 | (1.68–11.9) | 11.0 | (4.98–18.8) | 8.58 | (2.60–26.2) | n.s. |
| C18:1, c-Δ9 | 1838 | (463–9832) | 2720 | (462–7024) | 835 | (409–8162) | 1763 | (462–14,577) | n.s. |
| C18:1, c-Δ11 | 313 | (90.0–1583) | 368 | (90.4–910) | 164 | (83.9–1209) | 252 | (92.0–2262) | n.s. |
| C18:1, c-Δ12 | 9.66 | (3.99–14.9) | 8.76 | (3.06–15.2) | - | 8.63 | (2.73–25.4) | n.s. | |
| C18:1, c-Δ14 | - | - | 14.6 | (6.59–17.5) | 15.3 | (1.28–29.1) | n.s. | ||
| C20:1, c-Δ11 | 21.2 | (8.86–108.5) | 38.4 | (2.72–83.0) | 28.0 | (2.91–90.0) | 21.0 | (7.05–169.0) | n.s. |
| C24:1, c-Δ15 | 9.11 | (5.82–23.0) | 10.6a | (3.68–20.0) | 32.3a | (6.62–49.6) | 11.6 | (0.95–41.3) | 0.0447 |
| MUFA | 2457 | (658–13,231) | 3737 | (656–10,542) | 1282 | (576–11,535) | 2513 | (642–20,012) | n.s. |
| C18:2, c,c-Δ9,12 (LA) | 1331 | (163–6677) | 1734 | (131–5943) | 374 | (150–6361) | 768 | (145–11,915) | n.s. |
| C18:3, c,c,c-Δ6,9,12 (GLA) | 12.1 | (2.70–49.1) | 13.4 | (3.11–19.3) | 23.6 | (3.77–44.0) | 15.3 | (7.48–49.5) | n.s. |
| C18:3, c,c,c-Δ9,12,15 (ALA) | 180 | (43.7–291) | 77.2 | (16.0–256) | 90.4 | (6.74–376) | 64.9 | (6.40–699) | n.s. |
| C18:2, c,t-Δ9,11 (RA) | - | - | 53.2 | (25.0–97.2) | 33.6 | (2.65–252) | n.s. | ||
| C20:2, c,c-Δ11,14 | 13.9 | (4.51–51.3) | 24.7 | (6.47–41.5) | 17.4 | (6.14–46.6) | 16.2 | (2.39–78.3) | n.s. |
| C20:3, c,c,c-Δ8,11,14 | 28.5 | (22.7–50.3) | 33.3 | (22.5–47.5) | 40.3 | (30.7–64.2) | 37.2 | (17.5–91.5) | n.s. |
| C20:4, c,c,c,c-Δ5,8,11,14 (AA) | 828 | (684–1050) | 674a | (566–798) | 704b | (479–993) | 814a,b | (452–1946) | 0.0035 |
| C22:5, c,c,c,c,c-Δ7,10,13,16,19 (DPA) | 18.0 | (9.82–40.1) | 22.5 | (14.5–38.9) | 28.7 | (18.1–59.1) | 27.1 | (6.91–120.3) | n.s. |
| C22:6, c,c,c,c,c,c-Δ4,7,10,13,16,19 (DHA) | 107 | (75.7–186) | 91.7 | (65.1–144) | 95.2 | (51.6–161) | 101 | (61.7–323) | n.s. |
| PUFA | 2329 | (970 - 8395) | 2584 | (1040–7165) | 1162 | (945–7961) | 1872 | (818–14,530) | n.s. |
| n3PUFA | 150 | (85.6–517) | 158 | (107–439) | 128 | (91.7–567) | 149 | (78.9–1024) | n.s. |
| n6PUFA | 2176 | (885–7828) | 2424 | (933–6706) | 1027 | (854–7298) | 1707 | (727–13,205) | n.s. |
| n6/n3 | 12.6 | (8.24–15.1) | 13.9a,b | (8.73–17.7) | 9.60a | (5.80–16.6) | 11.4b | (6.48–15.3) | 0.0160 |
| Conjugated Fatty Acids | CONplus | Mplus | Gplus | GMplus | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Median | (Min–max) | Median | (Min–max) | Median | (Min–max) | Median | (Min–max) | p-Value | ||
| CFA | 18.7 a,b | (10.6–39.5) | 23.8 c,d | (17.0–60.5) | 86.3 a,c | (37.0–567) | 104 b,d | (27.2–954) | <0.0001 | |
| CD | 15.5 a,b | (8.48–37.2) | 20.0 c,d | (14.5–56.5) | 65.3 a,c | (29.7–544) | 68.5 b,d | (22.6–951) | 0.0001 | |
| tt | 7.97 a,b | (5.12–12.3) | 9.50 c,d | (6.28–19.4) | 38.0 a,c | (14.8–95.7) | 30.0 b,d | (12.4–154) | <0.0001 | |
| ct | 4.64 | (2.17–24.8) | 9.81 | (2.21–28.1) | 19.4 | (2.15–433) | 35.2 | (2.80–774) | 0.0192 | |
| cc | 2.93 a | (0.47–3.41) | 3.92 b | (0.25–9.07) | 10.0 a,b,c | (3.33–32.1) | 4.91c | (0.66–22.1) | 0.0005 | |
| c9t11CLA | 3.74 | (0.23–20.3) | 5.94 a | (0.62–20.9) | 19.4 | (2.15–391) | 31.4 a | (1.81–730) | 0.0060 | |
| CT | 2.15 a,b | (1.21–3.22) | 5.67 c | (2.23–7.24) | 16.0 a,c | (2.78–32.4) | 10.2 b | (0.30–175) | 0.0003 | |
| ttt | 1.56 a,b | (0.30–2.29) | 5.45 c | (0.30–7.24) | 15.8 a,c | (3.37–32.4) | 7.40 b | (2.29–37.7) | <0.0001 | |
| ttc/ctt | 0.74 | (0.51–1.21) | 0.34 | (0.34–0.34) | 1.90 | (0.43–3.29) | 0.70 | (0.12–26.4) | n.s. | |
| cct | - | - | 0.87 | (0.32–2.36) | 1.68 | (0.19–8.04) | 9.26 | (0.25–149) | n.s. | |
| Conjugated Fatty Acids | CONplus | Mplus | Gplus | GMplus | p-Value | |
|---|---|---|---|---|---|---|
| CD [%CFA] | 88.0 ± 7.2 | 82.2 ± 7.1 | 82.1 ± 11.1 | 79.2 ± 22.8 | n.s. | |
| tt [%CD] | 45.1 ± 13.7 | 45.5 ± 12.5 | 47.2 ± 18.4 | 48.4 ± 25.5 | n.s. | |
| ct [%CD] | 43.3 ± 17.6 | 37.4 ± 16.0 | 40.1 ± 22.5 | 50.5 ± 24.9 | n.s. | |
| cc [%CD] | 11.6 ± 6.7 | 17.1 ± 8.6 a | 13.6 ± 7.7 b | 4.06 (1.98–8.35) *a,b | 0.0002 | |
| c9t11CLA [%CD] | 29.7 ± 20.5 | 24.8 ± 15.5 | 33.5 ± 21.0 | 41.2 ± 24.7 | n.s. | |
| c9t11CLA [%ct] | 61.2 ± 29.8 | 62.1 ± 20.1 a | 80.8 ± 22.8 | 83.7 ± 13.2 a | 0.0104 | |
| CT [%CFA] | 12.0 ± 7.2 | 17.8 ± 7.1 | 17.9 ± 11.1 | 11.3 (3.3–38.5) * | n.s. | |
| ttt [%CT] | 70.3 ± 3.7 a | 85.5 ± 17.4 | 86.5 ± 12.7 | 71.9 ± 30.2 a | 0.0394 | |
| ttc/ctt [%CT] | 43.8 ± 31.6 | 4.87 ± 0.01 | 13.7 (4.6–40.6) * | 8.36 (2.52–27.7) * | n.s. | |
| cct [%CT] | - | 42.2 ± 34.9 | 19.5 ± 13.1 | 57.0 ± 30.1 | n.s. | |
| LOX Metabolites | CONplus | Mplus | Gplus | GMplus | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Median | Min–max | Median | Min–max | Median | Min–max | Median | Min–max | ||
| HODE | 354 | (175–747) | 312 a | (217–1098) | 688a | (295–2280) | 627 | (294–2398) | 0.0140 |
| 15-HETE | 575 | (355–1684) | 536 | (258–1128) | 879 | (256 – 2182) | 765 | (284–2343) | n.s. |
| 12-HETE | 445 | (270–539) | 495 | (236–2218) | 637 | (238–1075) | 430 | (144–6006) | n.s. |
| 5-HETE | 172 | (62.0–569) | 130 | (80.9–576) | 137 | (59.4–362) | 189 | (34.6–710) | n.s. |
| 12-HETE/15-HETE ratio | 0.8 | (0.3–0.9) | 1.0 | (0.4–8.6) | 0.6 | (0.3–1.5) | 0.6 | (0.3–3.9) | n.s. |
| 5-HETE/15-HETE ratio | 0.3 | (0.2–0.6) | 0.3 | (0.1–1.2) | 0.2 | (0.1–0.4) | 0.2 | (0.3–0.9) | n.s. |
| Compounds | CONplus | Mplus | Gplus | GMplus | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Median | (Min–max) | Median | (Min–max) | Median | (Min–max) | Median | (Min–max) | ||
| squalene | 112 | (32.7–204) | 71.7 | (33.0–272) | 62.8 | (15.9–741) | 47.3 | (8.78–207) | n.s. |
| cholesterol | 257 | (10.6–561) | 201 | (3.81–1283) | 409 | (61.6–3867) | 365 | (12.0–1502) | n.s. |
| cholesterol/squalene | 2.16a | (0.13–17.1) | 2.99 b | (0.10–7.60) | 5.83 | (0.33–43.6) | 8.20 a,b | (0.21–48.6) | 0.0040 |
| 7α-OH-Ch | 1.16 | (0.28–5.93) | 0.73 | (0.20–1.35) | 1.21 | (0.53–8.14) | 0.75 | (0.16–6.06) | n.s. |
| 7ß-OH-Ch | 3.06 | (1.00–4.17) | 3.44 | (2.00–9.32) | 3.66 | (2.37–54.6) | 3.09 | (2.09–20.0) | n.s. |
| 5α,6α-epoxy-Ch | 3.73 | (3.22–13.3) | 4.54 | (1.66–6.63) | 4.56 | (2.86–77.1) | 3.68 | (2.54–13.0) | n.s. |
| 25-OH-Ch | 0.08 | (0.06–3.27) | 0.14 | (0.11–1.22) | 0.16 | (0.06–4.56) | 0.14 | (0.06–0.47) | n.s. |
| 7-keto-Ch | 5.55 | (3.60–38.9) | 6.61 | (2.51–20.4) | 9.56 | (3.21–140) | 6.28 | (2.11–65.5) | n.s. |
| Σ oxy-Ch | 14.9 | (8.29–62.2) | 16.1 | (8.39–32.1) | 19.1 | (9.17–276) | 13.9 | (7.92–88.2) | n.s. |
| Actual Group | Number of Cases | Predicted Group Membership | |||
|---|---|---|---|---|---|
| CONplus | Mplus | Gplus | GMplus | ||
| CONplus | 5 | 4 80.0% | 1 20.0% | 0 0% | 0 0% |
| Mplus | 9 | 0 0% | 9 100% | 0 0% | 0 0% |
| Gplus | 16 | 0 0% | 0 0% | 15 93.8% | 1 6.2% |
| GMplus | 36 | 0 0% | 0 0% | 0 0% | 36 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Białek, A.; Jelińska, M.; Białek, M.; Lepionka, T.; Czerwonka, M.; Czauderna, M. The Effect of Diet Supplementation with Pomegranate and Bitter Melon on Lipidomic Profile of Serum and Cancerous Tissues of Rats with Mammary Tumours. Antioxidants 2020, 9, 243. https://doi.org/10.3390/antiox9030243
Białek A, Jelińska M, Białek M, Lepionka T, Czerwonka M, Czauderna M. The Effect of Diet Supplementation with Pomegranate and Bitter Melon on Lipidomic Profile of Serum and Cancerous Tissues of Rats with Mammary Tumours. Antioxidants. 2020; 9(3):243. https://doi.org/10.3390/antiox9030243
Chicago/Turabian StyleBiałek, Agnieszka, Małgorzata Jelińska, Małgorzata Białek, Tomasz Lepionka, Małgorzata Czerwonka, and Marian Czauderna. 2020. "The Effect of Diet Supplementation with Pomegranate and Bitter Melon on Lipidomic Profile of Serum and Cancerous Tissues of Rats with Mammary Tumours" Antioxidants 9, no. 3: 243. https://doi.org/10.3390/antiox9030243
APA StyleBiałek, A., Jelińska, M., Białek, M., Lepionka, T., Czerwonka, M., & Czauderna, M. (2020). The Effect of Diet Supplementation with Pomegranate and Bitter Melon on Lipidomic Profile of Serum and Cancerous Tissues of Rats with Mammary Tumours. Antioxidants, 9(3), 243. https://doi.org/10.3390/antiox9030243