
Immunity to Sda1 Protects against Infection by Sda1+ and Sda1− Serotypes of Group A Streptococcus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains and Culture Conditions
2.3. Polymerase Chain Reaction Amplification for the Detection of the Sda1 Gene
2.4. Cloning of Recombinant Protein
2.5. Western Blotting
2.6. Mouse Immunization and Challenge Experiment
2.7. Enzyme-Linked Immunosorbent Assay (ELISA) for Antibodies
2.8. Preparation of Rabbit Serum IgG against Sda1
2.9. Plasmid DNA Degradation Assay
2.10. NETs Degradation Assay
2.11. Statistical Analysis
3. Results
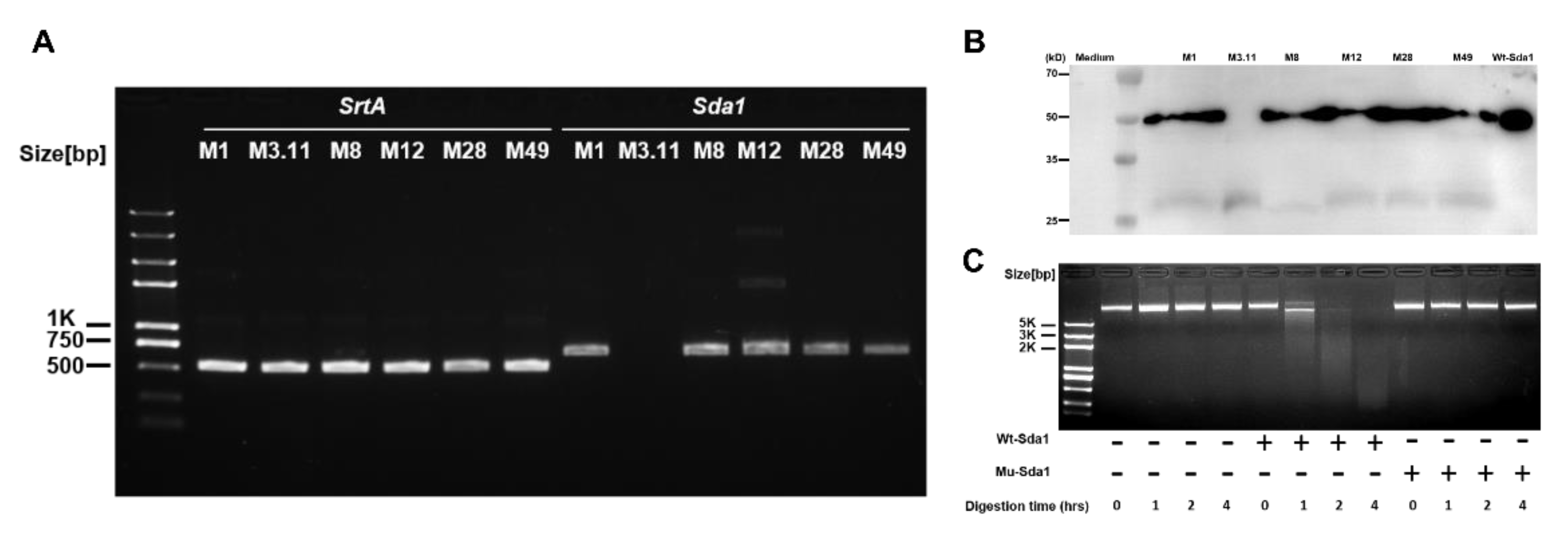
3.1. Sda1 Prevalence among GAS Isolates and Antibody Response to Sda1 in Mice and Humans
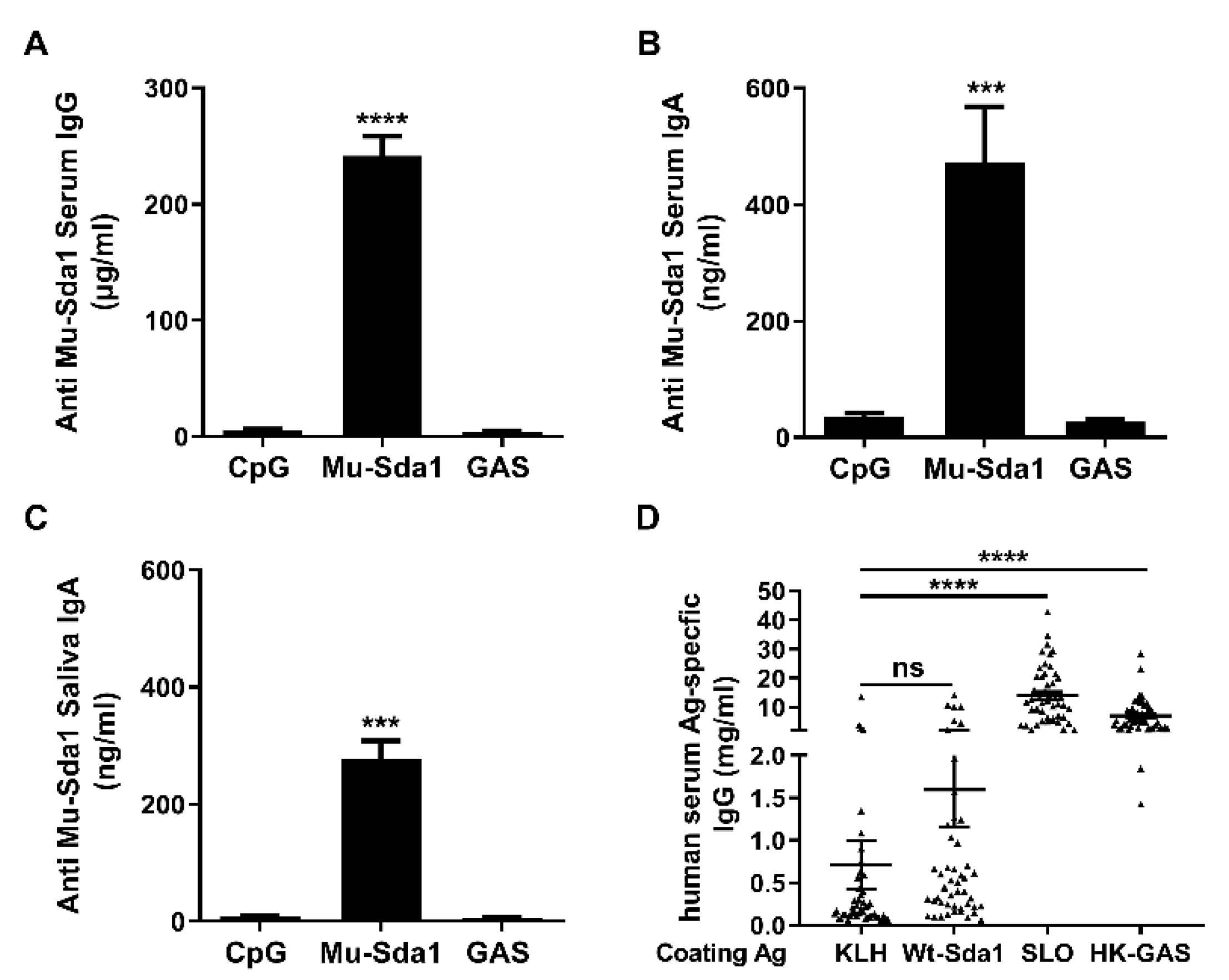
3.2. Antibody Response to Sda1 in Mice and Humans
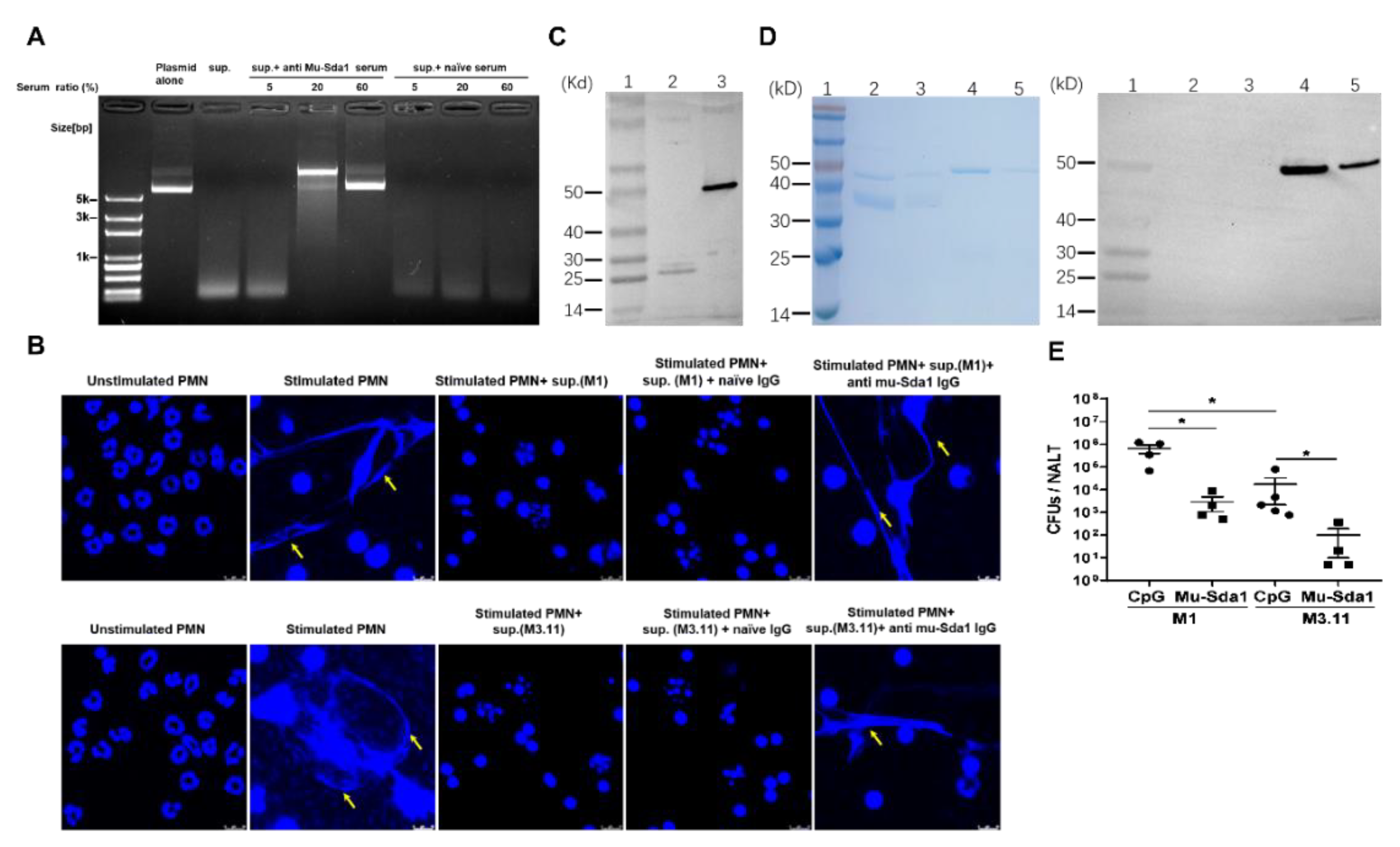
3.3. Anti-Sda1 Serum Neutralized Sda1-Inhibited Neutrophil NETosis
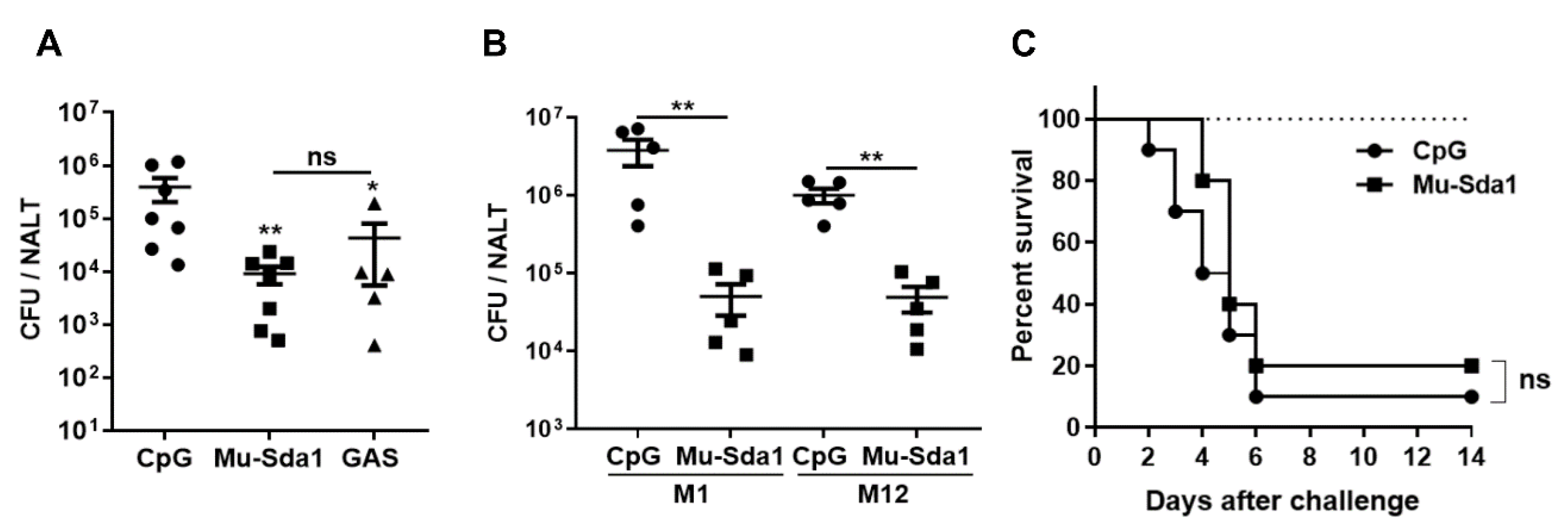
3.4. Immunization with Sda1 Prevented GAS Colonization in the Nasopharynx
3.5. Anti-Sda1 Prevented NETs Degradation and Mucosal Infection by the Sda1-Negative M3.11 Strain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanyahumbi, A.S.; Colquhoun, S.; Wyber, R.; Carapetis, J.R. Global Disease Burden of Group A Streptococcus. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease manifestations and pathogenic mechanisms of Group A Streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef] [Green Version]
- Henningham, A.; Barnett, T.C.; Maamary, P.G.; Walker, M.J. Pathogenesis of group A streptococcal infections. Discov. Med. 2012, 13, 329–342. [Google Scholar] [PubMed]
- Carapetis, J.R.; McDonald, M.; Wilson, N.J. Acute rheumatic fever. Lancet 2005, 366, 155–168. [Google Scholar] [CrossRef]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Porto, B.N.; Stein, R.T. Neutrophil Extracellular Traps in Pulmonary Diseases: Too Much of a Good Thing? Front. Immunol. 2016, 7, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumby, P.; Barbian, K.D.; Gardner, D.J.; Whitney, A.R.; Welty, D.M.; Long, R.D.; Bailey, J.R.; Parnell, M.J.; Hoe, N.P.; Adams, G.G.; et al. Extracellular deoxyribonuclease made by group A Streptococcus assists pathogenesis by enhancing evasion of the innate immune response. Proc. Natl. Acad. Sci. USA 2005, 102, 1679–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remmington, A.; Turner, C.E. The DNases of pathogenic Lancefield streptococci. Microbiology 2018, 164, 242–250. [Google Scholar] [CrossRef]
- Aziz, R.K.; Ismail, S.A.; Park, H.W.; Kotb, M. Post-proteomic identification of a novel phage-encoded streptodornase, Sda1, in invasive M1T1 Streptococcus pyogenes. Mol. Microbiol. 2004, 54, 184–197. [Google Scholar] [CrossRef]
- Buchanan, J.T.; Simpson, A.J.; Aziz, R.K.; Liu, G.Y.; Kristian, S.A.; Kotb, M.; Feramisco, J.; Nizet, V. DNase expression allows the pathogen group A Streptococcus to escape killing in neutrophil extracellular traps. Curr. Biol. 2006, 16, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.J.; Hollands, A.; Sanderson-Smith, M.L.; Cole, J.N.; Kirk, J.K.; Henningham, A.; McArthur, J.D.; Dinkla, K.; Aziz, R.K.; Kansal, R.G.; et al. DNase Sda1 provides selection pressure for a switch to invasive group A streptococcal infection. Nat. Med. 2007, 13, 981–985. [Google Scholar] [CrossRef]
- Uchiyama, S.; Andreoni, F.; Schuepbach, R.A.; Nizet, V.; Zinkernagel, A.S. DNase Sda1 allows invasive M1T1 Group A Streptococcus to prevent TLR9-dependent recognition. PLoS Pathog. 2012, 8, e1002736. [Google Scholar] [CrossRef]
- Keller, N.; Woytschak, J.; Heeb, L.E.M.; Marques Maggio, E.; Mairpady Shambat, S.; Snall, J.; Hyldegaard, O.; Boyman, O.; Norrby-Teglund, A.; Zinkernagel, A.S. Group A Streptococcal DNase Sda1 Impairs Plasmacytoid Dendritic Cells’ Type 1 Interferon Response. J. Investig. Dermatol. 2019, 139, 1284–1293. [Google Scholar] [CrossRef]
- Maamary, P.G.; Ben Zakour, N.L.; Cole, J.N.; Hollands, A.; Aziz, R.K.; Barnett, T.C.; Cork, A.J.; Henningham, A.; Sanderson-Smith, M.; McArthur, J.D.; et al. Tracing the evolutionary history of the pandemic group A streptococcal M1T1 clone. FASEB J. 2012, 26, 4675–4684. [Google Scholar] [CrossRef] [Green Version]
- Moon, A.F.; Krahn, J.M.; Lu, X.; Cuneo, M.J.; Pedersen, L.C. Structural characterization of the virulence factor Sda1 nuclease from Streptococcus pyogenes. Nucleic Acids Res. 2016, 44, 3946–3957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarnutzer, A.; Andreoni, F.; Keller, N.; Zurcher, C.; Norrby-Teglund, A.; Schupbach, R.A.; Zinkernagel, A.S. Human polyspecific immunoglobulin attenuates group A streptococcal virulence factor activity and reduces disease severity in a murine necrotizing fasciitis model. Clin. Microbiol. Infect. 2019, 25, 512-e7. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, N.; Bi, S.; Wang, X.; Wang, B. Co-Activation of Th17 and Antibody Responses Provides Efficient Protection against Mucosal Infection by Group A Streptococcus. PLoS ONE 2016, 11, e0168861. [Google Scholar] [CrossRef] [Green Version]
- Bi, S.; Xu, M.; Zhou, Y.; Xing, X.; Shen, A.; Wang, B. A Multicomponent Vaccine Provides Immunity against Local and Systemic Infections by Group A Streptococcus across Serotypes. mBio 2019, 10, e02600-19. [Google Scholar] [CrossRef] [Green Version]
- Vlaminckx, B.J.; Schuren, F.H.; Montijn, R.C.; Caspers, M.P.; Beitsma, M.M.; Wannet, W.J.; Schouls, L.M.; Verhoef, J.; Jansen, W.T. Dynamics in prophage content of invasive and noninvasive M1 and M28 Streptococcus pyogenes isolates in The Netherlands from 1959 to 1996. Infect. Immun. 2007, 75, 3673–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Race, P.R.; Bentley, M.L.; Melvin, J.A.; Crow, A.; Hughes, R.K.; Smith, W.D.; Sessions, R.B.; Kehoe, M.A.; McCafferty, D.G.; Banfield, M.J. Crystal structure of Streptococcus pyogenes sortase A: Implications for sortase mechanism. J. Biol. Chem. 2009, 284, 6924–6933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Wang, X.; Li, N.; Cui, H.; Hou, B.; Gao, B.; Cleary, P.P.; Wang, B. Sortase A induces Th17-mediated and antibody-independent immunity to heterologous serotypes of group A streptococci. PLoS ONE 2014, 9, e107638. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Francis, K.P.; Yu, J.; Cleary, P.P. Membranous cells in nasal-associated lymphoid tissue: A portal of entry for the respiratory mucosal pathogen group A streptococcus. J. Immunol. 2003, 171, 2532–2537. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Edwards, R.A.; Taylor, W.W.; Low, D.E.; McGeer, A.; Kotb, M. Mosaic prophages with horizontally acquired genes account for the emergence and diversification of the globally disseminated M1T1 clone of Streptococcus pyogenes. J. Bacteriol. 2005, 187, 3311–3318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, M.R.; Holden, M.T.; Coupland, P.; Chen, J.H.; Venturini, C.; Barnett, T.C.; Zakour, N.L.; Tse, H.; Dougan, G.; Yuen, K.Y.; et al. Emergence of scarlet fever Streptococcus pyogenes emm12 clones in Hong Kong is associated with toxin acquisition and multidrug resistance. Nat. Genet. 2015, 47, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Limbago, B.; Penumalli, V.; Weinrick, B.; Scott, J.R. Role of streptolysin O in a mouse model of invasive group A streptococcal disease. Infect. Immun. 2000, 68, 6384–6390. [Google Scholar] [CrossRef] [PubMed]
- Sriskandan, S.; Unnikrishnan, M.; Krausz, T.; Cohen, J. Mitogenic factor (MF) is the major DNase of serotype M89 Streptococcus pyogenes. Microbiology 2000, 146 Pt 11, 2785–2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, M.; Sakae, K.; Hashikawa, S.; Torii, K.; Hasegawa, T.; Horii, T.; Endo, M.; Okuno, R.; Murayama, S.; Hirasawa, K.; et al. Close correlation of streptococcal DNase B (sdaB) alleles with emm genotypes in Streptococcus pyogenes. Microbiol. Immunol. 2005, 49, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.H.; Lee, D.W.; Ryu, R.A.; Lee, Y.S.; Lee, S.H.; Kang, J.O.; Tae, K. Bacteriologic comparison of tonsil core in recurrent tonsillitis and tonsillar hypertrophy. Laryngoscope 2007, 117, 2146–2151. [Google Scholar] [CrossRef]
- Chang, A.; Khemlani, A.; Kang, H.; Proft, T. Functional analysis of Streptococcus pyogenes nuclease A (SpnA), a novel group A streptococcal virulence factor. Mol. Microbiol. 2011, 79, 1629–1642. [Google Scholar] [CrossRef]
- Cleary, P.P.; Prahbu, U.; Dale, J.B.; Wexler, D.E.; Handley, J. Streptococcal C5a peptidase is a highly specific endopeptidase. Infect. Immun. 1992, 60, 5219–5223. [Google Scholar] [CrossRef] [Green Version]
- Park, H.S.; Cleary, P.P. Active and passive intranasal immunizations with streptococcal surface protein C5a peptidase prevent infection of murine nasal mucosa-associated lymphoid tissue, a functional homologue of human tonsils. Infect. Immun. 2005, 73, 7878–7886. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Hernandez, T.; Pandey, M.; Henningham, A.; Cole, J.; Choudhury, B.; Cork, A.J.; Gillen, C.M.; Ghaffar, K.A.; West, N.P.; Silvestri, G.; et al. Differing Efficacies of Lead Group A Streptococcal Vaccine Candidates and Full-Length M Protein in Cutaneous and Invasive Disease Models. mBio 2016, 7, e00618-16. [Google Scholar] [CrossRef] [Green Version]
- Venturini, C.; Ong, C.L.; Gillen, C.M.; Ben-Zakour, N.L.; Maamary, P.G.; Nizet, V.; Beatson, S.A.; Walker, M.J. Acquisition of the Sda1-encoding bacteriophage does not enhance virulence of the serotype M1 Streptococcus pyogenes strain SF370. Infect. Immun. 2013, 81, 2062–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reglinski, M.; Lynskey, N.N.; Choi, Y.J.; Edwards, R.J.; Sriskandan, S. Development of a multicomponent vaccine for Streptococcus pyogenes based on the antigenic targets of IVIG. J. Infect. 2016, 72, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, C.F.; Tsao, N.; Hsieh, I.C.; Lin, Y.S.; Wu, J.J.; Hung, Y.T. Immunization with a streptococcal multiple-epitope recombinant protein protects mice against invasive group A streptococcal infection. PLoS ONE 2017, 12, e0174464. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, S.; Wang, J.; Xu, M.; Li, N.; Wang, B. Immunity to Sda1 Protects against Infection by Sda1+ and Sda1− Serotypes of Group A Streptococcus. Vaccines 2022, 10, 102. https://doi.org/10.3390/vaccines10010102
Bi S, Wang J, Xu M, Li N, Wang B. Immunity to Sda1 Protects against Infection by Sda1+ and Sda1− Serotypes of Group A Streptococcus. Vaccines. 2022; 10(1):102. https://doi.org/10.3390/vaccines10010102
Chicago/Turabian StyleBi, Shuai, Jie Wang, Meiyi Xu, Ning Li, and Beinan Wang. 2022. "Immunity to Sda1 Protects against Infection by Sda1+ and Sda1− Serotypes of Group A Streptococcus" Vaccines 10, no. 1: 102. https://doi.org/10.3390/vaccines10010102
APA StyleBi, S., Wang, J., Xu, M., Li, N., & Wang, B. (2022). Immunity to Sda1 Protects against Infection by Sda1+ and Sda1− Serotypes of Group A Streptococcus. Vaccines, 10(1), 102. https://doi.org/10.3390/vaccines10010102