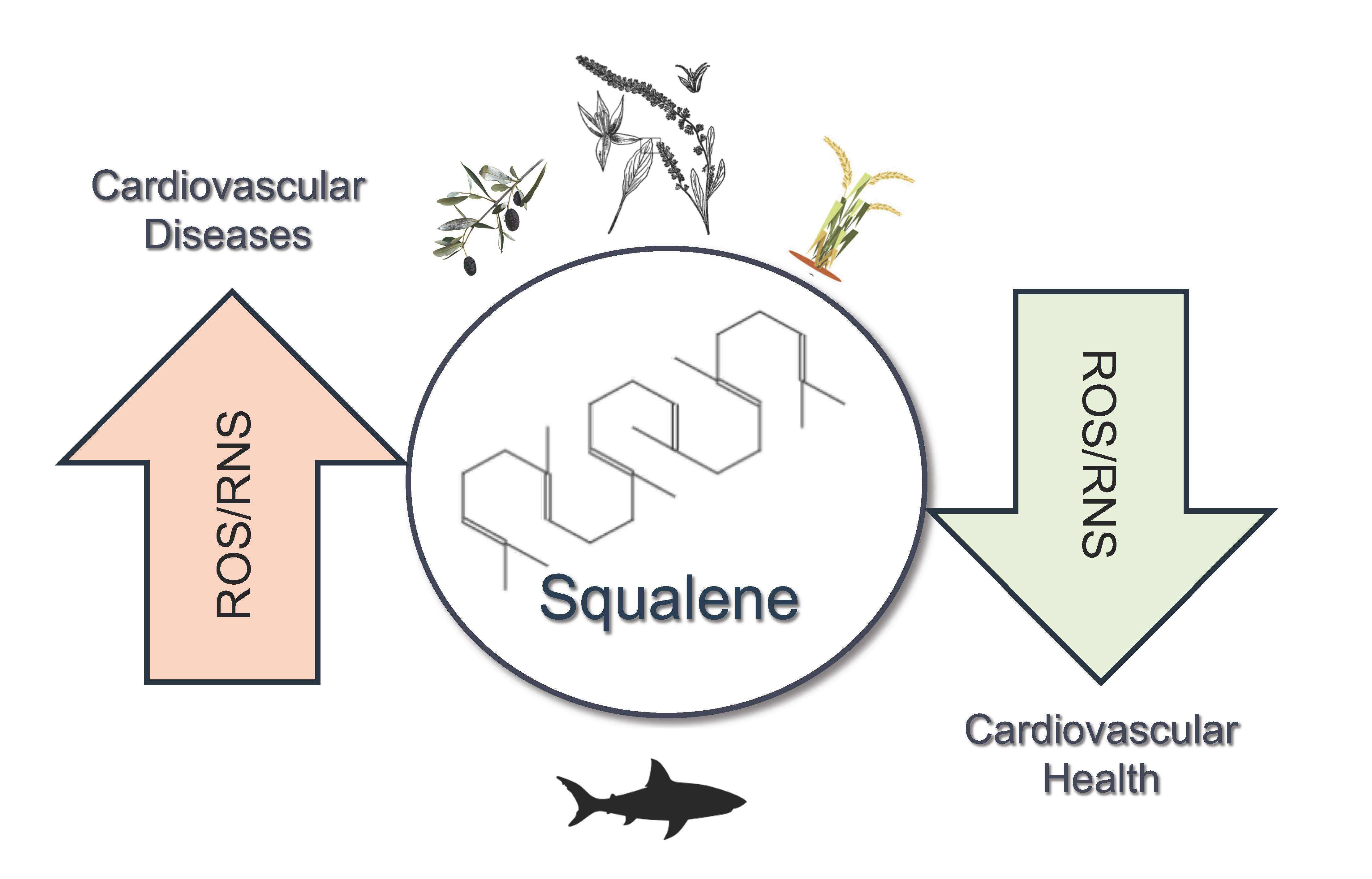
Squalene: More than a Step toward Sterols


 , ,
, ,
Abstract
:
1. Introduction
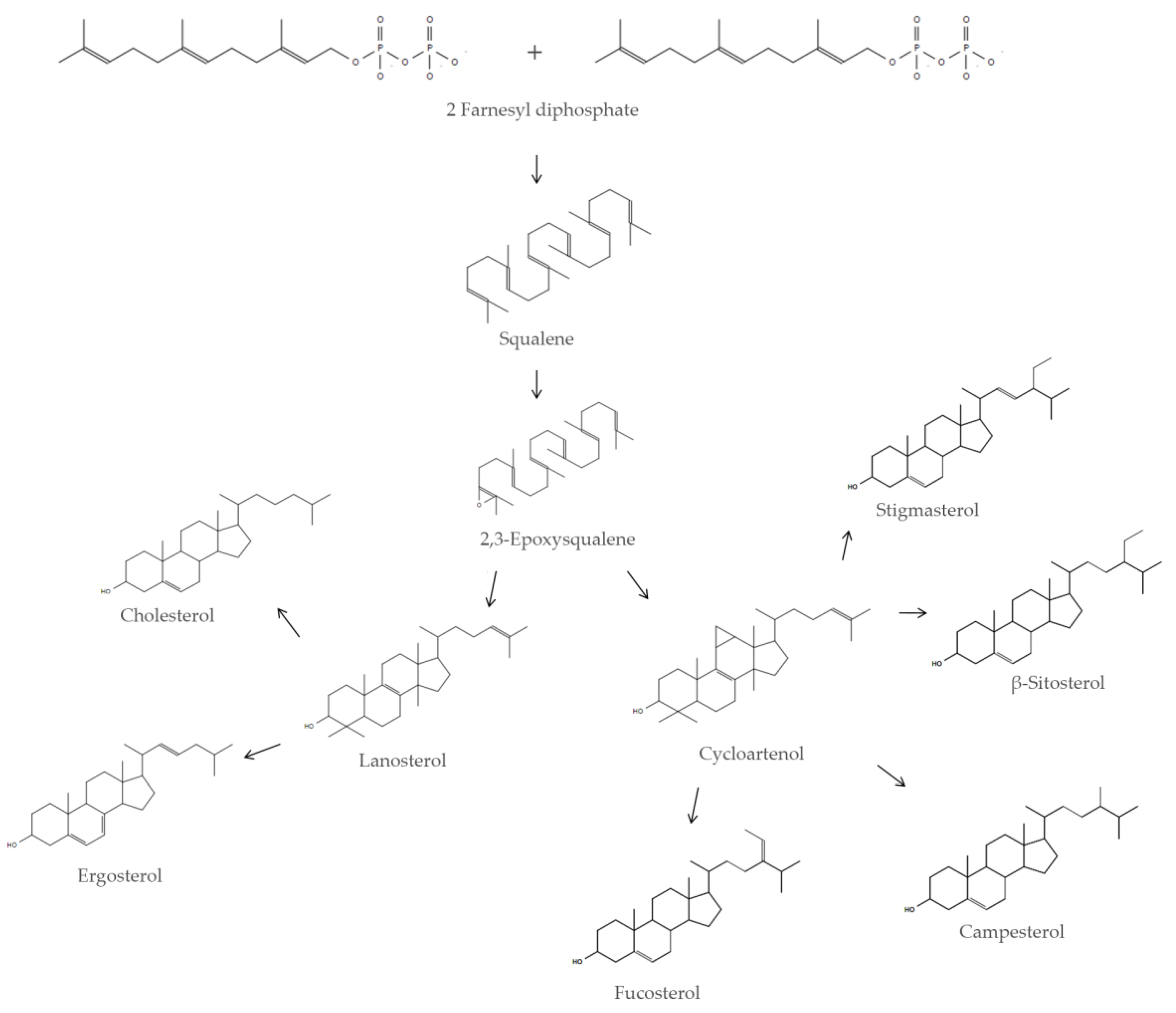
2. Sources of Squalene
2.1. Endogenous Sources of Squalene
2.2. Exogenous Sources of Squalene
3. ROS/RNS and Squalene
3.1. ROS/RNS and Squalene
3.2. Oxidative Stress in Skin and Squalene
3.3. Cardiovascular Antioxidant Systems and Squalene
4. Oxidative Stress Associated CVDs and Squalene
4.1. Hypercholesterolemia
4.2. Hypertension
4.3. Hyperglycemia-Dependent Endothelial Dysfunction
5. Pathological Implications of Squalene
5.1. Coronary Artery Disease
5.2. Metabolic Syndrome
5.3. Squalene Synthase Inhibitors
6. Nutraceutical Properties of SQ
7. Squalene: Healthy or Harmful in CVDs? Summary and Future Directions
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, S.-K.; Karadeniz, F. Chapter 14—Biological Importance and Applications of Squalene and Squalane. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Marine Medicinal Foods; Academic Press: Cambridge, MA, USA, 2012; Volume 65, pp. 223–233. [Google Scholar]
- Bhilwade, H.N.; Tatewaki, N.; Nishida, H.; Konishi, T. Squalene as novel food factor. Curr. Pharm. Biotechnol. 2010, 11, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, M. A highly unsaturated hydrocarbon in shark liver oil. J. Ind. Eng. Chem. 1916, 8, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Reddy, L.H.; Couvreur, P. Squalene: A natural triterpene for use in disease management and therapy. Adv. Drug Deliv. Rev. 2009, 61, 1412–1426. [Google Scholar] [CrossRef] [PubMed]
- Rezen, T.; Debeljak, N.; Kordis, D.; Rozman, D. New aspects on lanosterol 14alpha-demethylase and cytochrome P450 evolution: Lanosterol/cycloartenol diversification and lateral transfer. J. Mol. Evol. 2004, 59, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Tansey, T.R.; Shechter, I. Structure and regulation of mammalian squalene synthase. Biochimica et Biophysica Acta (BBA)—Mol. Cell Biol. Lipids 2000, 1529, 49–62. [Google Scholar] [CrossRef]
- Spanova, M.; Daum, G. Squalene—Biochemistry, molecular biology, process biotechnology, and applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1299–1320. [Google Scholar] [CrossRef]
- Kohno, Y.; Egawa, Y.; Itoh, S.; Nagaoka, S.; Takahashi, M.; Mukai, K. Kinetic study of quenching reaction of singlet oxygen and scavenging reaction of free radical by squalene in n-butanol. Biochim. Biophys. Acta 1995, 1256, 52–56. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Woods, J.A.; O’Brien, N.M. Squalene but not n-3 fatty acids protect against hydrogen peroxide-induced sister chromatid exchanges in Chinese hamster V79 cells. Nutr. Res. 2002, 22, 847–857. [Google Scholar] [CrossRef]
- Ott, D.B.; Lachance, P.A. Biochemical controls of liver cholesterol biosynthesis. Am. J. Clin. Nutr. 1981, 34, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Popa, I.; Băbeanu, N.; Niță, S.; Popa, O. Squalene-Natural resources and applications. Farmacia 2014, 62, 840–862. [Google Scholar]
- Saudek, C.D.; Frier, B.M.; Liu, G.C. Plasma squalene: Lipoprotein distribution and kinetic analysis. J. Lipid Res. 1978, 19, 827–835. [Google Scholar] [PubMed]
- Liu, G.C.; Ahrens, E.H.; Schreibman, P.H.; Crouse, J.R. Measurement of squalene in human tissues and plasma: Validation and application. J. Lipid Res. 1976, 17, 38–45. [Google Scholar] [PubMed]
- Shimizu, N.; Ito, J.; Kato, S.; Otoki, Y.; Goto, M.; Eitsuka, T.; Miyazawa, T.; Nakagawa, K. Oxidation of squalene by singlet oxygen and free radicals results in different compositions of squalene monohydroperoxide isomers. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.-R.; Lin, Y.-K.; Fang, J.-Y. Biological and pharmacological activities of squalene and related compounds: Potential uses in cosmetic dermatology. Molecules 2009, 14, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Gleiss, A.C.; Potvin, J.; Goldbogen, J.A. Physical trade-offs shape the evolution of buoyancy control in sharks. Proc. Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraujalis, P.; Venskutonis, P.R. Supercritical carbon dioxide extraction of squalene and tocopherols from amaranth and assessment of extracts antioxidant activity. J. Supercrit. Fluids 2013, 80, 78–85. [Google Scholar] [CrossRef]
- Wejnerowska, G.; Heinrich, P.; Gaca, J. Separation of squalene and oil from Amaranthus seeds by supercritical carbon dioxide. Sep. Purif. Technol. 2013, 110, 39–43. [Google Scholar] [CrossRef]
- De Vita, D.; Messore, A.; Toniolo, C.; Frezza, C.; Scipione, L.; Bertea, C.M.; Micera, M.; Di Sarno, V.; Madia, V.N.; Pindinello, I.; et al. Towards a new application of amaranth seed oil as an agent against Candida albicans. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef]
- Tikekar, R.V.; Ludescher, R.D.; Karwe, M.V. Processing stability of squalene in amaranth and antioxidant potential of amaranth extract. J. Agric. Food Chem. 2008, 56, 10675–10678. [Google Scholar] [CrossRef]
- Relas, H.; Gylling, H.; Miettinen, T.A. Dietary squalene increases cholesterol synthesis measured with serum non-cholesterol sterols after a single oral dose in humans. Atherosclerosis 2000, 152, 377–383. [Google Scholar] [CrossRef]
- Strandberg, T.E.; Tilvis, R.S.; Miettinen, T.A. Metabolic variables of cholesterol during squalene feeding in humans: Comparison with cholestyramine treatment. J. Lipid Res. 1990, 31, 1637–1643. [Google Scholar] [PubMed]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Food Chemistry, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-69933-0. [Google Scholar]
- Yardımcı, Ş.D.; Kaya, N.; Balci, M. Regioselectivity in the ene-reaction of singlet oxygen with cyclic alkenes: Photooxygenation of methyl-substituted 1,4-cyclohexadiene derivatives. Tetrahedron 2006, 62, 10633–10638. [Google Scholar] [CrossRef]
- Krasnovsky, A.A.; Kagan, V.E.; Minin, A.A. Quenching of singlet oxygen luminescence by fatty acids and lipids: Contribution of physical and chemical mechanisms. FEBS Lett. 1983, 155, 233–236. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Grande, M.A.; Gorinstein, S.; Espitia-Rangel, E.; Dávila-Ortiz, G.; Martínez-Ayala, A.L. Plant Sources, Extraction Methods, and Uses of Squalene. Available online: https://www.hindawi.com/journals/ija/2018/1829160/ (accessed on 12 March 2020).
- Nathan, J.A. Squalene and cholesterol in the balance at the ER membrane. Proc. Natl. Acad. Sci. USA 2020, 117, 8228–8230. [Google Scholar] [CrossRef] [PubMed]
- Aioi, A.; Shimizu, T.; Kuriyama, K. Effect of squalene on superoxide anion generation induced by a skin irritant, lauroylsarcosine. Int. J. Pharm. 1995, 113, 159–164. [Google Scholar] [CrossRef]
- Sabeena Farvin, K.H.; Anandan, R.; Kumar, S.H.S.; Shiny, K.S.; Sankar, T.V.; Thankappan, T.K. Effect of squalene on tissue defense system in isoproterenol-induced myocardial infarction in rats. Pharmacol. Res. 2004, 50, 231–236. [Google Scholar] [CrossRef]
- Ravi Kumar, S.; Narayan, B.; Sawada, Y.; Hosokawa, M.; Miyashita, K. Combined effect of astaxanthin and squalene on oxidative stress in vivo. Mol. Cell. Biochem. 2016, 417, 57–65. [Google Scholar] [CrossRef]
- Buddhan, S.; Sivakumar, R.; Dhandapani, N.; Ganesan, B.; Anandan, R. Protective effect of dietary squalene supplementation on mitochondrial function in liver of aged rats. Prostaglandins Leukot. Essent. Fatty Acids 2007, 76, 349–355. [Google Scholar] [CrossRef]
- Motawi, T.M.K.; Sadik, N.A.H.; Refaat, A. Cytoprotective effects of DL-alpha-lipoic acid or squalene on cyclophosphamide-induced oxidative injury: An experimental study on rat myocardium, testicles and urinary bladder. Food Chem. Toxicol. 2010, 48, 2326–2336. [Google Scholar] [CrossRef]
- Gabás Rivera, C.; Barranquero, C.; Martínez-Beamonte, R.; Navarro Ferrando, M.Á.; Surra, J.; Osada, J. Dietary Squalene Increases High Density Lipoprotein-Cholesterol and Paraoxonase 1 and Decreases Oxidative Stress in Mice. PLoS ONE 2014, 9, e104224. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Knock, G.A. NADPH oxidase in the vasculature: Expression, regulation and signalling pathways; role in normal cardiovascular physiology and its dysregulation in hypertension. Free Radic. Biol. Med. 2019, 145, 385–427. [Google Scholar] [CrossRef] [PubMed]
- Bretón-Romero, R.; González de Orduña, C.; Romero, N.; Sánchez-Gómez, F.J.; de Álvaro, C.; Porras, A.; Rodríguez-Pascual, F.; Laranjinha, J.; Radi, R.; Lamas, S. Critical role of hydrogen peroxide signaling in the sequential activation of p38 MAPK and eNOS in laminar shear stress. Free Radic. Biol. Med. 2012, 52, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Zhu, J.; Xiao, Q.; Zhang, L. Reactive oxygen species: Key regulators in vascular health and diseases. Br. J. Pharmacol. 2018, 175, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Luo, Z.; Pepe, A.E.; Margariti, A.; Zeng, L.; Xu, Q. Embryonic stem cell differentiation into smooth muscle cells is mediated by Nox4-produced H2O2. Am. J. Physiol. Cell Physiol. 2009, 296, C711–C723. [Google Scholar] [CrossRef] [Green Version]
- Rajaram, R.D.; Dissard, R.; Jaquet, V.; de Seigneux, S. Potential benefits and harms of NADPH oxidase type 4 in the kidneys and cardiovascular system. Nephrol. Dial. Transplant. 2019, 34, 567–576. [Google Scholar] [CrossRef]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [Green Version]
- Cervantes Gracia, K.; Llanas-Cornejo, D.; Husi, H. CVD and Oxidative Stress. J. Clin. Med. 2017, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The Vascular Endothelium and Human Diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- Endemann, D.H.; Schiffrin, E.L. Endothelial dysfunction. J. Am. Soc. Nephrol. 2004, 15, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Institute for Quality and Efficiency in Health Care (IQWiG). High Cholesterol: Overview; IQWiG: Cologne, Germany, 2017. [Google Scholar]
- Ibrahim, N.I.; Fairus, S.; Zulfarina, M.S.; Naina Mohamed, I. The Efficacy of Squalene in Cardiovascular Disease Risk-A Systematic Review. Nutrients 2020, 12, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khor, H.T.; Chieng, D.Y. Lipidaemic effects of tocotrienols, tocopherols and squalene: Studies in the hamster. Asia Pac. J. Clin. Nutr. 1997, 6, 36–40. [Google Scholar] [PubMed]
- Kritchevsky, D.; Moyer, A.W.; Tesar, W.C.; Logan, J.B.; Brown, R.A.; Richmond, G. Squalene feeding in experimental atherosclerosis. Circ. Res. 1954, 2, 340–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillén, N.; Acín, S.; Navarro, M.A.; Perona, J.S.; Arbonés-Mainar, J.M.; Arnal, C.; Sarría, A.J.; Surra, J.C.; Carnicer, R.; Orman, I.; et al. Squalene in a sex-dependent manner modulates atherosclerotic lesion which correlates with hepatic fat content in apoE-knockout male mice. Atherosclerosis 2008, 197, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Hien, H.T.M.; Ha, N.C.; Thom, L.T.; Hong, D.D. Squalene promotes cholesterol homeostasis in macrophage and hepatocyte cells via activation of liver X receptor (LXR) α and β. Biotechnol. Lett. 2017, 39, 1101–1107. [Google Scholar] [CrossRef]
- Hoang, T.M.H.; Nguyen, C.H.; Le, T.T.; Hoang, T.H.Q.; Ngo, T.H.T.; Hoang, T.L.A.; Dang, D.H. Squalene isolated from Schizochytrium mangrovei is a peroxisome proliferator-activated receptor-α agonist that regulates lipid metabolism in HepG2 cells. Biotechnol. Lett. 2016, 38, 1065–1071. [Google Scholar] [CrossRef]
- Granados-Principal, S.; Quiles, J.L.; Ramirez-Tortosa, C.L.; Ochoa-Herrera, J.; Perez-Lopez, P.; Pulido-Moran, M.; Ramirez-Tortosa, M.C. Squalene ameliorates atherosclerotic lesions through the reduction of CD36 scavenger receptor expression in macrophages. Mol. Nutr. Food Res. 2012, 56, 733–740. [Google Scholar] [CrossRef]
- Chan, P.; Tomlinson, B.; Lee, C.B.; Lee, Y.S. Effectiveness and safety of low-dose pravastatin and squalene, alone and in combination, in elderly patients with hypercholesterolemia. J. Clin. Pharmacol. 1996, 36, 422–427. [Google Scholar] [CrossRef]
- Shin, D.H.; Heo, H.J.; Lee, Y.J.; Kim, H.K. Amaranth squalene reduces serum and liver lipid levels in rats fed a cholesterol diet. Br. J. Biomed. Sci. 2004, 61, 11–14. [Google Scholar] [CrossRef]
- Pinheiro, L.C.; Oliveira-Paula, G.H. Sources and effects of oxidative stress in hypertension. Curr. Hypertens. Rev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, D.M.; Miroshnichenko, L.A.; Kulakova, S.N.; Pogojeva, A.V.; Zoloedov, V.I. Amaranth oil application for coronary heart disease and hypertension. Lipids Health Dis. 2007, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xu, X.; Bi, D.; Wang, X.; Zhang, X.; Dai, H.; Chen, S.; Zhang, W. Influence of squalene feeding on plasma leptin, testosterone & blood pressure in rats. Indian J. Med. Res. 2009, 129, 150–153. [Google Scholar] [PubMed]
- Fiorentino, T.V.; Prioletta, A.; Zuo, P.; Folli, F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5695–5703. [Google Scholar] [CrossRef] [PubMed]
- Widyawati, T.; Yusoff, N.A.; Asmawi, M.Z.; Ahmad, M. Antihyperglycemic Effect of Methanol Extract of Syzygium polyanthum (Wight.) Leaf in Streptozotocin-Induced Diabetic Rats. Nutrients 2015, 7, 7764–7780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, M.; Calzada, F.; Mendieta-Wejebe, J. Structure–Activity Relationship Study of Acyclic Terpenes in Blood Glucose Levels: Potential α-Glucosidase and Sodium Glucose Cotransporter (SGLT-1) Inhibitors. Molecules 2019, 24, 4020. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.E.; Jakulj, L.; Kastelein, J.J.P.; Stroes, E.S.G. LDL-C-lowering therapy: Current and future therapeutic targets. Curr. Cardiol. Rep. 2008, 10, 512–520. [Google Scholar] [CrossRef]
- Wierzbicki, A.S. New lipid-lowering agents. Expert Opin. Emerg. Drugs 2003, 8, 365–376. [Google Scholar] [CrossRef]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Independent association of serum squalene and noncholesterol sterols with coronary artery disease in postmenopausal women. J. Am. Coll. Cardiol. 2000, 35, 1185–1191. [Google Scholar] [CrossRef] [Green Version]
- Lupattelli, G.; De Vuono, S.; Mannarino, E. Patterns of cholesterol metabolism: Pathophysiological and therapeutic implications for dyslipidemias and the metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 620–627. [Google Scholar] [CrossRef]
- Peltola, P.; Pihlajamäki, J.; Koutnikova, H.; Ruotsalainen, E.; Salmenniemi, U.; Vauhkonen, I.; Kainulainen, S.; Gylling, H.; Miettinen, T.A.; Auwerx, J.; et al. Visceral obesity is associated with high levels of serum squalene. Obesity (Silver Spring) 2006, 14, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Kourounakis, A.P.; Katselou, M.G.; Matralis, A.N.; Ladopoulou, E.M.; Bavavea, E. Squalene synthase inhibitors: An update on the search for new antihyperlipidemic and antiatherosclerotic agents. Curr. Med. Chem. 2011, 18, 4418–4439. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chu, C.-L.; Chen, L.; Shui, D. Evaluation of potential inhibitors of squalene synthase based on virtual screening and in vitro studies. Comput. Biol. Chem. 2019, 80, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Wang, J.; Tian, J.; Tang, Y. Coronary Artery Disease: From Mechanism to Clinical Practice. Adv. Exp. Med. Biol. 2020, 1177, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Serum squalene in postmenopausal women without and with coronary artery disease. Atherosclerosis 1999, 146, 61–64. [Google Scholar] [CrossRef]
- Rajaratnam, R.A.; Gylling, H.; Miettinen, T.A. Impaired postprandial clearance of squalene and apolipoprotein B-48 in post-menopausal women with coronary artery disease. Clin. Sci. 1999, 97, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Gylling, H.; Hallikainen, M.; Rajaratnam, R.A.; Simonen, P.; Pihlajamäki, J.; Laakso, M.; Miettinen, T.A. The metabolism of plant sterols is disturbed in postmenopausal women with coronary artery disease. Metab. Clin. Exp. 2009, 58, 401–407. [Google Scholar] [CrossRef]
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428. [Google Scholar] [CrossRef]
- Samson, S.L.; Garber, A.J. Metabolic syndrome. Endocrinol. Metab. Clin. N. Am. 2014, 43, 1–23. [Google Scholar] [CrossRef]
- Das, L.; Bhaumik, E.; Raychaudhuri, U.; Chakraborty, R. Role of nutraceuticals in human health. J. Food Sci. Technol. 2012, 49, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Nasri, H.; Baradaran, A.; Shirzad, H.; Rafieian-Kopaei, M. New concepts in nutraceuticals as alternative for pharmaceuticals. Int. J. Prev. Med. 2014, 5, 1487–1499. [Google Scholar] [PubMed]
- Hamadate, N.; Matsumoto, Y.; Seto, K.; Yamamoto, T.; Yamaguchi, H.; Nakagawa, T.; Yamamoto, E.; Fukagawa, M.; Yazawa, K. Vascular effects and safety of supplementation with shark liver oil in middle-aged and elderly males. Exp. Ther. Med. 2015, 10, 641–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.J. Squalene: Potential chemopreventive agent. Expert Opin. Investig. Drugs 2000, 9, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.V.; Newmark, H.L.; Reddy, B.S. Chemopreventive effect of squalene on colon cancer. Carcinogenesis 1998, 19, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Maksimenko, A.; Dosio, F.; Mougin, J.; Ferrero, A.; Wack, S.; Reddy, L.H.; Weyn, A.-A.; Lepeltier, E.; Bourgaux, C.; Stella, B.; et al. A unique squalenoylated and nonpegylated doxorubicin nanomedicine with systemic long-circulating properties and anticancer activity. Proc. Natl. Acad. Sci. USA 2014, 111, E217–E226. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, S.; Devaki, T.; Manohar, B.M.; Babu, M.S. Effect of squalene on cyclophosphamide-induced toxicity. Clin. Chim. Acta 2006, 364, 335–342. [Google Scholar] [CrossRef]
- Narayan, B.H.; Tatewaki, N.; Giridharan, V.V.; Nishida, H.; Konishi, T. Modulation of doxorubicin-induced genotoxicity by squalene in Balb/c mice. Food Funct. 2010, 1, 174–179. [Google Scholar] [CrossRef]
- Cannon, M.V.; van Gilst, W.H.; de Boer, R.A. Emerging role of liver X receptors in cardiac pathophysiology and heart failure. Basic Res. Cardiol. 2016, 111, 3. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Study | Model | Results |
|---|---|---|
| Aioi A. et al., 1995 [28] | Keratinocytes from shaved dorsal skin of rats, measurements of O2− by reduction of equine ferricytochrome c after treatment with SQ and stressing agent | Inhibition of O2− production by addiction of 100 mg/mL SQ |
| Buddhan S. et al., 2006 [31] | Young and old male Wistar albino rats fed with a 2% SQ supplemented diet for 15 or 30 days | Inhibition of liver mitochondrial lipid peroxidation, increased levels of reduced glutathione and enhanced activities of glutathione peroxidase, glutathione S-transferase, superoxide dismutase and catalase |
| Motawi T. M. K. et al., 2010 [32] | Male Wistar albino rats were administered for 7 days with SQ (0.4 mL/rat) before and after CP treatment | Improvement of GPx activity and GSH levels in cardiac tissue |
| Gabás-Rivera C. et al., 2014 [33] | Wild-type, Apoa1- and Apoe- deficient C57BL/6J male mice fed with 1 g/kg SQ for 11 weeks | Decreased ROS in lipoproteins |
| Ravi Kumar S. et al., 2016 [30] | Male KK-Ay mice fed high fat/sucrose diets supplemented with 2% SQ in combination with astaxanthin for 4 weeks | Elevated mRNA expression of SOD1 and GPx1 in liver |
| Pathological Condition | Reference | Effects | |
|---|---|---|---|
| Positive effects of SQ | Hypercholesterolemia and atherosclerosis | Gabás-Rivera C. et al., 2014 [33] | Induction of paraoxonase-1 and reduction of ROS in lipoproteins in male mice fed with a SQ enriched diet |
| Kritchevsky D. et al., 1954 [48] | Only correlation between anti-atherosclerotic effect and SQ enriched diet | ||
| Guillén N. et al., 2007 [49] | Only correlation between reduced plasma cholesterol and triglycerides and SQ enriched diet in female mice ApoE-KO | ||
| Hien H.T.M. et al., 2017 [50] | Activation of LXR α and β in macrophages | ||
| Hoang T.M.H. et al., 2016 [51] | Activation of PPARα, induction of genes involved in lipid metabolism with hypolipidemic effect in HepG2 cells | ||
| Granados-Principal S. et al., 2012 [52] | Reduced expression of CD36 in monocytes and macrophages | ||
| Shin D.H. et al., 2004 [54] | Enhancement of cholesterol and biliary acid excretion in rats fed SQ enriched diet; inhibition of HMG-CoA reductase activity after i.p. injection of SQ in rats | ||
| Hamadate N. et al., 2015 [76] | Enhancement of arterial elasticity in middle-aged and elderly men fed with shark liver enriched diet | ||
| Hypertension | Martirosyan, D.M. et al., 2007 [56] | Reduction of systolic blood pressure after SQ-rich amaranth oil supplemented diets | |
| Liu Y. et al., 2009 [57] | Decrease of leptin levels and consequent reduction of plasma lipids and blood pressure | ||
| Hyperglycemia-induced endothelial dysfunction | Liu Y. et al., 2009 [57] | Lower glycemia in rats fed SQ supplemented diet | |
| Negative effects of SQ | Coronary Artery Disease | Rajaratnam R.A. et al., 2000 [63] | Only correlation between elevated plasma ratio of SQ to cholesterol and augmented risk for coronary artery disease in postmenopausal women |
| Metabolic syndrome | Peltola P. et al., 2006 [65] | Only correlation between high serum SQ levels and visceral obesity | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micera, M.; Botto, A.; Geddo, F.; Antoniotti, S.; Bertea, C.M.; Levi, R.; Gallo, M.P.; Querio, G. Squalene: More than a Step toward Sterols. Antioxidants 2020, 9, 688. https://doi.org/10.3390/antiox9080688
Micera M, Botto A, Geddo F, Antoniotti S, Bertea CM, Levi R, Gallo MP, Querio G. Squalene: More than a Step toward Sterols. Antioxidants. 2020; 9(8):688. https://doi.org/10.3390/antiox9080688
Chicago/Turabian StyleMicera, Marco, Alfonso Botto, Federica Geddo, Susanna Antoniotti, Cinzia Margherita Bertea, Renzo Levi, Maria Pia Gallo, and Giulia Querio. 2020. "Squalene: More than a Step toward Sterols" Antioxidants 9, no. 8: 688. https://doi.org/10.3390/antiox9080688
APA StyleMicera, M., Botto, A., Geddo, F., Antoniotti, S., Bertea, C. M., Levi, R., Gallo, M. P., & Querio, G. (2020). Squalene: More than a Step toward Sterols. Antioxidants, 9(8), 688. https://doi.org/10.3390/antiox9080688