

Differential Cell Line Susceptibility to the SARS-CoV-2 Omicron BA.1.1 Variant of Concern

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Propagation and Titration
2.2. Cell Culture Systems
2.3. Virus Infectivity Assay
2.4. RNA Extraction and Real-Time Reverse-Transcription PCR
2.5. Statistical Analysis
3. Results
3.1. Virus Propagation and Titration
3.2. Determination of Virus Infectivity Based on Cytopathic Effect
3.3. Determination of Virus Infectivity Based on Viral Titer
3.4. Determination of Virus Infectivity Based on the Viral Load
3.5. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses Nature Microbiology. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. COVID Dashboard. Available online: https://covid19.who.int/ason16.09.2022 (accessed on 16 September 2022).
- Romano, M.; Ruggiero, A.; Squeglia, F.; Maga, G.; Berisio, R. A structural view of SARS-CoV-2 RNA replication machinery: RNA synthesis, proofreading and final capping. Cells 2020, 9, 1267. [Google Scholar] [CrossRef] [PubMed]
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G.; O’Toole, Á.; et al. Assessing transmissibility of SARS-CoV-2 lineage B. 1.1. 7 in England. Nature 2021, 593, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Nie, J.; Wu, J.; Zhang, L.; Ding, R.; Wang, H.; Zhang, Y.; Li, T.; Liu, S.; Zhang, M.; et al. SARS-CoV-2 501Y. V2 variants lack higher infectivity but do have immune escape. Cell 2021, 184, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Arora, P.; Groß, R.; Seidel, A.; Hörnich, B.F.; Hahn, A.S.; Krüger, N.; Graichen, L.; Hofmann-Winkler, H.; Kempf, A.; et al. SARS-CoV-2 variants B. 1.351 and P. 1 escape from neutralizing antibodies. Cell 2021, 184, 2384–2393. [Google Scholar] [CrossRef]
- Dhar, M.S.; Marwal, R.; Vs, R.; Ponnusamy, K.; Jolly, B.; Bhoyar, R.C.; Sardana, V.; Naushin, S.; Rophina, M.; Mellan, T.A.; et al. Genomic characterization and epidemiology of an emerging SARS-CoV-2 variant in Delhi, India. Science 2021, 374, 995–999. [Google Scholar] [CrossRef]
- Mlcochova, P.; Kemp, S.A.; Dhar, M.S.; Papa, G.; Meng, B.; Ferreira, I.A.; Datir, R.; Collier, D.A.; Albecka, A.; Singh, S.; et al. SARS-CoV-2 B. 1.617. 2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef]
- He, X.; Hong, W.; Pan, X.; Lu, G.; Wei, X. SARS-CoV-2 Omicron variant: Characteristics and prevention. MedComm 2021, 2, 838–845. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, G. Sequence analysis of the emerging SARS-CoV-2 variant Omicron in South Africa. J. Med. Virol. 2022, 94, 1728–1733. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhi, H.; Teng, Y. The outbreak of SARS-CoV-2 Omicron lineages, immune escape and vaccine effectivity. J. Med. Virol. 2022. 10, 1002. [CrossRef]
- Mohapatra, R.K.; Tiwari, R.; Sarangi, A.K.; Sharma, S.K.; Khandia, R.; Saikumar, G.; Dhama, K. Twin combination of Omicron and Delta variants triggering a tsunami wave of ever high surges in COVID-19 cases: A challenging global threat with a special focus on the Indian subcontinent. J. Med. Virol. 2022, 94, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Leist, S.R.; Schäfer, A.; Martinez, D.R. Cell and animal models of SARS-CoV-2 pathogenesis and immunity. Dis. Models Mech. 2020, 13, dmm046581. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sarma, P.; Kaur, H.; Prajapat, M.; Bhattacharyya, A.; Avti, P.; Sehkhar, N.; Kaur, H.; Bansal, S.; Mahendiratta, S.; et al. Clinically relevant cell culture models and their significance in isolation, pathogenesis, vaccine development, repurposing and screening of new drugs for SARS-CoV-2: A systematic review. Tissue Cell 2021, 70, 101497. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Alfajaro, M.M.; DeWeirdt, P.C.; Hanna, R.E.; Lu-Culligan, W.J.; Cai, W.L.; Strine, M.S.; Zhang, S.M.; Graziano, V.R.; Schmitz, C.O.; et al. Genome-wide CRISPR screens reveal host factors critical for SARS-CoV-2 infection. Cell 2021, 184, 76–91. [Google Scholar] [CrossRef]
- Ren, X.; Glende, J.; Al-Falah, M.; de Vries, V.; Schwegmann-Wessels, C.; Qu, X.; Tan, L.; Tschernig, T.; Deng, H.; Naim, H.Y.; et al. Analysis of ACE2 in polarized epithelial cells: Surface expression and function as receptor for severe acute respiratory syndrome-associated coronavirus. J. Gen. Virol. 2006, 87, 1691–1695. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef] [Green Version]
- Ogando, N.S.; Dalebout, T.J.; Zevenhoven-Dobbe, J.C.; Limpens, R.W.; van der Meer, Y.; Caly, L.; Druce, J.; de Vries, J.J.; Kikkert, M.; Bárcena, M.; et al. SARS-coronavirus-2 replication in Vero E6 cells: Replication kinetics, rapid adaptation and cytopathology. J. Gen. Virol. 2020, 101, 925. [Google Scholar] [CrossRef]
- Meng, B.; Abdullahi, A.; Ferreira, I.A.; CITIID-NIHR BioResource COVID-19 Collaboration. Genotype to Phenotype Japan (G2P-Japan) Consortium, Ecuador-COVID19 Consortium. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagno, V. SARS-CoV-2 cellular tropism. Lancet Microbe 2020, 1, e2. [Google Scholar] [CrossRef]
- Yadav, P.D.; Gupta, N.; Potdar, V.; Mohandas, S.; Sahay, R.R.; Sarkale, P.; Shete, A.M.; Razdan, A.; Patil, D.Y.; Nyayanit, D.A.; et al. Isolation and genomic characterization of SARS-CoV-2 omicron variant obtained from human clinical specimens. Viruses 2022, 14, 461. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Wurtz, N.; Penant, G.; Jardot, P.; Duclos, N.; La Scola, B. Culture of SARS-CoV-2 in a panel of laboratory cell lines, permissivity, and differences in growth profile. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fan, X.; Bonenfant, G.; Cui, D.; Hossain, J.; Jiang, N.; Larson, G.; Currier, M.; Liddell, J.; Wilson, M.; et al. Susceptibility to SARS-CoV-2 of cell lines and substrates commonly used to diagnose and isolate influenza and other viruses. Emerg. Infect. Dis. 2021, 27, 1380. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.D.; Kode, S.S.; Keng, S.S.; Tare, D.S.; Diop, O.M.; Abraham, P.; Sharma, D.K.; Sangal, L.; Yadav, P.D.; Potdar, V.A. Replication of SARS-CoV-2 in cell lines used in public health surveillance programmes with special emphasis on biosafety. Indian J. Med. Res. 2022, 155, 129–135. [Google Scholar] [CrossRef] [PubMed]


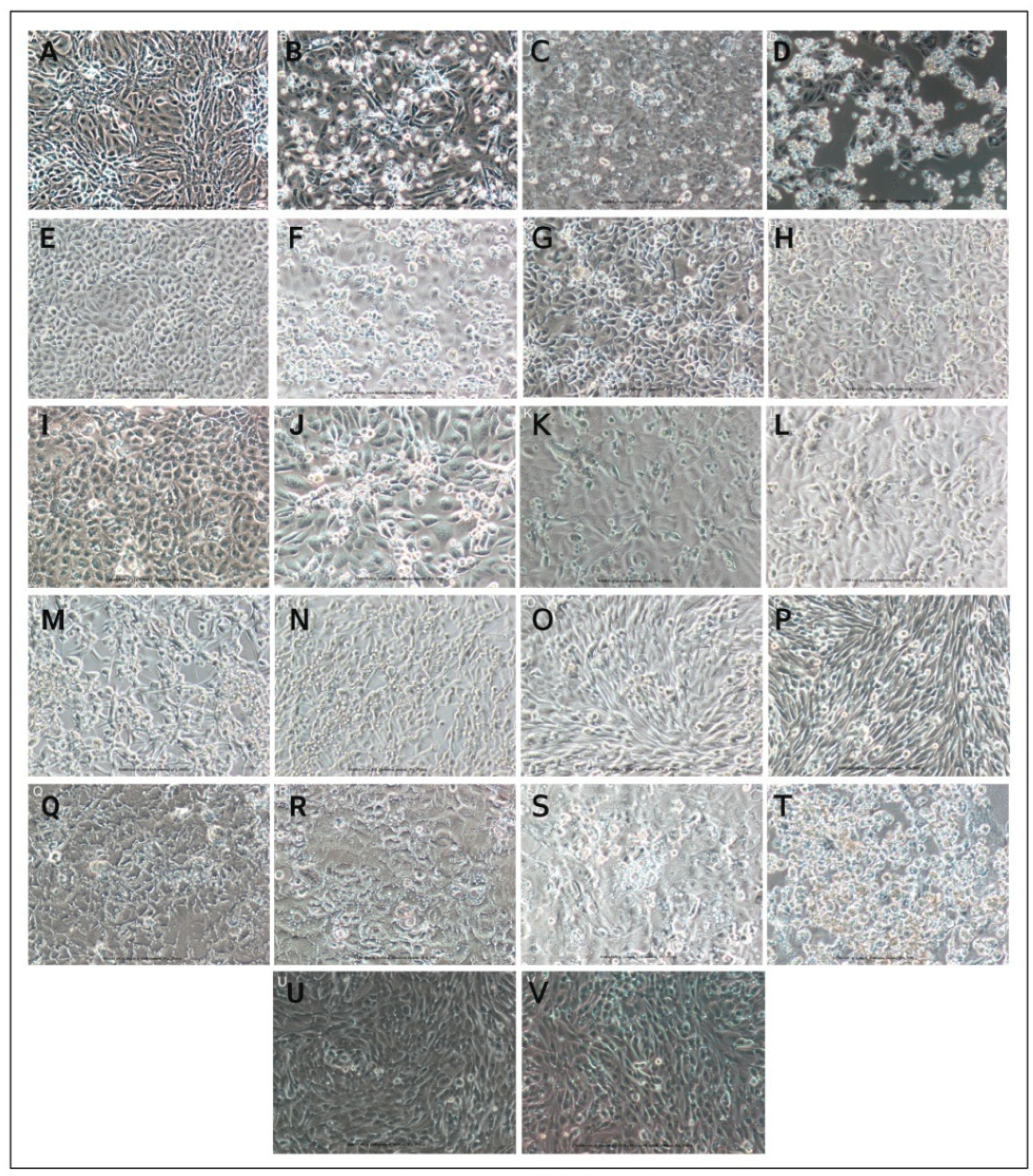{kind=link}
| Sr. No. | Cell Lines | Source | Cell Type | Tissue | Culture Medium | Culture Conditions | References |
|---|---|---|---|---|---|---|---|
| Monkey | |||||||
| 1 | Vero CCL-81 | Vero (ATCC CCL-81) | Epithelial | Kidney | MEM + 10% FBS | 37 °C, 5% CO2 | [15,26] |
| 2 | Vero E6 | ATCC no. CRL-1586 | Epithelial | Kidney | MEM + 10% FBS | 37 °C, 5% CO2 | [14,15,18,26] |
| 3 | Vero/hSLAM | 04091501-1VL | Epithelial | Kidney | DMEM + 10% FBS | 37 °C, 5% CO2 | [26] |
| 4 | MA-104 | ATCC no. CRL-2378 | Epithelial | Kidney | MEM + 10% FBS | 37 °C, 5% CO2 | [26] |
| 5 | LLC MK-2 | ATCC no. CCL-7 | Epithelial | Kidney | Medium 199 + 1% Horse serum | 37 °C, 5% CO2 | [15,26] |
| Human | |||||||
| 6 | A549 | ATCC no. CCL-185 | Epithelial-like | Lung adenocarcinoma | Nutrient mixture F-12 k HAM | 37 °C, 5% CO2 | [14,15,18] |
| 7 | RD | ATCC no. CCL-136 | Rhabdomyosarcoma | Muscle | MEM + 10% FBS | 37 °C, 5% CO2 | [Not available] |
| 8 | MRC-5 | ATCC no. CCL-171 | Fibroblast | Lung | MEM + 10% FBS | 37 °C, 5% CO2 | [26] |
| 9 | Caco-2 | ATCC no. HTB-37 | Epithelial | Colon cancer | MEM + 10% FBS | 37 °C, 5% CO2 | [15,26] |
| 10 | Calu-3 | ATCC no. HTB-55 | Epithelial | Lung adenocarcinoma | MEM + 10% FBS | 37 °C, 5% CO2 | [14,15] |
| Hamster | |||||||
| 11 | BHK-21 | ATCC no. CCL-10 | Fibroblast | Kidney | MEM + 10% FBS | 37 °C, 5% CO2 | [15,26] |
| Sr. No. | Cell Line | First Round of Infection (P-1) | Second Round of Infection (P-2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | ||
| 1 | Vero CCL-81 | N | 1+ | 2+ | 4+ | 4+ | N | 1+ | 2+ | 4+ | 4+ |
| 2 | Vero E6 | N | 1+ | 2+ | 4+ | 4+ | N | 1+ | 2+ | 4+ | 4+ |
| 3 | Vero/hSLAM | N | N | 1+ | 2+ | 4+ | N | N | 1+ | 2+ | 4+ |
| 4 | MA-104 | N | N | N | 2+ | 4+ | N | N | 1+ | 3+ | 4+ |
| 5 | LLC MK-2 | N | N | N | 1+ | 3+ | N | N | 1+ | 3+ | 4+ |
| 6 | A549 | N | N | N | N | N | N | N | N | N | N |
| 7 | RD | N | N | N | N | N | N | N | N | N | N |
| 8 | MRC-5 | N | N | N | N | N | N | N | N | N | N |
| 9 | Caco-2 | N | N | N | N | N | N | N | N | N | N |
| 10 | Calu-3 | N | N | N | 2+ | 4+ | N | N | 1+ | 2+ | 4+ |
| 11 | BHK-21 | N | N | N | N | N | N | N | N | N | N |
| Cell Line | First Round of Infection (P-1) | Second Round of Infection (P-2) | Permissiveness (Yes/No) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CPE Observation | Day of Start of CPE (PID) | TCID50/mL | Viral Loads (Copies/mL) | Times of Viral Loads Normalized to Baseline | CPE Observation | Day of Start of CPE (PID) | TCID50/mL | Viral Loads (Copies/mL) | Times of Viral Loads Normalized to Baseline | ||
| Monkey | |||||||||||
| Vero CCL-81 | CPE Observed | 2 | 105.33 | 1.96 × 1010 | 0.675 | CPE Observed | 2 | 105.66 | 2.83 × 1010 | 0.972 | Highly permissive |
| Vero E6 | CPE Observed | 2 | 104.67 | 1.53 × 1010 | 0.527 | CPE Observed | 2 | 104.67 | 3.72 × 1010 | 1.281 | Highly permissive |
| Vero/hSLAM | CPE Observed | 3 | 102.5 | 2.50 × 1010 | 0.859 | CPE Observed | 3 | 102.5 | 3.18 × 1010 | 1.094 | Permissive |
| MA-104 | CPE Observed | 4 | 102.67 | 1.01 × 1011 | 3.482 | CPE Observed | 3 | 103.67 | 1.36 × 1011 | 4.691 | Permissive |
| LLC MK-2 | CPE Observed | 4 | 102.67 | 1.59 × 1010 | 0.548 | CPE Observed | 3 | 103.23 | 4.35 × 109 | 0.15 | Permissive |
| Human | |||||||||||
| A549 | No CPE | NA | 0 | 1.65 × 109 | 0.057 | No CPE | NA | 0 | 3.64 × 107 | 0.001 | Permissive |
| RD | No CPE | NA | 0 | 2.55 × 109 | 0.088 | No CPE | NA | 0 | 5.76 × 109 | 0.198 | Permissive |
| MRC-5 | No CPE | NA | 0 | 6.12 × 108 | 0.021 | No CPE | NA | 0 | 2.70 × 107 | 0.001 | Permissive |
| Caco-2 | No CPE | NA | 103 | 5.04 × 109 | 0.173 | No CPE | NA | 103 | 2.06 × 109 | 0.071 | Permissive |
| Calu-3 | CPE Observed | 4 | 103.67 | 2.40 × 1010 | 0.826 | CPE Observed | 3 | 104.44 | 8.90 × 1010 | 3.06 | Highly permissive |
| Hamster | |||||||||||
| BHK-21 | No CPE | NA | 0 | 1.26 × 109 | 0.043 | No CPE | NA | 0 | 1.77 × 107 | 0.001 | Permissive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dighe, H.; Sarkale, P.; Patil, D.Y.; Mohandas, S.; Shete, A.M.; Sahay, R.R.; Lakra, R.; Patil, S.; Majumdar, T.; Gawande, P.; et al. Differential Cell Line Susceptibility to the SARS-CoV-2 Omicron BA.1.1 Variant of Concern. Vaccines 2022, 10, 1962. https://doi.org/10.3390/vaccines10111962
Dighe H, Sarkale P, Patil DY, Mohandas S, Shete AM, Sahay RR, Lakra R, Patil S, Majumdar T, Gawande P, et al. Differential Cell Line Susceptibility to the SARS-CoV-2 Omicron BA.1.1 Variant of Concern. Vaccines. 2022; 10(11):1962. https://doi.org/10.3390/vaccines10111962
Chicago/Turabian StyleDighe, Hitesh, Prasad Sarkale, Deepak Y. Patil, Sreelekshmy Mohandas, Anita M. Shete, Rima R. Sahay, Rajen Lakra, Savita Patil, Triparna Majumdar, Pranita Gawande, and et al. 2022. "Differential Cell Line Susceptibility to the SARS-CoV-2 Omicron BA.1.1 Variant of Concern" Vaccines 10, no. 11: 1962. https://doi.org/10.3390/vaccines10111962
APA StyleDighe, H., Sarkale, P., Patil, D. Y., Mohandas, S., Shete, A. M., Sahay, R. R., Lakra, R., Patil, S., Majumdar, T., Gawande, P., Yemul, J., Vedpathak, P., & Yadav, P. D. (2022). Differential Cell Line Susceptibility to the SARS-CoV-2 Omicron BA.1.1 Variant of Concern. Vaccines, 10(11), 1962. https://doi.org/10.3390/vaccines10111962