Effects of Melatonin Administration to Pregnant Ewes under Heat-Stress Conditions, in Redox Status and Reproductive Outcome



 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
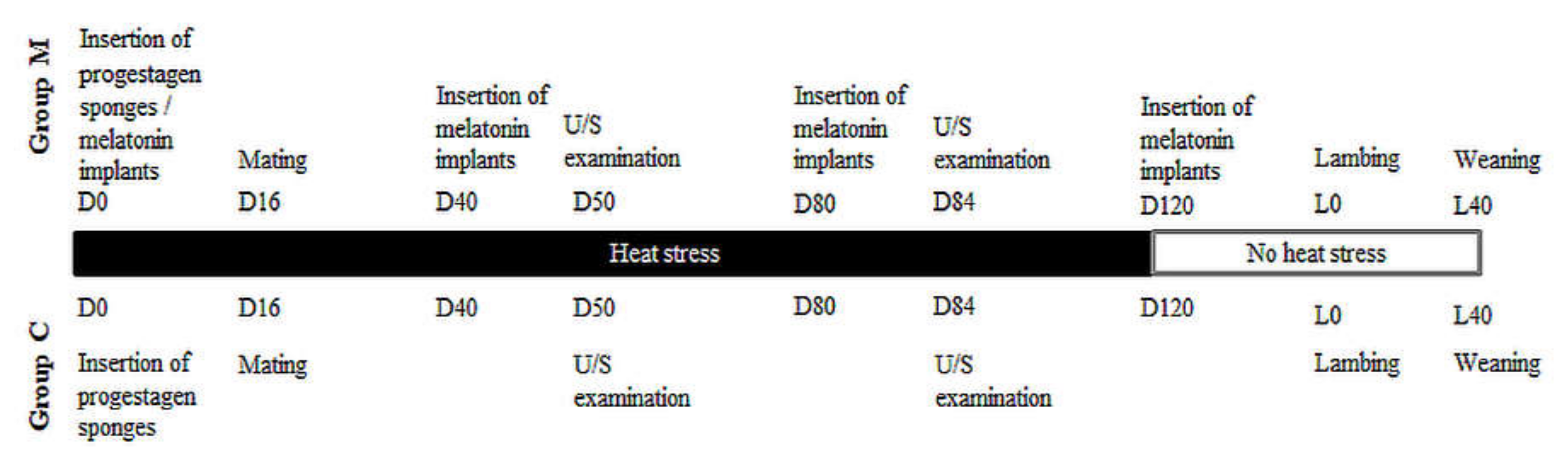
2.1. Experimental Overview—Animal Work
2.2. Laboratory Examinations
2.3. Data Management and Analysis
2.3.1. Temperature—Humidity Index
2.3.2. Measures for Reproductive Performance
- Mating rate: number of ewes marked by the ram during the two introductions into ewes/number of ewes exposed to the ram × 100.
- Pregnancy rate: number of ewes found pregnant at the ultrasonographic examination on D84/number of ewes exposed to the ram × 100.
- Lambing rate: number of ewes that lambed among those found pregnant at ultrasonographic examination on D84/number of ewes mated subsequently to application of reproductive control × 100.
- Total lambs per ewe: number of liveborn and stillborn lambs/number of ewes that lambed.
- Lamb bodyweight per ewe (kg): total weight (kg) of liveborn lambs/number of ewes that lambed.
2.3.3. Statistical Analysis
3. Results
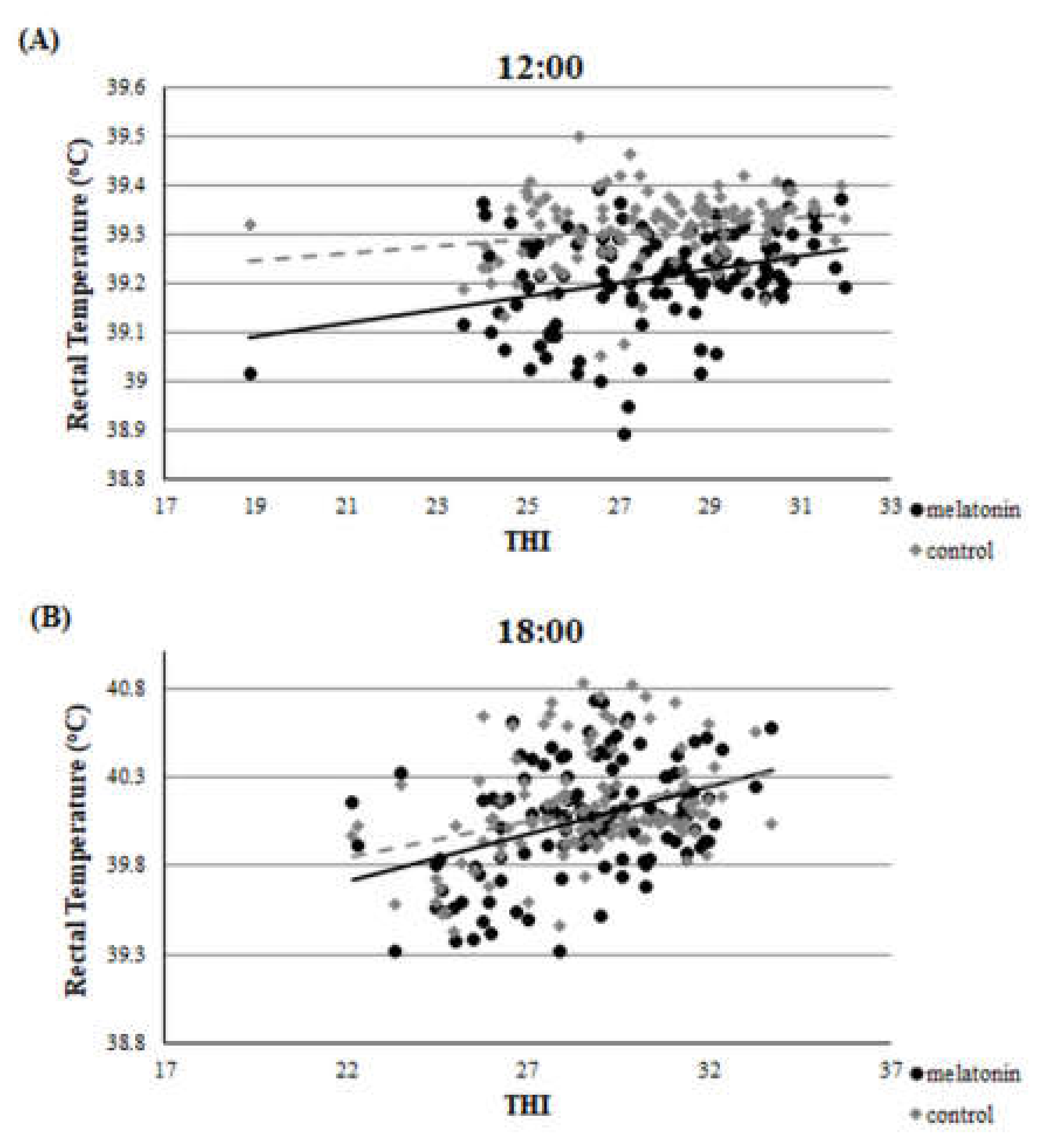
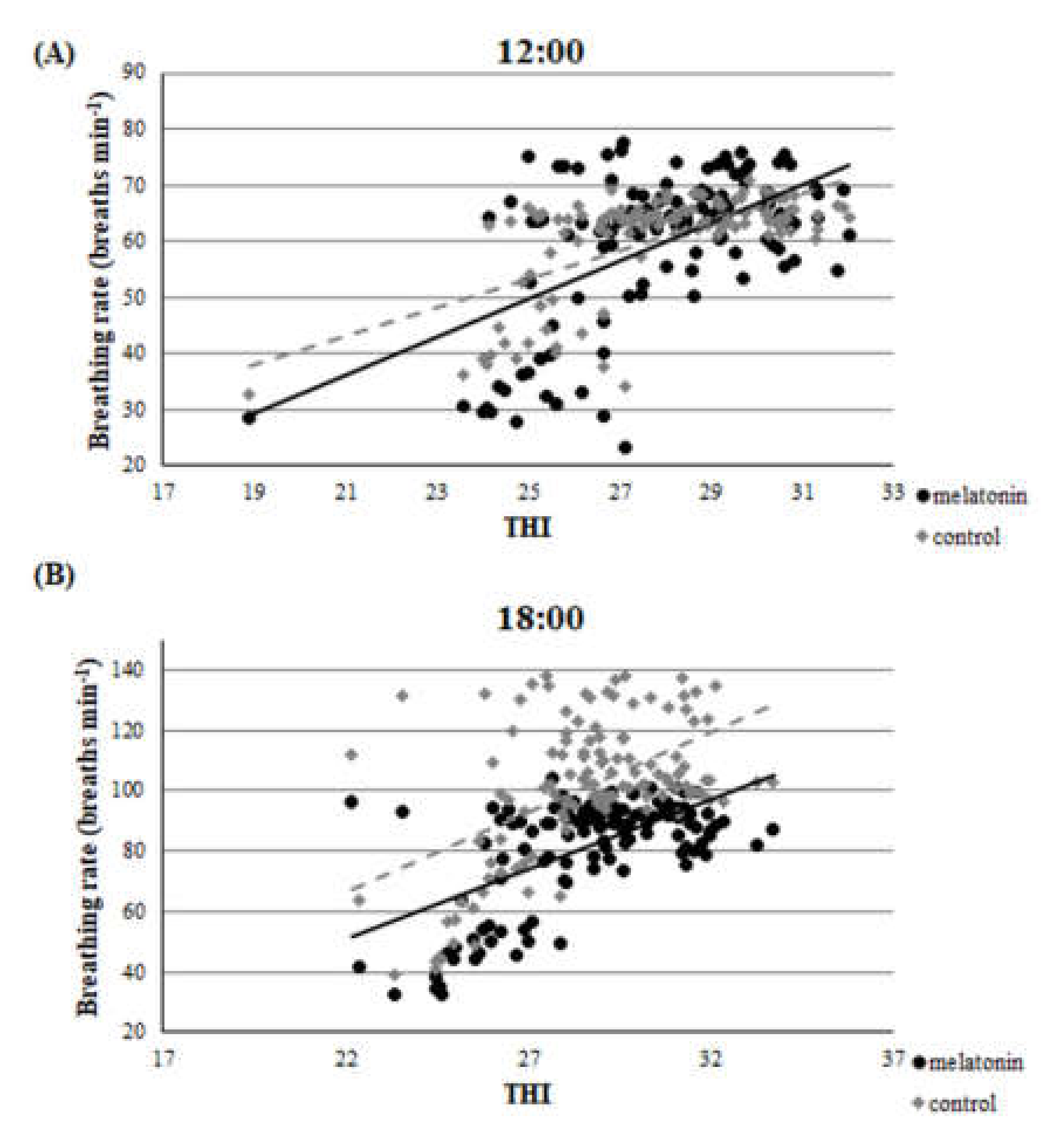
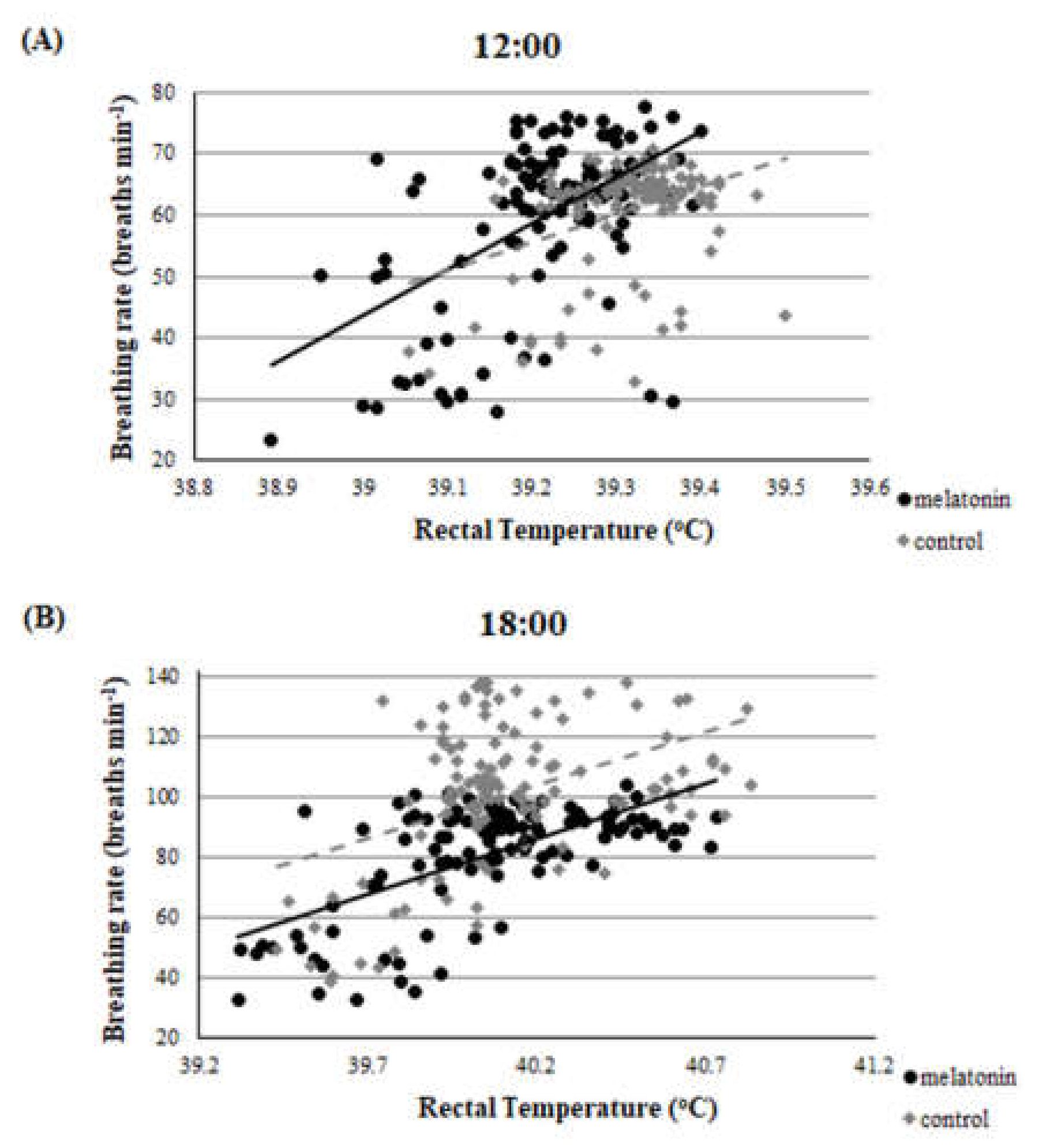
3.1. Clinical Findings—THI during the Heat Stress Period
3.2. Reproductive Performance
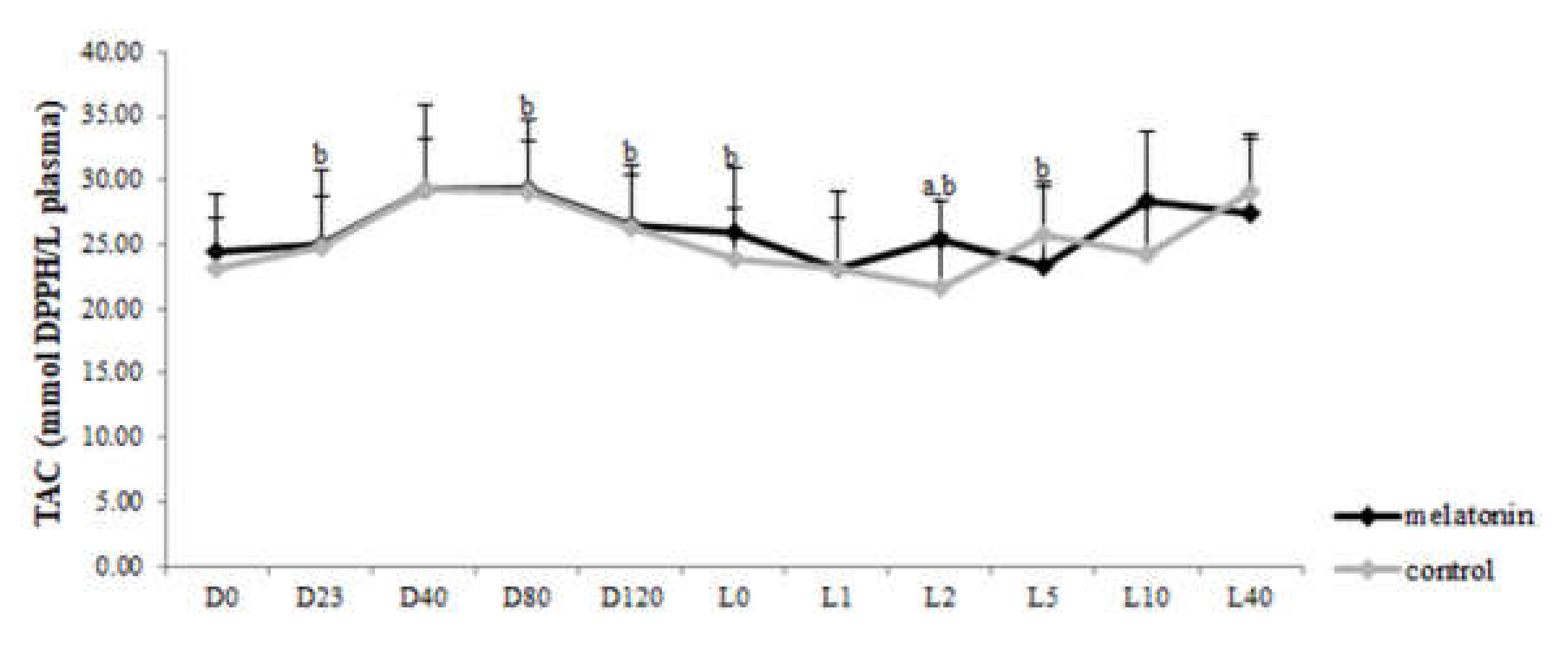
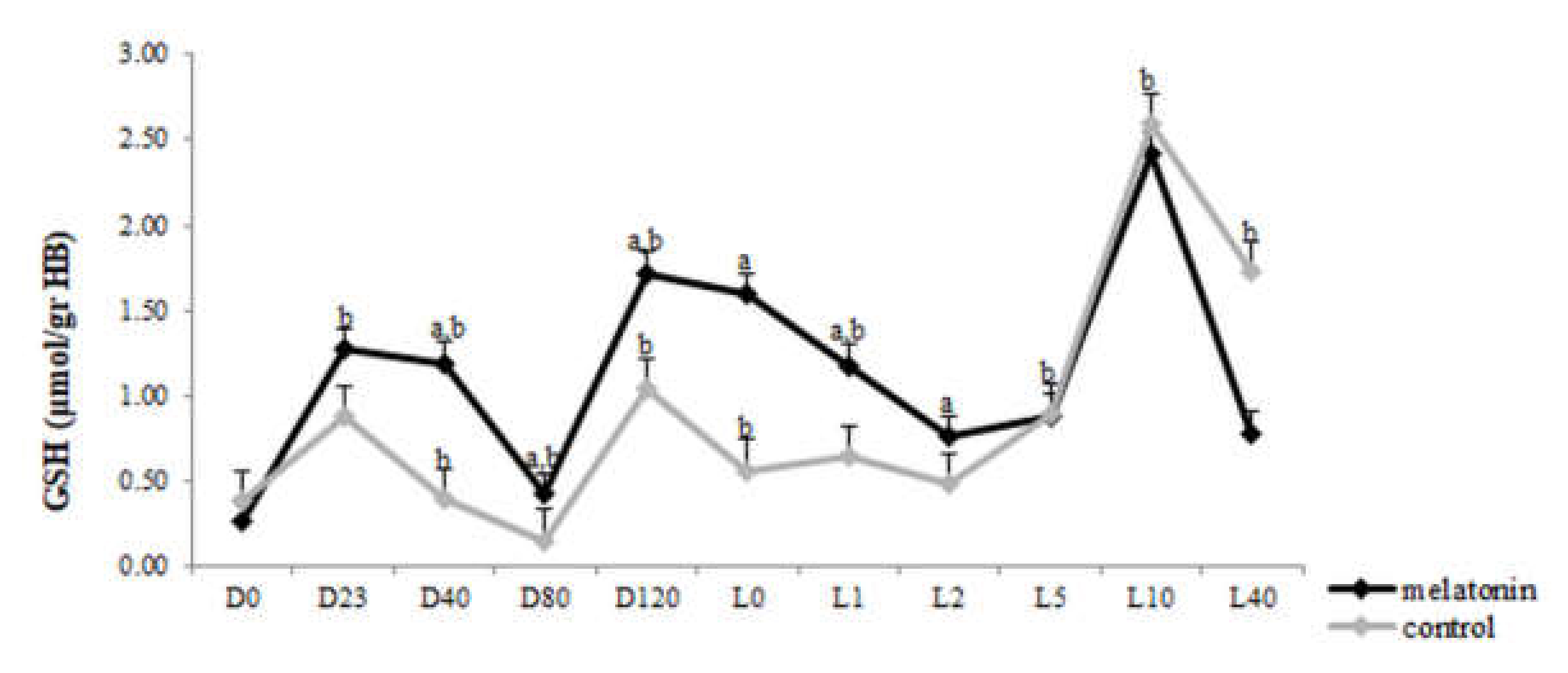
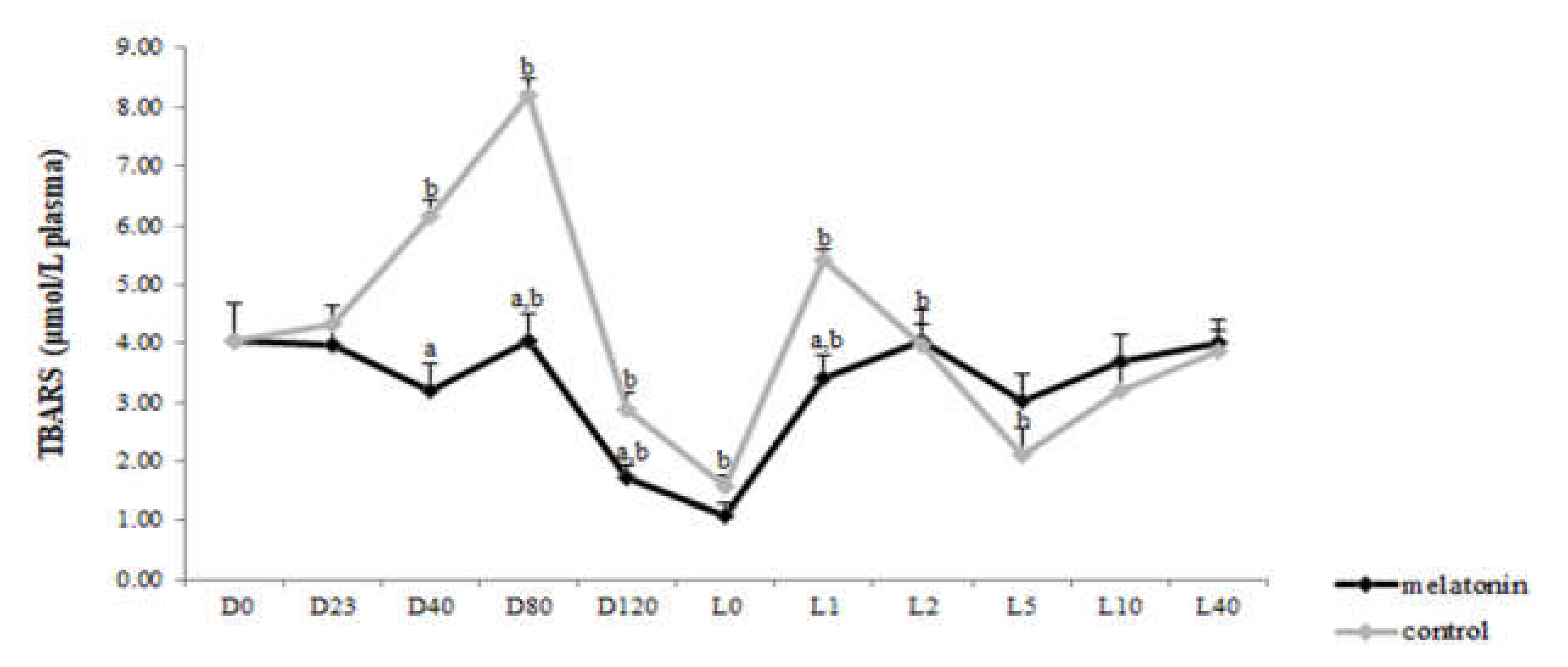
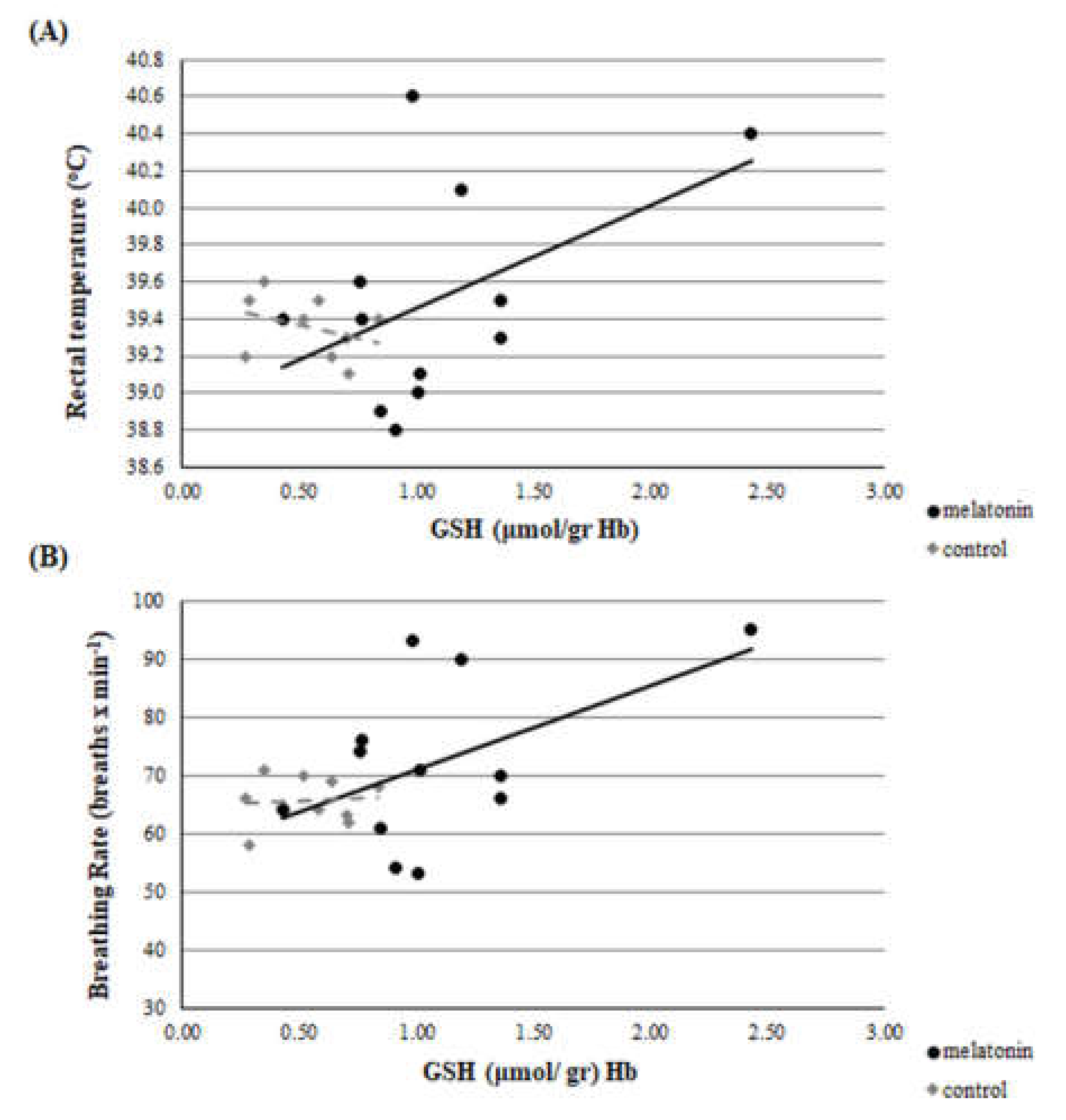
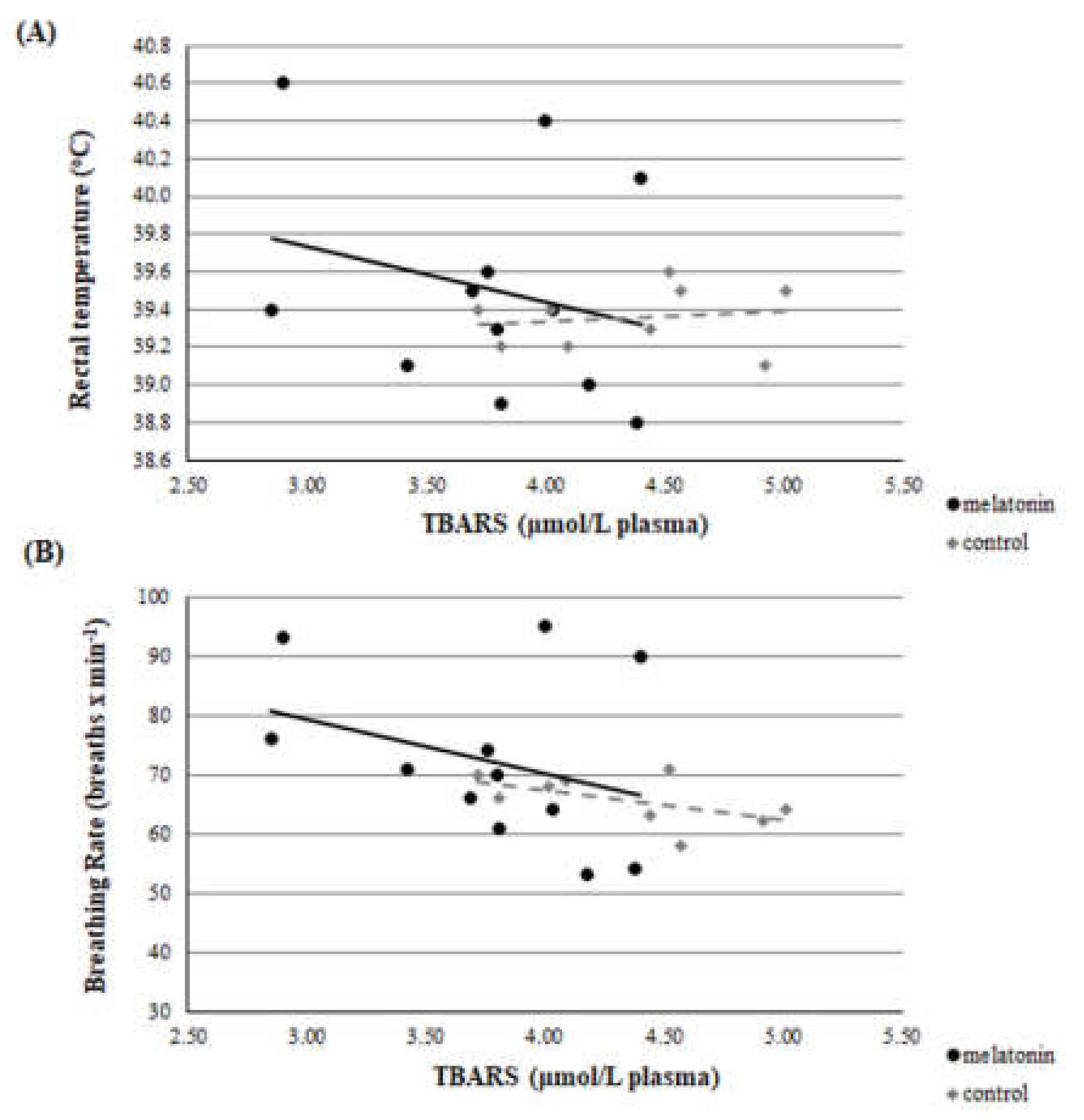
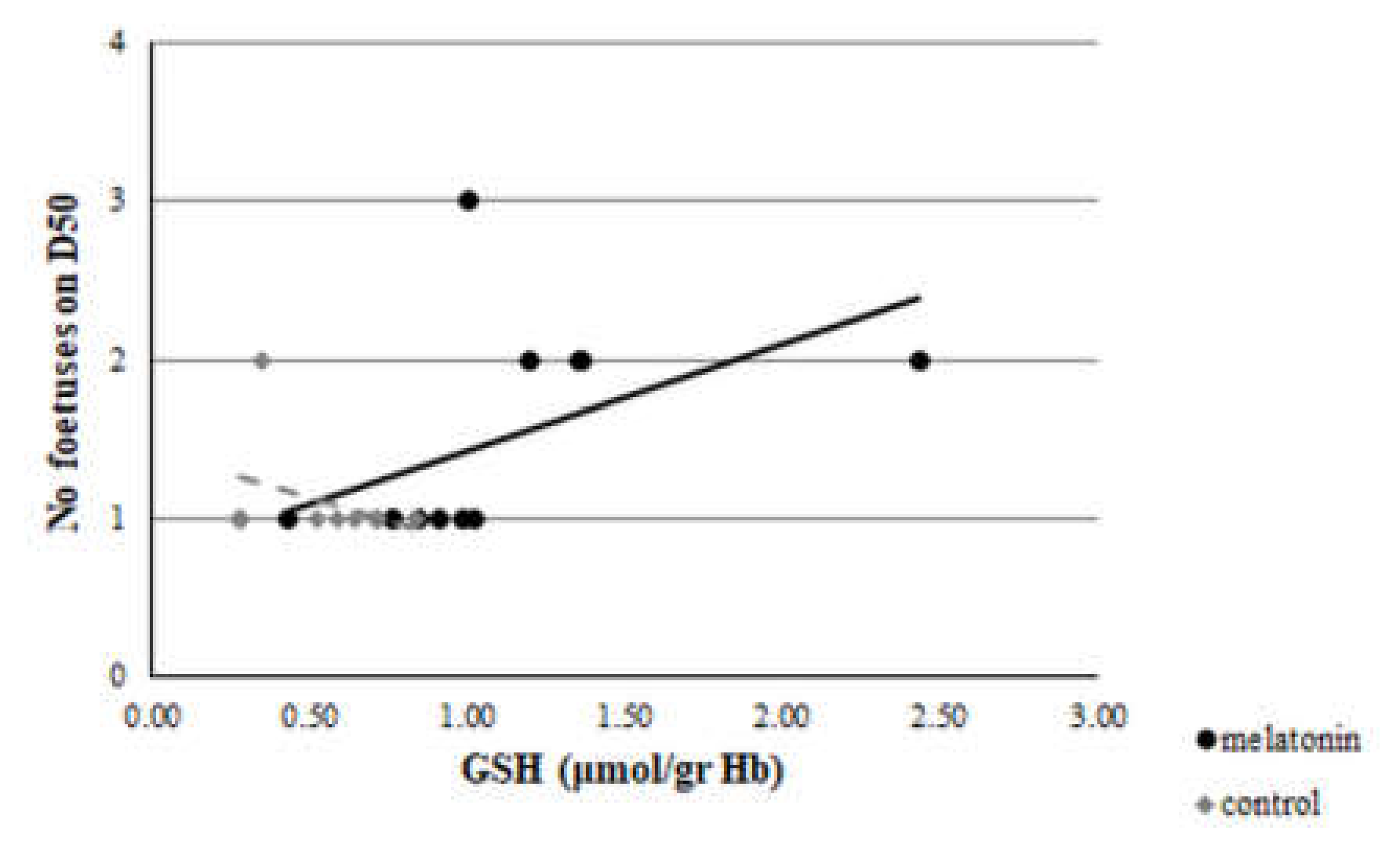
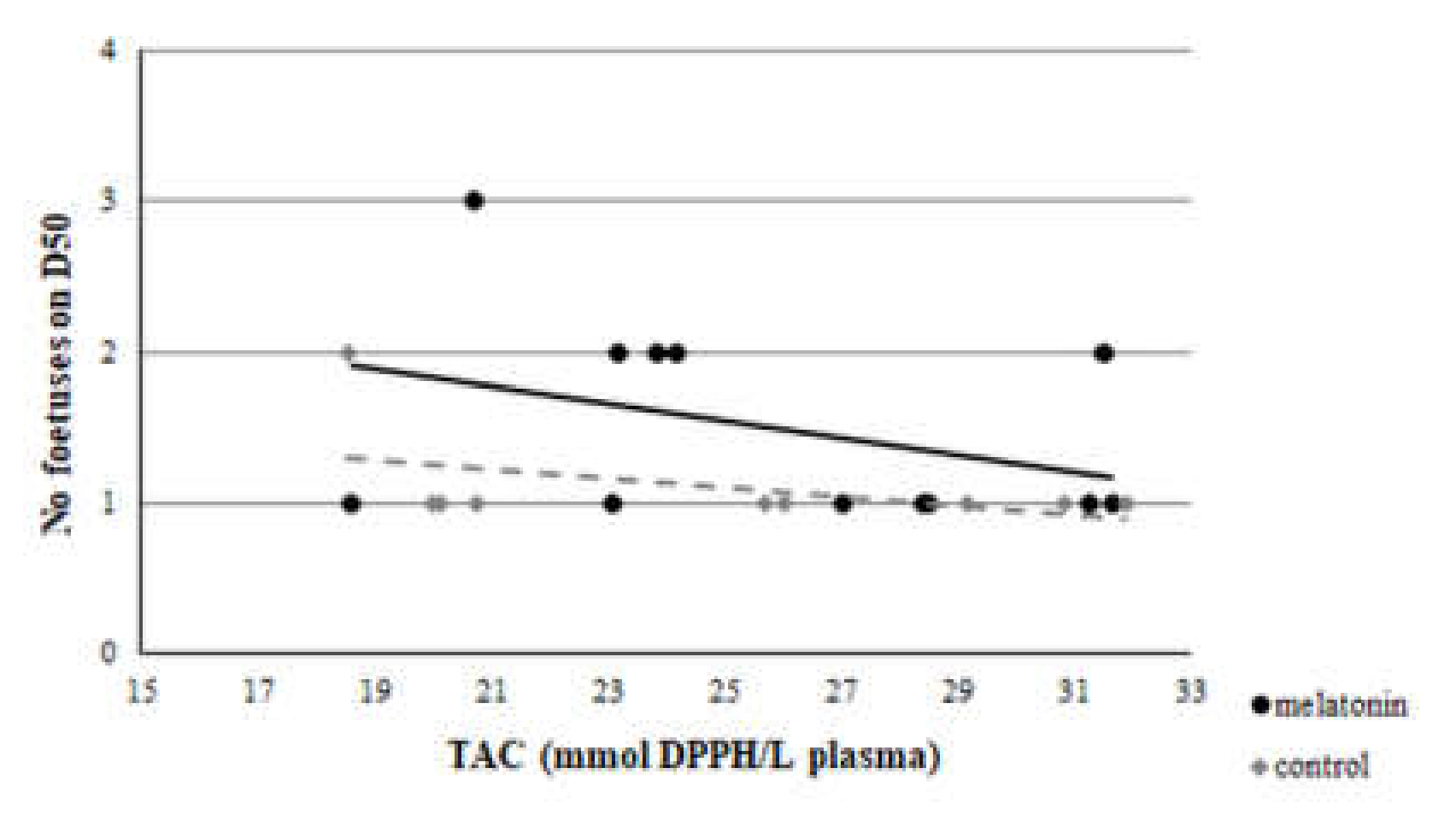
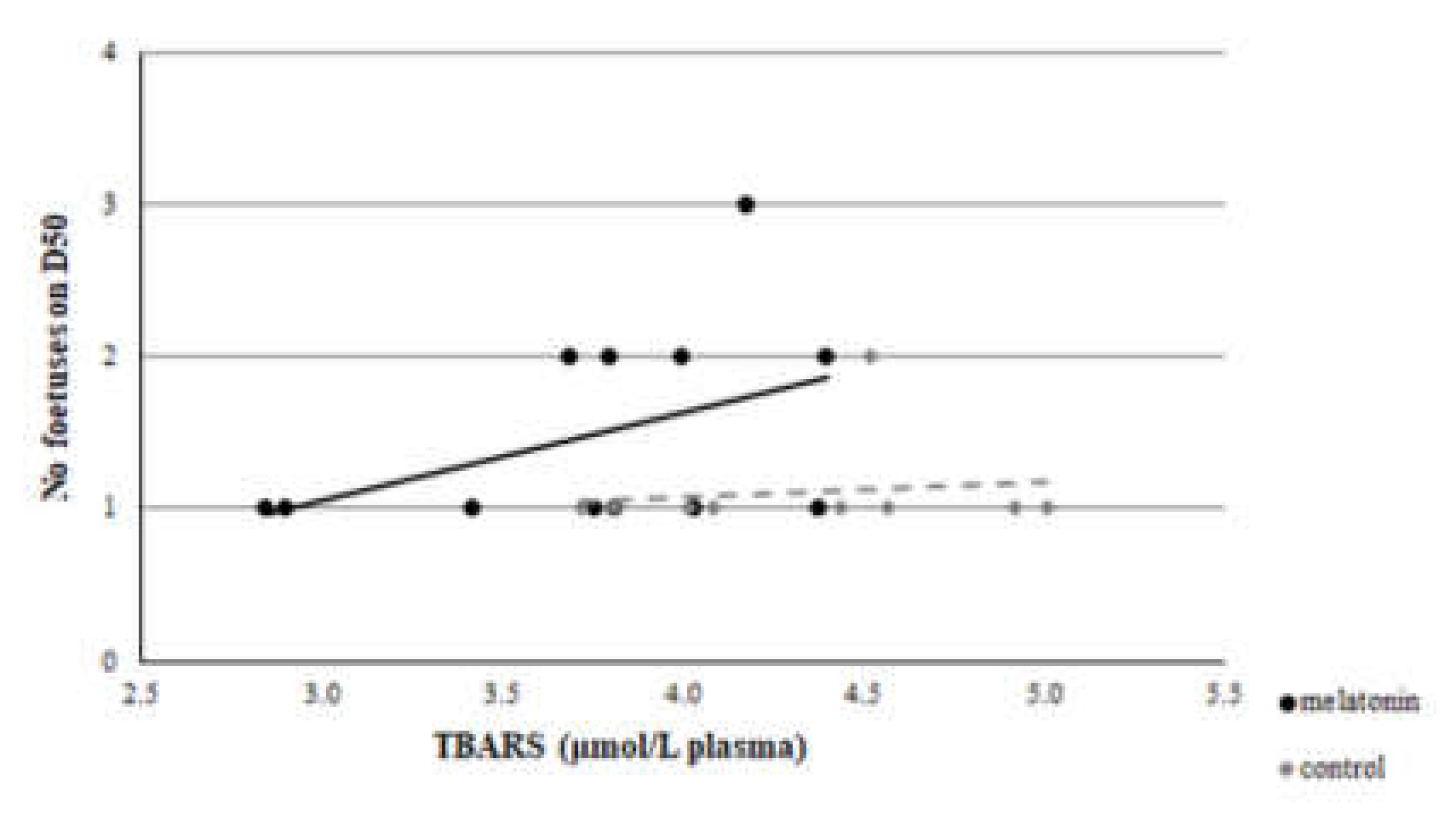
3.3. Redox Biomarkers
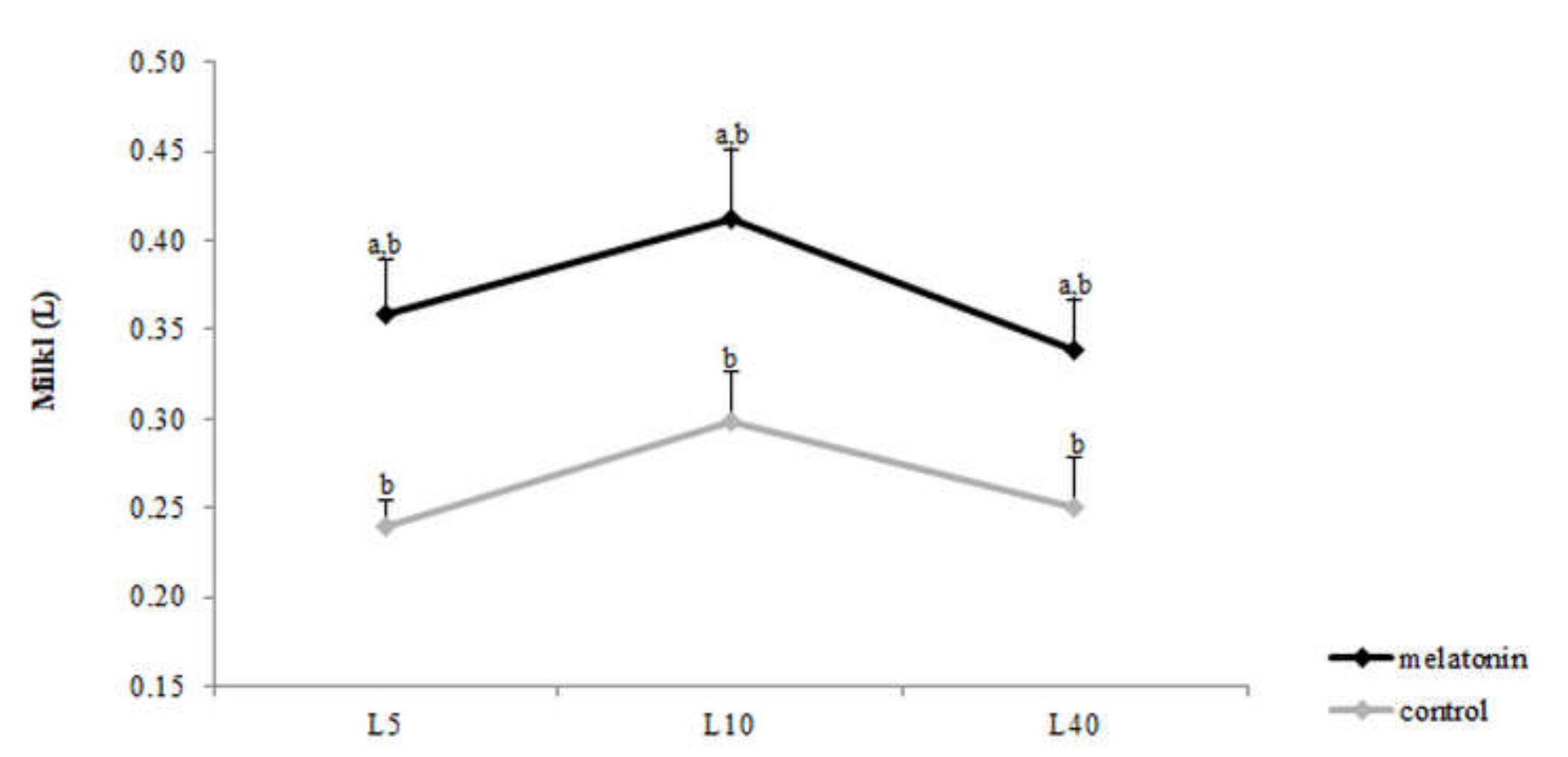
3.4. Milk Yield
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Garrel, C.; Fowler, P.A.; Al-Gubory, K.H. Developmental changes in antioxidant enzymatic defences against oxidative stress in sheep placentomes. J. Endocrinol. 2010, 205, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Ciliberti, M.G.; Albenzio, M.; Marino, R.; Santillo, A.; Sevi, A. Role of antioxidant molecules in milk of sheep. Small Rumin. Res. 2019, 180, 79–85. [Google Scholar] [CrossRef]
- Roth, Z.; Aroyo, A.; Yavin, S.; Arav, A. The antioxidant epigallocatechin gallate (EGCG) moderates the deleterious effects of maternal hyperthermia on follicle-enclosed oocytes in mice. Theriogenology 2008, 70, 887–897. [Google Scholar] [CrossRef]
- Lawrence, J.L.; Payton, R.R.; Godkin, J.D.; Saxton, A.M.; Schrick, F.N.; Edwards, J.L. Retinol improves development of bovine oocytes compromised by heat stress during maturation. J. Dairy Sci. 2004, 87, 2449–2454. [Google Scholar] [CrossRef]
- Mutinati, M.; Piccinno, M.; Roncetti, M.; Campanile, D.; Rizzo, A.; Sciorsci, R. Oxidative stress during pregnancy in the sheep. Reprod. Domest. Anim. 2013, 48, 353–357. [Google Scholar] [CrossRef]
- McDowell, R.E. Improvement of livestock production in warm climates. In Improvement of Livestock in Warm Climates; Cornell University: Ithaca, NY, USA, 1972; pp. 410–449. [Google Scholar]
- Bianca, W. Relative importance of dry- and wet-bulb temperatures in causing heat stress in cattle. Nature 1962, 195, 251–252. [Google Scholar] [CrossRef]
- Hayes, B.J.; Carrick, M.; Bowman, P.; Goddard, M.E. Genotype x environment interaction for milk production of daughters of Australian dairy sires from test-day records. J. Dairy Sci. 2003, 86, 3736–3744. [Google Scholar] [CrossRef]
- Finocchiaro, R.; Van Kaam, J.B.C.H.M.; Portolano, B.; Misztal, I. Effect of heat stress on production of mediterranean dairy sheep. J. Dairy Sci. 2005, 88, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Gaughan, J.; Cawdell-Smith, A.J. Impact of climate change on livestock production and reproduction. In Climate Change Impact on Livestock: Adaptation and Mitigation; Sejian, V., Gaughan, J., Baumgard, L., Prasad, C., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2015; pp. 51–60. [Google Scholar]
- Sejian, V.; Bhatta, R.; Gaughan, J.; Malik, P.K.; Naqvi, S.M.K.; Lal, R. Adapting Sheep Production to Climate Change. In Sheep Production Adapting to Climate Change; Sejian, V., Bhatta, R., Gaughan, J., Malik, P.K., Naqvi, S.M.K., Lal, R., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2017; Volume 1, pp. 1–29. [Google Scholar]
- Ozawa, M.; Hirabayashi, M.; Kanai, Y. Developmental competence and oxidative state of mouse zygotes heat-stressed maternally or in vitro. Reproduction 2002, 124, 683–689. [Google Scholar] [CrossRef]
- Matsuzuka, T.; Sakamoto, N.; Ozawa, M.; Ushitani, A.; Hirabayashi, M.; Kanai, Y. Alleviation of maternal hyperthermia-induced early embryonic death by administration of melatonin to mice. J. Pineal Res. 2005, 39, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Lorena, F.B. Melatonin: A multitasking molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [PubMed]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and Its Relation to the Immune System and Inflammation. Ann. N. Y. Acad. Sci. 2006, 917, 376–386. [Google Scholar] [CrossRef]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A pleiotropic molecule regulating inflammation. Biochem. Pharmacol. 2010, 80, 1844–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin-A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Abecia, J.A.; Forcada, F.; Casao, A.; Palacín, I. Effect of exogenous melatonin on the ovary, the embryo and the establishment of pregnancy in sheep. Animal 2008, 2, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.-X.; Maekawa, R.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar] [CrossRef]
- Casao, A.; Abecia, J.A.; Cebrián Pérez, J.A.; Muiño Blanco, T.; Vázquez, M.I.; Forcada, F. The effects of melatonin on in vitro oocyte competence and embryo development in sheep. Span. J. Agric. Res. 2010, 8, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Chemineau, P.; Malpaux, B.; Pelletier, J.; Leboeuf, B.; Delgadillo, J.A.; Deletang, F.; Pobel, T.; Brice, G. Emploi des implants de mélatonine et des traitements photopériodiques pour maîtriser la reproduction saisonnière chez les ovins et les caprins. Prod. Anim. 1996, 9, 45–60. [Google Scholar]
- Garcia-Ispierto, I.; Abdelfatah, A.; López-Gatius, F. Melatonin treatment at dry-off improves reproductive performance postpartum in high-producing dairy cows under heat stress conditions. Reprod. Domest. Anim. 2013, 48, 577–583. [Google Scholar] [CrossRef]
- Fthenakis, G.C.; Jones, J.E. The effect of inoculation of coagulase-negative Staphylococci into the ovine mammary gland. J. Comp. Pathol. 1990, 102, 211–219. [Google Scholar] [CrossRef]
- Fthenakis, G.C.; Papadopoulos, E.; Himonas, C. Effects of three anthelmintic regimes on milk yield of ewes and growth of lambs. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 2005, 52, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Veskoukis, A.S.; Kerasioti, E.; Priftis, A.; Kouka, P.; Spanidis, Y.; Makri, S.; Kouretas, D. A battery of translational biomarkers for the assessment of the in vitro and in vivo antioxidant action of plant polyphenolic compounds: The biomarker issue. Curr. Opin. Toxicol. 2019, 13, 99–109. [Google Scholar] [CrossRef]
- Janaszewska, A.; Bartosz, G. Assay of total antioxidant capacity: Comparison of four methods as applied to human blood plasma. Scand. J. Clin. Lab. Investig. 2002, 62, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kokkinos, D.; Nepka, C.; Barbanis, S.; Kouretas, D. Effects of xanthine oxidase inhibition on oxidative stress and swimming performance in rats. Appl. Physiol. Nutr. Metab. 2008, 33, 1140–1154. [Google Scholar] [CrossRef]
- Keles, M.S.; Taysi, S.; Sen, N.; Aksoy, H.; Akçay, F. Effect of corticosteroid therapy on serum and CSF malondialdehyde and antioxidant proteins in multiple sclerosis. Can. J. Neurol. Sci. 2001, 28, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Veskoukis, A.S.; Kyparos, A.; Nikolaidis, M.G.; Stagos, D.; Aligiannis, N.; Halabalaki, M.; Chronis, K.; Goutzourelas, N.; Skaltsounis, L.; Kouretas, D. The antioxidant effects of a polyphenol-rich grape pomace extract in vitro do not correspond in vivo using exercise as an oxidant stimulus. Oxid. Med. Cell. Longev. 2012, 2012, 185867. [Google Scholar] [CrossRef] [Green Version]
- Reddy, Y.N.; Murthy, S.; Krishna, D.R.; Prabhakar, M.C. Role of free radicals and antioxidants in tuberculosis patients. Indian J Tuberc. 2004, 51, 213–218. [Google Scholar]
- Veskoukis, A.S.; Kyparos, A.; Paschalis, V.; Nikolaidis, M.G. Spectrophotometric assays for measuring redox biomarkers in blood. Biomarkers 2016, 21, 208–217. [Google Scholar] [CrossRef]
- Marai, I.F.M.; Ayyat, M.S.; Abd El-Monem, U.M. Growth performance and reproductive traits at first parity of New Zealand White female rabbits as affected by heat stress and its alleviation under Egyptian conditions. Trop. Anim. Health Prod. 2001, 33, 451–462. [Google Scholar] [CrossRef]
- Habeeb, A.A.; Gad, A.E.; Atta, M.A. Temperature-Humidity Indices as Indicators to Heat Stress of Climatic Conditions with Relation to Production and Reproduction of Farm Animals. Int. J. Biotechnol. Recent Adv. 2018, 1, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Fthenakis, G.C. Effects of retention of foetal membranes on subsequent reproductive performance of dairy ewes. Theriogenology 2004, 61, 129–135. [Google Scholar] [CrossRef]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep-A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Carpentieri, A.; Díaz De Barboza, G.; Areco, V.; Peralta López, M.; Tolosa De Talamoni, N. New perspectives in melatonin uses. Pharmacol. Res. 2012, 65, 437–444. [Google Scholar] [CrossRef]
- Reiter, R.J. Oxidative processes and antioxidative defense mechanisms in the aging brain. FASEB J. 1995, 9, 526–533. [Google Scholar] [CrossRef]
- Kumar, A.; Mehrotra, S.; Singh, G.; Narayanan, K.; Das, G.; Soni, Y.; Singh, M.; Mahla, A.; Srivastava, N.; Verma, M. Sustained delivery of exogenous melatonin influences biomarkers of oxidative stress and total antioxidant capacity in summer-stressed anestrous water buffalo (Bubalus bubalis). Theriogenology 2015, 83, 1402–1407. [Google Scholar] [CrossRef]
- Tamura, H.; Nakamura, Y.; Terron, M.P.; Flores, L.J.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and pregnancy in the human. Reprod. Toxicol. 2008, 25, 291–303. [Google Scholar] [CrossRef]
- Sainz, R.M.; Reiter, R.J.; Mayo, J.C.; Cabrera, J.; Tan, D.X.; Qi, W.; Garcia, J.J. Changes in lipid peroxidation during pregnancy and after delivery in rats: Effect of pinealectomy. J. Reprod. Fertil. 2000, 119, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Tan, D.-X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and Reproduction Revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Hansen, P.J. Effects of heat stress on mammalian reproduction. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3341–3350. [Google Scholar] [CrossRef] [PubMed]
- Forcada, F.; Zarazaga, L.; Abecia, J.A. Effect of exogenous melatonin and plane of nutrition after weaning on estrous activity, endocrine status and ovulation rate in Salz ewes lambing in the seasonal anestrus. Theriogenology 1995, 43, 1179–1193. [Google Scholar] [CrossRef]
- Zarazaga, L.A.; Gatica, M.C.; Celi, I.; Guzmán, J.L.; Malpaux, B. Effect of melatonin implants on sexual activity in Mediterranean goat females without separation from males. Theriogenology 2009, 72, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Abecia, J.A.; Forcada, F.; Zúñiga, O. The effect of melatonin on the secretion of progesterone in sheep and on the development of ovine embryos in vitro. Vet. Res. Commun. 2002, 26, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Sugino, N.; Takiguchi, S.; Kashida, S.; Karube, A.; Nakamura, Y.; Kato, H. Superoxide dismutase expression in the human corpus luteum during the menstrual cycle and in early pregnancy. Mol. Hum. Reprod. 2000, 6, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manca, M.E.; Manunta, M.L.; Spezzigu, A.; Torres-rovira, L.; Gonzalez-bulnes, A.; Pasciu, V.; Piu, P.; Leoni, G.G.; Succu, S.; Chesneau, D.; et al. Melatonin deprival modifies follicular and corpus luteal growth dynamics in a sheep model. Reproduction 2014, 147, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Torres-Farfan, C.; Valenzuela, F.J.; Mondaca, M.; Valenzuela, G.J.; Krause, B.; Herrera, E.A.; Riquelme, R.; Llanos, A.J.; Seron-Ferre, M. Evidence of a role for melatonin in fetal sheep physiology: Direct actions of melatonin on fetal cerebral artery, brown adipose tissue and adrenal gland. J. Physiol. 2008, 586, 4017–4027. [Google Scholar] [CrossRef]
- Ealy, A.D.; Drost, M.; Hansen, P.J. Developmental Changes in Embryonic Resistance to Adverse Effects of Maternal Heat Stress in Cows. J. Dairy Sci. 1993, 76, 2899–2905. [Google Scholar] [CrossRef]
- Dutt, R.H. Detrimental effects of high ambient temperature on fertility and early embryo survival in sheep. Int. J. Biometeorol. 1964, 8, 47–56. [Google Scholar] [CrossRef]
- Abecia, J.A.; Forcada, F.; Vázquez, M.I.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Pérez-Pe, R.; Casao, A. Role of melatonin on embryo viability in sheep. Reprod. Fertil. Dev. 2019, 31, 82–92. [Google Scholar] [CrossRef]
- Lin, T.; Lee, J.E.; Kang, J.W.; Kim, S.Y.; Jin, D.I. The influence and role of melatonin on in vitro oocyte maturation and embryonic development in pig and cattle. Korean J. Agric. Sci. 2017, 44, 309–317. [Google Scholar]
- Pang, Y.; Zhao, S.; Sun, Y.; Jiang, X.; Hao, H.; Du, W.; Zhu, H. Protective effects of melatonin on the in vitro developmental competence of bovine oocytes. Anim. Sci. J. 2018, 89, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, F.; Zhang, L.; He, C.; Ji, P.; Wang, J.; Zhang, Z.; Lv, D.; Wusiman, A.; Wang, X.; et al. Beneficial effects of melatonin on the in vitro maturation of sheep oocytes and its relation to melatonin receptors. Int. J. Mol. Sci. 2017, 18, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.J.; Liu, W.J.; Wu, C.J.; Ma, F.H.; Ahmad, S.; Liu, B.R.; Han, L.; Jiang, X.P.; Zhang, S.J.; Yang, L.G. Melatonin suppresses apoptosis and stimulates proges-terone production by bovine granulosa cells via its receptors (MT1 and MT2). Theriogenology 2012, 78, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Talpur, H.S.; Chandio, I.B.; Brohi, R.D.; Worku, T.; Rehman, Z.; Bhattarai, D.; Ullah, F.; JiaJia, L.; Yang, L. Research progress on the role of melatonin and its receptors in animal reproduction: A comprehensive review. Reprod. Domest. Anim. 2018, 53, 831–849. [Google Scholar] [CrossRef]
- Matsuzuka, T.; Ozawa, M.; Nakamura, A.; Ushitani, A.; Hirabayashi, M.; Kanai, Y. Effects of heat stress on the redox status in the oviduct and early embryonic development in mice. J. Reprod. Dev. 2005, 51, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Aréchiga, C.F.; Ealy, A.D.; Hansen, P.J. Evidence That Glutathione is involved in Thermotolerance of Preimplantation Murine Embryos. Biol. Reprod. 1995, 52, 1296–1301. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef]
- Zhang, H.; Akbar, M.; Kim, H.Y. Melatonin: An endogenous negative modulator of 12-lipoxygenation in the rat pineal gland. Biochem. J. 1999, 344, 487–493. [Google Scholar] [CrossRef]
- Storr, M.; Koppitz, P.; Sibaev, A.; Saur, D.; Kurjak, M.; Franck, H.; Schusdziarra, V.; Allescher, H.-D. Melatonin reduces non-adrenergic, non-cholinergic relaxant neurotransmission by inhibition of nitric oxide synthase activity in the gastrointestinal tract of rodents in vitro. J. Pineal Res. 2002, 33, 101–108. [Google Scholar] [CrossRef]
- El-Sokkary, G.H. Melatonin and vitamin C administration ameliorate diazepam-induced oxidative stress and cell proliferation in the liver of rats. Cell Prolif. 2008, 41, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Guerrero, J.M.; Escames, G.; Pappolla, M.A.; Acuña-Castroviejo, D. Prophylactic actions of melatonin in oxidative neurotoxicity. Ann. N. Y. Acad. Sci. 1997, 825, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R.; Poindron, P. From birth to colostrum: Early steps leading to lamb survival. Reprod. Nutr. Dev. 2006, 46, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Lérias, J.R.; Hernández-Castellano, L.E.; Suárez-Trujillo, A.; Castro, N.; Pourlis, A.; Almeida, A.M. The mammary gland in small ruminants: Major morphological and functional events underlying milk production—A review. J. Dairy Res. 2014, 81, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Parisi, C.; Guerriero, G. Antioxidative Defense and Fertility Rate in the Assessment of Reprotoxicity Risk Posed by Global Warming. Antioxidants 2019, 8, 622. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Time of Day | Stage of the Heat Stress Period | |||||
|---|---|---|---|---|---|---|---|
| S1 | S2 | S3 | S4 | S5 | S6 | ||
| Rectal Temperature (°C) | |||||||
| M | 12.00 | 39.12 ± 0.02 a,m | 39.25 ± 0.02 a,m | 39.21 ± 0.02 a,m | 39.28 ± 0.01 a,m | 39.27 ± 0.01 a,m | 39.17 ± 0.02 a,m |
| 18.00 | 39.85 ± 0.03 a,n | 40.18 ± 0.02 a | 40.10 ± 0.02 a | 40.18 ± 0.02 a,n | 40.24 ± 0.02 a | 39.84 ± 0.03 a | |
| C | 12.00 | 39.30 ± 0.02 b,m | 39.33 ± 0.02 b,m | 39.30 ± 0.02 b,m | 39.32 ± 0.01 b,m | 39.35 ± 0.01 b,m | 39.27 ± 0.01 b,m |
| 18.00 | 40.05 ± 0.03b,n | 40.16 ± 0.02 b | 40.13 ± 0.02 b | 40.24 ± 0.02 b,n | 40.25 ± 0.03 b | 39.89 ± 0.03 b | |
| Breathing Rate (Breaths min−1) | |||||||
| M | 12.00 | 51.3 ± 1.1 c,o | 62.0 ± 0.7 c,o | 63.7 ± 0.9 c | 67.5 ± 0.6 c,o | 66.4 ± 0.6 c,o | 47.0 ± 1.1 c,o |
| 18.00 | 69.8 ± 1.2 c,p | 87.3 ± 0.7 c,p | 87.0 ± 1.0 c,p | 91.0 ± 0.6 c,p | 90.8 ± 0.6 c,p | 61.6 ± 1.6 c,p | |
| C | 12.00 | 57.0 ± 1.0 d,o | 66.3 ± 0.3 d,o | 61.8 ± 0.8 d | 63.4 ± 0.5 d,o | 63.8 ± 0.4 d,o | 51.8 ± 0.9 d,o |
| 18.00 | 88.9 ± 1.1 d,p | 104.0 ± 0.8 d,p | 112.9 ± 1.4 d,p | 115.7 ± 1.1 d,p | 115.7 ± 1.3 d,p | 69.2 ± 1.8 d,p | |
| Thermal Index | |||||||
| 12.00 | 27.9 ± 0.36 e | 29.7 ± 0.36 e | 28.1 ± 0.6 e | 28.5 ± 0.5 e | 27.3 ± 0.4 e | 25.9 ± 0.4 e | |
| 18.00 | 28.3 ± 0.38 e | 30.4 ± 0.57 e | 29.1 ± 0.5 e | 29.2 ± 0.4 e | 28.3 ± 0.4 e | 26.1 ± 0.4 e | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouroutzika, E.; Kouretas, D.; Papadopoulos, S.; Veskoukis, A.S.; Theodosiadou, E.; Makri, S.; Paliouras, C.; Michailidis, M.-L.; Caroprese, M.; Valasi, I. Effects of Melatonin Administration to Pregnant Ewes under Heat-Stress Conditions, in Redox Status and Reproductive Outcome. Antioxidants 2020, 9, 266. https://doi.org/10.3390/antiox9030266
Bouroutzika E, Kouretas D, Papadopoulos S, Veskoukis AS, Theodosiadou E, Makri S, Paliouras C, Michailidis M-L, Caroprese M, Valasi I. Effects of Melatonin Administration to Pregnant Ewes under Heat-Stress Conditions, in Redox Status and Reproductive Outcome. Antioxidants. 2020; 9(3):266. https://doi.org/10.3390/antiox9030266
Chicago/Turabian StyleBouroutzika, Efterpi, Dimitrios Kouretas, Serafeim Papadopoulos, Aristidis S. Veskoukis, Ekaterini Theodosiadou, Sotiria Makri, Charilaos Paliouras, Marios-Lazaros Michailidis, Mariangela Caroprese, and Irene Valasi. 2020. "Effects of Melatonin Administration to Pregnant Ewes under Heat-Stress Conditions, in Redox Status and Reproductive Outcome" Antioxidants 9, no. 3: 266. https://doi.org/10.3390/antiox9030266
APA StyleBouroutzika, E., Kouretas, D., Papadopoulos, S., Veskoukis, A. S., Theodosiadou, E., Makri, S., Paliouras, C., Michailidis, M. -L., Caroprese, M., & Valasi, I. (2020). Effects of Melatonin Administration to Pregnant Ewes under Heat-Stress Conditions, in Redox Status and Reproductive Outcome. Antioxidants, 9(3), 266. https://doi.org/10.3390/antiox9030266