Terpenoids as Potential Geroprotectors

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Extraction and Analysis of Terpenoids
3. Terpenoids as Potential Geroprotectors
3.1. Monoterpenes
3.1.1. Natural Sources
3.1.2. Lifespan Extension on Different Models
3.1.3. Effects on Stress-Resistance
3.1.4. Toxicity and Side Effects
3.1.5. Life Quality Effects
3.1.6. Suppression of Pro-Aging or Activation of Anti-Aging Molecular Targets or Pathways
3.1.7. Effects on Age-Related Diseases
3.1.8. Additional Activities
3.2. Sesquiterpenes
3.2.1. Natural Sources
3.2.2. Lifespan Extension on Different Models
3.2.3. Effects on Stress-Resistance
3.2.4. Effects on Aging Biomarkers
3.2.5. Toxicity and Side Effects
3.2.6. Life Quality Effects
3.2.7. Suppression of Pro-Aging or Activation of Anti-Aging Molecular Targets or Pathways
3.2.8. Effects on Age-Related Diseases
3.2.9. Additional Activities
3.3. Diterpenes
3.3.1. Natural Sources
3.3.2. Lifespan Extension on Different Models
3.3.3. Effects on Stress-Resistance
3.3.4. Effects on Aging Biomarkers
3.3.5. Toxicity and Side Effects
3.3.6. Life Quality Effects
3.3.7. Suppression of Pro-Aging or Activation of Anti-Aging Molecular Targets or Pathways
3.3.8. Effects on Age-Related Diseases
3.3.9. Additional Activities
3.4. Triterpenes
3.4.1. Natural Sources
3.4.2. Lifespan Extension on Different Models
3.4.3. Effects on Stress-Resistance
3.4.4. Effects on Aging Biomarkers
3.4.5. Toxicity and Side Effects
3.4.6. Life Quality Effects
3.4.7. Suppression of Pro-Aging or Activation of Anti-Aging Molecular Targets or Pathways
3.4.8. Effects on Age-Related Diseases
3.4.9. Additional Activities
3.5. Tetraterpenes or Carotenoids
3.5.1. Natural Sources
3.5.2. Lifespan Extension on Different Models
3.5.3. Effects on Stress-Resistance
3.5.4. Effects on Aging Biomarkers
3.5.5. Toxicity and Side Effects
3.5.6. Life Quality Effects
3.5.7. Suppression of Pro-Aging or Activation of Anti-Aging Molecular Targets or Pathways
3.5.8. Effects on Age-Related Diseases
3.6. Polyterpenes
3.7. Norisoprenoids
3.8. Sesterterpenes
3.9. Sesquarterpenes
4. Possible Prospect of Terpene using as Anti-Aging Drugs
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yazaki, K.; Arimura, G.I.; Ohnishi, T. ‘Hidden’ Terpenoids in Plants: Their Biosynthesis, Localization and Ecological Roles. Plant Cell Physiol. 2017, 58, 1615–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Smith, M.; Khatoon, T. Biological Activity of the Terpenoids and Their Derivatives. In Progress in Drug Research/Fortschritte der Arzneimittelforschung/Progrès des recherches pharmaceutiques; Gillette, J.R., Heinzelman, R.V., Szmuszkovicz, J., Leemann, H.G., Stich, K., Thomas, M., Martin-Smith, M., Khatoon, T., Kunz, W., Jucker, E., Eds.; Birkhäuser Basel: Basel, Switzerland, 1963; pp. 279–346. [Google Scholar] [CrossRef]
- Brahmkshatriya, P.P.; Brahmkshatriya, P.S. Terpenes: Chemistry, Biological Role, and Therapeutic Applications. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2665–2691. [Google Scholar] [CrossRef]
- Jaeger, R.; Cuny, E. Terpenoids with Special Pharmacological Significance: A Review. Nat. Prod. Commun. 2016, 11, 1934578X1601100946. [Google Scholar] [CrossRef] [Green Version]
- Kudryavtseva, A.; Krasnov, G.; Lipatova, A.; Alekseev, B.; Maganova, F.; Shaposhnikov, M.; Fedorova, M.; Snezhkina, A.; Moskalev, A. Effects of Abies sibirica terpenes on cancer- and aging-associated pathways in human cells. Oncotarget 2016, 7, 83744–83754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskalev, A.; Chernyagina, E.; Tsvetkov, V.; Fedintsev, A.; Shaposhnikov, M.; Krut’ko, V.; Zhavoronkov, A.; Kennedy, B.K. Developing criteria for evaluation of geroprotectors as a key stage toward translation to the clinic. Aging Cell 2016, 15, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Kempinski, C.; Chappell, J. Extraction and Analysis of Terpenes/Terpenoids. Curr. Protoc. Plant Biol. 2016, 1, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Piñeiro, Z.; Palma, M.; Barroso, C.G. Determination of terpenoids in wines by solid phase extraction and gas chromatography. Anal. Chim. Acta 2004, 513, 209–214. [Google Scholar] [CrossRef]
- Harman-Ware, A.E.; Sykes, R.; Peter, G.F.; Davis, M. Determination of Terpenoid Content in Pine by Organic Solvent Extraction and Fast-GC Analysis. Front. Energy Res. 2016, 4. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Riordan-Short, S.; Dang, T.T.; O’Brien, R.; Noestheden, M. Quantitation of Select Terpenes/Terpenoids and Nicotine Using Gas Chromatography-Mass Spectrometry with High-Temperature Headspace Sampling. ACS Omega 2020, 5, 5565–5573. [Google Scholar] [CrossRef]
- Azmir, J.; Zaidul, I.S.M.; Rahman, M.M.; Sharif, K.M.; Mohamed, A.; Sahena, F.; Jahurul, M.H.A.; Ghafoor, K.; Norulaini, N.A.N.; Omar, A.K.M. Techniques for extraction of bioactive compounds from plant materials: A review. J. Food Eng. 2013, 117, 426–436. [Google Scholar] [CrossRef]
- Marriott, P.J.; Shellie, R.; Cornwell, C. Gas chromatographic technologies for the analysis of essential oils. J. Chromatogr. 2001, 936, 1–22. [Google Scholar] [CrossRef]
- Pawliszyn, J. 1–Solid-Phase Microextraction in Perspective. In Handbook of Solid Phase Microextraction; Pawliszyn, J., Ed.; Elsevier: Oxford, AZ, USA, 2012; pp. 1–12. [Google Scholar] [CrossRef]
- Bajer, T.; Ligor, M.; Ligor, T.; Buszewski, B. Design of the extraction process for terpenes and other volatiles from allspice by solid-phase microextraction and hydrodistillation. J. Sep. Sci. 2016, 39, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Chemat, F.; Abert-Vian, M.; Fernandez, X. Microwave-Assisted Extraction of Essential Oils and Aromas. In Microwave-Assisted Extraction for Bioactive Compounds: Theory and Practice; Chemat, F., Cravotto, G., Eds.; Springer: Boston, MA, USA, 2013; pp. 53–68. [Google Scholar] [CrossRef]
- Martins, M.A.R.; Domańska, U.; Schröder, B.; Coutinho, J.A.P.; Pinho, S.P. Selection of Ionic Liquids to be Used as Separation Agents for Terpenes and Terpenoids. ACS Sustain. Chem. Eng. 2015, 4, 548–556. [Google Scholar] [CrossRef]
- Sun, Y.; Li, W.; Fitzloff, J.F.; van Breemen, R.B. Liquid chromatography/electrospray tandem mass spectrometry of terpenoid lactones in Ginkgo biloba. J. Mass Spectrom. 2005, 40, 373–379. [Google Scholar] [CrossRef]
- Tran, T.A.; Ho, M.T.; Song, Y.W.; Cho, M.; Cho, S.K. Camphor Induces Proliferative and Anti-senescence Activities in Human Primary Dermal Fibroblasts and Inhibits UV-Induced Wrinkle Formation in Mouse Skin. Phytother. Res. 2015, 29, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, T.; Kafil, H.S.; Asnaashari, S.; Farajnia, S.; Delazar, A.; Baek, S.C.; Hamishehkar, H.; Kim, K.H. Chemical Composition and Antimicrobial Activity of Essential Oils from the Aerial Parts of Pinus eldarica Grown in Northwestern Iran. Molecules 2019, 24, 3203. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.Y.; Chuang, C.H.; Chen, H.C.; Yang, K.M. Lime (Citrus aurantifolia (Christm.) Swingle) Essential Oils: Volatile Compounds, Antioxidant Capacity, and Hypolipidemic Effect. Foods 2019, 8, 398. [Google Scholar] [CrossRef] [Green Version]
- Md Othman, S.N.A.; Hassan, M.A.; Nahar, L.; Basar, N.; Jamil, S.; Sarker, S.D. Essential Oils from the Malaysian Citrus (Rutaceae) Medicinal Plants. Medicines 2016, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Papanastasiou, S.A.; Bali, E.-M.D.; Ioannou, C.S.; Papachristos, D.P.; Zarpas, K.D.; Papadopoulos, N.T. Toxic and hormetic-like effects of three components of citrus essential oils on adult Mediterranean fruit flies (Ceratitis capitata). PLoS ONE 2017, 12, e0177837. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Bedmar, Z.; Anter, J.; de La Cruz-Ares, S.; Muñoz-Serrano, A.; Alonso-Moraga, A.; Pérez-Guisado, J. Role of citrus juices and distinctive components in the modulation of degenerative processes: Genotoxicity, antigenotoxicity, cytotoxicity, and longevity in Drosophila. J. Toxicol. Environ. Health Part A 2011, 74, 1052–1066. [Google Scholar] [CrossRef]
- Gerofotis, C.D.; Ioannou, C.S.; Nakas, C.T.; Papadopoulos, N.T. The odor of a plant metabolite affects life history traits in dietary restricted adult olive flies. Sci. Rep. 2016, 6, 28540. [Google Scholar] [CrossRef]
- Mattson, M.P.; Son, T.G.; Camandola, S. Viewpoint: Mechanisms of action and therapeutic potential of neurohormetic phytochemicals. Dose Response 2007, 5, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Moayedi, Y.; Greenberg, S.A.; Jenkins, B.A.; Marshall, K.L.; Dimitrov, L.V.; Nelson, A.M.; Owens, D.M.; Lumpkin, E.A. Camphor white oil induces tumor regression through cytotoxic T cell-dependent mechanisms. Mol. Carcinog. 2019, 58, 722–734. [Google Scholar] [CrossRef]
- Nikolić, B.; Vasilijević, B.; Mitić-Ćulafić, D.; Vuković-Gačić, B.; Knežević-Vukćević, J. Comparative study of genotoxic, antigenotoxic and cytotoxic activities of monoterpenes camphor, eucalyptol and thujone in bacteria and mammalian cells. Chem. Biol. Interact. 2015, 242, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.; Kanimozhi, G.; Madahavan, N.R.; Agilan, B.; Ganesan, M.; Prasad, N.R.; Rathinaraj, P. Alpha-pinene attenuates UVA-induced photoaging through inhibition of matrix metalloproteinases expression in mouse skin. Life Sci. 2019, 217, 110–118. [Google Scholar] [CrossRef]
- Wu, Z.; Tan, B.; Liu, Y.; Dunn, J.; Martorell Guerola, P.; Tortajada, M.; Cao, Z.; Ji, P. Chemical Composition and Antioxidant Properties of Essential Oils from Peppermint, Native Spearmint and Scotch Spearmint. Molecules 2019, 24, 2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foti, M.C.; Ingold, K.U. Mechanism of inhibition of lipid peroxidation by gamma-terpinene, an unusual and potentially useful hydrocarbon antioxidant. J. Agric. Food Chem. 2003, 51, 2758–2765. [Google Scholar] [CrossRef] [Green Version]
- Pratheeshkumar, P.; Raphael, T.J.; Kuttan, G. Protective role of perillic acid against radiation-induced oxidative stress, cytokine profile, DNA damage, and intestinal toxicity in mice. J. Environ. Pathol. Toxicol. Oncol. 2010, 29, 199–212. [Google Scholar] [CrossRef]
- Singh, B.K.; Tripathi, M.; Chaudhari, B.P.; Pandey, P.K.; Kakkar, P. Natural terpenes prevent mitochondrial dysfunction, oxidative stress and release of apoptotic proteins during nimesulide-hepatotoxicity in rats. PLoS ONE 2012, 7, e34200. [Google Scholar] [CrossRef] [Green Version]
- Mohamad, R.H.; El-Bastawesy, A.M.; Abdel-Monem, M.G.; Noor, A.M.; Al-Mehdar, H.A.; Sharawy, S.M.; El-Merzabani, M.M. Antioxidant and anticarcinogenic effects of methanolic extract and volatile oil of fennel seeds (Foeniculum vulgare). J. Med. Food 2011, 14, 986–1001. [Google Scholar] [CrossRef]
- Gomes-Carneiro, M.R.; Felzenszwalb, I.; Paumgartten, F.J. Mutagenicity testing (+/-)-camphor, 1,8-cineole, citral, citronellal, (-)-menthol and terpineol with the Salmonella/microsome assay. Mutat. Res. 1998, 416, 129–136. [Google Scholar] [CrossRef] [Green Version]
- De Groot, A. Limonene Hydroperoxides. Dermatitis 2019, 30, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Kopečná, M.; Macháček, M.; Nováčková, A.; Paraskevopoulos, G.; Roh, J.; Vávrová, K. Esters of terpene alcohols as highly potent, reversible, and low toxic skin penetration enhancers. Sci. Rep. 2019, 9, 14617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisson, J.F.; Menut, C.; d’Alessio, P. Anti-inflammatory senescence actives 5203-L molecule to promote healthy aging and prolongation of lifespan. Rejuvenation Res. 2008, 11, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alessio, P.A.; Bisson, J.F.; Bene, M.C. Anti-stress effects of d-limonene and its metabolite perillyl alcohol. Rejuvenation Res. 2014, 17, 145–149. [Google Scholar] [CrossRef]
- Ostan, R.; Bene, M.C.; Spazzafumo, L.; Pinto, A.; Donini, L.M.; Pryen, F.; Charrouf, Z.; Valentini, L.; Lochs, H.; Bourdel-Marchasson, I.; et al. Impact of diet and nutraceutical supplementation on inflammation in elderly people. Results from the RISTOMED study, an open-label randomized control trial. Clin. Nutr. 2016, 35, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, G.A.d.; Souza, D.S.d.; Lima, B.S.; Vasconcelos, C.M.L.d.; Araújo, A.A.d.S.; Durço, A.O.; Quintans-Junior, L.J.; Almeida, J.R.G.d.S.; Oliveira, A.P.; Santana-Filho, V.J.d.; et al. Bradycardic and Antiarrhythmic Effects of the D-Limonene in Rats. Arq. Bras. De Cardiol. 2019, 113, 925–932. [Google Scholar] [CrossRef]
- Kesbiç, O.S.; Acar, Ü.; Yilmaz, S.; Aydin, Ö.D. Effects of bergamot (Citrus bergamia) peel oil-supplemented diets on growth performance, haematology and serum biochemical parameters of Nile tilapia (Oreochromis niloticus). Fish. Physiol. Biochem. 2020, 46, 103–110. [Google Scholar] [CrossRef]
- Hwang, E.; Ngo, H.T.T.; Park, B.; Seo, S.A.; Yang, J.E.; Yi, T.H. Myrcene, an Aromatic Volatile Compound, Ameliorates Human Skin Extrinsic Aging via Regulation of MMPs Production. Am. J. Chin. Med. 2017, 45, 1113–1124. [Google Scholar] [CrossRef]
- Nevzorova, Y.A.; Grossmann, J.; Trautwein, C. Anti-tumorigenic and anti-angiogenic effects of natural conifer Abies sibirica terpenoids in vivo and in vitro. Biomed. Pharm. 2017, 89, 386–395. [Google Scholar] [CrossRef]
- Srinivasan, S.; Muruganathan, U. Antidiabetic efficacy of citronellol, a citrus monoterpene by ameliorating the hepatic key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2016, 250, 38–46. [Google Scholar] [CrossRef]
- Raphael, T.J.; Kuttan, G. Effect of naturally occurring monoterpenes carvone, limonene and perillic acid in the inhibition of experimental lung metastasis induced by B16F-10 melanoma cells. J. Exp. Clin. Cancer Res. 2003, 22, 419–424. [Google Scholar] [PubMed]
- Raphael, T.J.; Kuttan, G. Immunomodulatory activity of naturally occurring monoterpenes carvone, limonene, and perillic acid. Immunopharmacol. Immunotoxicol. 2003, 25, 285–294. [Google Scholar] [CrossRef] [PubMed]
- El-Minshawy, A.M.; Abdelgaleil, S.A.M.; Gadelhak, G.G.; Al-Eryan, M.A.; Rabab, R.A. Effects of monoterpenes on mortality, growth, fecundity, and ovarian development of Bactrocera zonata (Saunders) (Diptera: Tephritidae). Environ. Sci. Pollut. Res. Int. 2018, 25, 15671–15679. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.H.; Salazar, D.; Hakim, I.A. Pharmacokinetics of perillic acid in humans after a single dose administration of a citrus preparation rich in d-limonene content. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1472–1476. [Google Scholar]
- Rufino, A.T.; Ribeiro, M.; Sousa, C.; Judas, F.; Salgueiro, L.; Cavaleiro, C.; Mendes, A.F. Evaluation of the anti-inflammatory, anti-catabolic and pro-anabolic effects of E-caryophyllene, myrcene and limonene in a cell model of osteoarthritis. Eur. J. Pharm. 2015, 750, 141–150. [Google Scholar] [CrossRef]
- Hassan, S.B.; Gali-Muhtasib, H.; Goransson, H.; Larsson, R. Alpha terpineol: A potential anticancer agent which acts through suppressing NF-kappaB signalling. Anticancer Res. 2010, 30, 1911–1919. [Google Scholar]
- Moghimi, M.; Parvardeh, S.; Zanjani, T.M.; Ghafghazi, S. Protective effect of alpha-terpineol against impairment of hippocampal synaptic plasticity and spatial memory following transient cerebral ischemia in rats. Iran J. Basic Med. Sci 2016, 19, 960–969. [Google Scholar]
- Wu, Z.L.; Yin, Z.Q.; Du, Y.H.; Feng, R.Z.; Ye, K.C.; Wei, Q.; Hu, Y.; He, L.; Liao, L.; Wang, Y. gamma-terpineol inhibits cell growth and induces apoptosis in human liver cancer BEL-7402 cells in vitro. Int. J. Clin. Exp. Pathol. 2014, 7, 6524–6533. [Google Scholar]
- Bardon, S.; Picard, K.; Martel, P. Monoterpenes inhibit cell growth, cell cycle progression, and cyclin D1 gene expression in human breast cancer cell lines. Nutr. Cancer 1998, 32, 1–7. [Google Scholar] [CrossRef]
- Vallianou, I.; Peroulis, N.; Pantazis, P.; Hadzopoulou-Cladaras, M. Camphene, a plant-derived monoterpene, reduces plasma cholesterol and triglycerides in hyperlipidemic rats independently of HMG-CoA reductase activity. PLoS ONE 2011, 6, e20516. [Google Scholar] [CrossRef] [Green Version]
- Pereira Fde, O.; Mendes, J.M.; Lima, I.O.; Mota, K.S.; Oliveira, W.A.; Lima Ede, O. Antifungal activity of geraniol and citronellol, two monoterpenes alcohols, against Trichophyton rubrum involves inhibition of ergosterol biosynthesis. Pharm. Biol. 2015, 53, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves-Silva, J.M.; Zuzarte, M.; Goncalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Salgueiro, L. Unveiling the bioactive potential of the essential oil of a Portuguese endemism, St. Impressa. J. Ethnopharmacol. 2019, 244, 112120. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, T.; Singh, S.; Nishad, I.; Kumar, A.; Tiwari, N.; Tandon, S.; Saikia, D.; Verma, R.S. Chemical composition and antimicrobial activity of the essential oil of senescent leaves of guava (Psidium guajava L.). Nat. Prod. Res. 2019, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Pant, A.; Mishra, V.; Saikia, S.K.; Shukla, V.; Asthana, J.; Akhoon, B.A.; Pandey, R. Beta-caryophyllene modulates expression of stress response genes and mediates longevity in Caenorhabditis elegans. Exp. Gerontol. 2014, 57, 81–95. [Google Scholar] [CrossRef]
- Santhanasabapathy, R.; Vasudevan, S.; Anupriya, K.; Pabitha, R.; Sudhandiran, G. Farnesol quells oxidative stress, reactive gliosis and inflammation during acrylamide-induced neurotoxicity: Behavioral and biochemical evidence. Neuroscience 2015, 308, 212–227. [Google Scholar] [CrossRef]
- Kaur, D.; Pahwa, P.; Goel, R.K. Protective Effect of Nerolidol Against Pentylenetetrazol-Induced Kindling, Oxidative Stress and Associated Behavioral Comorbidities in Mice. Neurochem. Res. 2016, 41, 2859–2867. [Google Scholar] [CrossRef]
- Javed, H.; Azimullah, S.; Abul Khair, S.B.; Ojha, S.; Haque, M.E. Neuroprotective effect of nerolidol against neuroinflammation and oxidative stress induced by rotenone. BMC Neurosci. 2016, 17, 58. [Google Scholar] [CrossRef] [Green Version]
- Leite, G.O.; Ecker, A.; Seeger, R.L.; Krum, B.N.; Lugokenski, T.H.; Fachinetto, R.; Sudati, J.H.; Barbosa, N.V.; Wagner, C. Protective effect of (-)-alpha-bisabolol on rotenone-induced toxicity in Drosophila melanogaster. Can. J. Physiol. Pharm. 2018, 96, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Meeran, M.F.N.; Laham, F.; Al-Taee, H.; Azimullah, S.; Ojha, S. Protective effects of alpha-bisabolol on altered hemodynamics, lipid peroxidation, and nonenzymatic antioxidants in isoproterenol-induced myocardial infarction: In vivo and in vitro evidences. J. Biochem. Mol. Toxicol. 2018, 32, e22200. [Google Scholar] [CrossRef]
- Sampaio, T.L.; Menezes, R.R.; da Costa, M.F.; Meneses, G.C.; Arrieta, M.C.; Chaves Filho, A.J.; de Morais, G.B.; Liborio, A.B.; Alves, R.S.; Evangelista, J.S.; et al. Nephroprotective effects of (-)-alpha-bisabolol against ischemic-reperfusion acute kidney injury. Phytomedicine 2016, 23, 1843–1852. [Google Scholar] [CrossRef]
- Meng, X.; Li, N.; Zhang, Y.; Fan, D.; Yang, C.; Li, H.; Guo, D.; Pan, S. Beneficial Effect of beta-Elemene Alone and in Combination with Hyperbaric Oxygen in Traumatic Brain Injury by Inflammatory Pathway. Transl. Neurosci. 2018, 9, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Han, J.S. The antioxidant xanthorrhizol prevents amyloid-beta-induced oxidative modification and inactivation of neprilysin. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Calleja, M.A.; Vieites, J.M.; Montero-Melendez, T.; Torres, M.I.; Faus, M.J.; Gil, A.; Suarez, A. The antioxidant effect of beta-caryophyllene protects rat liver from carbon tetrachloride-induced fibrosis by inhibiting hepatic stellate cell activation. Br. J. Nutr. 2013, 109, 394–401. [Google Scholar] [CrossRef]
- Lnenickova, K.; Svobodova, H.; Skalova, L.; Ambroz, M.; Novak, F.; Matouskova, P. The impact of sesquiterpenes beta-caryophyllene oxide and trans-nerolidol on xenobiotic-metabolizing enzymes in mice in vivo. Xenobiotica 2018, 48, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Yi, M.; Wang, R.; Huang, Y.; Chen, M. Protective Effects of Costunolide Against D-Galactosamine and Lipopolysaccharide-Induced Acute Liver Injury in Mice. Front. Pharm. 2018, 9, 1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliza, J.; Daisy, P.; Ignacimuthu, S. Antioxidant activity of costunolide and eremanthin isolated from Costus speciosus (Koen ex. Retz) Sm. Chem. Biol. Interact. 2010, 188, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Cheong, C.U.; Yeh, C.S.; Hsieh, Y.W.; Lee, Y.R.; Lin, M.Y.; Chen, C.Y.; Lee, C.H. Protective Effects of Costunolide against Hydrogen Peroxide-Induced Injury in PC12 Cells. Molecules 2016, 21, 898. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Chen, Y.; Zhang, J.; Wang, L.; Jin, Z.; Huang, H.; Man, S.; Gao, W. Evaluation of protective effects of costunolide and dehydrocostuslactone on ethanol-induced gastric ulcer in mice based on multi-pathway regulation. Chem. Biol. Interact. 2016, 250, 68–77. [Google Scholar] [CrossRef]
- Jang, Y.J.; Back, M.J.; Fu, Z.; Lee, J.H.; Won, J.H.; Ha, H.C.; Lee, H.K.; Jang, J.M.; Choi, J.M.; Kim, D.K. Protective effect of sesquiterpene lactone parthenolide on LPS-induced acute lung injury. Arch. Pharm. Res. 2016, 39, 1716–1725. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, S.; Huang, R.; Wei, L.; Tan, S.; Liang, S.; Tian, Y.; Wu, X.; Lu, Z.; Huang, Q. Helenalin attenuates alcohol-induced hepatic fibrosis by enhancing ethanol metabolism, inhibiting oxidative stress and suppressing HSC activation. Fitoterapia 2014, 95, 203–213. [Google Scholar] [CrossRef]
- Wang, X.; Lan, Y.L.; Xing, J.S.; Lan, X.Q.; Wang, L.T.; Zhang, B. Alantolactone plays neuroprotective roles in traumatic brain injury in rats via anti-inflammatory, anti-oxidative and anti-apoptosis pathways. Am. J. Transl. Res. 2018, 10, 368–380. [Google Scholar] [PubMed]
- Seo, J.Y.; Lim, S.S.; Kim, J.; Lee, K.W.; Kim, J.S. Alantolactone and Isoalantolactone Prevent Amyloid β25-35-induced Toxicity in Mouse Cortical Neurons and Scopolamine-induced Cognitive Impairment in Mice. Phytother. Res. 2017, 31, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, Z.; Gao, J.; Xie, J.; Yang, L.; Hu, S. Downregulation effects of beta-elemene on the levels of plasma endotoxin, serum TNF-alpha, and hepatic CD14 expression in rats with liver fibrosis. Front. Med. 2011, 5, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Oon, S.F.; Nallappan, M.; Kassim, N.K.; Shohaimi, S.; Sa’ariwijaya, M.S.; Tee, T.T.; Cheah, Y.H. Hypolipidemic activities of xanthorrhizol purified from centrifugal TLC. Biochem. Biophys. Res. Commun. 2016, 478, 1403–1408. [Google Scholar] [CrossRef] [Green Version]
- Harb, A.A.; Bustanji, Y.K.; Abdalla, S.S. Hypocholesterolemic effect of beta-caryophyllene in rats fed cholesterol and fat enriched diet. J. Clin. Biochem. Nutr. 2018, 62, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Baldissera, M.D.; Souza, C.F.; Grando, T.H.; Stefani, L.M.; Monteiro, S.G. β-caryophyllene reduces atherogenic index and coronary risk index in hypercholesterolemic rats: The involvement of cardiac oxidative damage. Chem. Biol. Interact. 2017, 270, 9–14. [Google Scholar] [CrossRef]
- Tian, X.; Peng, J.; Zhong, J.; Yang, M.; Pang, J.; Lou, J.; Li, M.; An, R.; Zhang, Q.; Xu, L.; et al. β-Caryophyllene protects in vitro neurovascular unit against oxygen-glucose deprivation and re-oxygenation-induced injury. J. Neurochem. 2016, 139, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.M.; Palmeira, C.M.; Oliveira, M.M.; Santos, D.; Simoes, A.M.; Rocha, S.M.; Coimbra, M.A.; Peixoto, F. Nerolidol effects on mitochondrial and cellular energetics. Toxicol. In Vitro 2012, 26, 189–196. [Google Scholar] [CrossRef]
- Piculo, F.; Guiraldeli Macedo, C.; de Andrade, S.F.; Luis Maistro, E. In vivo genotoxicity assessment of nerolidol. J. Appl. Toxicol. 2011, 31, 633–639. [Google Scholar] [CrossRef]
- Rigo, A.; Vinante, F. The antineoplastic agent alpha-bisabolol promotes cell death by inducing pores in mitochondria and lysosomes. Apoptosis 2016, 21, 917–927. [Google Scholar] [CrossRef]
- Oliveira, G.; Machado, K.C.; Machado, K.C.; da Silva, A.; Feitosa, C.M.; de Castro Almeida, F.R. Non-clinical toxicity of beta-caryophyllene, a dietary cannabinoid: Absence of adverse effects in female Swiss mice. Regul. Toxicol. Pharm. 2018, 92, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, D.; Levy, R.; Carroll, B. Toxicological Evaluation of beta-Caryophyllene Oil: Subchronic Toxicity in Rats. Int. J. Toxicol. 2016, 35, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Singireesu, S.; Misra, S.; Mondal, S.K.; Yerramsetty, S.; Sahu, N.; K, S.B. Costunolide induces micronuclei formation, chromosomal aberrations, cytostasis, and mitochondrial-mediated apoptosis in Chinese hamster ovary cells. Cell Biol. Toxicol. 2018, 34, 125–142. [Google Scholar] [CrossRef]
- Berges, C.; Fuchs, D.; Opelz, G.; Daniel, V.; Naujokat, C. Helenalin suppresses essential immune functions of activated CD4+ T cells by multiple mechanisms. Mol. Immunol. 2009, 46, 2892–2901. [Google Scholar] [CrossRef]
- Supornsilchai, V.; Soder, O.; Svechnikov, K. Sesquiterpene lactone helenalin suppresses Leydig and adrenocortical cell steroidogenesis by inhibiting expression of the steroidogenic acute regulatory protein. Reprod. Toxicol. 2006, 22, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.M.; Lin, J.Y. Farnesol, a sesquiterpene alcohol in herbal plants, exerts anti-inflammatory and antiallergic effects on ovalbumin-sensitized and -challenged asthmatic mice. Evid. Based Complement. Altern. Med. 2015, 2015, 387357. [Google Scholar] [CrossRef] [Green Version]
- Ku, C.M.; Lin, J.Y. Farnesol, a sesquiterpene alcohol in essential oils, ameliorates serum allergic antibody titres and lipid profiles in ovalbumin-challenged mice. Allergol. Immunopathol. (Madr.) 2016, 44, 149–159. [Google Scholar] [CrossRef]
- De Oliveira Junior, W.M.; Benedito, R.B.; Pereira, W.B.; de Arruda Torres, P.; Ramos, C.A.; Costa, J.P.; da Rocha Tome, A.; de Sousa, D.P.; de Freitas, R.M.; de Fatima Formiga Melo Diniz, M.; et al. Farnesol: Antinociceptive effect and histopathological analysis of the striatum and hippocampus of mice. Fundam. Clin. Pharm. 2013, 27, 419–426. [Google Scholar] [CrossRef]
- Sharma, C.; Al Kaabi, J.M.; Nurulain, S.M.; Goyal, S.N.; Kamal, M.A.; Ojha, S. Polypharmacological Properties and Therapeutic Potential of beta-Caryophyllene: A Dietary Phytocannabinoid of Pharmaceutical Promise. Curr. Pharm. Des. 2016, 22, 3237–3264. [Google Scholar] [CrossRef]
- Joo, J.H.; Jetten, A.M. Molecular mechanisms involved in farnesol-induced apoptosis. Cancer Lett. 2010, 287, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Biazi, B.I.; Zanetti, T.A.; Baranoski, A.; Corveloni, A.C.; Mantovani, M.S. Cis-Nerolidol Induces Endoplasmic Reticulum Stress and Cell Death in Human Hepatocellular Carcinoma Cells through Extensive CYP2C19 and CYP1A2 Oxidation. Basic Clin. Pharm. Toxicol. 2017, 121, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Cho, S.K.; Kim, K.D.; Nam, D.; Chung, W.S.; Jang, H.J.; Lee, S.G.; Shim, B.S.; Sethi, G.; Ahn, K.S. β-Caryophyllene oxide potentiates TNFα-induced apoptosis and inhibits invasion through down-modulation of NF-κB-regulated gene products. Apoptosis 2014, 19, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, H.; Yao, Y.; Ren, Y.; Lin, J.; Hu, J.; Zheng, M.; Song, X.; Zhao, T.; Chen, Y.Y.; et al. beta-Elemene Enhances GAP-43 Expression and Neurite Outgrowth by Inhibiting RhoA Kinase Activation in Rats with Spinal Cord Injury. Neuroscience 2018, 383, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Hong, L.L.; Lin, J.S.; Yao, X.Y.; Wu, D.H.; Lin, X.P.; Zhang, J.M.; Zhang, X.B.; Zeng, Y.M. β-Elemene inhibits the proliferation of primary human airway granulation fibroblasts by down-regulating canonical Wnt/beta-catenin pathway. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Kang, Y.; Zou, H.; Cheng, X.; Xie, T.; Shi, L.; Zhang, H. β-Elemene attenuates macrophage activation and proinflammatory factor production via crosstalk with Wnt/β-catenin signaling pathway. Fitoterapia 2018, 124, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jiang, Z.Y.; Zhou, Y.L.; Qiu, H.H.; Wang, G.; Luo, Y.; Liu, J.B.; Liu, X.W.; Bu, W.Q.; Song, J.; et al. β-Elemene regulates endoplasmic reticulum stress to induce the apoptosis of NSCLC cells through PERK/IRE1α/ATF6 pathway. Biomed. Pharm. 2017, 93, 490–497. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Luo, H.; Luo, Z.; Zhang, T.; Yang, N.; Long, X.; Xie, H.; Qiu, W.; Zhang, B.; et al. β-Elemene acts as an antitumor factor and downregulates the expression of survivin, Bcl-xL and Mta-1. Mol. Med. Rep. 2012, 6, 989–995. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; An, J.M.; Chung, W.Y.; Park, K.K.; Hwang, J.K.; Kim du, S.; Seo, S.R.; Seo, J.T. Xanthorrhizol induces apoptosis through ROS-mediated MAPK activation in human oral squamous cell carcinoma cells and inhibits DMBA-induced oral carcinogenesis in hamsters. Phytother. Res. 2013, 27, 493–498. [Google Scholar] [CrossRef]
- Kang, Y.J.; Park, K.K.; Chung, W.Y.; Hwang, J.K.; Lee, S.K. Xanthorrhizol, a natural sesquiterpenoid, induces apoptosis and growth arrest in HCT116 human colon cancer cells. J. Pharm. Sci. 2009, 111, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Handayani, T.; Sakinah, S.; Nallappan, M.; Pihie, A.H. Regulation of p53-, Bcl-2- and caspase-dependent signaling pathway in xanthorrhizol-induced apoptosis of HepG2 hepatoma cells. Anticancer Res. 2007, 27, 965–971. [Google Scholar]
- Lou, J.; Cao, G.; Li, R.; Liu, J.; Dong, Z.; Xu, L. β-Caryophyllene Attenuates Focal Cerebral Ischemia-Reperfusion Injury by Nrf2/HO-1 Pathway in Rats. Neurochem. Res. 2016, 41, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.I.; Hong, J.M.; Choi, J.W.; Choi, H.S.; Kwak, J.H.; Lee, D.U.; Kook Lee, S.; Lee, S.M. β-Caryophyllene alleviates D-galactosamine and lipopolysaccharide-induced hepatic injury through suppression of the TLR4 and RAGE signaling pathways. Eur. J. Pharm. 2015, 764, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Gonzalez, I.; Madrigal-Bujaidar, E.; Castro-Garcia, S. Antigenotoxic capacity of beta-caryophyllene in mouse, and evaluation of its antioxidant and GST induction activities. J. Toxicol. Sci. 2014, 39, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Dong, Z.; Liu, S. β-Caryophyllene ameliorates the Alzheimer-like phenotype in APP/PS1 Mice through CB2 receptor activation and the PPARγ pathway. Pharmacology 2014, 94, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, D.; Li, M.; Wang, B. Costunolide ameliorates lipoteichoic acid-induced acute lung injury via attenuating MAPK signaling pathway. Int. Immunopharmacol. 2018, 61, 283–289. [Google Scholar] [CrossRef]
- Hu, M.; Liu, L.; Yao, W. Activation of p53 by costunolide blocks glutaminolysis and inhibits proliferation in human colorectal cancer cells. Gene 2018, 678, 261–269. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Zhao, L.; Shi, M.; Wei, Z.; Yang, Z.; Guo, C.; Fu, Y. Costunolide protects lipopolysaccharide/d-galactosamine-induced acute liver injury in mice by inhibiting NF-kappaB signaling pathway. J. Surg. Res. 2017, 220, 40–45. [Google Scholar] [CrossRef]
- Rayan, N.A.; Baby, N.; Pitchai, D.; Indraswari, F.; Ling, E.A.; Lu, J.; Dheen, T. Costunolide inhibits proinflammatory cytokines and iNOS in activated murine BV2 microglia. Front. Biosci. (Elite Ed.) 2011, 3, 1079–1091. [Google Scholar] [CrossRef]
- Choi, S.H.; Im, E.; Kang, H.K.; Lee, J.H.; Kwak, H.S.; Bae, Y.T.; Park, H.J.; Kim, N.D. Inhibitory effects of costunolide on the telomerase activity in human breast carcinoma cells. Cancer Lett. 2005, 227, 153–162. [Google Scholar] [CrossRef]
- Juliana, C.; Fernandes-Alnemri, T.; Wu, J.; Datta, P.; Solorzano, L.; Yu, J.W.; Meng, R.; Quong, A.A.; Latz, E.; Scott, C.P.; et al. Anti-inflammatory compounds parthenolide and Bay 11-7082 are direct inhibitors of the inflammasome. J. Biol. Chem. 2010, 285, 9792–9802. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Xiao, C.; Sun, M.; Tan, M.; Hu, L.; Yu, Q. Parthenolide Inhibits STAT3 Signaling by Covalently Targeting Janus Kinases. Molecules 2018, 23, 1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Yuan, C.; Tian, C.; Li, C.; Nie, F.; Song, X.; Zeng, R.; Wu, D.; Hao, X.; Li, L. The plant sesquiterpene lactone parthenolide inhibits Wnt/beta-catenin signaling by blocking synthesis of the transcriptional regulators TCF4/LEF1. J. Biol. Chem. 2018, 293, 5335–5344. [Google Scholar] [CrossRef] [Green Version]
- Kwak, S.W.; Park, E.S.; Lee, C.S. Parthenolide induces apoptosis by activating the mitochondrial and death receptor pathways and inhibits FAK-mediated cell invasion. Mol. Cell. Biochem. 2014, 385, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.; McGowan, A.; DiNatale, G.J.; Chiramanewong, T.; Cai, T.; Connor, R.E. Hsp72 Is an Intracellular Target of the alpha, beta-Unsaturated Sesquiterpene Lactone, Parthenolide. ACS Omega 2017, 2, 7267–7274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwicker, P.; Schultze, N.; Niehs, S.; Albrecht, D.; Methling, K.; Wurster, M.; Wachlin, G.; Lalk, M.; Lindequist, U.; Haertel, B. Differential effects of Helenalin, an anti-inflammatory sesquiterpene lactone, on the proteome, metabolome and the oxidative stress response in several immune cell types. Toxicol. In Vitro 2017, 40, 45–54. [Google Scholar] [CrossRef]
- Lim, C.B.; Fu, P.Y.; Ky, N.; Zhu, H.S.; Feng, X.; Li, J.; Srinivasan, K.G.; Hamza, M.S.; Zhao, Y. NF-kappaB p65 repression by the sesquiterpene lactone, Helenalin, contributes to the induction of autophagy cell death. BMC Complement. Altern. Med. 2012, 12, 93. [Google Scholar] [CrossRef] [Green Version]
- Lyss, G.; Knorre, A.; Schmidt, T.J.; Pahl, H.L.; Merfort, I. The anti-inflammatory sesquiterpene lactone helenalin inhibits the transcription factor NF-kappaB by directly targeting p65. J. Biol. Chem. 1998, 273, 33508–33516. [Google Scholar] [CrossRef] [Green Version]
- Tornhamre, S.; Schmidt, T.J.; Nasman-Glaser, B.; Ericsson, I.; Lindgren, J.A. Inhibitory effects of helenalin and related compounds on 5-lipoxygenase and leukotriene C(4) synthase in human blood cells. Biochem. Pharm. 2001, 62, 903–911. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, L.; Cheung, H.Y. The inhibitory effect of helenalin on telomerase activity is attributed to the alkylation of the CYS445 residue: Evidence from QM/MM simulations. J. Mol. Graph. Model. 2014, 51, 97–103. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Wang, S.; He, Y.; Huo, Y.; Yang, Z.; Cao, X. Alantolactone induces apoptosis and suppresses migration in MCF7 human breast cancer cells via the p38 MAPK, NFkappaB and Nrf2 signaling pathways. Int. J. Mol. Med. 2018, 42, 1847–1856. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.Y.; Lim, S.S.; Kim, J.R.; Lim, J.S.; Ha, Y.R.; Lee, I.A.; Kim, E.J.; Park, J.H.; Kim, J.S. Nrf2-mediated induction of detoxifying enzymes by alantolactone present in Inula helenium. Phytother. Res. 2008, 22, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.C.; Yu, J.Q.; Yin, Y.; Liu, Y.W.; Zou, G.L. Alantolactone induces activation of apoptosis in human hepatoma cells. Food Chem. Toxicol. 2012, 50, 3313–3319. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Jin, S.E.; Kim, O.S.; Shin, H.K.; Jeong, S.J. Alantolactone from Saussurea lappa Exerts Antiinflammatory Effects by Inhibiting Chemokine Production and STAT1 Phosphorylation in TNF-alpha and IFN-gamma-induced in HaCaT cells. Phytother. Res. 2015, 29, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yang, L.; Kang, Y.; Chen, M.; Lin, S.; Xiang, Y.; Li, C.; Dai, X.; Huang, X.; Liang, G.; et al. Alantolactone sensitizes human pancreatic cancer cells to EGFR inhibitors through the inhibition of STAT3 signaling. Mol. Carcinog. 2019, 58, 565–576. [Google Scholar] [CrossRef]
- Kim, H.L.; Jung, Y.; Park, J.; Youn, D.H.; Kang, J.; Lim, S.; Lee, B.S.; Jeong, M.Y.; Choe, S.K.; Park, R.; et al. Farnesol Has an Anti-obesity Effect in High-Fat Diet-Induced Obese Mice and Induces the Development of Beige Adipocytes in Human Adipose Tissue Derived-Mesenchymal Stem Cells. Front. Pharm. 2017, 8, 654. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.Y.; Hwang, S.T.; Sethi, G.; Fan, L.; Arfuso, F.; Ahn, K.S. Potential Anti-Inflammatory and Anti-Cancer Properties of Farnesol. Molecules 2018, 23, 2827. [Google Scholar] [CrossRef] [Green Version]
- Szucs, G.; Murlasits, Z.; Torok, S.; Kocsis, G.F.; Paloczi, J.; Gorbe, A.; Csont, T.; Csonka, C.; Ferdinandy, P. Cardioprotection by farnesol: Role of the mevalonate pathway. Cardiovasc. Drugs 2013, 27, 269–277. [Google Scholar] [CrossRef]
- De Carvalho, R.B.F.; De Almeida, A.A.C.; Campelo, N.B.; Lellis, D.; Nunes, L.C.C. Nerolidol and its Pharmacological Application in Treating Neurodegenerative Diseases: A Review. Recent Pat. Biotechnol. 2018, 12, 158–168. [Google Scholar] [CrossRef]
- Chan, W.K.; Tan, L.T.; Chan, K.G.; Lee, L.H.; Goh, B.H. Nerolidol: A Sesquiterpene Alcohol with Multi-Faceted Pharmacological and Biological Activities. Molecules 2016, 21, 529. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, D.V.; Salgado, P.R.; de Carvalho, F.L.; Salvadori, M.G.; Penha, A.R.; Leite, F.C.; Borges, C.J.; Piuvezam, M.R.; Pordeus, L.C.; Sousa, D.P.; et al. Nerolidol exhibits antinociceptive and anti-inflammatory activity: Involvement of the GABAergic system and proinflammatory cytokines. Fundam Clin. Pharm. 2016, 30, 14–22. [Google Scholar] [CrossRef]
- Fernandes, M.Y.D.; Carmo, M.; Fonteles, A.A.; Neves, J.C.S.; Silva, A.; Pereira, J.F.; Ferreira, E.O.; Lima, N.M.R.; Neves, K.R.T.; Andrade, G.M. (-)-alpha-bisabolol prevents neuronal damage and memory deficits through reduction of proinflammatory markers induced by permanent focal cerebral ischemia in mice. Eur. J. Pharm. 2019, 842, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Maurya, A.K.; Singh, M.; Dubey, V.; Srivastava, S.; Luqman, S.; Bawankule, D.U. alpha-(-)-bisabolol reduces pro-inflammatory cytokine production and ameliorates skin inflammation. Curr. Pharm. Biotechnol. 2014, 15, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Rocha, N.F.; Rios, E.R.; Carvalho, A.M.; Cerqueira, G.S.; Lopes Ade, A.; Leal, L.K.; Dias, M.L.; de Sousa, D.P.; de Sousa, F.C. Anti-nociceptive and anti-inflammatory activities of (-)-alpha-bisabolol in rodents. Naunyn Schmiedebergs Arch. Pharm. 2011, 384, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Leite Gde, O.; Leite, L.H.; Sampaio Rde, S.; Araruna, M.K.; de Menezes, I.R.; da Costa, J.G.; Campos, A.R. (-)-alpha-Bisabolol attenuates visceral nociception and inflammation in mice. Fitoterapia 2011, 82, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Jin, D.Q.; Mok, H.; Oh, S.J.; Lee, J.U.; Hwang, J.K.; Ha, I.; Han, J.S. Antioxidant and antiinflammatory activities of xanthorrhizol in hippocampal neurons and primary cultured microglia. J. Neurosci. Res. 2005, 82, 831–838. [Google Scholar] [CrossRef]
- Kim, M.B.; Kim, C.; Song, Y.; Hwang, J.K. Antihyperglycemic and Anti-Inflammatory Effects of Standardized Curcuma xanthorrhiza Roxb. Extract and Its Active Compound Xanthorrhizol in High-Fat Diet-Induced Obese Mice. Evid. Based Complement. Altern. Med. 2014, 2014, 205915. [Google Scholar] [CrossRef] [Green Version]
- Oon, S.F.; Nallappan, M.; Tee, T.T.; Shohaimi, S.; Kassim, N.K.; Sa’ariwijaya, M.S.; Cheah, Y.H. Xanthorrhizol: A review of its pharmacological activities and anticancer properties. Cancer Cell Int. 2015, 15, 100. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Chen, X.; Ma, J.; Hassan, W.; Wu, H.; Ling, J.; Shang, J. beta-Elemene attenuates atherosclerosis in apolipoprotein E-deficient mice via restoring NO levels and alleviating oxidative stress. Biomed. Pharm. 2017, 95, 1789–1798. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu, J.; Huo, W.M.; Duan, W.L.; Wang, X.; Shang, J. beta-Elemene reduces the progression of atherosclerosis in rabbits. Chin. J. Nat. Med. 2015, 13, 415–420. [Google Scholar] [CrossRef]
- Jiang, Z.; Jacob, J.A.; Loganathachetti, D.S.; Nainangu, P.; Chen, B. beta-Elemene: Mechanistic Studies on Cancer Cell Interaction and Its Chemosensitization Effect. Front. Pharm. 2017, 8, 105. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Ling, C.; Li, W.; Jiang, H.; Zhi, Q.; Jiang, M. Molecular Mechanisms of Anti-cancer Activities of β-elemene: Targeting Hallmarks of Cancer. Anticancer Agents Med. Chem. 2016, 16, 1426–1434. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lv, Y.; Tian, X.; Lou, J.; An, R.; Zhang, Q.; Li, M.; Xu, L.; Dong, Z. Neuroprotective Effect of beta-Caryophyllene on Cerebral Ischemia-Reperfusion Injury via Regulation of Necroptotic Neuronal Death and Inflammation: In Vivo and In Vitro. Front. Neurosci. 2017, 11, 583. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.; Javed, H.; Azimullah, S.; Haque, M.E. beta-Caryophyllene, a phytocannabinoid attenuates oxidative stress, neuroinflammation, glial activation, and salvages dopaminergic neurons in a rat model of Parkinson disease. Mol. Cell. Biochem. 2016, 418, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Viveros-Paredes, J.M.; Gonzalez-Castaneda, R.E.; Gertsch, J.; Chaparro-Huerta, V.; Lopez-Roa, R.I.; Vazquez-Valls, E.; Beas-Zarate, C.; Camins-Espuny, A.; Flores-Soto, M.E. Neuroprotective Effects of beta-Caryophyllene against Dopaminergic Neuron Injury in a Murine Model of Parkinson’s Disease Induced by MPTP. Pharmaceuticals 2017, 10, 60. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.J.; Kim, J.M.; Lee, J.C.; Kim, W.K.; Chun, H.S. Protective effect of beta-caryophyllene, a natural bicyclic sesquiterpene, against cerebral ischemic injury. J. Med. Food 2013, 16, 471–480. [Google Scholar] [CrossRef]
- Machado, K.D.C.; Islam, M.T.; Ali, E.S.; Rouf, R.; Uddin, S.J.; Dev, S.; Shilpi, J.A.; Shill, M.C.; Reza, H.M.; Das, A.K.; et al. A systematic review on the neuroprotective perspectives of beta-caryophyllene. Phytother. Res. 2018, 32, 2376–2388. [Google Scholar] [CrossRef]
- Saraswati, S.; Alhaider, A.A.; Abdelgadir, A.M. Costunolide suppresses an inflammatory angiogenic response in a subcutaneous murine sponge model. APMIS 2018, 126, 257–266. [Google Scholar] [CrossRef]
- Butturini, E.; Di Paola, R.; Suzuki, H.; Paterniti, I.; Ahmad, A.; Mariotto, S.; Cuzzocrea, S. Costunolide and Dehydrocostuslactone, two natural sesquiterpene lactones, ameliorate the inflammatory process associated to experimental pleurisy in mice. Eur. J. Pharm. 2014, 730, 107–115. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, R.T.; Zhang, P.; Zhang, N.; Yang, C.L.; Yue, L.T.; Li, X.L.; Liu, Y.; Li, H.; Du, J.; et al. Parthenolide inhibits the initiation of experimental autoimmune neuritis. J. Neuroimmunol. 2017, 305, 154–161. [Google Scholar] [CrossRef]
- Popiolek-Barczyk, K.; Kolosowska, N.; Piotrowska, A.; Makuch, W.; Rojewska, E.; Jurga, A.M.; Pilat, D.; Mika, J. Parthenolide Relieves Pain and Promotes M2 Microglia/Macrophage Polarization in Rat Model of Neuropathy. Neural Plast 2015, 2015, 676473. [Google Scholar] [CrossRef]
- Bahabadi, M.; Mohammadalipour, A.; Karimi, J.; Sheikh, N.; Solgi, G.; Goudarzi, F.; Hashemnia, M.; Khodadadi, I. Hepatoprotective effect of parthenolide in rat model of nonalcoholic fatty liver disease. Immunopharmacol. Immunotoxicol. 2017, 39, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Khare, P.; Datusalia, A.K.; Sharma, S.S. Parthenolide, an NF-kappaB Inhibitor Ameliorates Diabetes-Induced Behavioural Deficit, Neurotransmitter Imbalance and Neuroinflammation in Type 2 Diabetes Rat Model. Neuromol. Med. 2017, 19, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, Q. Parthenolide could become a promising and stable drug with anti-inflammatory effects. Nat. Prod. Res. 2015, 29, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhao, J.; Tan, R.; Zhou, H.; Lin, Z.; Zheng, M.; Romas, E.; Xu, J.; Sims, N.A. Parthenolide inhibits pro-inflammatory cytokine production and exhibits protective effects on progression of collagen-induced arthritis in a rat model. Scand. J. Rheumatol. 2015, 44, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R.E.; Archer, M.C. Farnesol induces thyroid hormone receptor (THR) beta1 but inhibits THR-mediated signaling in MCF-7 human breast cancer cells. Biochem. Biophys. Res. Commun. 2006, 343, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, H.; Ren, Y.; Yao, Y.; Hu, J.; Zheng, M.; Ding, Y.; Chen, Y.Y.; Shen, Y.; Wang, L.L.; et al. Local Delivery of beta-Elemene Improves Locomotor Functional Recovery by Alleviating Endoplasmic Reticulum Stress and Reducing Neuronal Apoptosis in Rats with Spinal Cord Injury. Cell Physiol. Biochem. 2018, 49, 595–609. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, J.; Hou, L.; Cui, X. The effect of beta-elemene on alpha-tubulin polymerization in human hepatoma HepG2 cells. Chin. J. Cancer Res. 2013, 25, 770–776. [Google Scholar] [CrossRef]
- Wu, L.; Wang, G.; Tang, S.; Long, G.; Yin, T. Protection of endothelial cells, inhibition of neointimal hyperplasia by beta-elemene in an injured artery. Cardiovasc. Drugs 2011, 25, 233–242. [Google Scholar] [CrossRef]
- Varga, Z.V.; Matyas, C.; Erdelyi, K.; Cinar, R.; Nieri, D.; Chicca, A.; Nemeth, B.T.; Paloczi, J.; Lajtos, T.; Corey, L.; et al. beta-Caryophyllene protects against alcoholic steatohepatitis by attenuating inflammation and metabolic dysregulation in mice. Br. J. Pharm. 2018, 175, 320–334. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Levy, R.M. beta-Caryophyllene promotes osteoblastic mineralization, and suppresses osteoclastogenesis and adipogenesis in mouse bone marrow cultures in vitro. Exp. Med. 2016, 12, 3602–3606. [Google Scholar] [CrossRef]
- Basha, R.H.; Sankaranarayanan, C. beta-Caryophyllene, a natural sesquiterpene, modulates carbohydrate metabolism in streptozotocin-induced diabetic rats. Acta Histochem. 2014, 116, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Tambe, Y.; Tsujiuchi, H.; Honda, G.; Ikeshiro, Y.; Tanaka, S. Gastric cytoprotection of the non-steroidal anti-inflammatory sesquiterpene, beta-caryophyllene. Planta Med. 1996, 62, 469–470. [Google Scholar] [CrossRef] [PubMed]
- Anggakusuma; Yanti; Lee, M.; Hwang, J.K. Estrogenic activity of xanthorrhizol isolated from curcuma xanthorrhiza ROXB. Biol. Pharm. Bull. 2009, 32, 1892–1897. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.Y.; Shim, J.S.; Rukayadi, Y.; Hwang, J.K. Antibacterial activity of xanthorrhizol isolated from Curcuma xanthorrhiza Roxb. against foodborne pathogens. J. Food Prot. 2008, 71, 1926–1930. [Google Scholar] [CrossRef]
- Wellwood, C.R.; Cole, R.A. Relevance of carnosic acid concentrations to the selection of rosemary, Rosmarinus officinalis (L.), accessions for optimization of antioxidant yield. J. Agric. Food Chem. 2004, 52, 6101–6107. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, C. Characterization of phenolic composition in Lamiaceae spices by LC-ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [CrossRef]
- Birtic, S.; Dussort, P.; Pierre, F.X.; Bily, A.C.; Roller, M. Carnosic acid. Phytochemistry 2015, 115, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kang, Y.-G.; Lee, J.-Y.; Choi, D.-H.; Cho, Y.-U.; Shin, J.-M.; Park, J.S.; Lee, J.H.; Kim, W.G.; Seo, D.B.; et al. The natural phytochemical dehydroabietic acid is an anti-aging reagent that mediates the direct activation of SIRT1. Mol. Cell Endocrinol. 2015, 412, 216–225. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, Y.K.; Lee, D.S.; Yoo, J.E.; Shin, M.S.; Yamabe, N.; Kim, S.N.; Lee, S.; Kim, K.H.; Lee, H.J.; et al. Abietic acid isolated from pine resin (Resina Pini) enhances angiogenesis in HUVECs and accelerates cutaneous wound healing in mice. J. Ethnopharmacol. 2017, 203, 279–287. [Google Scholar] [CrossRef]
- Momtazi-Borojeni, A.A.; Esmaeili, S.A.; Abdollahi, E.; Sahebkar, A. A Review on the Pharmacology and Toxicology of Steviol Glycosides Extracted from Stevia rebaudiana. Curr. Pharm. Des. 2017, 23, 1616–1622. [Google Scholar] [CrossRef]
- Bucknall, R.A.; Moores, H.; Simms, R.; Hesp, B. Antiviral effects of aphidicolin, a new antibiotic produced by Cephalosporium aphidicola. Antimicrob. Agents Chemother. 1973, 4, 294–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Zhang, X.; Su, Z.; Xiao, J.; Lv, M.; Cao, Y.; Chen, Y. Carnosol Improved Lifespan and Healthspan by Promoting Antioxidant Capacity in Caenorhabditis elegans. Oxid. Med. Cell. Longev. 2019, 2019, 5958043. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zhang, X.; Xiao, J.; Zhong, Q.; Kuang, Y.; Cao, Y.; Chen, Y. Effects on longevity extension and mechanism of action of carnosic acid in Caenorhabditis elegans. Food Funct. 2019, 10, 1398–1410. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I.; Halliwell, B.; Aeschbach, R.; Loligers, J. Antioxidant and pro-oxidant properties of active rosemary constituents: Carnosol and carnosic acid. Xenobiotica 1992, 22, 257–268. [Google Scholar] [CrossRef]
- Loussouarn, M.; Krieger-Liszkay, A.; Svilar, L.; Bily, A.; Birtic, S.; Havaux, M. Carnosic Acid and Carnosol, Two Major Antioxidants of Rosemary, Act through Different Mechanisms. Plant. Physiol. 2017, 175, 1381–1394. [Google Scholar] [CrossRef] [Green Version]
- Minnunni, M.; Wolleb, U.; Mueller, O.; Pfeifer, A.; Aeschbacher, H.U. Natural antioxidants as inhibitors of oxygen species induced mutagenicity. Mutat. Res. 1992, 269, 193–200. [Google Scholar] [CrossRef]
- Del Bano, M.J.; Castillo, J.; Benavente-Garcia, O.; Lorente, J.; Martin-Gil, R.; Acevedo, C.; Alcaraz, M. Radioprotective-antimutagenic effects of rosemary phenolics against chromosomal damage induced in human lymphocytes by gamma-rays. J. Agric. Food Chem. 2006, 54, 2064–2068. [Google Scholar] [CrossRef]
- Fahim, F.A.; Esmat, A.Y.; Fadel, H.M.; Hassan, K.F. Allied studies on the effect of Rosmarinus officinalis L. on experimental hepatotoxicity and mutagenesis. Int. J. Food Sci. Nutr. 1999, 50, 413–427. [Google Scholar] [CrossRef]
- Dao, T.T.; Le, T.V.; Nguyen, P.H.; Thuong, P.T.; Minh, P.T.; Woo, E.R.; Lee, K.Y.; Oh, W.K. SIRT1 inhibitory diterpenoids from the Vietnamese medicinal plant Croton tonkinensis. Planta Med. 2010, 76, 1011–1014. [Google Scholar] [CrossRef]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Chen, P.Y.; Cheng, T.H.; Chao, H.H.; Liu, J.C.; Chen, J.J. Cafestol Inhibits Cyclic-Strain-Induced Interleukin-8, Intercellular Adhesion Molecule-1, and Monocyte Chemoattractant Protein-1 Production in Vascular Endothelial Cells. Oxid. Med. Cell. Longev. 2018, 2018, 7861518. [Google Scholar] [CrossRef] [Green Version]
- Romo Vaquero, M.; Garcia Villalba, R.; Larrosa, M.; Yanez-Gascon, M.J.; Fromentin, E.; Flanagan, J.; Roller, M.; Tomas-Barberan, F.A.; Espin, J.C.; Garcia-Conesa, M.T. Bioavailability of the major bioactive diterpenoids in a rosemary extract: Metabolic profile in the intestine, liver, plasma, and brain of Zucker rats. Mol. Nutr. Food Res. 2013, 57, 1834–1846. [Google Scholar] [CrossRef] [PubMed]
- Cavin, C.; Holzhaeuser, D.; Scharf, G.; Constable, A.; Huber, W.W.; Schilter, B. Cafestol and kahweol, two coffee specific diterpenes with anticarcinogenic activity. Food Chem. Toxicol. 2002, 40, 1155–1163. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Mabkhot, Y.; Badshah, S.L. Bioactivity Profile of the Diterpene Isosteviol and its Derivatives. Molecules 2019, 24, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curry, L.L.; Roberts, A.; Brown, N. Rebaudioside A: Two-generation reproductive toxicity study in rats. Food Chem. Toxicol. 2008, 46, S21–S30. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.; Bryant, A. Short versus long duration infusions of paclitaxel for any advanced adenocarcinoma. Cochrane Database Syst. Rev. 2011. [Google Scholar] [CrossRef] [Green Version]
- Ohmori, K.; Kawamura, Y. Cell transformation activities of abietic acid and dehydroabietic acid: Safety assessment of possible contaminants in paper and paperboard for food contact use. Food Addit. Contam. Part. A Chem. Anal. Control Expo. Risk Assess. 2009, 26, 568–573. [Google Scholar] [CrossRef]
- Christianson-Heiska, I.L.; Haavisto, T.; Paranko, J.; Bergelin, E.; Isomaa, B. Effects of the wood extractives dehydroabietic acid and betulinol on reproductive physiology of zebrafish (Danio rerio)-a two-generation study. Aquat. Toxicol. 2008, 86, 388–396. [Google Scholar] [CrossRef]
- Pandelides, Z.; Guchardi, J.; Holdway, D. Dehydroabietic acid (DHAA) alters metabolic enzyme activity and the effects of 17beta-estradiol in rainbow trout (Oncorhynchus mykiss). Ecotoxicol. Environ. Saf. 2014, 101, 168–176. [Google Scholar] [CrossRef]
- Stegelmeier, B.L.; Gardner, D.R.; James, L.F.; Panter, K.E.; Molyneux, R.J. The toxic and abortifacient effects of ponderosa pine. Vet. Pathol. 1996, 33, 22–28. [Google Scholar] [CrossRef]
- Shimojo, Y.; Kosaka, K.; Noda, Y.; Shimizu, T.; Shirasawa, T. Effect of rosmarinic acid in motor dysfunction and life span in a mouse model of familial amyotrophic lateral sclerosis. J. Neurosci. Res. 2010, 88, 896–904. [Google Scholar] [CrossRef]
- Ninomiya, K.; Matsuda, H.; Shimoda, H.; Nishida, N.; Kasajima, N.; Yoshino, T.; Morikawa, T.; Yoshikawa, M. Carnosic acid, a new class of lipid absorption inhibitor from sage. Bioorg. Med. Chem. Lett. 2004, 14, 1943–1946. [Google Scholar] [CrossRef] [PubMed]
- Fukuma, Y.; Sakai, E.; Nishishita, K.; Okamoto, K.; Tsukuba, T. Cafestol has a weaker inhibitory effect on osteoclastogenesis than kahweol and promotes osteoblast differentiation. BioFactors 2015, 41, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Poeckel, D.; Greiner, C.; Verhoff, M.; Rau, O.; Tausch, L.; Hornig, C.; Steinhilber, D.; Schubert-Zsilavecz, M.; Werz, O. Carnosic acid and carnosol potently inhibit human 5-lipoxygenase and suppress pro-inflammatory responses of stimulated human polymorphonuclear leukocytes. Biochem. Pharm. 2008, 76, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, K.; Mimura, J.; Itoh, K.; Satoh, T.; Shimojo, Y.; Kitajima, C.; Maruyama, A.; Yamamoto, M.; Shirasawa, T. Role of Nrf2 and p62/ZIP in the neurite outgrowth by carnosic acid in PC12h cells. J. Biochem. 2010, 147, 73–81. [Google Scholar] [CrossRef]
- Johnson, J.J. Carnosol: A promising anti-cancer and anti-inflammatory agent. Cancer Lett. 2011, 305, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.P.; Jeong, H.G. The coffee diterpene kahweol induces heme oxygenase-1 via the PI3K and p38/Nrf2 pathway to protect human dopaminergic neurons from 6-hydroxydopamine-derived oxidative stress. FEBS Lett. 2008, 582, 2655–2662. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Guo, M.; Song, X.; Zhang, Z.; Jiang, H.; Wang, W.; Fu, Y.; Cao, Y.; Zhu, L.; Zhang, N. Stevioside plays an anti-inflammatory role by regulating the NF-kappaB and MAPK pathways in S. aureus-infected mouse mammary glands. Inflammation 2014, 37, 1837–1846. [Google Scholar] [CrossRef]
- Boonkaewwan, C.; Ao, M.; Toskulkao, C.; Rao, M.C. Specific immunomodulatory and secretory activities of stevioside and steviol in intestinal cells. J. Agric. Food Chem. 2008, 56, 3777–3784. [Google Scholar] [CrossRef]
- Yingkun, N.; Zhenyu, W.; Jing, L.; Xiuyun, L.; Huimin, Y. Stevioside protects LPS-induced acute lung injury in mice. Inflammation 2013, 36, 242–250. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Guo, J.H.; Wu, Z.L.; Xiong, Y.; Zhou, W.L. Innate immune responses of epididymal epithelial cells to Staphylococcus aureus infection. Immunol. Lett. 2008, 119, 84–90. [Google Scholar] [CrossRef]
- Boonkaewwan, C.; Burodom, A. Anti-inflammatory and immunomodulatory activities of stevioside and steviol on colonic epithelial cells. J. Sci. Food Agric. 2013, 93, 3820–3825. [Google Scholar] [CrossRef] [PubMed]
- Boonkaewwan, C.; Toskulkao, C.; Vongsakul, M. Anti-Inflammatory and Immunomodulatory Activities of Stevioside and Its Metabolite Steviol on THP-1 Cells. J. Agric. Food Chem. 2006, 54, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Yarimizu, T.; Mitamura, T.; Suzuki, S.; Sakamoto, S. Protective effects of an antiulcer agent, ecabet sodium on colorectal carcinogenesis in rodents. Oncol. Rep. 1998, 5, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Hirai, S.; Goto, T.; Kuroyanagi, K.; Lee, J.Y.; Uemura, T.; Ezaki, Y.; Takahashi, N.; Kawada, T. Dehydroabietic acid, a phytochemical, acts as ligand for PPARs in macrophages and adipocytes to regulate inflammation. Biochem. Biophys. Res. Commun. 2008, 369, 333–338. [Google Scholar] [CrossRef]
- De Oliveira, M.R. The Dietary Components Carnosic Acid and Carnosol as Neuroprotective Agents: A Mechanistic View. Mol. Neurobiol. 2016, 53, 6155–6168. [Google Scholar] [CrossRef]
- Mellbye, F.B.; Jeppesen, P.B.; Hermansen, K.; Gregersen, S. Cafestol, a Bioactive Substance in Coffee, Stimulates Insulin Secretion and Increases Glucose Uptake in Muscle Cells: Studies in Vitro. J. Nat. Prod. 2015, 78, 2447–2451. [Google Scholar] [CrossRef]
- Mellbye, F.B.; Jeppesen, P.B.; Shokouh, P.; Laustsen, C.; Hermansen, K.; Gregersen, S. Cafestol, a Bioactive Substance in Coffee, Has Antidiabetic Properties in KKAy Mice. J. Nat. Prod. 2017, 80, 2353–2359. [Google Scholar] [CrossRef] [PubMed]
- Moeenfard, M.; Cortez, A.; Machado, V.; Costa, R.; Luis, C.; Coelho, P.; Soares, R.; Alves, A.; Borges, N.; Santos, A. Anti-Angiogenic Properties of Cafestol and Kahweol Palmitate Diterpene Esters. J. Cell. Biochem. 2016, 117, 2748–2756. [Google Scholar] [CrossRef] [Green Version]
- McCall, A.L.; Millington, W.R.; Wurtman, R.J. Blood-brain barrier transport of caffeine: Dose-related restriction of adenine transport. Life Sci. 1982, 31, 2709–2715. [Google Scholar] [CrossRef]
- Trinh, K.; Andrews, L.; Krause, J.; Hanak, T.; Lee, D.; Gelb, M.; Pallanck, L. Decaffeinated coffee and nicotine-free tobacco provide neuroprotection in Drosophila models of Parkinson’s disease through an NRF2-dependent mechanism. J. Neurosci. 2010, 30, 5525–5532. [Google Scholar] [CrossRef]
- Gregersen, S.; Jeppesen, P.B.; Holst, J.J.; Hermansen, K. Antihyperglycemic effects of stevioside in type 2 diabetic subjects. Metabolism 2004, 53, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, P.B.; Gregersen, S.; Poulsen, C.R.; Hermansen, K. Stevioside acts directly on pancreatic beta cells to secrete insulin: Actions independent of cyclic adenosine monophosphate and adenosine triphosphate-sensitive K+-channel activity. Metabolism 2000, 49, 208–214. [Google Scholar] [CrossRef]
- Jeppesen, P.B.; Gregersen, S.; Rolfsen, S.E.; Jepsen, M.; Colombo, M.; Agger, A.; Xiao, J.; Kruhoffer, M.; Orntoft, T.; Hermansen, K. Antihyperglycemic and blood pressure-reducing effects of stevioside in the diabetic Goto-Kakizaki rat. Metabolism 2003, 52, 372–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Hou, K.; Qin, P.; Liu, H.; Yi, B.; Yang, W.; Wu, W. RNA-Seq for gene identification and transcript profiling of three Stevia rebaudiana genotypes. BMC Genom. 2014, 15, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melis, M.S. Stevioside effect on renal function of normal and hypertensive rats. J. Ethnopharmacol. 1992, 36, 213–217. [Google Scholar] [CrossRef]
- Li, R.; Morris-Natschke, S.L.; Lee, K.H. Clerodane diterpenes: Sources, structures, and biological activities. Nat. Prod. Rep. 2016, 33, 1166–1226. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.C.; Chen, C.F.; Chiou, W.F. Andrographolide prevents oxygen radical production by human neutrophils: Possible mechanism(s) involved in its anti-inflammatory effect. Br. J. Pharm. 2002, 135, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Kishore, V.; Yarla, N.S.; Bishayee, A.; Putta, S.; Malla, R.; Neelapu, N.R.; Challa, S.; Das, S.; Shiralgi, Y.; Hegde, G.; et al. Multi-targeting Andrographolide and its Natural Analogs as Potential Therapeutic Agents. Curr. Top. Med. Chem. 2017, 17, 845–857. [Google Scholar] [CrossRef]
- Xia, Y.F.; Ye, B.Q.; Li, Y.D.; Wang, J.G.; He, X.J.; Lin, X.; Yao, X.; Ma, D.; Slungaard, A.; Hebbel, R.P.; et al. Andrographolide attenuates inflammation by inhibition of NF-kappa B activation through covalent modification of reduced cysteine 62 of p50. J. Immunol. 2004, 173, 4207–4217. [Google Scholar] [CrossRef] [Green Version]
- Batkhuu, J.; Hattori, K.; Takano, F.; Fushiya, S.; Oshiman, K.; Fujimiya, Y. Suppression of NO production in activated macrophages in vitro and ex vivo by neoandrographolide isolated from Andrographis paniculata. Biol. Pharm. Bull. 2002, 25, 1169–1174. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Wang, G.; Pan, W. Andrographolide Inhibits Proliferation and Metastasis of SGC7901 Gastric Cancer Cells. Biomed. Res. Int. 2017, 2017, 6242103. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, M.; Chattopadhyay, S.; Choudhuri, T.; Bera, R.; Kumar, S.; Chakraborty, B.; Mukherjee, S.K. Cytotoxicity and cell cycle arrest induced by andrographolide lead to programmed cell death of MDA-MB-231 breast cancer cell line. J. Biomed. Sci. 2016, 23, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannaste, A.; Laanisto, L.; Pazouki, L.; Copolovici, L.; Suhorutsenko, M.; Azeem, M.; Toom, L.; Borg-Karlson, A.K.; Niinemets, U. Diterpenoid fingerprints in pine foliage across an environmental and chemotypic matrix: Isoabienol content is a key trait differentiating chemotypes. Phytochemistry 2018, 147, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Hillier, S.G.; Lathe, R. Terpenes, hormones and life: Isoprene rule revisited. J. Endocrinol. 2019, 242, R9–R22. [Google Scholar] [CrossRef]
- Kushiro, T.; Ebizuka, Y. 1.18–Triterpenes. In Comprehensive Natural Products II’; Liu, H.-W., Mander, L., Eds.; Elsevier: Oxford, ME, USA, 2010; pp. 673–708. [Google Scholar] [CrossRef]
- Jesus, J.A.; Lago, J.H.; Laurenti, M.D.; Yamamoto, E.S.; Passero, L.F. Antimicrobial activity of oleanolic and ursolic acids: An update. Evid. Based Complement. Altern. Med. 2015, 2015, 620472. [Google Scholar] [CrossRef]
- Salvador, J.A.R.; Leal, A.S.; Valdeira, A.S.; Goncalves, B.M.F.; Alho, D.P.S.; Figueiredo, S.A.C.; Silvestre, S.M.; Mendes, V.I.S. Oleanane-, ursane-, and quinone methide friedelane-type triterpenoid derivatives: Recent advances in cancer treatment. Eur. J. Med. Chem. 2017, 142, 95–130. [Google Scholar] [CrossRef]
- Ammon, H.P. Boswellic Acids and Their Role in Chronic Inflammatory Diseases. Adv. Exp. Med. Biol. 2016, 928, 291–327. [Google Scholar] [CrossRef]
- Sharma, H.; Kumar, P.; Deshmukh, R.R.; Bishayee, A.; Kumar, S. Pentacyclic triterpenes: New tools to fight metabolic syndrome. Phytomedicine 2018, 50, 166–177. [Google Scholar] [CrossRef]
- Siddique, H.R.; Saleem, M. Beneficial health effects of lupeol triterpene: A review of preclinical studies. Life Sci. 2011, 88, 285–293. [Google Scholar] [CrossRef]
- Seo, D.Y.; Lee, S.R.; Heo, J.W.; No, M.H.; Rhee, B.D.; Ko, K.S.; Kwak, H.B.; Han, J. Ursolic acid in health and disease. Korean J. Physiol. Pharm. 2018, 22, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Lou-Bonafonte, J.M.; Martinez-Beamonte, R.; Sanclemente, T.; Surra, J.C.; Herrera-Marcos, L.V.; Sanchez-Marco, J.; Arnal, C.; Osada, J. Current Insights into the Biological Action of Squalene. Mol. Nutr. Food Res. 2018, e1800136. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Karadeniz, F. Biological importance and applications of squalene and squalane. Adv. Food Nutr. Res. 2012, 65, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Popa, O.; Babeanu, N.E.; Popa, I.; Nita, S.; Dinu-Parvu, C.E. Methods for obtaining and determination of squalene from natural sources. Biomed. Res. Int. 2015, 2015, 367202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhilwade, H.N.; Tatewaki, N.; Nishida, H.; Konishi, T. Squalene as novel food factor. Curr. Pharm. Biotechnol. 2010, 11, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Camino, M.C.; Cert, A. Quantitative determination of hydroxy pentacyclic triterpene acids in vegetable oils. J. Agric. Food Chem. 1999, 47, 1558–1562. [Google Scholar] [CrossRef] [PubMed]
- Jager, S.; Trojan, H.; Kopp, T.; Laszczyk, M.N.; Scheffler, A. Pentacyclic triterpene distribution in various plants–rich sources for a new group of multi-potent plant extracts. Molecules 2009, 14, 2016–2031. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.G.; Lee, C.C.; Dong, Z.; Lee, K.W. Ginsenosides and their metabolites: A review of their pharmacological activities in the skin. Arch. Derm. Res. 2015, 307, 397–403. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, P.; Wang, Y.; Wu, L.; Wang, X.; Zhang, K.; Liu, Q. In Vitro and In Vivo Activity of Fomitopsis Pinicola (Sw. Ex Fr.) Karst Chloroform (Fpkc) Extract Against S180 Tumor Cells. Cell Physiol. Biochem. 2017, 44, 2042–2056. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Swanson, K.; Eliaz, I.; McClintick, J.N.; Sandusky, G.E.; Sliva, D. Pachymic acid inhibits growth and induces apoptosis of pancreatic cancer in vitro and in vivo by targeting ER stress. PLoS ONE 2015, 10, e0122270. [Google Scholar] [CrossRef]
- Wang, J.; Cao, B.; Zhao, H.; Feng, J. Emerging Roles of Ganoderma Lucidum in Anti-Aging. Aging Dis. 2017, 8, 691–707. [Google Scholar] [CrossRef] [Green Version]
- Crowley, V.M.; Ayi, K.; Lu, Z.; Liby, K.T.; Sporn, M.; Kain, K.C. Synthetic oleanane triterpenoids enhance blood brain barrier integrity and improve survival in experimental cerebral malaria. Malar J. 2017, 16, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega-Munoz, M.; Rodriguez-Serrano, F.; De Los Reyes-Berbel, E.; Mut-Salud, N.; Hernandez-Mateo, F.; Rodriguez-Lopez, A.; Garrido, J.M.; Lopez-Jaramillo, F.J.; Santoyo-Gonzalez, F. Biological Evaluation and Docking Studies of Synthetic Oleanane-type Triterpenoids. ACS Omega 2018, 3, 11455–11468. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, B.; Zhang, Z.; Wei, Y.; Xu, Z.; Qin, S.; Liu, N.; Zhao, R.; Peng, J.; Yang, G.; et al. Synthesis and Discovery Novel Anti-Cancer Stem Cells Compounds Derived from the Natural Triterpenoic Acids. J. Med. Chem. 2018, 61, 10814–10833. [Google Scholar] [CrossRef] [PubMed]
- Satheesan, J.; Narayanan, A.K.; Sakunthala, M. Induction of root colonization by Piriformospora indica leads to enhanced asiaticoside production in Centella asiatica. Mycorrhiza 2012, 22, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Negi, H.; Shukla, A.; Khan, F.; Pandey, R. 3beta-Hydroxy-urs-12-en-28-oic acid prolongs lifespan in C. elegans by modulating JNK-1. Biochem. Biophys. Res. Commun. 2016, 480, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Negi, H.; Saikia, S.K.; Pandey, R. 3beta-Hydroxy-urs-12-en-28-oic Acid Modulates Dietary Restriction Mediated Longevity and Ameliorates Toxic Protein Aggregation in C. Elegans. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 1614–1619. [Google Scholar] [CrossRef]
- Vayndorf, E.M.; Lee, S.S.; Liu, R.H. Whole apple extracts increase lifespan, healthspan and resistance to stress in Caenorhabditis elegans. J. Funct. Foods 2013, 5, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Altun, D.; Ayar, A.; Uysal, H.; Kara, A.A.; Unal, E.L. Extended longevity of Drosophila melanogaster by water and ethanol extracts of Stachys lavandulifolia. Pharm. Biol. 2010, 48, 1291–1296. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, L.; Zhou, L. Oleanolic acid activates daf-16 to increase lifespan in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2015, 468, 843–849. [Google Scholar] [CrossRef]
- Jattujan, P.; Chalorak, P.; Siangcham, T.; Sangpairoj, K.; Nobsathian, S.; Poomtong, T.; Sobhon, P.; Meemon, K. Holothuria scabra extracts possess anti-oxidant activity and promote stress resistance and lifespan extension in Caenorhabditis elegans. Exp. Gerontol. 2018, 110, 158–171. [Google Scholar] [CrossRef]
- Cao, X.; Sun, Y.; Lin, Y.; Pan, Y.; Farooq, U.; Xiang, L.; Qi, J. Antiaging of Cucurbitane Glycosides from Fruits of Momordica charantia L. Oxid. Med. Cell. Longev. 2018, 2018, 1538632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Choi, S.H.; Kwon, O.S.; Shin, T.J.; Lee, J.H.; Lee, B.H.; Yoon, I.S.; Pyo, M.K.; Rhim, H.; Lim, Y.H.; et al. Effects of ginsenosides, active ingredients of Panax ginseng, on development, growth, and life span of Caenorhabditis elegans. Biol. Pharm. Bull. 2007, 30, 2126–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papaevgeniou, N.; Sakellari, M.; Jha, S.; Tavernarakis, N.; Holmberg, C.I.; Gonos, E.S.; Chondrogianni, N. 18alpha-Glycyrrhetinic Acid Proteasome Activator Decelerates Aging and Alzheimer’s Disease Progression in Caenorhabditis elegans and Neuronal Cultures. Antioxid. Redox. Signal. 2016, 25, 855–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.K.; Vatsa, N.; Nelson, V.K.; Kumar, V.; Kumar, S.S.; Mandal, S.C.; Pal, M.; Jana, N.R. Azadiradione Restores Protein Quality Control and Ameliorates the Disease Pathogenesis in a Mouse Model of Huntington’s Disease. Mol. Neurobiol. 2018, 55, 6337–6346. [Google Scholar] [CrossRef]
- Kiaei, M.; Kipiani, K.; Petri, S.; Chen, J.; Calingasan, N.Y.; Beal, M.F. Celastrol blocks neuronal cell death and extends life in transgenic mouse model of amyotrophic lateral sclerosis. Neurodegener Dis. 2005, 2, 246–254. [Google Scholar] [CrossRef]
- Tsai, C.W.; Tsai, R.T.; Liu, S.P.; Chen, C.S.; Tsai, M.C.; Chien, S.H.; Hung, H.S.; Lin, S.Z.; Shyu, W.C.; Fu, R.H. Neuroprotective Effects of Betulin in Pharmacological and Transgenic Caenorhabditis elegans Models of Parkinson’s Disease. Cell Transpl. 2017, 26, 1903–1918. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yao, H.; Chen, X.; Wang, Z.; Xiang, Y.; Xia, J.; Liu, Y.; Wang, Y. Ginsenoside Rg1 Decreases Oxidative Stress and Down-Regulates Akt/mTOR Signalling to Attenuate Cognitive Impairment in Mice and Senescence of Neural Stem Cells Induced by D-Galactose. Neurochem. Res. 2018, 43, 430–440. [Google Scholar] [CrossRef]
- Smina, T.P.; Joseph, J.; Janardhanan, K.K. Ganoderma lucidum total triterpenes prevent gamma-radiation induced oxidative stress in Swiss albino mice In Vivo. Redox Rep. 2016, 21, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.M.; Park, S.N. Inhibitory Effect of Lupeol on MMPs Expression using Aged Fibroblast through Repeated UVA Irradiation. Photochem. Photobiol. 2019, 95, 587–594. [Google Scholar] [CrossRef]
- Bahrami, S.A.; Bakhtiari, N. Ursolic acid regulates aging process through enhancing of metabolic sensor proteins level. Biomed. Pharm. 2016, 82, 8–14. [Google Scholar] [CrossRef]
- Buddhan, S.; Sivakumar, R.; Dhandapani, N.; Ganesan, B.; Anandan, R. Protective effect of dietary squalene supplementation on mitochondrial function in liver of aged rats. Prostaglandins Leukot Essent Fat. Acids 2007, 76, 349–355. [Google Scholar] [CrossRef]
- Sun, J.; Jiao, C.; Ma, Y.; Chen, J.; Wu, W.; Liu, S. Anti-ageing effect of red ginseng revealed by urinary metabonomics using RRLC-Q-TOF-MS. Phytochem. Anal. 2018, 29, 387–397. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Ling, L.; Wei, T.; Wang, Y.; Xiong, Z. Ginsenoside Rg1 improves fertility and reduces ovarian pathological damages in premature ovarian failure model of mice. Exp. Biol. Med. (Maywood) 2017, 242, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhavadas, S.; Subramanian, S. Combination of Spirulina with glycyrrhizin prevents cognitive dysfunction in aged obese rats. Indian J. Pharm. 2015, 47, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Nagai, N.; Yagyu, S.; Hata, A.; Nirengi, S.; Kotani, K.; Moritani, T.; Sakane, N. Maslinic acid derived from olive fruit in combination with resistance training improves muscle mass and mobility functions in the elderly. J. Clin. Biochem. Nutr. 2019, 64, 224–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, N.J. Dietary safety of cycloastragenol from Astragalus spp.: Subchronic toxicity and genotoxicity studies. Food Chem. Toxicol. 2014, 64, 322–334. [Google Scholar] [CrossRef]
- Akbarsha, M.A.; Palanisamy, M.; Murugaian, P.; Lakshmi Latha, P.N. Ursolic acid generates symplasts in rat spermatogenic clones. Phytother. Res. 1998, 12, 32–36. [Google Scholar] [CrossRef]
- Cho, S.; Choi, C.W.; Lee, D.H.; Won, C.H.; Kim, S.M.; Lee, S.; Lee, M.J.; Chung, J.H. High-dose squalene ingestion increases type I procollagen and decreases ultraviolet-induced DNA damage in human skin in vivo but is associated with transient adverse effects. Clin. Exp. Derm. 2009, 34, 500–508. [Google Scholar] [CrossRef]
- Lu, J.; Zheng, Y.L.; Wu, D.M.; Luo, L.; Sun, D.X.; Shan, Q. Ursolic acid ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by D-galactose. Biochem. Pharm. 2007, 74, 1078–1090. [Google Scholar] [CrossRef]
- Park, H.J.; Kwon, H.; Lee, S.; Jung, J.W.; Ryu, J.H.; Jang, D.S.; Lee, Y.C.; Kim, D.H. Echinocystic Acid Facilitates Neurite Outgrowth in Neuroblastoma Neuro2a Cells and Enhances Spatial Memory in Aged Mice. Biol. Pharm. Bull. 2017, 40, 1724–1729. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.W.; Choi, H.J.; Kim, N.J.; Kim, D.H. Anxiolytic-like effects of ginsenosides Rg3 and Rh2 from red ginseng in the elevated plus-maze model. Planta Med. 2009, 75, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.C.; Kang, Y.S.; Noh, E.B.; Seo, B.W.; Seo, D.Y.; Park, G.D.; Kim, S.H. Concurrent treatment with ursolic acid and low-intensity treadmill exercise improves muscle atrophy and related outcomes in rats. Korean J. Physiol. Pharm. 2018, 22, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Sung, B.; Kang, Y.J.; Kim, D.H.; Lee, Y.; Hwang, S.Y.; Yoon, J.H.; Yoo, M.A.; Kim, C.M.; Chung, H.Y.; et al. The combination of ursolic acid and leucine potentiates the differentiation of C2C12 murine myoblasts through the mTOR signaling pathway. Int. J. Mol. Med. 2015, 35, 755–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, S.M.; Dyle, M.C.; Bullard, S.A.; Dierdorff, J.M.; Murry, D.J.; Fox, D.K.; Bongers, K.S.; Lira, V.A.; Meyerholz, D.K.; Talley, J.J.; et al. Identification and Small Molecule Inhibition of an Activating Transcription Factor 4 (ATF4)-dependent Pathway to Age-related Skeletal Muscle Weakness and Atrophy. J. Biol. Chem. 2015, 290, 25497–25511. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, S.D.; Suneja, M.; Ebert, S.M.; Bongers, K.S.; Fox, D.K.; Malmberg, S.E.; Alipour, F.; Shields, R.K.; Adams, C.M. mRNA expression signatures of human skeletal muscle atrophy identify a natural compound that increases muscle mass. Cell Metab. 2011, 13, 627–638. [Google Scholar] [CrossRef] [Green Version]
- Staats, S.; Wagner, A.E.; Luersen, K.; Kunstner, A.; Meyer, T.; Kahns, A.K.; Derer, S.; Graspeuntner, S.; Rupp, J.; Busch, H.; et al. Dietary ursolic acid improves health span and life span in male Drosophila melanogaster. BioFactors 2019, 45, 169–186. [Google Scholar] [CrossRef]
- Gill, B.S.; Kumar, S.; Navgeet. Triterpenes in cancer: Significance and their influence. Mol. Biol. Rep. 2016, 43, 881–896. [Google Scholar] [CrossRef]
- Lee, S.G.; Kim, M.M. Pachymic acid promotes induction of autophagy related to IGF-1 signaling pathway in WI-38 cells. Phytomedicine 2017, 36, 82–87. [Google Scholar] [CrossRef]
- Li, B.; Wu, G.L.; Dai, W.; Wang, G.; Su, H.Y.; Shen, X.P.; Zhan, R.; Xie, J.M.; Wang, Z.; Qin, Z.H.; et al. Aescin-induced reactive oxygen species play a pro-survival role in human cancer cells via ATM/AMPK/ULK1-mediated autophagy. Acta Pharm. Sin. 2018, 39, 1874–1884. [Google Scholar] [CrossRef]
- Sohretoglu, D.; Zhang, C.; Luo, J.; Huang, S. ReishiMax inhibits mTORC1/2 by activating AMPK and inhibiting IGFR/PI3K/Rheb in tumor cells. Signal. Transduct. Target. 2019, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Zhang, B.; Li, P.; Wen, X.; Yang, J. Maslinic Acid Inhibits Colon Tumorigenesis by the AMPK-mTOR Signaling Pathway. J. Agric. Food Chem. 2019, 67, 4259–4272. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Wang, Y.; Qin, Y.; Gong, X.G. Oleanolic acid induces autophagic death in human gastric cancer cells In Vitro and In Vivo. Cell Biol. Int. 2016, 40, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, N.; Zhang, H.; Liu, C.; Dong, X.; Wang, X.; Zhu, Y.; Xu, C.; Liu, L.; Yang, S.; et al. Celastrol prevents cadmium-induced neuronal cell death by blocking reactive oxygen species-mediated mammalian target of rapamycin pathway. Br. J. Pharm. 2017, 174, 82–100. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, Y.; Zeng, S.; Lu, J.; Jiang, T.M.; Li, Y.M. Protective effect of astragaloside IV on lipopolysaccharide-induced cardiac dysfunction via downregulation of inflammatory signaling in mice. Immunopharmacol. Immunotoxicol. 2015, 37, 428–433. [Google Scholar] [CrossRef]
- Dai, Y.; Desano, J.; Tang, W.; Meng, X.; Meng, Y.; Burstein, E.; Lawrence, T.S.; Xu, L. Natural proteasome inhibitor celastrol suppresses androgen-independent prostate cancer progression by modulating apoptotic proteins and NF-kappaB. PLoS ONE 2010, 5, e14153. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, K.; Inoue, M.; Matsumoto, Y.; Sakakibara, C.; Miyataka, H.; Matsumoto, H.; Arihara, S. Lanostane triterpenoids and triterpene glycosides from the fruit body of Fomitopsis pinicola and their inhibitory activity against COX-1 and COX-2. J. Nat. Prod. 2005, 68, 69–73. [Google Scholar] [CrossRef]
- Shishodia, S.; Majumdar, S.; Banerjee, S.; Aggarwal, B.B. Ursolic acid inhibits nuclear factor-kappaB activation induced by carcinogenic agents through suppression of IkappaBalpha kinase and p65 phosphorylation: Correlation with down-regulation of cyclooxygenase 2, matrix metalloproteinase 9, and cyclin D1. Cancer Res. 2003, 63, 4375–4383. [Google Scholar]
- Gao, J.; Tang, X.; Dou, H.; Fan, Y.; Zhao, X.; Xu, Q. Hepatoprotective activity of Terminalia catappa L. leaves and its two triterpenoids. J. Pharm. Pharm. 2004, 56, 1449–1455. [Google Scholar] [CrossRef]
- Lee, W.; Kim, J.; Park, E.K.; Bae, J.S. Maslinic Acid Ameliorates Inflammation via the Downregulation of NF-kappaB and STAT-1. Antioxidants 2020, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Fukumitsu, S.; Villareal, M.O.; Fujitsuka, T.; Aida, K.; Isoda, H. Anti-inflammatory and anti-arthritic effects of pentacyclic triterpenoids maslinic acid through NF-kappaB inactivation. Mol. Nutr. Food Res. 2016, 60, 399–409. [Google Scholar] [CrossRef]
- Yang, X.W.; Ma, L.Y.; Zhou, Q.L.; Xu, W.; Zhang, Y.B. SIRT1 activator isolated from artificial gastric juice incubate of total saponins in stems and leaves of Panax ginseng. Bioorg. Med. Chem. Lett. 2018, 28, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Bakhtiari, N.; Mirzaie, S.; Hemmati, R.; Moslemee-Jalalvand, E.; Noori, A.R.; Kazemi, J. Mounting evidence validates Ursolic Acid directly activates SIRT1: A powerful STAC which mimic endogenous activator of SIRT1. Arch. Biochem. Biophys. 2018, 650, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bakhtiari, N.; Hosseinkhani, S.; Tashakor, A.; Hemmati, R. Ursolic acid ameliorates aging-metabolic phenotype through promoting of skeletal muscle rejuvenation. Med. Hypotheses 2015, 85, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sudhahar, V.; Kumar, S.A.; Varalakshmi, P.; Sundarapandiyan, R. Mitigating role of lupeol and lupeol linoleate on hepatic lipemic-oxidative injury and lipoprotein peroxidation in experimental hypercholesterolemia. Mol. Cell. Biochem. 2007, 295, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Asha, R.; Gayathri Devi, V.; Abraham, A. Lupeol, a pentacyclic triterpenoid isolated from Vernonia cinerea attenuate selenite induced cataract formation in Sprague Dawley rat pups. Chem. Biol. Interact. 2016, 245, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Huang, R.; Zhang, S.; Wei, L.; Zhuo, L.; Wu, X.; Tang, A.; Huang, Q. Beneficial effects of asiaticoside on cognitive deficits in senescence-accelerated mice. Fitoterapia 2013, 87, 69–77. [Google Scholar] [CrossRef]
- Laszczyk, M.N. Pentacyclic triterpenes of the lupane, oleanane and ursane group as tools in cancer therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.; Madan, E.; Nigam, N.; Roy, P.; George, J.; Shukla, Y. Induction of apoptosis by lupeol in human epidermoid carcinoma A431 cells through regulation of mitochondrial, Akt/PKB and NFkappaB signaling pathways. Cancer Biol. 2009, 8, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- De Los Reyes, M.M.; Oyong, G.G.; VA, S.N.; Shen, C.C.; Ragasa, C.Y. Cytotoxic Compounds from Wrightia pubescens (R.Br.). Pharmacogn. Res. 2018, 10, 9–15. [Google Scholar] [CrossRef]
- Qu, L.; Li, S.; Zhuo, Y.; Chen, J.; Qin, X.; Guo, G. Anticancer effect of triterpenes from Ganoderma lucidum in human prostate cancer cells. Oncol. Lett. 2017, 14, 7467–7472. [Google Scholar] [CrossRef] [Green Version]
- Valdes, K.; Morales, J.; Rodriguez, L.; Gunther, G. Potential use of nanocarriers with pentacyclic triterpenes in cancer treatments. Nanomedicine 2016, 11, 3139–3156. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Alvarado, H.L.; Abrego, G.; Martins-Gomes, C.; Garduno-Ramirez, M.L.; Garcia, M.L.; Calpena, A.C.; Souto, E.B. In Vitro Cytotoxicity of Oleanolic/Ursolic Acids-Loaded in PLGA Nanoparticles in Different Cell Lines. Pharmaceutics 2019, 11, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthilkumar, S.; Yogeeta, S.K.; Subashini, R.; Devaki, T. Attenuation of cyclophosphamide induced toxicity by squalene in experimental rats. Chem. Biol. Interact. 2006, 160, 252–260. [Google Scholar] [CrossRef]
- Senthilkumar, S.; Devaki, T.; Manohar, B.M.; Babu, M.S. Effect of squalene on cyclophosphamide-induced toxicity. Clin. Chim. Acta 2006, 364, 335–342. [Google Scholar] [CrossRef]
- Narayan Bhilwade, H.; Tatewaki, N.; Konishi, T.; Nishida, M.; Eitsuka, T.; Yasui, H.; Inanami, O.; Handa, O.; Naito, Y.; Ikekawa, N.; et al. The Adjuvant Effect of Squalene, an Active Ingredient of Functional Foods, on Doxorubicin-Treated Allograft Mice. Nutr. Cancer 2019, 71, 1153–1164. [Google Scholar] [CrossRef]
- Arias, J.L.; Reddy, L.H.; Othman, M.; Gillet, B.; Desmaele, D.; Zouhiri, F.; Dosio, F.; Gref, R.; Couvreur, P. Squalene based nanocomposites: A new platform for the design of multifunctional pharmaceutical theragnostics. ACS Nano 2011, 5, 1513–1521. [Google Scholar] [CrossRef]
- Bui, D.T.; Nicolas, J.; Maksimenko, A.; Desmaele, D.; Couvreur, P. Multifunctional squalene-based prodrug nanoparticles for targeted cancer therapy. Chem. Commun. (Camb.) 2014, 50, 5336–5338. [Google Scholar] [CrossRef]
- Kotelevets, L.; Chastre, E.; Caron, J.; Mougin, J.; Bastian, G.; Pineau, A.; Walker, F.; Lehy, T.; Desmaele, D.; Couvreur, P. A Squalene-Based Nanomedicine for Oral Treatment of Colon Cancer. Cancer Res. 2017, 77, 2964–2975. [Google Scholar] [CrossRef] [Green Version]
- Gaudin, A.; Yemisci, M.; Eroglu, H.; Lepetre-Mouelhi, S.; Turkoglu, O.F.; Donmez-Demir, B.; Caban, S.; Sargon, M.F.; Garcia-Argote, S.; Pieters, G.; et al. Squalenoyl adenosine nanoparticles provide neuroprotection after stroke and spinal cord injury. Nat. Nanotechnol. 2014, 9, 1054–1062. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, J.; Zheng, K.; Shen, H.; Chen, X. Long-term ginsenoside Rg1 supplementation improves age-related cognitive decline by promoting synaptic plasticity associated protein expression in C57BL/6J mice. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.F.; Li, Q.; Li, Y. Long-term ginsenoside administration prevents memory loss in aged female C57BL/6J mice by modulating the redox status and up-regulating the plasticity-related proteins in hippocampus. Neuroscience 2011, 183, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhou, L.; Li, W.; Liu, Y.; Wang, J.; He, P. Protective effects of ginsenosides Rg1 and Rb1 on an Alzheimer’s disease mouse model: A metabolomics study. J. Chromatogr. B Anal. Technol. Biomed. Life Sci 2015, 985, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Kaundal, M.; Akhtar, M.; Deshmukh, R. Lupeol Isolated from Betula alnoides Ameliorates Amyloid Beta Induced Neuronal Damage via Targeting Various Pathological Events and Alteration in Neurotransmitter Levels in Rat’s Brain. J. Neurol. Neurosci. 2017, 8, 195. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.C.; Chang, C.H.; Liao, V.H. Anti-Parkinsonian effects of beta-amyrin are regulated via LGG-1 involved autophagy pathway in Caenorhabditis elegans. Phytomedicine 2017, 36, 118–125. [Google Scholar] [CrossRef]
- Farvin, K.H.; Anandan, R.; Kumar, S.H.; Shiny, K.S.; Mathew, S.; Sankar, T.V.; Nair, P.G. Cardioprotective effect of squalene on lipid profile in isoprenaline-induced myocardial infarction in rats. J. Med. Food 2006, 9, 531–536. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, X.; Bi, D.; Wang, X.; Zhang, X.; Dai, H.; Chen, S.; Zhang, W. Influence of squalene feeding on plasma leptin, testosterone & blood pressure in rats. Indian J. Med. Res. 2009, 129, 150–153. [Google Scholar]
- Du, Y.; Ko, K.M. Effects of pharmacological preconditioning by emodin/oleanolic acid treatment and/or ischemic preconditioning on mitochondrial antioxidant components as well as the susceptibility to ischemia-reperfusion injury in rat hearts. Mol. Cell. Biochem. 2006, 288, 135–142. [Google Scholar] [CrossRef]
- Du, Y.; Ko, K.M. Oleanolic acid protects against myocardial ischemia-reperfusion injury by enhancing mitochondrial antioxidant mechanism mediated by glutathione and alpha-tocopherol in rats. Planta Med. 2006, 72, 222–227. [Google Scholar] [CrossRef]
- Ibrahim, N.; Fairus, S.; Zulfarina, M.S.; Naina Mohamed, I. The Efficacy of Squalene in Cardiovascular Disease Risk-A Systematic Review. Nutrients 2020, 12, 414. [Google Scholar] [CrossRef] [Green Version]
- Guan, T.; Qian, Y.; Tang, X.; Huang, M.; Huang, L.; Li, Y.; Sun, H. Maslinic acid, a natural inhibitor of glycogen phosphorylase, reduces cerebral ischemic injury in hyperglycemic rats by GLT-1 up-regulation. J. Neurosci. Res. 2011, 89, 1829–1839. [Google Scholar] [CrossRef]
- Ravi Kumar, S.; Narayan, B.; Sawada, Y.; Hosokawa, M.; Miyashita, K. Combined effect of astaxanthin and squalene on oxidative stress in vivo. Mol. Cell. Biochem. 2016, 417, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Ramu, R.; Shirahatti, P.S.; Nanjunda Swamy, S.; Zameer, F.; Dhananjaya, B.L.; Prasad, N. Assessment of In Vivo Antidiabetic Properties of Umbelliferone and Lupeol Constituents of Banana (Musa sp. var. Nanjangud Rasa Bale) Flower in Hyperglycaemic Rodent Model. PLoS ONE 2016, 11, e0151135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarse, K.; Bossecker, A.; Muller-Kuhrt, L.; Siems, K.; Hernandez, M.A.; Berendsohn, W.G.; Birringer, M.; Ristow, M. The phytochemical glaucarubinone promotes mitochondrial metabolism, reduces body fat, and extends lifespan of Caenorhabditis elegans. Horm. Metab. Res. 2011, 43, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhai, C.; Liu, Q.; Wang, X.; Ren, Z.; Zhang, Y.; Zhang, Y.; Wu, Q.; Sun, S.; Li, S.; et al. Cycloastragenol, a triterpene aglycone derived from Radix astragali, suppresses the accumulation of cytoplasmic lipid droplet in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2014, 450, 306–311. [Google Scholar] [CrossRef]
- Cao, S.; Dong, X.L.; Ho, M.X.; Yu, W.X.; Wong, K.C.; Yao, X.S.; Wong, M.S. Oleanolic Acid Exerts Osteoprotective Effects and Modulates Vitamin D Metabolism. Nutrients 2018, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Pokorny, J.; Borkova, L.; Urban, M. Click Reactions in Chemistry of Triterpenes–Advances Towards Development of Potential Therapeutics. Curr. Med. Chem. 2018, 25, 636–658. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, J.; Liu, T.; Fang, J.; Wan, J.; Zhao, J.; Li, W.; Liu, J.; Zhao, X.; Chen, S. Anti-viral effects of urosolic acid on guinea pig cytomegalovirus in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2012, 32, 883–887. [Google Scholar] [CrossRef]
- Kazakova, O.B.; Giniyatullina, G.V.; Yamansarov, E.Y.; Tolstikov, G.A. Betulin and ursolic acid synthetic derivatives as inhibitors of Papilloma virus. Bioorg. Med. Chem. Lett. 2010, 20, 4088–4090. [Google Scholar] [CrossRef]
- Chang, C.D.; Lin, P.Y.; Hsu, J.L.; Shih, W.L. Ursolic Acid Suppresses Hepatitis B Virus X Protein-mediated Autophagy and Chemotherapeutic Drug Resistance. Anticancer Res. 2016, 36, 5097–5107. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Li, S.; Liao, Q.; Zhang, Y.; Sun, R.; Zhu, X.; Zhang, Q.; Wang, J.; Wu, X.; Fang, X.; et al. Oleanolic acid and ursolic acid: Novel hepatitis C virus antivirals that inhibit NS5B activity. Antivir. Res. 2013, 98, 44–53. [Google Scholar] [CrossRef]
- Jimenez-Arellanes, A.; Luna-Herrera, J.; Cornejo-Garrido, J.; Lopez-Garcia, S.; Castro-Mussot, M.E.; Meckes-Fischer, M.; Mata-Espinosa, D.; Marquina, B.; Torres, J.; Hernandez-Pando, R. Ursolic and oleanolic acids as antimicrobial and immunomodulatory compounds for tuberculosis treatment. BMC Complement. Altern. Med. 2013, 13, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.; Li, Y.; Zhang, L.; Cheng, A.; Liu, Y.; Lou, H. Retigeric acid B enhances the efficacy of azoles combating the virulence and biofilm formation of Candida albicans. Biol. Pharm. Bull. 2012, 35, 1794–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids Functionality, Sources, and Processing by Supercritical Technology: A Review. J. Chem. 2016, 2016, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Alcaino, J.; Baeza, M.; Cifuentes, V. Carotenoid Distribution in Nature. Subcell Biochem. 2016, 79, 3–33. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharm. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- O’Neill, M.E.; Carroll, Y.; Corridan, B.; Olmedilla, B.; Granado, F.; Blanco, I.; Van den Berg, H.; Hininger, I.; Rousell, A.M.; Chopra, M.; et al. A European carotenoid database to assess carotenoid intakes and its use in a five-country comparative study. Br. J. Nutr. 2001, 85, 499–507. [Google Scholar] [CrossRef]
- Perry, A.; Rasmussen, H.; Johnson, E.J. Xanthophyll (lutein, zeaxanthin) content in fruits, vegetables and corn and egg products. J. Food Compos. Anal. 2009, 22, 9–15. [Google Scholar] [CrossRef]
- Lashmanova, E.A.; Kuzivanova, O.A.; Dymova, O.V.; Moskalev, A.A. The Effects of Cloudberry Fruit Extract on Drosophila melanogaster Lifespan and Stress Resistance. Adv. Gerontol. 2019, 9, 254–260. [Google Scholar] [CrossRef]
- Weinrich, T.; Xu, Y.; Wosu, C.; Harvey, P.J.; Jeffery, G. Mitochondrial Function, Mobility and Lifespan Are Improved in Drosophila melanogaster by Extracts of 9-cis-beta-Carotene from Dunaliella salina. Mar. Drugs 2019, 17, 279. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Dai, D.; Li, W. Extension of Life Span and Improvement of Vitality in Drosophila melanogaster by Supplementation with Lycopene Mycelium Powder from Blakeslea Trispora. Food Sci. Technol. Res. 2013, 19, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Lashmanova, E.; Proshkina, E.; Zhikrivetskaya, S.; Shevchenko, O.; Marusich, E.; Leonov, S.; Melerzanov, A.; Zhavoronkov, A.; Moskalev, A. Fucoxanthin increases lifespan of Drosophila melanogaster and Caenorhabditis Elegans. Pharm. Res. 2015, 100, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Massie, H.R.; Ferreira, J.R., Jr.; DeWolfe, L.K. Effect of dietary beta-carotene on the survival of young and old mice. Gerontology 1986, 32, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.; Sultan, L.D.; Saada, A.; Hirschberg, J.; Osterzetser-Biran, O.; Gruenbaum, Y. Astaxanthin extends lifespan via altered biogenesis of the mitochondrial respiratory chain complex III. Biorxiv 2019. [Google Scholar] [CrossRef]
- Nicoll, A.; Locarnini, S. Review: Present and future directions in the treatment of chronic hepatitis B infection. J. Gastroenterol. Hepatol. 1997, 12, 843–854. [Google Scholar] [CrossRef]
- SJ, S.; Veerabhadrappa, B.; Subramaniyan, S.; Dyavaiah, M. Astaxanthin enhances the longevity of Saccharomyces cerevisiae by decreasing oxidative stress and apoptosis. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazaki, K.; Yoshikoshi, C.; Oshiro, S.; Yanase, S. Supplemental cellular protection by a carotenoid extends lifespan via Ins/IGF-1 signaling in Caenorhabditis elegans. Oxid. Med. Cell. Longev. 2011, 2011, 596240. [Google Scholar] [CrossRef] [Green Version]
- Dhinaut, J.; Balourdet, A.; Teixeira, M.; Chogne, M.; Moret, Y. A dietary carotenoid reduces immunopathology and enhances longevity through an immune depressive effect in an insect model. Sci. Rep. 2017, 7, 12429. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Han, S.; Wang, H.; Wang, T. Lutein extends the lifespan of Drosophila melanogaster. Arch. Gerontol. Geriatr. 2014, 58, 153–159. [Google Scholar] [CrossRef]
- Guvatova, Z.; Dalina, A.; Marusich, E.; Pudova, E.; Kobelyatskaya, A.; Krasnov, G.; Kudryavtseva, A.; Leonov, S.; Moskalev, A. Protective effects of carotenoid fucoxanthin in fibroblasts cellular senescence. Mech. Ageing Dev. 2020, 111260. [Google Scholar] [CrossRef]
- Dambroise, E.; Monnier, L.; Ruisheng, L.; Aguilaniu, H.; Joly, J.S.; Tricoire, H.; Rera, M. Two phases of aging separated by the Smurf transition as a public path to death. Sci. Rep. 2016, 6, 23523. [Google Scholar] [CrossRef] [Green Version]
- Gil, P.; Farinas, F.; Casado, A.; Lopez-Fernandez, E. Malondialdehyde: A possible marker of ageing. Gerontology 2002, 48, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Koh, K.; Evans, J.M.; Hendricks, J.C.; Sehgal, A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc. Natl. Acad. Sci. USA 2006, 103, 13843–13847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mander, B.A.; Winer, J.R.; Walker, M.P. Sleep and Human Aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.W.; Ingold, K.U. beta-Carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573. [Google Scholar] [CrossRef]
- Albanes, D.; Heinonen, O.P.; Huttunen, J.K.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Haapakoski, J.; Rautalahti, M.; Hartman, A.M.; Palmgren, J. Effects of alpha-tocopherol and beta-carotene supplements on cancer incidence in the Alpha-Tocopherol Beta-Carotene Cancer Prevention Study. Am. J. Clin. Nutr. 1995, 62, 1427S–1430S. [Google Scholar] [CrossRef]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Crespo, J.; Montero, Z.; Fuentes, J.L.; Reig Garcia-Galbis, M.; Garbayo, I.; Vilchez, C.; Martinez-Espinosa, R.M. Exploring the Valuable Carotenoids for the Large-Scale Production by Marine Microorganisms. Mar. Drugs 2018, 16, 203. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, A. Carotenoids and other pigments as natural colorants. Pure Appl. Chem. 2006, 78, 1477–1491. [Google Scholar] [CrossRef]
- Yeum, K.-J.; Aldini, G.; Russell, R.M.; Krinsky, N.I. Antioxidant/Pro-oxidant Actions of Carotenoids. In Carotenoids: Volume 5: Nutrition and Health; Britton, G., Pfander, H., Liaaen-Jensen, S., Eds.; Birkhäuser Basel: Basel, Switzerland, 2009; pp. 235–268. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress—Implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids activate the antioxidant response element transcription system. Mol. Cancer 2005, 4, 177–186. [Google Scholar]
- Liu, C.L.; Chiu, Y.T.; Hu, M.L. Fucoxanthin enhances HO-1 and NQO1 expression in murine hepatic BNL CL.2 cells through activation of the Nrf2/ARE system partially by its pro-oxidant activity. J. Agric. Food Chem. 2011, 59, 11344–11351. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.K.; Lee, S.J.; Na, H.J.; Ha, K.S.; Han, J.A.; Lee, H.; Kwon, Y.G.; Chung, C.K.; Kim, Y.M. beta-Carotene inhibits inflammatory gene expression in lipopolysaccharide-stimulated macrophages by suppressing redox-based NF-kappaB activation. Exp. Mol. Med. 2005, 37, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Hong, P.; Zheng, X. beta-carotene attenuates lipopolysaccharide-induced inflammation via inhibition of the NF-kappaB, JAK2/STAT3 and JNK/p38 MAPK signaling pathways in macrophages. Anim. Sci. J. 2019, 90, 140–148. [Google Scholar] [CrossRef]
- Liu, C.; Russell, R.M.; Wang, X.D. Low dose beta-carotene supplementation of ferrets attenuates smoke-induced lung phosphorylation of JNK, p38 MAPK, and p53 proteins. J. Nutr. 2004, 134, 2705–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satomi, Y.; Nishino, H. Implication of mitogen-activated protein kinase in the induction of G1 cell cycle arrest and gadd45 expression by the carotenoid fucoxanthin in human cancer cells. Biochim. Biophys. Acta 2009, 1790, 260–266. [Google Scholar] [CrossRef]
- Satomi, Y. Fucoxanthin induces GADD45A expression and G1 arrest with SAPK/JNK activation in LNCap human prostate cancer cells. Anticancer Res. 2012, 32, 807–813. [Google Scholar]
- Plyusnina, E.N.; Shaposhnikov, M.V.; Moskalev, A.A. Increase of Drosophila melanogaster lifespan due to D-GADD45 overexpression in the nervous system. Biogerontology 2011, 12, 211–226. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.R.; Hosokawa, M.; Miyashita, K. Fucoxanthin: A marine carotenoid exerting anti-cancer effects by affecting multiple mechanisms. Mar. Drugs 2013, 11, 5130–5147. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer chemoprevention by carotenoids. Molecules 2012, 17, 3202–3242. [Google Scholar] [CrossRef]
- Pangestuti, R.; Vo, T.S.; Ngo, D.H.; Kim, S.K. Fucoxanthin ameliorates inflammation and oxidative reponses in microglia. J. Agric. Food Chem. 2013, 61, 3876–3883. [Google Scholar] [CrossRef] [PubMed]
- Mounien, L.; Tourniaire, F.; Landrier, J.F. Anti-Obesity Effect of Carotenoids: Direct Impact on Adipose Tissue and Adipose Tissue-Driven Indirect Effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbaraly, N.T.; Faure, H.; Gourlet, V.; Favier, A.; Berr, C. Plasma carotenoid levels and cognitive performance in an elderly population: Results of the EVA Study. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.J.; Vishwanathan, R.; Johnson, M.A.; Hausman, D.B.; Davey, A.; Scott, T.M.; Green, R.C.; Miller, L.S.; Gearing, M.; Woodard, J.; et al. Relationship between Serum and Brain Carotenoids, alpha-Tocopherol, and Retinol Concentrations and Cognitive Performance in the Oldest Old from the Georgia Centenarian Study. J. Aging Res. 2013, 2013, 951786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.J.; McDonald, K.; Caldarella, S.M.; Chung, H.Y.; Troen, A.M.; Snodderly, D.M. Cognitive findings of an exploratory trial of docosahexaenoic acid and lutein supplementation in older women. Nutr. Neurosci. 2008, 11, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Perrig, W.J.; Perrig, P.; Stahelin, H.B. The relation between antioxidants and memory performance in the old and very old. J. Am. Geriatr. Soc. 1997, 45, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Nyssen, O.P.; Syed, A.; Jansen, E.; Bueno-de-Mesquita, B.; Gallo, V. Vitamin A and carotenoids and the risk of Parkinson’s disease: A systematic review and meta-analysis. Neuroepidemiology 2014, 42, 25–38. [Google Scholar] [CrossRef]
- Li, F.J.; Shen, L.; Ji, H.F. Dietary intakes of vitamin E, vitamin C, and beta-carotene and risk of Alzheimer’s disease: A meta-analysis. J. Alzheimers Dis. 2012, 31, 253–258. [Google Scholar] [CrossRef]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T.H. Carotenoids and cardiovascular health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar] [CrossRef]
- Kulczyński, B.; Gramza-Michałowska, A.; Kobus-Cisowska, J.; Kmiecik, D. The role of carotenoids in the prevention and treatment of cardiovascular disease—Current state of knowledge. J. Funct. Foods 2017, 38, 45–65. [Google Scholar] [CrossRef]
- Sluijs, I.; Cadier, E.; Beulens, J.W.; van der, A.D.; Spijkerman, A.M.; van der Schouw, Y.T. Dietary intake of carotenoids and risk of type 2 diabetes. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Montonen, J.; Knekt, P.; Jarvinen, R.; Reunanen, A. Dietary antioxidant intake and risk of type 2 diabetes. Diabetes Care 2004, 27, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Arnlov, J.; Zethelius, B.; Riserus, U.; Basu, S.; Berne, C.; Vessby, B.; Alfthan, G.; Helmersson, J. Serum and dietary beta-carotene and alpha-tocopherol and incidence of type 2 diabetes mellitus in a community-based study of Swedish men: Report from the Uppsala Longitudinal Study of Adult Men (ULSAM) study. Diabetologia 2009, 52, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyne, T.; Ibiebele, T.I.; Baade, P.D.; Dobson, A.; McClintock, C.; Dunn, S.; Leonard, D.; Shaw, J. Diabetes mellitus and serum carotenoids: Findings of a population-based study in Queensland, Australia. Am. J. Clin. Nutr. 2005, 82, 685–693. [Google Scholar] [CrossRef]
- Sugiura, M.; Nakamura, M.; Ogawa, K.; Ikoma, Y.; Yano, M. High-serum carotenoids associated with lower risk for developing type 2 diabetes among Japanese subjects: Mikkabi cohort study. BMJ Open Diabetes Res. Care 2015, 3, e000147. [Google Scholar] [CrossRef] [Green Version]
- Arteni, A.A.; Fradot, M.; Galzerano, D.; Mendes-Pinto, M.M.; Sahel, J.A.; Picaud, S.; Robert, B.; Pascal, A.A. Structure and Conformation of the Carotenoids in Human Retinal Macular Pigment. PLoS ONE 2015, 10, e0135779. [Google Scholar] [CrossRef]
- Buscemi, S.; Corleo, D.; Di Pace, F.; Petroni, M.L.; Satriano, A.; Marchesini, G. The Effect of Lutein on Eye and Extra-Eye Health. Nutrients 2018, 10, 1321. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.J.; Hu, Y.N.; Lin, S.; Ma, W.J.; Li, X.R. Application of Lutein and Zeaxanthin in nonproliferative diabetic retinopathy. Int. J. Ophthalmol. 2011, 4, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Moschos, M.M.; Dettoraki, M.; Tsatsos, M.; Kitsos, G.; Kalogeropoulos, C. Effect of carotenoids dietary supplementation on macular function in diabetic patients. Eye Vis. (Lond.) 2017, 4, 23. [Google Scholar] [CrossRef]
- Yamaguchi, M. Role of carotenoid beta-cryptoxanthin in bone homeostasis. J. Biomed. Sci. 2012, 19, 36. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.; Rao, A.V.; Rao, L.G. Lycopene II—Effect on osteoblasts: The carotenoid lycopene stimulates cell proliferation and alkaline phosphatase activity of SaOS-2 cells. J. Med. Food 2003, 6, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Virdee, S.S.; Thomas, M.B. A practitioner’s guide to gutta-percha removal during endodontic retreatment. Br. Dent. J. 2017, 222, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Belladonna, F.G.; Calasans-Maia, M.D.; Novellino Alves, A.T.; de Brito Resende, R.F.; Souza, E.M.; Silva, E.J.; Fidel, S.R.; De-Deus, G. Biocompatibility of a self-adhesive gutta-percha-based material in subcutaneous tissue of mice. J. Endod. 2014, 40, 1869–1873. [Google Scholar] [CrossRef]
- Marending, M.; Bubenhofer, S.B.; Sener, B.; De-Deus, G. Primary assessment of a self-adhesive gutta-percha material. Int. Endod. J. 2013, 46, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, D.J.; Duncan, H.F. Root canal filling using Resilon: A review. Br. Dent. J. 2011, 211, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Qian, M.C. Development of C13-norisoprenoids, carotenoids and other volatile compounds in Vitis vinifera L. Cv. Pinot noir grapes. Food Chem. 2016, 192, 633–641. [Google Scholar] [CrossRef]
- Crupi, P.; Coletta, A.; Antonacci, D. Analysis of carotenoids in grapes to predict norisoprenoid varietal aroma of wines from Apulia. J. Agric. Food Chem. 2010, 58, 9647–9656. [Google Scholar] [CrossRef]
- Wirth, J.; Guo, W.; Baumes, R.; Gunata, Z. Volatile compounds released by enzymatic hydrolysis of glycoconjugates of leaves and grape berries from Vitis vinifera Muscat of Alexandria and Shiraz cultivars. J. Agric. Food Chem. 2001, 49, 2917–2923. [Google Scholar] [CrossRef]
- Song, J.; Shellie, K.C.; Wang, H.; Qian, M.C. Influence of deficit irrigation and kaolin particle film on grape composition and volatile compounds in Merlot grape (Vitis vinifera L.). Food Chem. 2012, 134, 841–850. [Google Scholar] [CrossRef]
- Mathieu, S.; Terrier, N.; Procureur, J.; Bigey, F.; Gunata, Z. A carotenoid cleavage dioxygenase from Vitis vinifera L.: Functional characterization and expression during grape berry development in relation to C13-norisoprenoid accumulation. J. Exp. Bot. 2005, 56, 2721–2731. [Google Scholar] [CrossRef]
- Ishikura, H.; Mochizuki, T.; Izumi, Y.; Usui, T.; Sawada, H.; Uchino, H. Differentiation of mouse leukemic M1 cells induced by polyprenoids. Leuk Res. 1984, 8, 843–852. [Google Scholar] [CrossRef]
- Sakagami, H.; Hashimoto, K.; Suzuki, F.; Ishihara, M.; Kikuchi, H.; Katayama, T.; Satoh, K. Tumor-specificity and type of cell death induced by vitamin K2 derivatives and prenylalcohols. Anticancer Res. 2008, 28, 151–158. [Google Scholar]
- Hsieh, C.L.; Tseng, M.H.; Shao, Y.Y.; Chang, J.Y.; Kuo, C.C.; Chang, C.Y.; Kuo, Y.H. C35 terpenoids from the bark of Calocedrus macrolepis var. formosana with activity against human cancer cell lines. J. Nat. Prod. 2006, 69, 1611–1613. [Google Scholar] [CrossRef] [PubMed]
- Zwenger, S.; Basu, C. Plant terpenoids: Applications and future potentials. Biotechnol. Mol. Biol. Rev. 2008, 3, 1. [Google Scholar]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and Medicinal Uses of Terpenes. Med. Plants 2019. [Google Scholar] [CrossRef]
- Dönertaş, H.M.; Fuentealba, M.; Partridge, L.; Thornton, J.M. Identifying Potential Ageing-Modulating Drugs In Silico. Trends Endocrinol. Metab. 2019, 30, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Cruz, G.S.; Wanderley-Teixeira, V.; Oliveira, J.V.; CG, D.A.; Cunha, F.M.; Teixeira, A.A.C.; Guedes, C.A.; Dutra, K.A.; Barbosa, D.R.S.; Breda, M.O. Effect of trans-anethole, limonene and your combination in nutritional components and their reflection on reproductive parameters and testicular apoptosis in Spodoptera frugiperda (Lepidoptera: Noctuidae). Chem. Biol. Interact. 2017, 263, 74–80. [Google Scholar] [CrossRef]
- Gray, C.A.; Runyon, J.B.; Jenkins, M.J.; Giunta, A.D. Mountain Pine Beetles Use Volatile Cues to Locate Host Limber Pine and Avoid Non-Host Great Basin Bristlecone Pine. PLoS ONE 2015, 10, e0135752. [Google Scholar] [CrossRef]
- Tang, X.P.; Guo, X.H.; Geng, D.; Weng, L.J. d-Limonene protects PC12 cells against corticosterone-induced neurotoxicity by activating the AMPK pathway. Environ. Toxicol. Pharm. 2019, 70, 103192. [Google Scholar] [CrossRef]
- Miller, J.A.; Lang, J.E.; Ley, M.; Nagle, R.; Hsu, C.H.; Thompson, P.A.; Cordova, C.; Waer, A.; Chow, H.H. Human breast tissue disposition and bioactivity of limonene in women with early-stage breast cancer. Cancer Prev. Res. 2013, 6, 577–584. [Google Scholar] [CrossRef] [Green Version]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Oday, O.H.; Lateef, A.; Hassan, S.K.; Rashid, S.; Ali, N.; Zeeshan, M.; et al. D-limonene suppresses doxorubicin-induced oxidative stress and inflammation via repression of COX-2, iNOS, and NFkappaB in kidneys of Wistar rats. Exp. Biol. Med. (Maywood) 2014, 239, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, L.P.; Daphney, C.M.; Oppong-Damoah, A.; Uchakin, P.N.; Abney, S.E.; Uchakina, O.N.; Khusial, R.D.; Akil, A.; Murnane, K.S. The cannabinoid receptor 2 agonist, beta-caryophyllene, improves working memory and reduces circulating levels of specific proinflammatory cytokines in aged male mice. Behav. Brain Res. 2019, 372, 112012. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Hurtado, P.; Gonzalez-Castaneda, R.E.; Beas-Zarate, C.; Flores-Soto, M.E.; Viveros-Paredes, J.M. beta-Caryophyllene Reduces DNA Oxidation and the Overexpression of Glial Fibrillary Acidic Protein in the Prefrontal Cortex and Hippocampus of d-Galactose-Induced Aged BALB/c Mice. J. Med. Food 2020, 23, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Pant, A.; Trivedi, S.; Pandey, R. Curcumin and beta-caryophellene attenuate cadmium quantum dots induced oxidative stress and lethality in Caenorhabditis elegans model system. Environ. Toxicol. Pharm. 2016, 42, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, D.; Chen, Y.; Yang, M. beta-Caryophyllene inhibits high glucose-induced oxidative stress, inflammation and extracellular matrix accumulation in mesangial cells. Int. Immunopharmacol. 2020, 84, 106556. [Google Scholar] [CrossRef]
- Younis, N.S.; Mohamed, M.E. beta-Caryophyllene as a Potential Protective Agent Against Myocardial Injury: The Role of Toll-Like Receptors. Molecules 2019, 24, 1929. [Google Scholar] [CrossRef] [Green Version]
- Youssef, D.A.; El-Fayoumi, H.M.; Mahmoud, M.F. Beta-caryophyllene alleviates diet-induced neurobehavioral changes in rats: The role of CB2 and PPAR-gamma receptors. Biomed. Pharm. 2019, 110, 145–154. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strzadala, L.; Szumny, A. beta-caryophyllene and beta-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Park, N.H.; Kang, Y.G.; Kim, S.H.; Bae, I.H.; Lee, S.H.; Kim, D.Y.; Hwang, J.S.; Kim, Y.J.; Lee, T.R.; Lee, E.S. Dehydroabietic Acid Induces Regeneration of Collagen Fibers in Ultraviolet B-Irradiated Human Dermal Fibroblasts and Skin Equivalents. Ski. Pharmacol. Physiol. 2019, 32, 109–116. [Google Scholar] [CrossRef]
- Kamaya, Y.; Tokita, N.; Suzuki, K. Effects of dehydroabietic acid and abietic acid on survival, reproduction, and growth of the crustacean Daphnia magna. Ecotoxicol. Environ. Saf. 2005, 61, 83–88. [Google Scholar] [CrossRef]
- Xing, L.; Gutierrez-Villagomez, J.M.; Da Fonte, D.F.; Venables, M.J.; Trudeau, V.L. Dehydroabietic acid cytotoxicity in goldfish radial glial cells In Vitro. Aquat. Toxicol. 2016, 180, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Rojo, A.I.; Salinas, M.; Diaz, R.; Gallardo, G.; Alam, J.; De Galarreta, C.M.; Cuadrado, A. Regulation of heme oxygenase-1 expression through the phosphatidylinositol 3-kinase/Akt pathway and the Nrf2 transcription factor in response to the antioxidant phytochemical carnosol. J. Biol. Chem. 2004, 279, 8919–8929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, T.; McKercher, S.R.; Lipton, S.A. Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic. Biol. Med. 2013, 65, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.; Xu, G.; Zhan, X.; Gao, Y.; Wang, Z.; Fu, S.; Qin, N.; Hou, X.; Ai, Y.; Wang, C.; et al. Carnosol inhibits inflammasome activation by directly targeting HSP90 to treat inflammasome-mediated diseases. Cell Death Dis. 2020, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Azimi-Nezhad, M.; Borji, A.; Samini, M.; Farkhondeh, T. Protective effects of carnosol against oxidative stress induced brain damage by chronic stress in rats. BMC Complement. Altern. Med. 2017, 17, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, K.C.; Chuang, J.J.; Hsieh, C.W.; Wung, B.S.; Huang, G.D.; Jian, T.Y.; Sun, Y.W. Dual mechanisms of NF-kappaB inhibition in carnosol-treated endothelial cells. Toxicol. Appl. Pharm. 2010, 245, 21–35. [Google Scholar] [CrossRef]
- Satoh, T.; Lipton, S. Recent advances in understanding NRF2 as a druggable target: Development of pro-electrophilic and non-covalent NRF2 activators to overcome systemic side effects of electrophilic drugs like dimethyl fumarate. F1000Research 2017, 6, 2138. [Google Scholar] [CrossRef] [Green Version]
- Lipton, S.A.; Rezaie, T.; Nutter, A.; Lopez, K.M.; Parker, J.; Kosaka, K.; Satoh, T.; McKercher, S.R.; Masliah, E.; Nakanishi, N. Therapeutic advantage of pro-electrophilic drugs to activate the Nrf2/ARE pathway in Alzheimer’s disease models. Cell Death Dis. 2016, 7, e2499. [Google Scholar] [CrossRef]
- Farr, S.A.; Niehoff, M.L.; Ceddia, M.A.; Herrlinger, K.A.; Lewis, B.J.; Feng, S.; Welleford, A.; Butterfield, D.A.; Morley, J.E. Effect of botanical extracts containing carnosic acid or rosmarinic acid on learning and memory in SAMP8 mice. Physiol. Behav. 2016, 165, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Taram, F.; Ignowski, E.; Duval, N.; Linseman, D.A. Neuroprotection Comparison of Rosmarinic Acid and Carnosic Acid in Primary Cultures of Cerebellar Granule Neurons. Molecules 2018, 23, 2956. [Google Scholar] [CrossRef] [Green Version]
- Ishitobi, H.; Sanada, Y.; Kato, Y.; Ikuta, Y.; Shibata, S.; Yamasaki, S.; Lotz, M.K.; Matsubara, K.; Miyaki, S.; Adachi, N. Carnosic acid attenuates cartilage degeneration through induction of heme oxygenase-1 in human articular chondrocytes. Eur. J. Pharm. 2018, 830, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Hryb, A.B.; Pazini, F.L.; Kaster, M.P.; Rodrigues, A.L.S. Therapeutic Potential of Ursolic Acid to Manage Neurodegenerative and Psychiatric Diseases. CNS Drugs 2017, 31, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.X.; Wink, M. Evidence for Anti-Inflammatory Activity of Isoliquiritigenin, 18beta Glycyrrhetinic Acid, Ursolic Acid, and the Traditional Chinese Medicine Plants Glycyrrhiza glabra and Eriobotrya japonica, at the Molecular Level. Medicine 2019, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Gharibi, S.; Bakhtiari, N.; Elham Moslemee, J.; Bakhtiari, F. Ursolic Acid Mediates Hepatic Protection through Enhancing of anti-aging Biomarkers. Curr. Aging Sci. 2018, 11, 16–23. [Google Scholar] [CrossRef]
- Geerlofs, L.; He, Z.; Xiao, S.; Xiao, Z.C. Repeated dose (90 days) oral toxicity study of ursolic acid in Han-Wistar rats. Toxicol. Rep. 2020, 7, 610–623. [Google Scholar] [CrossRef]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Shanmugam, M.K.; Kumar, A.P.; Yap, C.T.; Sethi, G.; Bishayee, A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer 2019, 125, 1228–1246. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.F.; Wan, X.L.; Xu, Y.; Liu, J. Repeated oral administration of oleanolic acid produces cholestatic liver injury in mice. Molecules 2013, 18, 3060–3071. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Liu, J.; Song, L.; Liu, Z.; Han, G.; Yuan, D.; Wang, T.; Dun, Y.; Zhou, Z.; Liu, Z.; et al. Oleanolic acid rejuvenates testicular function through attenuating germ cell DNA damage and apoptosis via deactivation of NF-kappaB, p53 and p38 signalling pathways. J. Pharm. Pharm. 2017, 69, 295–304. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, R. Oleanolic acid and related triterpenoids from olives on vascular function: Molecular mechanisms and therapeutic perspectives. Curr. Med. Chem. 2015, 22, 1414–1425. [Google Scholar] [CrossRef]
- Fernandez-Aparicio, A.; Schmidt-RioValle, J.; Perona, J.S.; Correa-Rodriguez, M.; Castellano, J.M.; Gonzalez-Jimenez, E. Potential Protective Effect of Oleanolic Acid on the Components of Metabolic Syndrome: A Systematic Review. J. Clin. Med. 2019, 8, 1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayeleso, T.B.; Matumba, M.G.; Mukwevho, E. Oleanolic Acid and Its Derivatives: Biological Activities and Therapeutic Potential in Chronic Diseases. Molecules 2017, 22, 1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Gonzalez, M.; Lozano-Mena, G.; Juan, M.E.; Garcia-Granados, A.; Planas, J.M. Assessment of the safety of maslinic acid, a bioactive compound from Olea europaea L. Mol. Nutr. Food Res. 2013, 57, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Qiu, L.; Wang, F. 18alpha-Glycyrrhetinic acid (GA) ameliorates fructose-induced nephropathy in mice by suppressing oxidative stress, dyslipidemia and inflammation. Biomed. Pharm. 2020, 125, 109702. [Google Scholar] [CrossRef] [PubMed]
- Lefaki, M.; Papaevgeniou, N.; Tur, J.A.; Vorgias, C.E.; Sykiotis, G.P.; Chondrogianni, N. The dietary triterpenoid 18alpha-Glycyrrhetinic acid protects from MMC-induced genotoxicity through the ERK/Nrf2 pathway. Redox Biol. 2020, 28, 101317. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, Y.; Mizoguchi, K. Neuropharmacological efficacy of the traditional Japanese Kampo medicine yokukansan and its active ingredients. Pharmacol. Ther. 2016, 166, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.Y.; Wang, W.J.; Dou, J.H.; Gong, L.K. Research progress on the protective effects of licorice-derived 18beta-glycyrrhetinic acid against liver injury. Acta Pharm. Sin. 2020. [Google Scholar] [CrossRef]
- Kowalska, A.; Kalinowska-Lis, U. 18beta-Glycyrrhetinic acid: Its core biological properties and dermatological applications. Int. J. Cosmet. Sci. 2019, 41, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.Z.; Chen, H.M.; Yu, X.T.; Zhang, X.; Feng, X.X.; Kang, X.H.; Li, W.J.; Huang, N.; Luo, H.; Su, Z.R. The protective effect of 18beta-Glycyrrhetinic acid against UV irradiation induced photoaging in mice. Exp. Gerontol. 2015, 61, 147–155. [Google Scholar] [CrossRef]
- Li, X.; Sun, R.; Liu, R. Natural products in licorice for the therapy of liver diseases: Progress and future opportunities. Pharm. Res. 2019, 144, 210–226. [Google Scholar] [CrossRef]
- Al-Saeedi, F.J. Study of the cytotoxicity of asiaticoside on rats and tumour cells. BMC Cancer 2014, 14, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Yan Ni, H.; Si Wei, C.; Wen Juan, W.; Xu, N.; Cui, S.; Liu, X.H.; Zhang, H.; Yue Nan, L.; Liu, S.; et al. Antidepressant-like effect of asiaticoside in mice. Pharm. Biochem. Behav. 2008, 89, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.W.; Wang, W.J.; Li, W.J.; Wang, R.; Li, Y.L.; Huang, Y.N.; Liang, X. Anxiolytic-like effect of asiaticoside in mice. Pharm. Biochem. Behav. 2006, 85, 339–344. [Google Scholar] [CrossRef]
- Namviriyachote, N.; Lipipun, V.; Akkhawattanangkul, Y.; Charoonrut, P.; Ritthidej, G.C. Development of polyurethane foam dressing containing silver and asiaticoside for healing of dermal wound. Asian J. Pharm. Sci. 2019, 14, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Yu, L.; Zhang, X.; Wu, Q.; Wang, D.; Wang, X.; Xia, C.; Feng, H. Asiaticoside attenuates lipopolysaccharide-induced acute lung injury via down-regulation of NF-kappaB signaling pathway. Int. Immunopharmacol. 2015, 26, 181–187. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, S.; Zhang, Z.; Xu, H.; Zhang, W.; Xu, D.; Lin, B.; Mei, Y. Asiaticoside Alleviates Cerebral Ischemia-Reperfusion Injury via NOD2/Mitogen-Activated Protein Kinase (MAPK)/Nuclear Factor kappa B (NF-kappaB) Signaling Pathway. Med. Sci. Monit. 2020, 26, e920325. [Google Scholar] [CrossRef]
- Luo, Y.; Fu, C.; Wang, Z.; Zhang, Z.; Wang, H.; Liu, Y. Asiaticoside attenuates the effects of spinal cord injury through antioxidant and antiinflammatory effects, and inhibition of the p38MAPK mechanism. Mol. Med. Rep. 2015, 12, 8294–8300. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Yu, H.; Chen, S.; Ma, C.; Ma, X.; Xu, L.; Ma, Z.; Qu, R.; Ma, S. Asiaticoside attenuates diabetes-induced cognition deficits by regulating PI3K/Akt/NF-kappaB pathway. Behav. Brain Res. 2015, 292, 288–299. [Google Scholar] [CrossRef]
- Razali, N.N.M.; Ng, C.T.; Fong, L.Y. Cardiovascular Protective Effects of Centella asiatica and Its Triterpenes: A Review. Planta Med. 2019, 85, 1203–1215. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Xia, H.; Guo, Y.; Qian, X.; Zou, X.; Yang, H.; Yin, M.; Liu, H. Ginsenoside Rb1 retards aging process by regulating cell cycle, apoptotic pathway and metabolism of aging mice. J. Ethnopharmacol. 2020, 255, 112746. [Google Scholar] [CrossRef]
- Qi, R.; Jiang, R.; Xiao, H.; Wang, Z.; He, S.; Wang, L.; Wang, Y. Ginsenoside Rg1 protects against d-galactose induced fatty liver disease in a mouse model via FOXO1 transcriptional factor. Life Sci. 2020, 254, 117776. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Z.; Meng, F.J.; Cai, H.Q.; Diao, X.B.; Zhang, B.; Bai, X.P. Ginsenoside Rg3 protects heart against isoproterenol-induced myocardial infarction by activating AMPK mediated autophagy. Cardiovasc. Diagn 2020, 10, 153–160. [Google Scholar] [CrossRef]
- Huang, Q.; Lou, T.; Wang, M.; Xue, L.; Lu, J.; Zhang, H.; Zhang, Z.; Wang, H.; Jing, C.; Zhao, D.; et al. Compound K inhibits autophagy-mediated apoptosis induced by oxygen and glucose deprivation/reperfusion via regulating AMPK-mTOR pathway in neurons. Life Sci. 2020, 254, 117793. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Hua, K.F.; Hsu, W.H.; Suzuki, Y.; Chu, L.J.; Lee, Y.C.; Takahata, A.; Lee, S.L.; Wu, C.C.; Nikolic-Paterson, D.J.; et al. IgA Nephropathy Benefits from Compound K Treatment by Inhibiting NF-kappaB/NLRP3 Inflammasome and Enhancing Autophagy and SIRT1. J. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; She, C.; Li, Q.; Zhang, D.; Xu, J.X.; Li, M.H.; Li, P.; Xu, H.B. Ginsenoside Rh3 activates Nrf2 signaling and protects endometrial cells from oxygen and glucose deprivation-reoxygenation. Aging (Albany Ny) 2020, 12, 6109–6119. [Google Scholar] [CrossRef] [PubMed]
- Go, G.Y.; Jo, A.; Seo, D.W.; Kim, W.Y.; Kim, Y.K.; So, E.Y.; Chen, Q.; Kang, J.S.; Bae, G.U.; Lee, S.J. Ginsenoside Rb1 and Rb2 upregulate Akt/mTOR signaling-mediated muscular hypertrophy and myoblast differentiation. J. Ginseng Res. 2020, 44, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.M.; Yao, Q.; Chen, C. Ginseng compounds: An update on their molecular mechanisms and medical applications. Curr. Vasc. Pharm. 2009, 7, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Hong, Y.; Tran, Q.; Cho, H.; Kim, M.; Kim, C.; Kwon, S.H.; Park, S.; Park, J.; Park, J. A new role for the ginsenoside RG3 in antiaging via mitochondria function in ultraviolet-irradiated human dermal fibroblasts. J. Ginseng Res. 2019, 43, 431–441. [Google Scholar] [CrossRef]
- Kim, D.H.; Park, C.H.; Park, D.; Choi, Y.J.; Park, M.H.; Chung, K.W.; Kim, S.R.; Lee, J.S.; Chung, H.Y. Ginsenoside Rc modulates Akt/FoxO1 pathways and suppresses oxidative stress. Arch. Pharm Res. 2014, 37, 813–820. [Google Scholar] [CrossRef]
- Sandner, G.; Mueller, A.S.; Zhou, X.; Stadlbauer, V.; Schwarzinger, B.; Schwarzinger, C.; Wenzel, U.; Maenner, K.; van der Klis, J.D.; Hirtenlehner, S.; et al. Ginseng Extract Ameliorates the Negative Physiological Effects of Heat Stress by Supporting Heat Shock Response and Improving Intestinal Barrier Integrity: Evidence from Studies with Heat-Stressed Caco-2 Cells, C. elegans and Growing Broilers. Molecules 2020, 25, 835. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.Y.; Kim, Y.G.; Sin, S.J.; Koo, H.; Cheon, K.; Kim, D. Preventive Effect of Mebicar and Ginsenoside Rg1 on Neurobehavioral and Immunological Disruptions Caused by Intermittent Unpredictable Stress in Mice. Neuroimmunomodulation 2018, 25, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.P.; Behm, B. Ginseng: A Qualitative Review of Benefits for Palliative Clinicians. Am. J. Hosp. Palliat. Care 2019, 36, 630–659. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.J.; Robien, K.; Zota, A.R. Serum Folate, Vitamin B-12, Vitamin A, gamma-Tocopherol, alpha-Tocopherol, and Carotenoids Do Not Modify Associations between Cadmium Exposure and Leukocyte Telomere Length in the General US Adult Population. J. Nutr. 2017, 147, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Boccardi, V.; Arosio, B.; Cari, L.; Bastiani, P.; Scamosci, M.; Casati, M.; Ferri, E.; Bertagnoli, L.; Ciccone, S.; Rossi, P.D.; et al. Beta-carotene, telomerase activity and Alzheimer’s disease in old age subjects. Eur J. Nutr. 2020, 59, 119–126. [Google Scholar] [CrossRef]
- Cao, Y.; Wittert, G.; Taylor, A.W.; Adams, R.; Appleton, S.; Shi, Z. Nutrient patterns and chronic inflammation in a cohort of community dwelling middle-aged men. Clin. Nutr. 2017, 36, 1040–1047. [Google Scholar] [CrossRef]
- Dolara, P.; Bigagli, E.; Collins, A. Antioxidant vitamins and mineral supplementation, life span expansion and cancer incidence: A critical commentary. Eur. J. Nutr. 2012, 51, 769–781. [Google Scholar] [CrossRef]
- Min, K.B.; Min, J.Y. Association between leukocyte telomere length and serum carotenoid in US adults. Eur. J. Nutr. 2017, 56, 1045–1052. [Google Scholar] [CrossRef]
- Yuan, C.; Fondell, E.; Ascherio, A.; Okereke, O.I.; Grodstein, F.; Hofman, A.; Willett, W.C. Long-Term Intake of Dietary Carotenoids Is Positively Associated with Late-Life Subjective Cognitive Function in a Prospective Study in US Women. J. Nutr. 2020. [Google Scholar] [CrossRef]
- Mazidi, M.; Katsiki, N.; George, E.S.; Banach, M. Tomato and Lycopene Consumption Is Inversely Associated with Total and Cause-Specific Mortality: A Population-based Cohort Study, on behalf of the International Lipid Expert Panel (ILEP). Br. J. Nutr. 2019. [Google Scholar] [CrossRef] [Green Version]
- Nouri, M.; Nasr-Esfahani, M.H.; Tarrahi, M.J.; Amani, R. The Effect of Lycopene Supplementation on Mood Status and Quality of Life in Infertile Men: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Int. J. Fertil Steril 2020, 14, 17–22. [Google Scholar] [CrossRef]
- Zhai, L.L.; Tang, Z.G. Lycopene improves sperm quality: A promising nutrient for the treatment of male infertility. Phytother. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Huang, C.; Chen, Z. A review for the pharmacological effect of lycopene in central nervous system disorders. Biomed. Pharm. 2019, 111, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, Y.; Wang, Q. Lycopene can reduce prostate-specific antigen velocity in a phase II clinical study in Chinese population. Chin. Med. J. (Engl.) 2014, 127, 2143–2146. [Google Scholar] [PubMed]
- Jiang, X.; Wang, G.; Lin, Q.; Tang, Z.; Yan, Q.; Yu, X. Fucoxanthin prevents lipopolysaccharide-induced depressive-like behavior in mice via AMPK- NF-kappaB pathway. Metab. Brain Dis. 2019, 34, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Mathison, B.D.; Hayek, M.G.; Zhang, J.; Reinhart, G.A.; Chew, B.P. Astaxanthin modulates age-associated mitochondrial dysfunction in healthy dogs. J. Anim. Sci. 2013, 91, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Kawamura, M.; Kawashima, Y.; Uemura, T.; Maoka, T. Effect of astaxanthin-rich extract derived from Paracoccus carotinifaciens on the status of stress and sleep in adults. J. Clin. Biochem. Nutr. 2020, 66, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Yan, Q.; Liu, F.; Jing, C.; Ding, L.; Zhang, L.; Pang, C. Chronic trans-astaxanthin treatment exerts antihyperalgesic effect and corrects co-morbid depressive like behaviors in mice with chronic pain. Neurosci. Lett. 2018, 662, 36–43. [Google Scholar] [CrossRef]
- Comhaire, F.H.; El Garem, Y.; Mahmoud, A.; Eertmans, F.; Schoonjans, F. Combined conventional/antioxidant “Astaxanthin” treatment for male infertility: A double blind, randomized trial. Asian J. 2005, 7, 257–262. [Google Scholar] [CrossRef]
- Nishida, Y.; Nawaz, A.; Kado, T.; Takikawa, A.; Igarashi, Y.; Onogi, Y.; Wada, T.; Sasaoka, T.; Yamamoto, S.; Sasahara, M.; et al. Astaxanthin stimulates mitochondrial biogenesis in insulin resistant muscle via activation of AMPK pathway. J. Cachexia Sarcopenia Muscle 2020, 11, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Zeni, A.L.B.; Camargo, A.; Dalmagro, A.P. Lutein prevents corticosterone-induced depressive-like behavior in mice with the involvement of antioxidant and neuroprotective activities. Pharm. Biochem. Behav. 2019, 179, 63–72. [Google Scholar] [CrossRef]
- Cooke, M.C.; Coates, A.M.; Buckley, E.S.; Buckley, J.D. Lutein Intake and Blood Lutein Concentration Are Positively Associated with Physical Activity in Adults: A Systematic Review. Nutrients 2018, 10, 1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindbergh, C.A.; Lv, J.; Zhao, Y.; Mewborn, C.M.; Puente, A.N.; Terry, D.P.; Renzi-Hammond, L.M.; Hammond, B.R.; Liu, T.; Miller, L.S. The effects of lutein and zeaxanthin on resting state functional connectivity in older Caucasian adults: A randomized controlled trial. Brain Imaging Behav. 2020, 14, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Gerger, P.; Pai, R.K.; Stuckenschneider, T.; Falkenreck, J.; Weigert, H.; Stahl, W.; Weber, B.; Nelles, G.; Spazzafumo, L.; Schneider, S.; et al. Associations of Lipophilic Micronutrients with Physical and Cognitive Fitness in Persons with Mild Cognitive Impairment. Nutrients 2019, 11, 902. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.P.; Sun, L.; Yu, H.S.; Liang, L.P.; Li, W.; Ding, H.; Song, X.B.; Zhang, L.J. The Pharmacological Effects of Lutein and Zeaxanthin on Visual Disorders and Cognition Diseases. Molecules 2017, 22, 610. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.A. Zeaxanthin: Review of Toxicological Data and Acceptable Daily Intake. J. Ophthalmol. 2016, 2016, 3690140. [Google Scholar] [CrossRef] [Green Version]
- Unno, K.; Sugiura, M.; Ogawa, K.; Takabayashi, F.; Toda, M.; Sakuma, M.; Maeda, K.; Fujitani, K.; Miyazaki, H.; Yamamoto, H.; et al. Beta-cryptoxanthin, plentiful in Japanese mandarin orange, prevents age-related cognitive dysfunction and oxidative damage in senescence-accelerated mouse brain. Biol. Pharm. Bull. 2011, 34, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Pearce, K.; Tremellen, K. Influence of nutrition on the decline of ovarian reserve and subsequent onset of natural menopause. Hum. Fertil (Camb.) 2016, 19, 173–179. [Google Scholar] [CrossRef]
- Pearce, K.; Tremellen, K. Influence of Betaeta-Cryptoxanthin Supplementation on Ovarian Reserve and Fertility Status in Aged Wistar Rats. J. Diet. Suppl. 2020, 17, 273–285. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Akdemir, F.; Tuzcu, M.; Sahin, N.; Yilmaz, I.; Juturu, V. beta-Cryptoxanthin ameliorates metabolic risk factors by regulating NF-kappaB and Nrf2 pathways in insulin resistance induced by high-fat diet in rodents. Food Chem. Toxicol. 2017, 107, 270–279. [Google Scholar] [CrossRef]
- Zhang, F.; Shi, D.; Wang, X.; Zhang, Y.; Duan, W.; Li, Y. beta-cryptoxanthin alleviates myocardial ischaemia/reperfusion injury by inhibiting NF-kappaB-mediated inflammatory signalling in rats. Arch. Physiol. Biochem. 2020, 1–8. [Google Scholar] [CrossRef]
- Mukherjee, S.; Date, A.; Patravale, V.; Korting, H.C.; Roeder, A.; Weindl, G. Retinoids in the treatment of skin aging: An overview of clinical efficacy and safety. Clin. Interv. Aging 2006, 1, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.S.; Lim, Y.R.; Lee, K.; Lee, J.; Lee, J.H.; Lee, I.S. Terpenes from Forests and Human Health. Toxicol. Res. 2017, 33, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Partridge, L.; Fuentealba, M.; Kennedy, B.K. The quest to slow ageing through drug discovery. Nat. Rev. Drug Discov. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sander, M.; Oxlund, B.; Jespersen, A.; Krasnik, A.; Mortensen, E.L.; Westendorp, R.G.J.; Rasmussen, L.J. The challenges of human population ageing. Age Ageing 2015, 44, 185–187. [Google Scholar] [CrossRef] [Green Version]
- Partridge, L.; Deelen, J.; Slagboom, P.E. Facing up to the global challenges of ageing. Nature 2018, 561, 45–56. [Google Scholar] [CrossRef]
- Kontis, V.; Bennett, J.E.; Mathers, C.D.; Li, G.; Foreman, K.; Ezzati, M. Future life expectancy in 35 industrialised countries: Projections with a Bayesian model ensemble. Lancet 2017, 389, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Crimmins, E.M. Lifespan and Healthspan: Past, Present, and Promise. Gerontologist 2015, 55, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Ogura, S.; Jakovljevic, M.M. Editorial: Global Population Aging–Health Care, Social and Economic Consequences. Front. Public Health 2018, 6, 335. [Google Scholar] [CrossRef]
- Christensen, K.; Doblhammer, G.; Rau, R.; Vaupel, J.W. Ageing populations: The challenges ahead. Lancet 2009, 374, 1196–1208. [Google Scholar] [CrossRef] [Green Version]






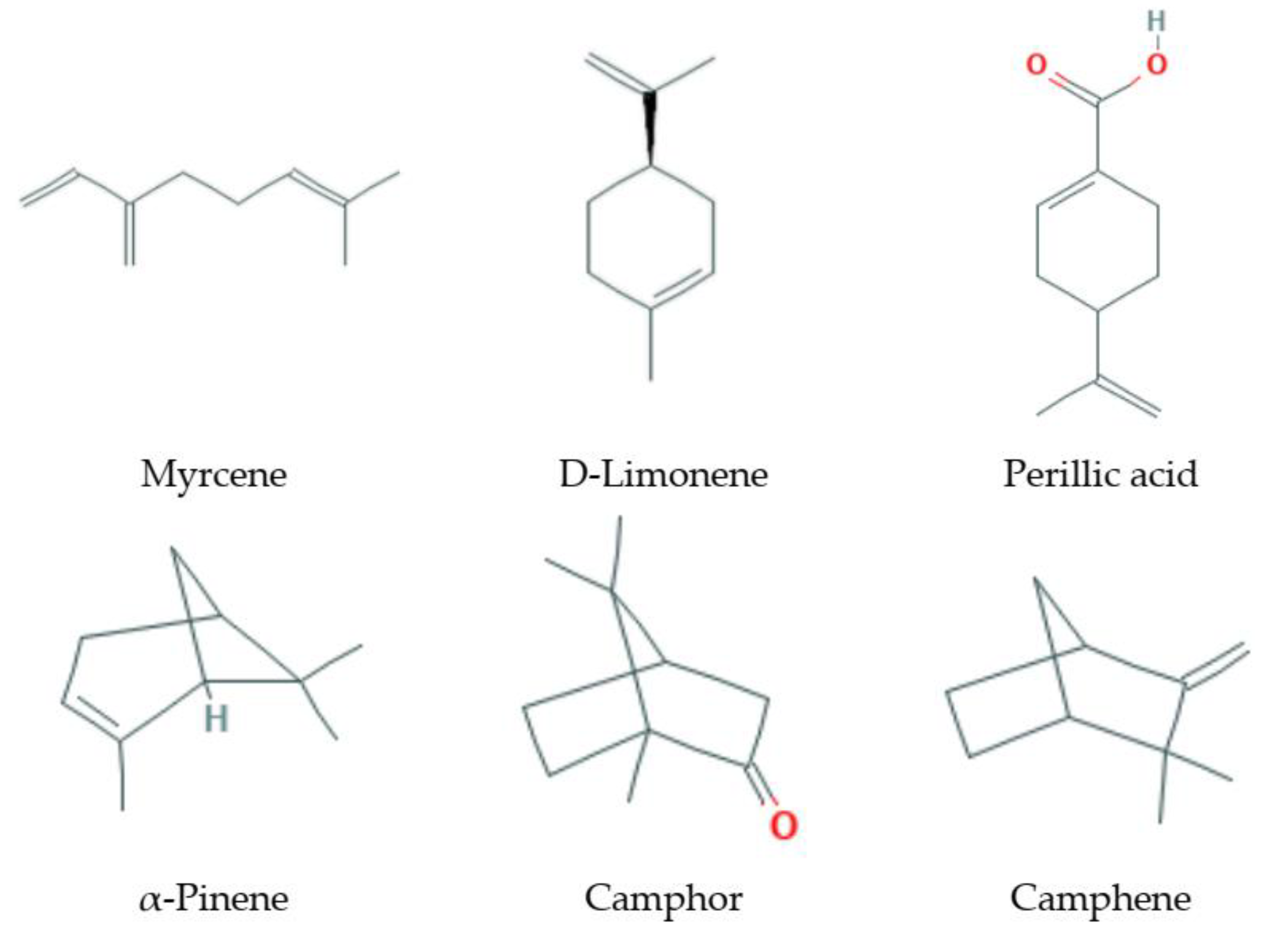{kind=link}
{kind=link}
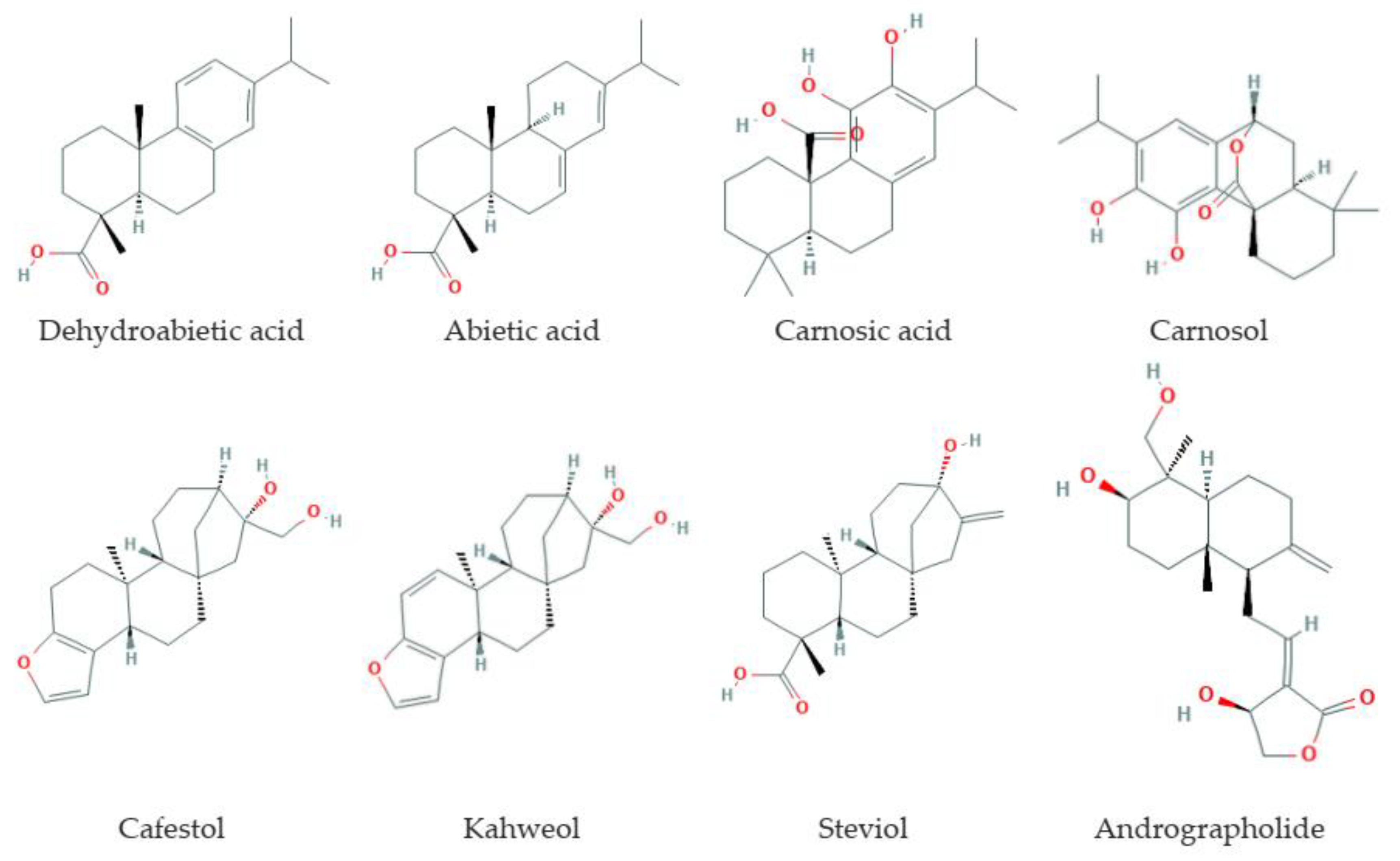{kind=link}
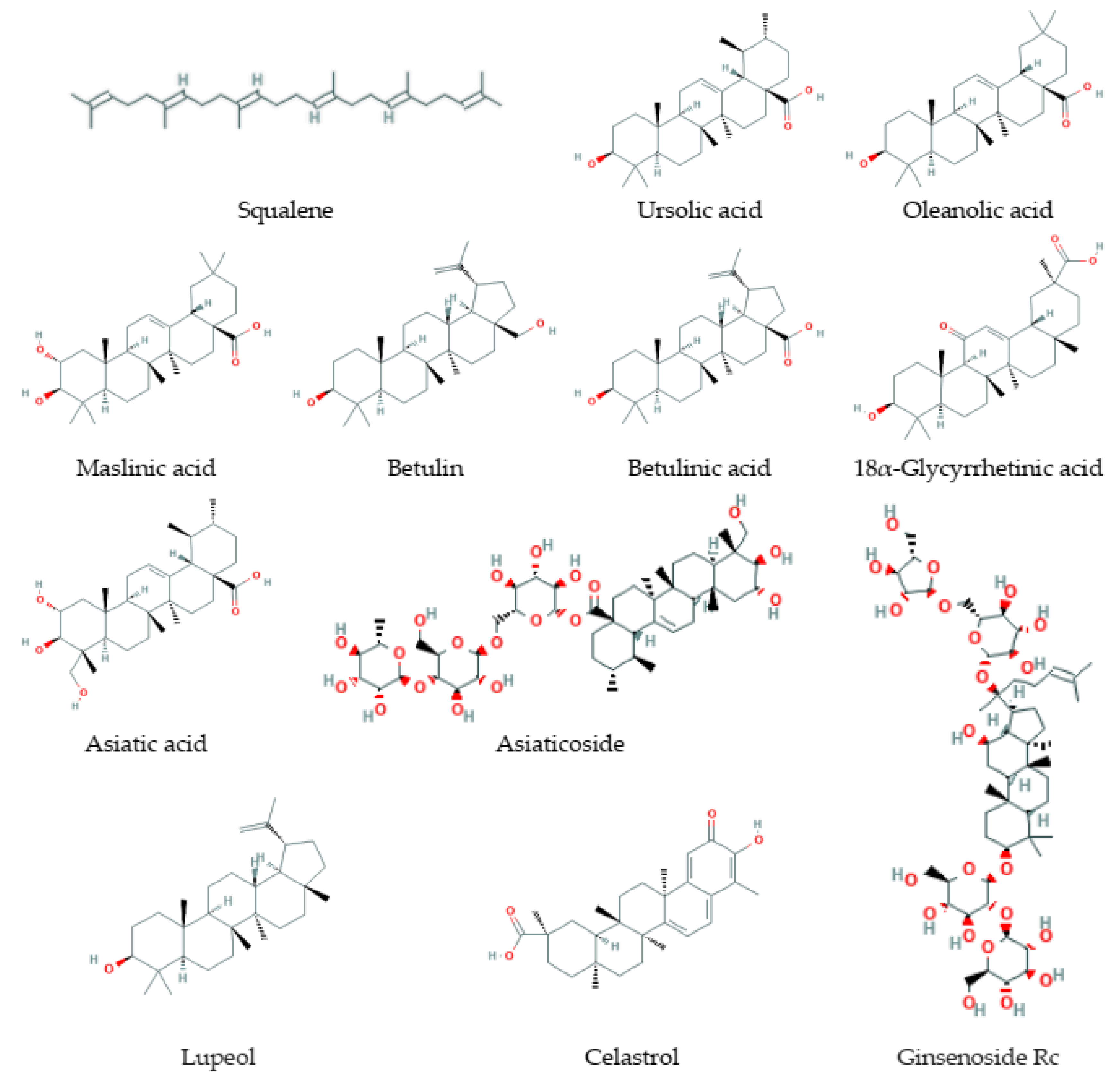{kind=link}
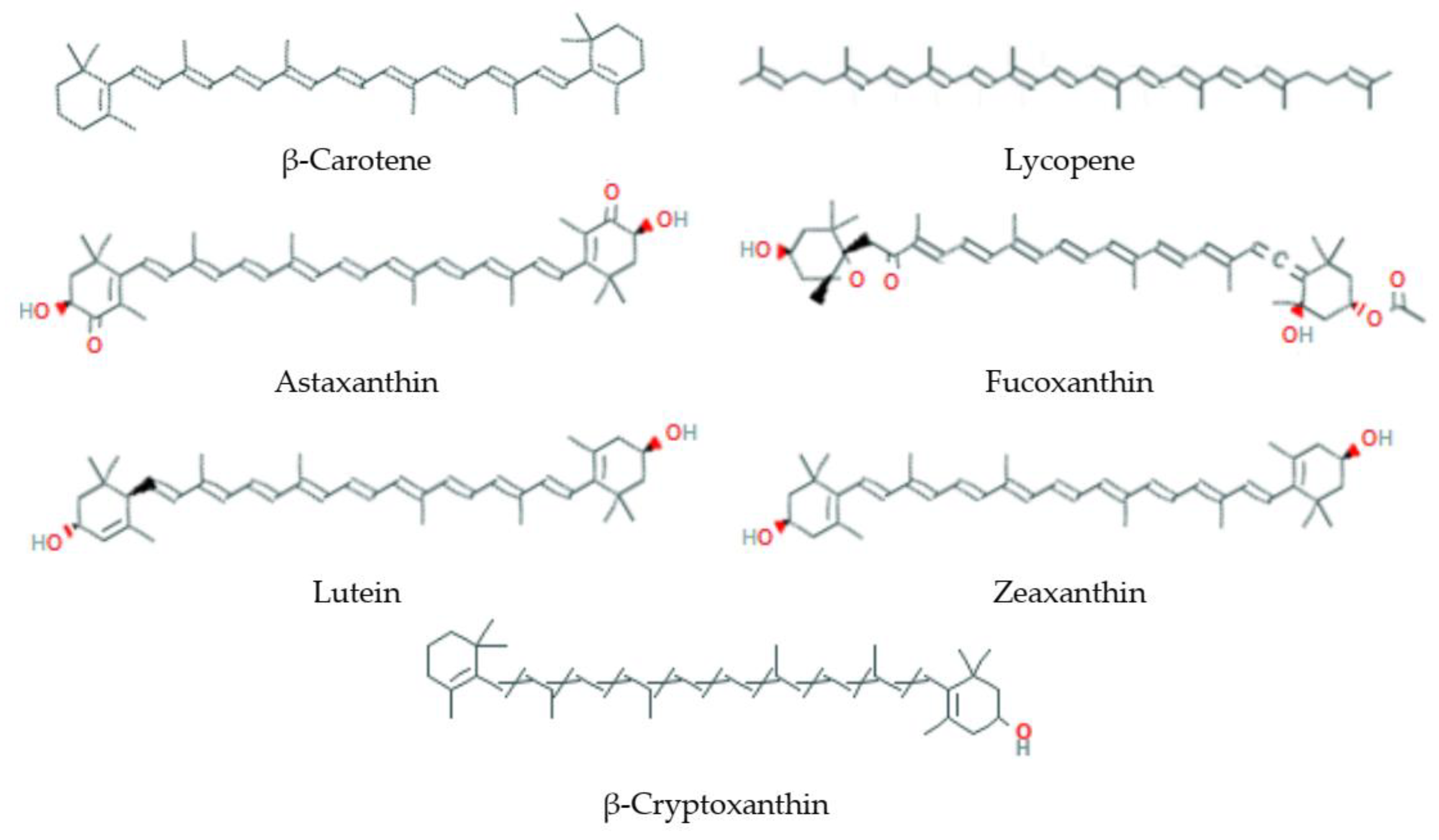{kind=link}
| Compound (Molecular Formula) | Compliance with Criteria of Geroprotectors * | References | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Primary Criteria | Secondary Criteria | |||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | ||
| Monoterpenes (C10H16) | ||||||||||
| Limonene | + | + | - | - | + | + | + | + | + | [22,23,28,35,37,38,39,40,41,44,46,47,48,413,414,415,416,417] |
| Sesquiterpenes (C15H24) | ||||||||||
| β-Caryophyllene | + | + | + | + | + | + | 0 | + | + | [58,67,79,85,86,93,96,105,147,150,164,418,419,420,421,422,423,424] |
| Diterpenes ((C10H16)2) | ||||||||||
| Dehydroabietic acid | + | + | - | - | - | + | 0 | + | 0 | [172,190,191,192,193,207,208,425,426,427] |
| Carnosol | + | + | + | + | + | + | 0 | + | + | [176,181,185,186,199,209,428,429,430,431,432] |
| Carnosic acid | + | + | + | + | + | + | 0 | + | + | [177,185,194,195,197,209,429,433,434,435,436,437] |
| Triterpenes ((C10H16)3) | ||||||||||
| Squalene | 0 | + | 0 | + | 0 | + | 0 | + | 0 | [236,266,273,308,309,310,314,320,321,324,326] |
| –Ursolic acid | + | + | + | - | + | + | + | + | + | [231,250,251,265,281,297,298,302,438,439,440,441,442,443] |
| Oleanolic acid | + | 0 | + | - | + | + | 0 | + | + | [231,254,302,322,323,330,444,445,446,447,448] |
| Maslinic acid | 0 | + | + | + | + | + | 0 | + | 0 | [231,270,294,295,325,449] |
| 18α-Glycyrrhetinic acid | + | 0 | - | - | + | + | 0 | + | 0 | [231,258,439,450,451,452,453,454,455,456] |
| Asiaticoside | 0 | + | + | 0 | + | + | 0 | + | 0 | [231,301,302,457,458,459,460,461,462,463,464,465] |
| Ginsenosides | + | + | + | - | + | + | 0 | + | + | [257,262,268,466,467,468,469,470,471,472,473,474,475,476,477,478] |
| Tetraterpenes/carotenoids ((C10H16)4) | ||||||||||
| β-Carotene | - | + | + | - | + | + | - | + | - | [344,346,347,360,361,362,365,370,376,378,383,384,479,480,481,482,483,484] |
| Lycopene | + | + | + | + | + | + | - | + | 0 | [345,348,362,365,366,376,378,479,484,485,486,487,488,489] |
| Fucoxanthin | + | + | + | + | + | + | + | + | - | [346,349,362,365,367,371,372,377,378,490] |
| Astaxanthin | + | + | + | - | + | + | + | + | + | [326,350,351,352,362,365,376,378,491,492,493,494,495] |
| Lutein | + | + | + | + | + | + | 0 | + | + | [353,362,365,376,393,394,395,481,484,496,497,498,499,500] |
| Zeaxanthin | - | + | + | + | + | + | 0 | + | 0 | [348,362,365,376,378,393,394,395,481,484,498,499,500,501] |
| β-Cryptoxanthin | 0 | + | + | 0 | + | + | 0 | + | 0 | [362,376,378,479,483,484,499,502,503,504,505,506] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proshkina, E.; Plyusnin, S.; Babak, T.; Lashmanova, E.; Maganova, F.; Koval, L.; Platonova, E.; Shaposhnikov, M.; Moskalev, A. Terpenoids as Potential Geroprotectors. Antioxidants 2020, 9, 529. https://doi.org/10.3390/antiox9060529
Proshkina E, Plyusnin S, Babak T, Lashmanova E, Maganova F, Koval L, Platonova E, Shaposhnikov M, Moskalev A. Terpenoids as Potential Geroprotectors. Antioxidants. 2020; 9(6):529. https://doi.org/10.3390/antiox9060529
Chicago/Turabian StyleProshkina, Ekaterina, Sergey Plyusnin, Tatyana Babak, Ekaterina Lashmanova, Faniya Maganova, Liubov Koval, Elena Platonova, Mikhail Shaposhnikov, and Alexey Moskalev. 2020. "Terpenoids as Potential Geroprotectors" Antioxidants 9, no. 6: 529. https://doi.org/10.3390/antiox9060529
APA StyleProshkina, E., Plyusnin, S., Babak, T., Lashmanova, E., Maganova, F., Koval, L., Platonova, E., Shaposhnikov, M., & Moskalev, A. (2020). Terpenoids as Potential Geroprotectors. Antioxidants, 9(6), 529. https://doi.org/10.3390/antiox9060529