Potential Role of Phenolic Extracts of Mentha in Managing Oxidative Stress and Alzheimer’s Disease



Abstract
:1. Development of Alzheimer’s Disease
2. Cellular and Molecular Pathways Implicated in AD
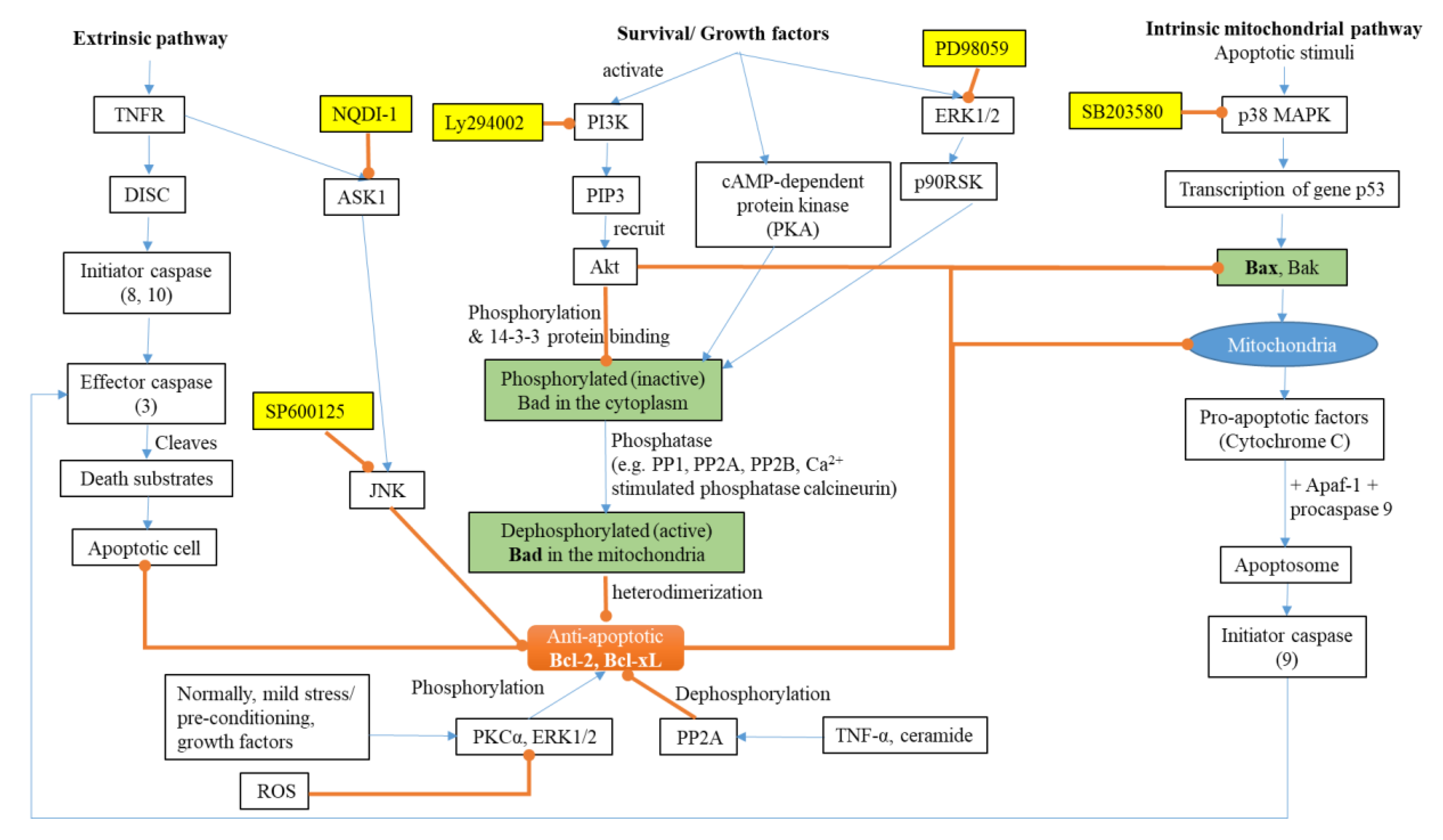
2.1. Apoptosis and AD
2.2. The Redox System and AD
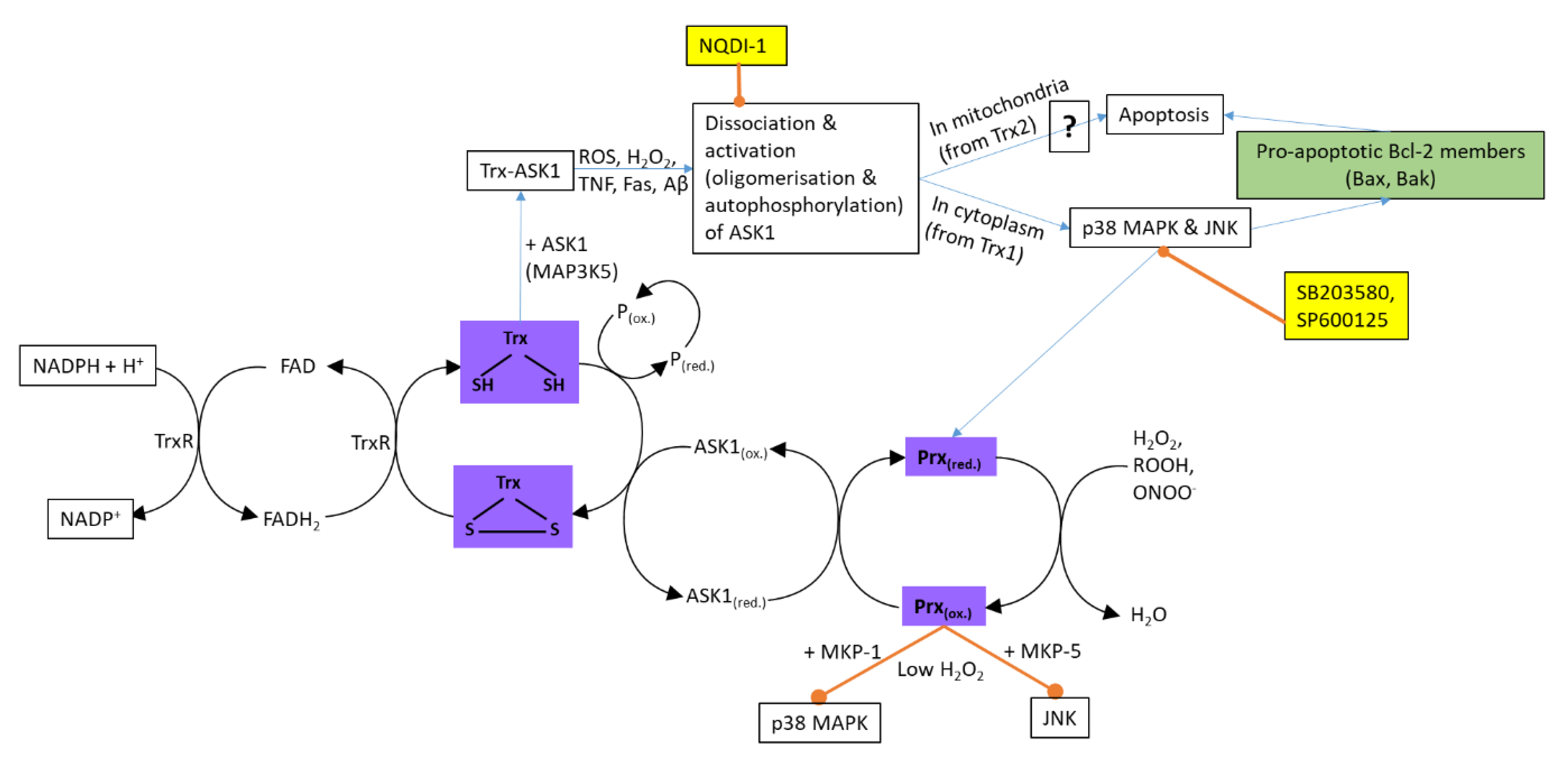
2.2.1. Thioredoxin and Peroxiredoxin
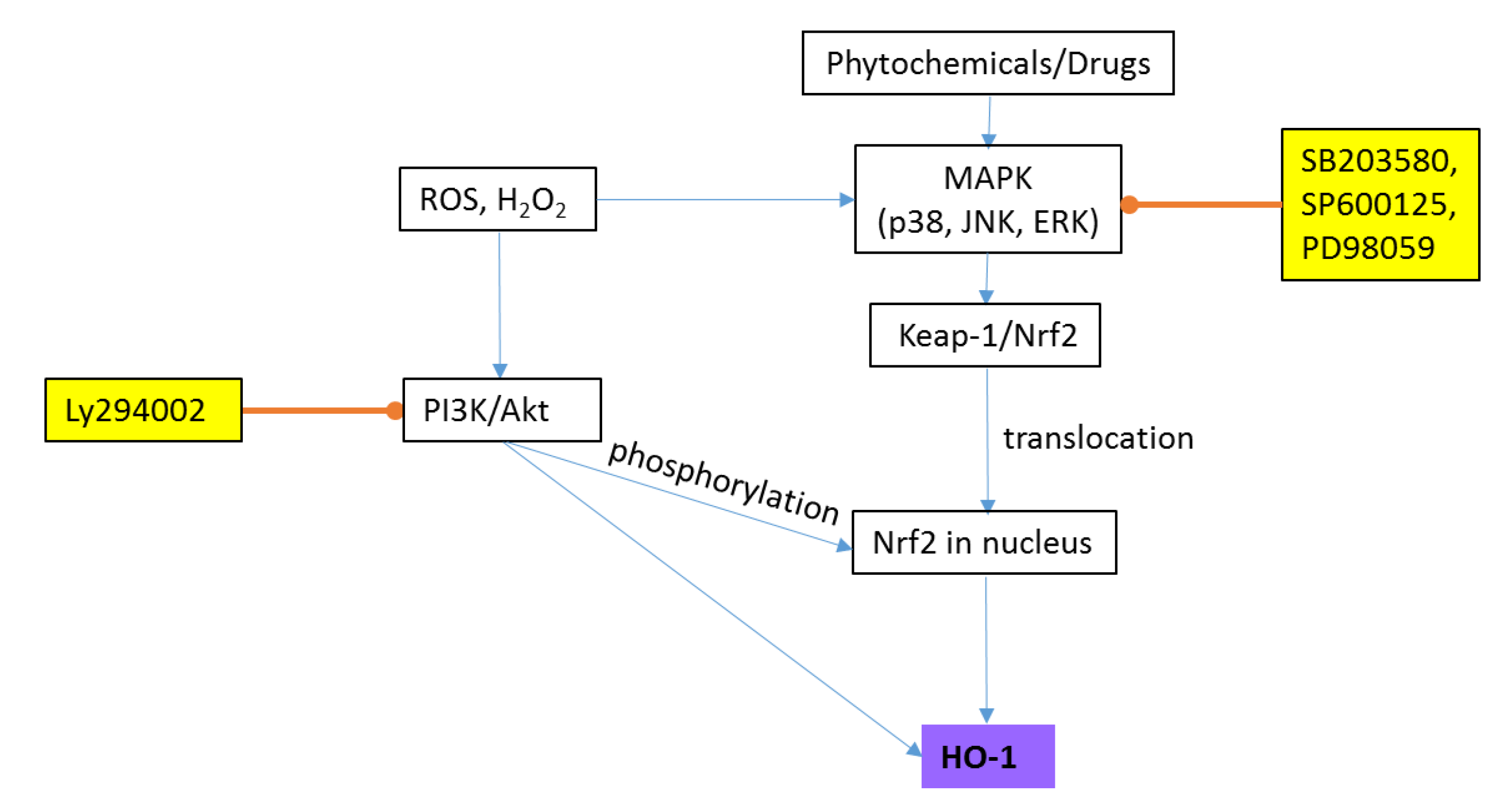
2.2.2. Heme Oxygenase-1 (HO-1)
3. Treatment of AD
4. History and Uses of Mentha
5. Chemical Composition and General Bioactivities of Mentha
6. Bioactivities of Mentha Extracts Related to CNS, Oxidative Stress and AD
7. Variation in Phenolic Composition
8. Toxicity of Mentha Extracts
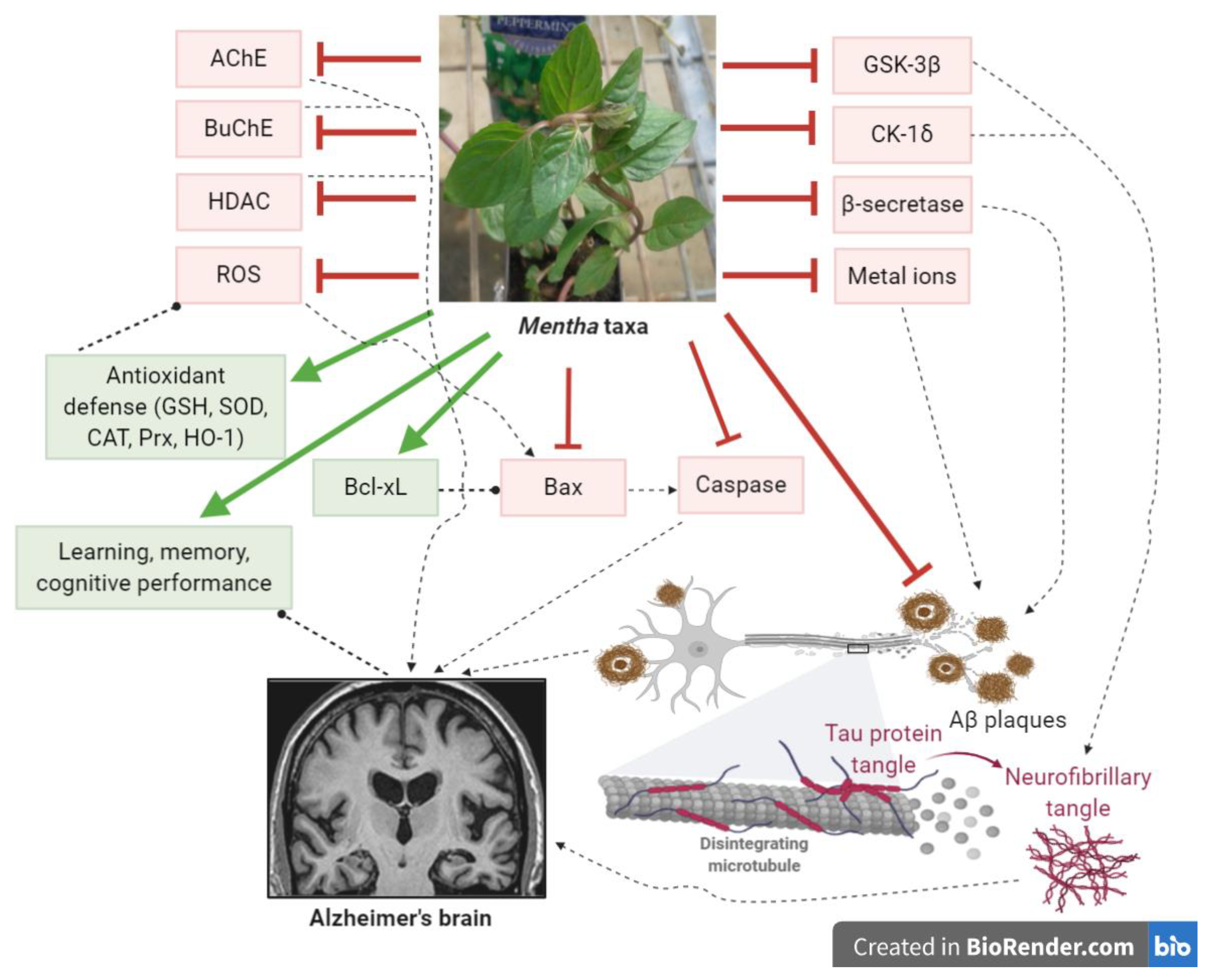
9. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Korolev, I.O. Alzheimer’s disease: A clinical and basic science review. Med. Stud. Res. J. 2014, 4, 24–33. [Google Scholar]
- Long, J.M.; Holtzman, D.M. Alzheimer disease: An update on pathobiology and treatment strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Disease International. World Alzheimer Report 2019: Attitudes to Dementia; Alzheimer’s Disease Internationals: London, UK, 2019. [Google Scholar]
- 2019 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2019, 15, 321–387. [CrossRef]
- Dementia Australia Home Page. Available online: https://www.dementia.org.au/statistics (accessed on 1 June 2020).
- Abyad, A. Alzheimer’s in the Middle East. JSM Alzheimers Dis. Relat. Dement. 2015, 2, 1–4. [Google Scholar]
- Farrag, A.-K.; Farwiz, H.M.; Khedr, E.H.; Mahfouz, R.M.; Omran, S.M. Prevalence of Alzheimer’s disease and other dementing disorders: Assiut-Upper Egypt study. Dement. Geriatr. Cogn. Disord. 1998, 9, 323–328. [Google Scholar] [CrossRef]
- Zaitoun, A.M.; Sarhan, A.-A.M.M.M.; Selim, A.M.; Mousa, G.R. Epidemiological study of dementia after retirement. Egypt. J. Neurol. Neurosurg. 2008, 45, 65–74. [Google Scholar]
- Jia, L.; Quan, M.; Fu, Y.; Zhao, T.; Li, Y.; Wei, C.; Tang, Y.; Qin, Q.; Wang, F.; Qiao, Y. Dementia in China: Epidemiology, clinical management, and research advances. Lancet Neurol. 2020, 19, 81–92. [Google Scholar] [CrossRef]
- Alzheimer Society Canada Home Page. Available online: https://alzheimer.ca/en/Home/About-dementia/What-is-dementia/Dementia-numbers (accessed on 1 July 2020).
- Rizzi, L.; Rosset, I.; Roriz-Cruz, M. Global epidemiology of dementia: Alzheimer’s and vascular types. Biomed Res. Int. 2014, 2014, 908915. [Google Scholar] [CrossRef]
- Obied, H.K.; Prenzler, P.D.; Omar, S.H.; Ismael, R.; Servili, M.; Esposto, S.; Taticchi, A.; Selvaggini, R.; Urbani, S. Pharmacology of olive biophenols. In Advances in Molecular Toxicology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 6, pp. 195–242. [Google Scholar]
- Désiré, L.; Bourdin, J.; Loiseau, N.; Peillon, H.; Picard, V.; De Oliveira, C.; Bachelot, F.; Leblond, B.; Taverne, T.; Beausoleil, E. RAC1 inhibition targets amyloid precursor protein processing by γ-secretase and decreases Aβ production in vitro and in vivo. J. Biol. Chem. 2005, 280, 37516–37525. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Fu, A.K.Y.; Ip, N.Y. Synaptic dysfunction in Alzheimer’s disease: Mechanisms and therapeutic strategies. Pharmacol. Ther. 2019, 195, 186–198. [Google Scholar] [CrossRef]
- Panza, F.; Lozupone, M.; Logroscino, G.; Imbimbo, B.P. A critical appraisal of amyloid-β-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Khan, M.; Shah, S.A.; Saeed, K.; Kim, M.O. Natural antioxidant anthocyanins—A hidden therapeutic candidate in metabolic disorders with major focus in neurodegeneration. Nutrients 2019, 11, 1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moneim, A.E.A. Oxidant/antioxidant imbalance and the risk of Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Gunn, A.P.; Wong, B.X.; Johanssen, T.; Griffith, J.C.; Masters, C.L.; Bush, A.I.; Barnham, K.J.; Duce, J.A.; Cherny, R.A. Amyloid-β peptide Aβ3pE-42 induces lipid peroxidation, membrane permeabilization, and calcium influx in neurons. J. Biol. Chem. 2016, 291, 6134–6145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwon, A.R.; Park, J.S.; Arumugam, T.V.; Kwon, Y.K.; Chan, S.L.; Kim, S.H.; Baik, S.H.; Yang, S.; Yun, Y.K.; Choi, Y.; et al. Oxidative lipid modification of nicastrin enhances amyloidogenic γ-secretase activity in Alzheimer’s disease. Aging Cell 2012, 11, 559–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenough, M.A.; Camakaris, J.; Bush, A.I. Metal dyshomeostasis and oxidative stress in Alzheimer’s disease. Neurochem. Int. 2013, 62, 540–555. [Google Scholar] [CrossRef]
- Wyss-Coray, T.; Rogers, J. Inflammation in Alzheimer disease—A brief review of the basic science and clinical literature. Cold Spring Harb. Perspect. Med. 2012, 2, 1–23. [Google Scholar] [CrossRef]
- Xu, K.; Dai, X.-L.; Huang, H.-C.; Jiang, Z.-F. Targeting HDACs: A promising therapy for Alzheimer’s disease. Oxidative Med. Cell. Longev. 2011, 2011, 1–5. [Google Scholar] [CrossRef] [Green Version]
- De Ruijter, A.J.; Van Gennip, A.H.; Caron, H.N.; Kemp, S.; Van Kuilenburg, A.B. Histone deacetylases (HDACs): Characterization of the classical HDAC family. Biochem. J. 2003, 370, 737–749. [Google Scholar] [CrossRef]
- Wojsiat, J.; Zoltowska, K.M.; Laskowska-Kaszub, K.; Wojda, U. Oxidant/antioxidant imbalance in Alzheimer’s disease: Therapeutic and diagnostic prospects. Oxidative Med. Cell. Longev. 2018, 2018, 6435861. [Google Scholar] [CrossRef] [Green Version]
- Obulesu, M.; Lakshmi, M.J. Apoptosis in Alzheimer’s disease: An understanding of the physiology, pathology and therapeutic avenues. Neurochem. Res. 2014, 39, 2301–2312. [Google Scholar] [CrossRef]
- Rang, H.P.; Ritter, J.M.; Flower, R.J.; Henderson, G. Rang & Dale’s Pharmacology; Elsevier: London, UK, 2015. [Google Scholar]
- Portt, L.; Norman, G.; Clapp, C.; Greenwood, M.; Greenwood, M.T. Anti-apoptosis and cell survival: A review. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 238–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohn, T.T.; Head, E. Caspase activation in Alzheimer’s disease: Early to rise and late to bed. Rev. Neurosci. 2008, 19, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Shimohama, S. Apoptosis in Alzheimer’s disease—An update. Apoptosis 2000, 5, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Howells, C.C.; Baumann, W.T.; Samuels, D.C.; Finkielstein, C.V. The Bcl-2-associated death promoter (BAD) lowers the threshold at which the Bcl-2-interacting domain death agonist (BID) triggers mitochondria disintegration. J. Theor. Biol. 2011, 271, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, W.; Jiang, Z.-G.; Ghanbari, H.A. Oxidative stress and neurodegenerative disorders. Int. J. Mol. Sci. 2013, 14, 24438–24475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, H.; Noguchi, T.; Takeda, K.; Ichijo, H. Pathophysiological roles of ASK1-MAP kinase signaling pathways. J. Biochem. Mol. Biol. 2007, 40, 1–6. [Google Scholar] [CrossRef]
- Tan, Y.; Ruan, H.; Demeter, M.R.; Comb, M.J. p90RSK blocks Bad-mediated cell death via a protein kinase C-dependent pathway. J. Biol. Chem. 1999, 274, 34859–34867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Xu, S. ERK1/2 MAP kinases in cell survival and apoptosis. IUBMB Life 2006, 58, 621–631. [Google Scholar] [CrossRef]
- Song, J.; Park, K.A.; Lee, W.T.; Lee, J.E. Apoptosis signal regulating kinase 1 (ASK1): Potential as a therapeutic target for Alzheimer’s disease. Int. J. Mol. Sci. 2014, 15, 2119–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latimer, H.R.; Veal, E.A. Peroxiredoxins in regulation of MAPK signalling pathways; sensors and barriers to signal transduction. Mol. Cells 2016, 39, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, glutaredoxins, and peroxiredoxins—Molecular mechanisms and health significance: From cofactors to antioxidants to redox signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef] [PubMed]
- Andoh, T.; Chock, P.B.; Chiueh, C.C. The roles of thioredoxin in protection against oxidative stress-induced apoptosis in SH-SY5Y cells. J. Biol. Chem. 2002, 277, 9655–9660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Al-Lamki, R.; Bai, L.; Streb, J.W.; Miano, J.M.; Bradley, J.; Min, W. Thioredoxin-2 inhibits mitochondria-located ASK1-mediated apoptosis in a JNK-independent manner. Circ. Res. 2004, 94, 1483–1491. [Google Scholar] [CrossRef] [Green Version]
- Akterin, S.; Cowburn, R.F.; Miranda-Vizuete, A.; Jiménez, A.; Bogdanovic, N.; Winblad, B.; Cedazo-Minguez, A. Involvement of glutaredoxin-1 and thioredoxin-1 in β-amyloid toxicity and Alzheimer’s disease. Cell Death Differ. 2006, 13, 1454–1465. [Google Scholar] [CrossRef] [Green Version]
- Di Domenico, F.; Sultana, R.; Tiu, G.F.; Scheff, N.N.; Perluigi, M.; Cini, C.; Butterfield, D.A. Protein levels of heat shock proteins 27, 32, 60, 70, 90 and thioredoxin-1 in amnestic mild cognitive impairment: An investigation on the role of cellular stress response in the progression of Alzheimer disease. Brain Res. 2010, 1333, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Lovell, M.A.; Xie, C.; Gabbita, S.P.; Markesbery, W.R. Decreased thioredoxin and increased thioredoxin reductase levels in Alzheimer’s disease brain. Free Radic. Biol. Med. 2000, 28, 418–427. [Google Scholar] [CrossRef]
- Jarvis, R.M.; Hughes, S.M.; Ledgerwood, E.C. Peroxiredoxin 1 functions as a signal peroxidase to receive, transduce, and transmit peroxide signals in mammalian cells. Free Radic. Biol. Med. 2012, 53, 1522–1530. [Google Scholar] [CrossRef]
- Li, B.; Ishii, T.; Tan, C.P.; Soh, J.-W.; Goff, S.P. Pathways of induction of peroxiredoxin I expression in osteoblasts: Roles of p38 mitogen-activated protein kinase and protein kinase C. J. Biol. Chem. 2002, 277, 12418–12422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krapfenbauer, K.; Engidawork, E.; Cairns, N.; Fountoulakis, M.; Lubec, G. Aberrant expression of peroxiredoxin subtypes in neurodegenerative disorders. Brain Res. 2003, 967, 152–160. [Google Scholar] [CrossRef]
- Cumming, R.C.; Dargusch, R.; Fischer, W.H.; Schubert, D. Increase in expression levels and resistance to sulfhydryl oxidation of peroxiredoxin isoforms in amyloid β-resistant nerve cells. J. Biol. Chem. 2007, 282, 30523–30534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Fountoulakis, M.; Cairns, N.; Lubec, G. Protein levels of human peroxiredoxin subtypes in brains of patients with Alzheimer’s disease and Down syndrome. In Protein Expression in Down Syndrome Brain; Springer: Vienna, Austria, 2001; pp. 223–235. [Google Scholar] [CrossRef]
- Pae, H.-O.; Kim, E.-C.; Chung, H.-T. Integrative survival response evoked by heme oxygenase-1 and heme metabolites. J. Clin. Biochem. Nutr. 2008, 42, 197–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, L.; Yang, C.; Gu, M.; Zheng, D.; Lin, L.; Wang, X.; Lan, A.; Hu, F.; Feng, J. PI3K/Akt signaling pathway-induced heme oxygenase-1 upregulation mediates the adaptive cytoprotection of hydrogen peroxide preconditioning against oxidative injury in PC12 cells. Int. J. Mol. Med. 2012, 30, 314–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennery, P.A. Signaling function of heme oxygenase proteins. Antioxid. Redox Signal. 2014, 20, 1743–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J. Neurol. Neurosurg. Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef]
- Adsersen, A.; Gauguin, B.; Gudiksen, L.; Jäger, A.K. Screening of plants used in Danish folk medicine to treat memory dysfunction for acetylcholinesterase inhibitory activity. J. Ethnopharmacol. 2006, 104, 418–422. [Google Scholar] [CrossRef]
- Imbimbo, B.P.; Watling, M. Investigational BACE inhibitors for the treatment of Alzheimer’s disease. Expert Opin. Investig. Drugs 2019, 28, 967–975. [Google Scholar] [CrossRef]
- Vladimir-Knežević, S.; Blažeković, B.; Kindl, M.; Vladić, J.; Lower-Nedza, A.D.; Brantner, A.H. Acetylcholinesterase inhibitory, antioxidant and phytochemical properties of selected medicinal plants of the Lamiaceae family. Molecules 2014, 19, 767–782. [Google Scholar] [CrossRef] [Green Version]
- Cicero, A.F.; Fogacci, F.; Banach, M. Botanicals and phytochemicals active on cognitive decline: The clinical evidence. Pharmacol. Res. 2018, 130, 204–212. [Google Scholar] [CrossRef]
- Shu, Y.-Z. Recent natural products based drug development: A pharmaceutical industry perspective. J. Nat. Prod. 1998, 61, 1053–1071. [Google Scholar] [CrossRef]
- Heinrich, M.; Lee Teoh, H. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.P.; Chin, Y.-W.; Kinghorn, A.D. The role of pharmacognosy in modern medicine and pharmacy. Curr. Drug Targets 2006, 7, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Poison Control - National Capital Poison Center Home Page. Available online: https://www.poison.org/articles/tacrine-171 (accessed on 1 June 2020).
- Declercq, L.D.; Vandenberghe, R.; Van Laere, K.; Verbruggen, A.; Bormans, G. Drug development in Alzheimer’s disease: The contribution of PET and SPECT. Front. Pharmacol. 2016, 7, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Tucker, A.O.; Naczi, R.F.C. Mentha: An Overview of its Classification and Relationships. In Mint: The Genus Mentha; Lawrence, B.M., Ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 3–39. [Google Scholar]
- Jabeen, A.; Guo, B.; Abbasi, B.H.; Shinwari, Z.K.; Mahmood, T. Phylogenetics of selected Mentha species on the basis of rps8, rps11 and rps14 chloroplast genes. J. Med. Plants Res. 2012, 6, 30–36. [Google Scholar] [CrossRef]
- Mahendran, G.; Rahman, L.U. Ethnomedicinal, phytochemical and pharmacological updates on peppermint (Mentha × piperita L.)—A review. Phytother. Res. 2020, 1–52. [Google Scholar] [CrossRef]
- Peixoto, I.T.A.; Furletti, V.F.; Anibal, P.C.; Duarte, M.C.T.; Höfling, J.F. Potential pharmacological and toxicological basis of the essential oil from Mentha spp. Revista de Ciências Farmacêuticas Básica e Aplicada 2010, 30, 235–239. [Google Scholar]
- Naghibi, F.; Mosaddegh, M.; Mohammadi Motamed, M.; Ghorbani, A. Labiatae family in folk medicine in Iran: From ethnobotany to pharmacology. Iran. J. Pharm. Res. 2010, 4, 63–79. [Google Scholar] [CrossRef]
- Karousou, R.; Balta, M.; Hanlidou, E.; Kokkini, S. “Mints”, smells and traditional uses in Thessaloniki (Greece) and other Mediterranean countries. J. Ethnopharmacol. 2007, 109, 248–257. [Google Scholar] [CrossRef]
- Mata, A.T.; Proença, C.; Ferreira, A.R.; Serralheiro, M.L.M.; Nogueira, J.M.F.; Araújo, M.E.M. Antioxidant and antiacetylcholinesterase activities of five plants used as Portuguese food spices. Food Chem. 2007, 103, 778–786. [Google Scholar] [CrossRef]
- Ayyobi, H.; Peyvast, G.-A.; Olfati, J.-A. Effect of drying methods on essential oil yield, total phenol content and antioxidant capacity of peppermint and dill. Ratar. I Povrt. 2014, 51, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Fatiha, B.; Didier, H.; Naima, G.; Khodir, M.; Martin, K.; Léocadie, K.; Caroline, S.; Mohamed, C.; Pierre, D. Phenolic composition, in vitro antioxidant effects and tyrosinase inhibitory activity of three Algerian Mentha species: M. spicata (L.), M. pulegium (L.) and M. rotundifolia (L.) Huds (Lamiaceae). Ind. Crop. Prod. 2015, 74, 722–730. [Google Scholar] [CrossRef]
- Olsen, H.T.; Stafford, G.I.; van Staden, J.; Christensen, S.B.; Jäger, A.K. Isolation of the MAO-inhibitor naringenin from Mentha aquatica L. J. Ethnopharmacol. 2008, 117, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Biswas, N.N.; Saha, S.; Ali, M. Antioxidant, antimicrobial, cytotoxic and analgesic activities of ethanolic extract of Mentha arvensis L. Asian Pac. J. Trop. Biomed. 2014, 4, 792–797. [Google Scholar] [CrossRef]
- Kapp, K. Polyphenolic and Essential Oil Composition of Mentha and Their Antimicrobial Effect. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2015. [Google Scholar]
- Brahmi, F.; Khodir, M.; Mohamed, C.; Pierre, D. Chemical composition and biological activities of Mentha species. In Aromatic and Medicinal Plants-Back to Nature; El-Shemy, H., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Hanafy, D.M.; Prenzler, P.D.; Burrows, G.E.; Ryan, D.; Nielsen, S.; El Sawi, S.A.; El Alfy, T.S.; Abdelrahman, E.H.; Obied, H.K. Biophenols of mints: Antioxidant, acetylcholinesterase, butyrylcholinesterase and histone deacetylase inhibition activities targeting Alzheimer’s disease treatment. J. Funct. Foods 2017, 33, 345–362. [Google Scholar] [CrossRef]
- Salin, O.; Tormakangas, L.; Leinonen, M.; Saario, E.; Hagstrom, M.; Ketola, R.A.; Saikku, P.; Vuorela, H.; Vuorela, P.M. Corn mint (Mentha arvensis) extract diminishes acute Chlamydia pneumoniae infection in vitro and in vivo. J. Agric. Food Chem. 2011, 59, 12836–12842. [Google Scholar] [CrossRef] [PubMed]
- Kosar, M.; Dorman, H.D.; Can Baser, K.H.; Hiltunen, R. Screening of free radical scavenging compounds in water extracts of Mentha samples using a postcolumn derivatization method. J. Agric. Food Chem. 2004, 52, 5004–5010. [Google Scholar] [CrossRef]
- Stagos, D.; Portesis, N.; Spanou, C.; Mossialos, D.; Aligiannis, N.; Chaita, E.; Panagoulis, C.; Reri, E.; Skaltsounis, L.; Tsatsakis, A.M.; et al. Correlation of total polyphenolic content with antioxidant and antibacterial activity of 24 extracts from Greek domestic Lamiaceae species. Food Chem. Toxicol. 2012, 50, 4115–4124. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Ioele, G.; Statti, G.A.; Marrelli, M.; Ragno, G.; Menichini, F. Antiproliferative activity against human tumor cell lines and toxicity test on Mediterranean dietary plants. Food Chem. Toxicol. 2008, 46, 3325–3332. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Sosa, S.; Marrelli, M.; Menichini, F.; Statti, G.A.; Uzunov, D.; Tubaro, A.; Menichini, F.; Loggia, R.D. In vivo anti-inflammatory and in vitro antioxidant activities of Mediterranean dietary plants. J. Ethnopharmacol. 2008, 116, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Baris, O.; Karadayi, M.; Yanmis, D.; Guvenalp, Z.; Bal, T.; Gulluce, M. Isolation of 3 flavonoids from Mentha longifolia (L.) Hudson subsp. longifolia and determination of their genotoxic potentials by using the E. coli WP2 test system. J. Food Sci. 2011, 76, T212–T217. [Google Scholar] [CrossRef]
- Orhan, F.; Barış, Ö.; Yanmış, D.; Bal, T.; Güvenalp, Z.; Güllüce, M. Isolation of some luteolin derivatives from Mentha longifolia (L.) Hudson subsp. longifolia and determination of their genotoxic potencies. Food Chem. 2012, 135, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Kapp, K.; Hakala, E.; Orav, A.; Pohjala, L.; Vuorela, P.; Püssa, T.; Vuorela, H.; Raal, A. Commercial peppermint (Mentha × piperita L.) teas: Antichlamydial effect and polyphenolic composition. Food Res. Int. 2013, 53, 758–766. [Google Scholar] [CrossRef]
- Dorman, H.D.; Kosar, M.; Kahlos, K.; Holm, Y.; Hiltunen, R. Antioxidant properties and composition of aqueous extracts from Mentha species, hybrids, varieties, and cultivars. J. Agric. Food Chem. 2003, 51, 4563–4569. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Proenca, C.; Serralheiro, M.L.M.; Araujo, M.E.M. The in vitro screening for acetylcholinesterase inhibition and antioxidant activity of medicinal plants from Portugal. J. Ethnopharmacol. 2006, 108, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Jäger, A.K.; Almqvist, J.P.; Vangsøe, S.A.K.; Stafford, G.I.; Adsersen, A.; Van Staden, J. Compounds from Mentha aquatica with affinity to the GABA-benzodiazepine receptor. S. Afr. J. Bot. 2007, 73, 518–521. [Google Scholar] [CrossRef]
- Moss, M.; Hewitt, S.; Moss, L.; Wesnes, K. Modulation of cognitive performance and mood by aromas of peppermint and ylang-ylang. Int. J. Neurosci. 2008, 118, 59–77. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C. Neuroprotective phenolics in medicinal plants. Arch. Pharmacal Res. 2010, 33, 1611–1632. [Google Scholar] [CrossRef] [PubMed]
- Sabogal-Guáqueta, A.M.; Muñoz-Manco, J.I.; Ramírez-Pineda, J.R.; Lamprea-Rodriguez, M.; Osorio, E.; Cardona-Gómez, G.P. The flavonoid quercetin ameliorates Alzheimer’s disease pathology and protects cognitive and emotional function in aged triple transgenic Alzheimer’s disease model mice. Neuropharmacology 2015, 93, 134–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, H.T.; Samarat, K.; Takamura, Y.; Azo-Oussou, A.F.; Nakazono, Y.; Vestergaard, M.d.C. Polyphenols modulate Alzheimer’s amyloid beta aggregation in a structure-dependent manner. Nutrients 2019, 11, 756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, V.; Martín, S.; Gómez--Serranillos, M.P.; Carretero, M.E.; Jäger, A.K.; Calvo, M.I. Neuroprotective and neurochemical properties of mint extracts. Phytother. Res. 2010, 24, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Gürbüz, P.; Martinez, A.; Pérez, C.; Martínez-González, L.; Göger, F.; Ayran, İ. Potential anti-Alzheimer effects of selected Lamiaceae plants through polypharmacology on glycogen synthase kinase-3β, β-secretase, and casein kinase 1δ. Ind. Crop. Prod. 2019, 138, 1–6. [Google Scholar] [CrossRef]
- Benabdallah, A.; Rahmoune, C.; Boumendjel, M.; Aissi, O.; Messaoud, C. Total phenolic content and antioxidant activity of six wild Mentha species (Lamiaceae) from northeast of Algeria. Asian Pac. J. Trop. Biomed. 2016, 6, 760–766. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Zengin, G.; Bahadori, S.; Dinparast, L.; Movahhedin, N. Phenolic composition and functional properties of wild mint (Mentha longifolia var. calliantha (Stapf) Briq.). Int. J. Food Prop. 2018, 21, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Hanafy, D.M.; Prenzler, P.D.; Burrows, G.E.; Gurusinghe, S.; Thejer, B.M.; Obied, H.K.; Hill, R.A. Neuroprotective activity of Mentha species on hydrogen peroxide-induced apoptosis in SH-SY5Y cells. Nutrients 2020, 12, 1366. [Google Scholar] [CrossRef]
- Lee, H.; Yeom, M.; Shin, S.; Jeon, K.; Park, D.; Jung, E. Protective effects of aqueous extract of Mentha suaveolens against oxidative stress-induced damages in human keratinocyte HaCaT cells. Evid. Based Complementary Altern. Med. 2019, 2019, 1–8. [Google Scholar] [CrossRef]
- Joshi, H.; Bhadania, M. Evaluation of freeze dried extract of Mentha piperita in management of cognitive dysfunctions in mice. Alzheimers Dement. 2014, 10, 461. [Google Scholar] [CrossRef]
- Hassan, H.A.; Hafez, H.S.; Goda, M.S. Mentha piperita as a pivotal neuro-protective agent against gamma irradiation induced DNA fragmentation and apoptosis: Mentha extract as a neuroprotective against gamma irradiation. Cytotechnology 2013, 65, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Arshad, H.; Kalam, N.A.; Anshu, M.; Hasin, A.M.; Shadma, W. Effect of the aqueous extract of Mentha arvensis on haloperidol induced catalepsy in albino mice. J. Clin. Diagn. Res 2012, 6, 542–546. [Google Scholar]
- Farr, S.A.; Niehoff, M.L.; Ceddia, M.A.; Herrlinger, K.A.; Lewis, B.J.; Feng, S.; Welleford, A.; Butterfield, D.A.; Morley, J.E. Effect of botanical extracts containing carnosic acid or rosmarinic acid on learning and memory in SAMP8 mice. Physiol. Behav. 2016, 165, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Herrlinger, K.A.; Nieman, K.M.; Sanoshy, K.D.; Fonseca, B.A.; Lasrado, J.A.; Schild, A.L.; Maki, K.C.; Wesnes, K.A.; Ceddia, M.A. Spearmint extract improves working memory in men and women with age-associated memory impairment. J. Altern. Complementary Med. 2018, 24, 37–47. [Google Scholar] [CrossRef]
- Oinonen, P.P.; Jokela, J.K.; Hatakka, A.I.; Vuorela, P.M. Linarin, a selective acetylcholinesterase inhibitor from Mentha arvensis. Fitoterapia 2006, 77, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Fan, P.; Perez, R.G.; Lou, H. Neuroprotective effects of linarin through activation of the PI3K/Akt pathway in amyloid-β-induced neuronal cell death. Bioorganic Med. Chem. 2011, 19, 4021–4027. [Google Scholar] [CrossRef] [PubMed]
- Riachi, L.G.; De Maria, C.A.B. Peppermint antioxidants revisited. Food Chem. 2015, 176, 72–81. [Google Scholar] [CrossRef]
- Chien, M.-Y.; Chuang, C.-H.; Chern, C.-M.; Liou, K.-T.; Liu, D.-Z.; Hou, Y.-C.; Shen, Y.-C. Salvianolic acid A alleviates ischemic brain injury through the inhibition of inflammation and apoptosis and the promotion of neurogenesis in mice. Free Radic. Biol. Med. 2016, 99, 508–519. [Google Scholar] [CrossRef]
- Lee, H.J.; Cho, H.-S.; Park, E.; Kim, S.; Lee, S.-Y.; Kim, C.-S.; Kim, D.K.; Kim, S.-J.; Chun, H.S. Rosmarinic acid protects human dopaminergic neuronal cells against hydrogen peroxide-induced apoptosis. Toxicology 2008, 250, 109–115. [Google Scholar] [CrossRef]
- Rong, H.; Liang, Y.; Niu, Y. Rosmarinic acid attenuates β-amyloid-induced oxidative stress via Akt/GSK-3β/Fyn-mediated Nrf2 activation in PC12 cells. Free Radic. Biol. Med. 2018, 120, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, T.; Ono, K.; Murase, A.; Yamada, M. Phenolic compounds prevent Alzheimer’s pathology through different effects on the amyloid-β aggregation pathway. Am. J. Pathol. 2009, 175, 2557–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.J.; Sowndhararajan, K.; Park, H.-B.; Kim, S.-Y.; Kim, S.; Kim, D.H.; Choi, J.W.; Jang, D.S.; Ryu, J.H.; Park, S.J. Danshensu attenuates scopolamine and amyloid-β-induced cognitive impairments through the activation of PKA-CREB signaling in mice. Neurochem. Int. 2019, 131, 104537. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, W.; Xu, L.; Chen, L. In Salvia miltiorrhiza, phenolic acids possess protective properties against amyloid β-induced cytotoxicity, and tanshinones act as acetylcholinesterase inhibitors. Environ. Toxicol. Pharmacol. 2011, 31, 443–452. [Google Scholar] [CrossRef]
- Chong, C.-M.; Zhou, Z.-Y.; Razmovski-Naumovski, V.; Cui, G.-Z.; Zhang, L.-Q.; Sa, F.; Hoi, P.-M.; Chan, K.; Lee, S.M.-Y. Danshensu protects against 6-hydroxydopamine-induced damage of PC12 cells in vitro and dopaminergic neurons in zebrafish. Neurosci. Lett. 2013, 543, 121–125. [Google Scholar] [CrossRef]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a potent antioxidant: Implications for neurodegenerative disorders. Oxidative Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef]
- Hajialyani, M.; Hosein Farzaei, M.; Echeverría, J.; Nabavi, S.M.; Uriarte, E.; Sobarzo-Sánchez, E. Hesperidin as a neuroprotective agent: A review of animal and clinical evidence. Molecules 2019, 24, 648. [Google Scholar] [CrossRef] [Green Version]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Lin, M.-T.; Zhu, Y.-D.; Xu, H.-L.; Zhao, Y.-Z. Recent Trends in potential therapeutic applications of the dietary flavonoid didymin. Molecules 2018, 23, 2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dourado, N.S.; Souza, C.d.S.; de Almeida, M.M.A.; Bispo da Silva, A.; dos Santos, B.L.; Silva, V.D.A.; De Assis, A.M.; da Silva, J.S.; Souza, D.O.; Costa, M.d.F.D.; et al. Neuroimmunomodulatory and neuroprotective effects of the flavonoid apigenin in in vitro models of neuroinflammation associated with Alzheimer’s disease. Front. Aging Neurosci. 2020, 12. [Google Scholar] [CrossRef]
- Zhang, F.; Li, F.; Chen, G. Neuroprotective effect of apigenin in rats after contusive spinal cord injury. Neurol. Sci. 2014, 35, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.J.; Li, F.; Yu, P.H.; Chan, S.W. Neuroprotective effects of luteolin against apoptosis induced by 6-hydroxydopamine on rat pheochromocytoma PC12 cells. Pharm. Biol. 2013, 51, 190–196. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.; Ghafoor, K. Total phenols and antioxidant activities of leaf and stem extracts from coriander, mint and parsley grown in Saudi Arabia. Pak. J. Bot. 2011, 43, 2235–2237. [Google Scholar]
- Benedec, D.; Vlase, L.; Oniga, I.; Mot, A.C.; Silaghi-Dumitrescu, R.; Hanganu, D.; Tiperciuc, B.; Crişan, G. LC-MS analysis and antioxidant activity of phenolic compounds from two indigenous species of Mentha. Note I. Farmacia 2013, 61, 262–267. [Google Scholar]
- Biswas, A.K.; Chatli, M.K.; Sahoo, J. Antioxidant potential of curry (Murraya koenigii L.) and mint (Mentha spicata) leaf extracts and their effect on colour and oxidative stability of raw ground pork meat during refrigeration storage. Food Chem. 2012, 133, 467–472. [Google Scholar] [CrossRef]
- Malik, B.; Sharma, N.R.; Soni, G. Influence of agro-climatic conditions on antioxidant potential of Mentha species. J. Pharm. Res. 2013, 7, 427–432. [Google Scholar] [CrossRef]
- Yi, W.; Wetzstein, H.Y. Biochemical, biological and histological evaluation of some culinary and medicinal herbs grown under greenhouse and field conditions. J. Sci. Food Agric. 2010, 90, 1063–1070. [Google Scholar] [CrossRef]
- Antolovich, M.; Prenzler, P.; Robards, K.; Ryan, D. Sample preparation in the determination of phenolic compounds in fruits. Analyst 2000, 125, 989–1009. [Google Scholar] [CrossRef]
- Patonay, K.; Szalontai, H.; Csugány, J.; Szabó-Hudák, O.; Kónya, E.P.; Németh, É.Z. Comparison of extraction methods for the assessment of total polyphenol content and in vitro antioxidant capacity of horsemint (Mentha longifolia (L.) L.). J. Appl. Res. Med. Aromat. Plants 2019, 15. [Google Scholar] [CrossRef]
- Bimakr, M.; Rahman, R.A.; Taip, F.S.; Ganjloo, A.; Salleh, L.M.; Selamat, J.; Hamid, A.; Zaidul, I.S.M. Comparison of different extraction methods for the extraction of major bioactive flavonoid compounds from spearmint (Mentha spicata L.) leaves. Food Bioprod. Process. 2011, 89, 67–72. [Google Scholar] [CrossRef]
- Sulaiman, S.F.; Sajak, A.A.B.; Ooi, K.L.; Seow, E.W. Effect of solvents in extracting polyphenols and antioxidants of selected raw vegetables. J. Food Compos. Anal. 2011, 24, 506–515. [Google Scholar] [CrossRef]
- Arslan, D.; Özcan, M.M.; Mengeş, H.O. Evaluation of drying methods with respect to drying parameters, some nutritional and colour characteristics of peppermint (Mentha x piperita L.). Energy Convers. Manag. 2010, 51, 2769–2775. [Google Scholar] [CrossRef]
- Hayat, K. Impact of drying methods on the functional properties of peppermint (Mentha piperita L.) leaves. Sci. Lett. 2020, 8, 36–42. [Google Scholar]
- Safaiee, P.; Taghipour, A.; Vahdatkhoram, F.; Movagharnejad, K. Extraction of phenolic compounds from Mentha aquatica: The effects of sonication time, temperature and drying method. Chem. Pap. 2019, 73, 3067–3073. [Google Scholar] [CrossRef]
- Fletcher, R.S.; Slimmon, T.; Kott, L.S. Environmental factors affecting the accumulation of rosmarinic acid in spearmint (Mentha spicata L.) and peppermint (Mentha piperita L.). Open Agric. J. 2010, 4, 10–16. [Google Scholar] [CrossRef]
- Voirin, B.; Saunois, A.; Bayet, C. Free flavonoid aglycones from Mentha × piperita: Developmental, chemotaxonomical and physiological aspects. Biochem. Syst. Ecol. 1994, 22, 95–99. [Google Scholar] [CrossRef]
- Neves, J.M. Concerns regarding the toxicity of Mentha x piperita. Ann. Med. Chem. Res. 2005, 60, 87–100. [Google Scholar]
- Caro, D.C.; Rivera, D.E.; Ocampo, Y.; Franco, L.A.; Salas, R.D. Pharmacological evaluation of Mentha spicata L. and Plantago major L., medicinal plants used to treat anxiety and insomnia in Colombian Caribbean Coast. Evid Based Complement Altern. Med 2018, 2018, 5921514. [Google Scholar] [CrossRef] [PubMed]
- Akdogan, M.; Kilinç, I.; Oncu, M.; Karaoz, E.; Delibas, N. Investigation of biochemical and histopathological effects of Mentha piperita L. and Mentha spicata L. on kidney tissue in rats. Hum. Exp. Toxicol. 2003, 22, 213–219. [Google Scholar] [CrossRef]
- Akdogan, M.; Ozguner, M.; Aydin, G.; Gokalp, O. Investigation of biochemical and histopathological effects of Mentha piperita Labiatae and Mentha spicata Labiatae on liver tissue in rats. Hum. Exp. Toxicol. 2004, 23, 21–28. [Google Scholar] [CrossRef]
- Akdogan, M.; Ozguner, M.; Kocak, A.; Oncu, M.; Cicek, E. Effects of peppermint teas on plasma testosterone, follicle-stimulating hormone, and luteinizing hormone levels and testicular tissue in rats. Urology 2004, 64, 394–398. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
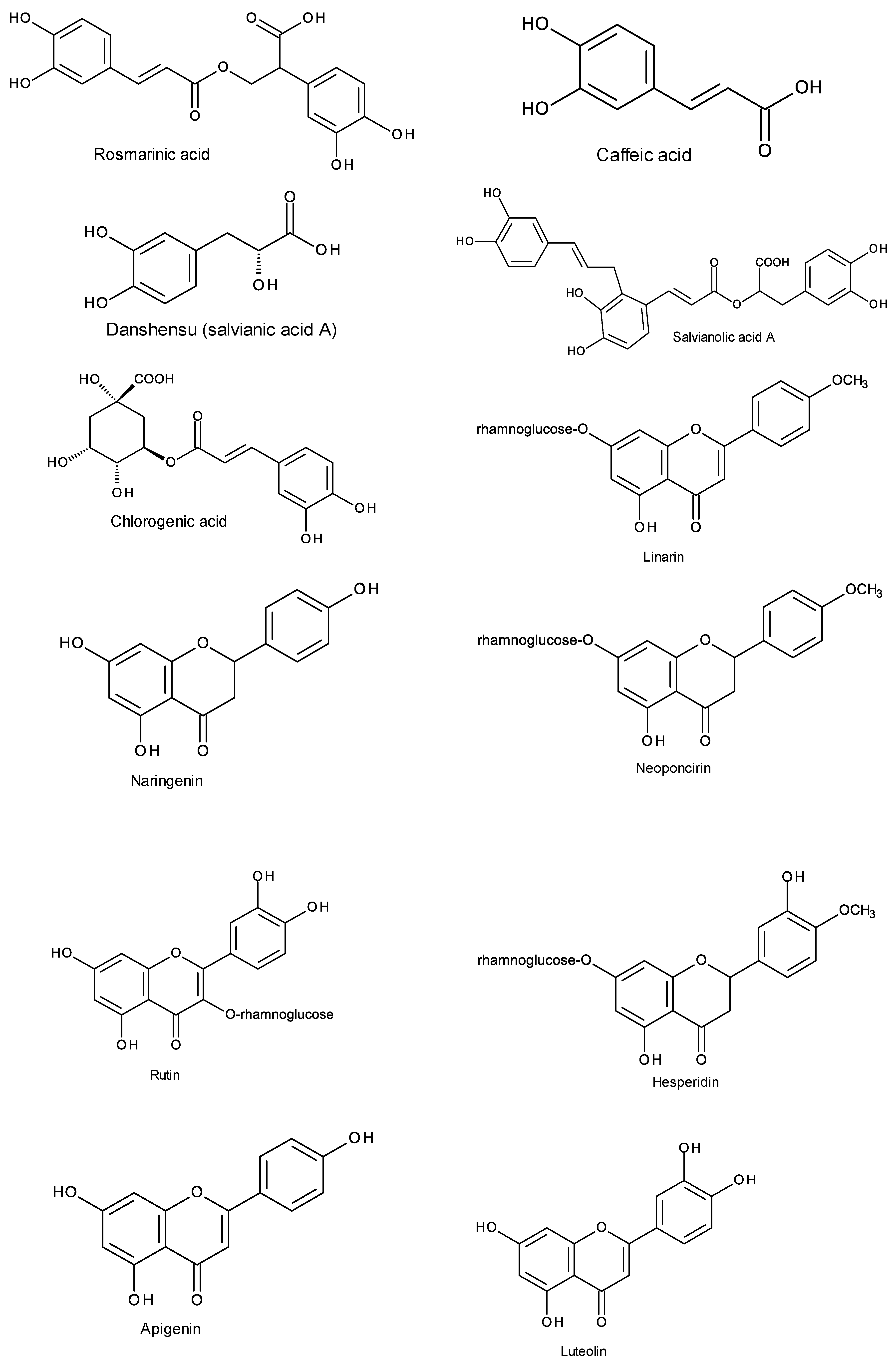{kind=link}
| Plant 1 | Extraction Solvent | Chemical Composition | References |
|---|---|---|---|
| M. aquatica | 50% aqueous methanol | danshensu, coumaroylquinic acid, caffeic acid, rosmarinic acid, salvianolic acid J, luteolin-7-O-diglucuronide, eriocitrin, apigenin-7-O-diglucuronide, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, apigenin-7-O-rutinoside, hesperidin, apigenin-7-O-glucuronide | [74] |
| 70% aqueous ethanol | naringenin | [70] | |
| M. arvensis | 50% aqueous methanol | chlorogenic acid, salvianolic acid I/J, rosmarinic acid, hesperidin | [74] |
| 80% aqueous methanol | linarin | [101] | |
| M. australis | 50% aqueous methanol | danshensu, caftaric acid, chlorogenic acid, lithospermic acid, rosmarinic acid, narirutin, isosakuranetin, linarin, neoponcirin | [74] |
| M. diemenica | 50% aqueous methanol | danshensu, lithospermic acid, salvianolic acid A/H/I, rosmarinic acid, luteolin-7-O-diglucuronide, luteolin-7-O-glucuronide, apigenin-7-O-rutinoside, linarin | [74] |
| M. gentilis | 50% aqueous methanol | rosmarinic acid, luteolin-7-O-rutinoside, apigenin-7-O-rutinoside, hesperidin, neoponcirin, acacetin-7-O-(6″-O-acetyl)-glucosylrhamnosylglucoside, linarin | [74] |
| M. longifolia | Ethanol | rosmarinic acid, chlorogenic acid, caffeic acid | [54] |
| Mentha longifolia var. calliantha | Ethanol or water | gallic acid, protocatechuic acid, p-hydroxybenzoic acid, chlorogenic acid, caffeic acid, syringic acid, vanillin, p-coumaric acid, sinapic acid, o-coumaric acid, rutin | [93] |
| M. pulegium | 50% aqueous methanol | danshensu, caftaric acid, caffeic acid, chicoric acid, salvianolic acid B isomer, rosmarinic acid, salvianolic acid C, luteolin-7-O-diglucuronide, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, apigenin-7-O-glucuronide | [74] |
| Ethanol | rosmarinic acid, chlorogenic acid, caffeic acid | [54] | |
| M. requienii | 50% aqueous methanol | danshensu, caftaric acid, chlorogenic acid, caffeic acid, chicoric acid, salvianolic acid B isomer, rosmarinic acid, salvianolic acid C, rutin | [74] |
| M. spicata | 50% aqueous methanol | danshensu, chlorogenic acid, caffeic acid, rosmarinic acid, salvianolic acid B/C, luteolin-7-O-diglucuronide, eriocitrin, apigenin-7-O-diglucuronide, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, narirutin, apigenin-7-O-glucuronide, neoponcirin | [74] |
| M. spicata var. crispa | 50% aqueous methanol | danshensu, caftaric acid, chlorogenic acid, caffeic acid, rosmarinic acid, salvianolic acid J, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, apigenin-7-O-rutinoside, hesperidin, neoponcirin | [74] |
| M. suaveolens | 50% aqueous methanol | danshensu, coumaroylquinic acid, chlorogenic acid, caffeic acid, rosmarinic acid, salvianolic acid B/J, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, hesperidin | [74] |
| M. suaveolens var. variegate | 50% aqueous methanol | danshensu, coumaroylquinic acid, chlorogenic acid, caffeic acid, rosmarinic acid, salvianolic acid A, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide | [74] |
| M. viridis | 50% aqueous methanol | danshensu, caffeic acid, salvianolic acid F/J, rosmarinic acid, eriocitrin, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, narirutin, apigenin-7-O-rutinoside, hesperidin, neoponcirin | [74] |
| M. × niliaca | 50% aqueous methanol | danshensu, rosmarinic acid, salvianolic acid B/E, eriocitrin, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, narirutin, apigenin-7-O-rutinoside, hesperidin, neoponcirin | [74] |
| M. × piperita | 50% aqueous methanol | danshensu, caftaric acid, chlorogenic acid, caffeic acid, salvianolic acid F/J, rosmarinic acid, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, narirutin, hesperidin, neoponcirin | [74] |
| Ethanol | rosmarinic acid, chlorogenic acid, caffeic acid, ferulic acid | [54] | |
| 70% aqueous methanol | luteolin diglucuronide, eriocitrin, luteolin rutinoside, salvianolic acid H/I/J, luteolin-7-O-glucuronide | [91] | |
| M. × piperita var. citrata | 50% aqueous methanol | rosmarinic acid, salvianolic acid J, apigenin-7-O-diglucuronide, eriocitrin, luteolin-7-O-diglucuronide, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, apigenin-7-O-rutinoside, narirutin, apigenin-7-O-glucuronide, hesperidin | [74] |
| M. × piperita var. officinalis | 50% aqueous methanol | danshensu, coumaroylquinic acid, chlorogenic acid, salvianolic acid B/F, rosmarinic acid, luteolin-7-O-diglucuronide, luteolin-7-O-rutinoside, narirutin, linarin, hesperidin, apigenin-7-O-rutinoside, neoponcirin | [74] |
| M. × piperita f. citrata ʻChocolate’ | 50% aqueous methanol | chlorogenic acid, caffeic acid, rosmarinic acid, salvianolic acid B, luteolin-7-O-diglucuronide, eriocitrin, luteolin-7-O-rutinoside, apigenin-7-O-diglucuronide, apigenin-7-O-glucuronide, luteolin-7-O-glucuronide, apigenin-7-O-rutinoside, hesperidin | [74] |
| M. × piperita f. citrata ʻBasil’ | 50% aqueous methanol | danshensu, rosmarinic acid, salvianolic acid B, luteolin-7-O-diglucuronide, eriocitrin, apigenin-7-O-diglucuronide, luteolin-7-O-rutinoside, luteolin-7-O-glucuronide, apigenin-7-O-rutinoside, hesperidin, apigenin-7-O-glucuronide, neoponcirin | [74] |
| Extraction/Drying Method | Advantages | Disadvantages | References |
|---|---|---|---|
| conventional Soxhlet extraction | ↑ crude extract yield Simple | ↓ flavonoids at ↓ concentration time and solvent consuming not suitable for thermo-sensitive compounds | [125] |
| supercritical carbon dioxide extraction | ↑ flavonoids at ↑ concentration extract selective soluble components | ↓ crude extract yield CO2 is not suitable for extracting polar compounds | [125] |
| accelerated solvent extractor | ↑ TPC 3 | [77] | |
| hydrodistillation extraction | ↑ TPC | [83] | |
| ultrasonic methanol extraction | ↑ TPC | methanol is highly restricted in food or preservative industry | [124] |
| Soxhlet 70% ethanol extraction | ↑ TPC | [124] | |
| extraction at 60 °C | ↑ TPC | [119] | |
| 70% acetone extraction | ↑ TPC | [126] | |
| ultrasonic 70% ethanol extraction | ↓ TPC | [124] | |
| 70% ethanol extraction | ↓ TPC | [126] | |
| 70% methanol extraction | ↓ TPC | [126] | |
| water extraction | ↓ TPC | [126] | |
| freeze-drying 5 min. at 60 °C | ↑ TPC | ↑ cost and time consuming | [129] |
| microwave oven drying | ↑ TPC shortens drying time | [127,128] | |
| direct sunlight drying at 20–30 °C | ↑ TPC | samples get polluted, time consuming | [127] |
| oven drying at 50 °C | ↓ TPC | not suitable for thermo-sensitive compounds | [127] |
| oven drying at 80 °C | ↓ TPC | time consuming | [128] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanafy, D.M.; Burrows, G.E.; Prenzler, P.D.; Hill, R.A. Potential Role of Phenolic Extracts of Mentha in Managing Oxidative Stress and Alzheimer’s Disease. Antioxidants 2020, 9, 631. https://doi.org/10.3390/antiox9070631
Hanafy DM, Burrows GE, Prenzler PD, Hill RA. Potential Role of Phenolic Extracts of Mentha in Managing Oxidative Stress and Alzheimer’s Disease. Antioxidants. 2020; 9(7):631. https://doi.org/10.3390/antiox9070631
Chicago/Turabian StyleHanafy, Doaa M., Geoffrey E. Burrows, Paul D. Prenzler, and Rodney A. Hill. 2020. "Potential Role of Phenolic Extracts of Mentha in Managing Oxidative Stress and Alzheimer’s Disease" Antioxidants 9, no. 7: 631. https://doi.org/10.3390/antiox9070631
APA StyleHanafy, D. M., Burrows, G. E., Prenzler, P. D., & Hill, R. A. (2020). Potential Role of Phenolic Extracts of Mentha in Managing Oxidative Stress and Alzheimer’s Disease. Antioxidants, 9(7), 631. https://doi.org/10.3390/antiox9070631