

In Silico Discovery of GPCRs and GnRHRs as Novel Binding Receptors of SARS-CoV-2 Spike Protein Could Explain Neuroendocrine Disorders in COVID-19

 , , , ,
, , , ,  , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
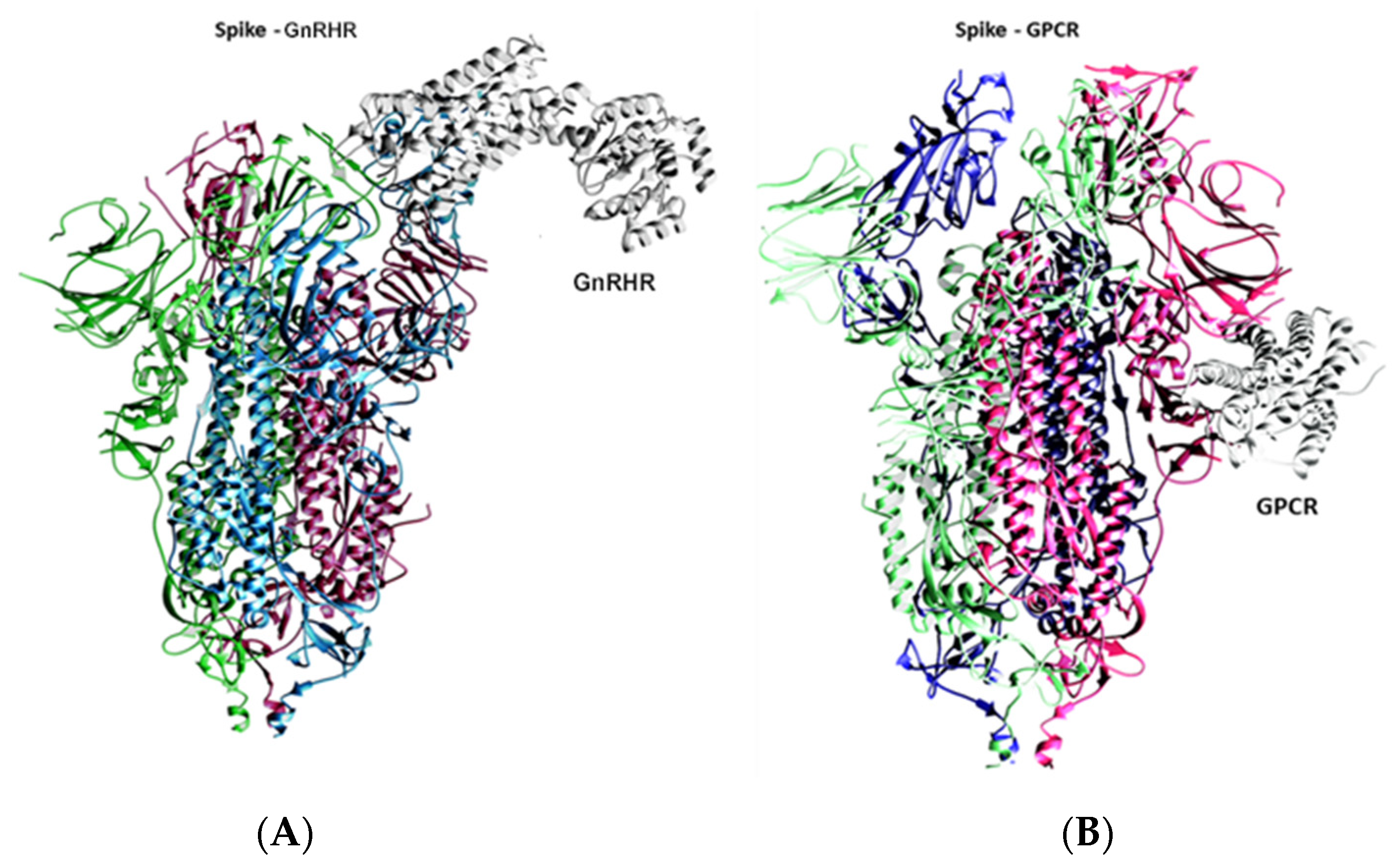
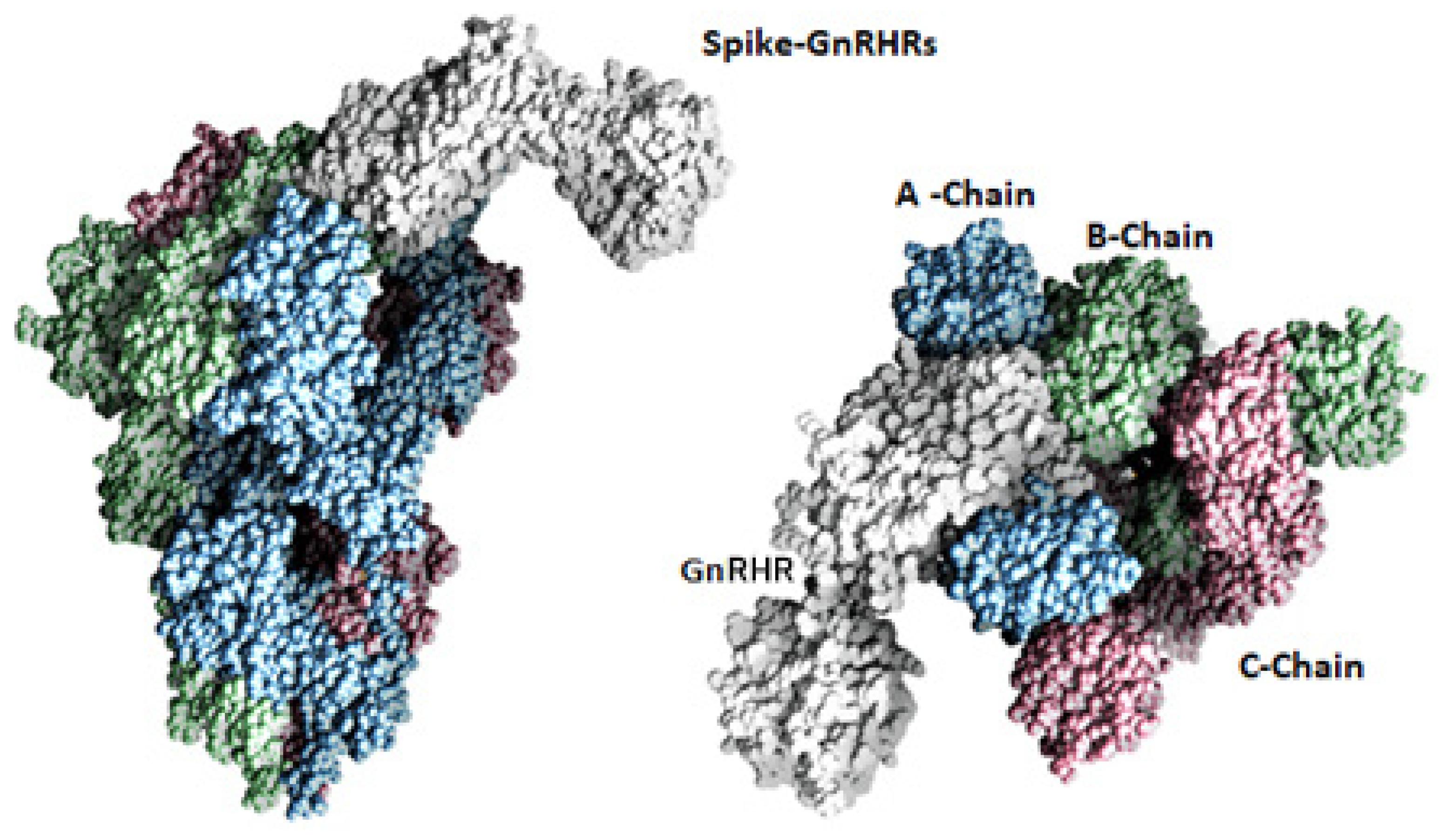
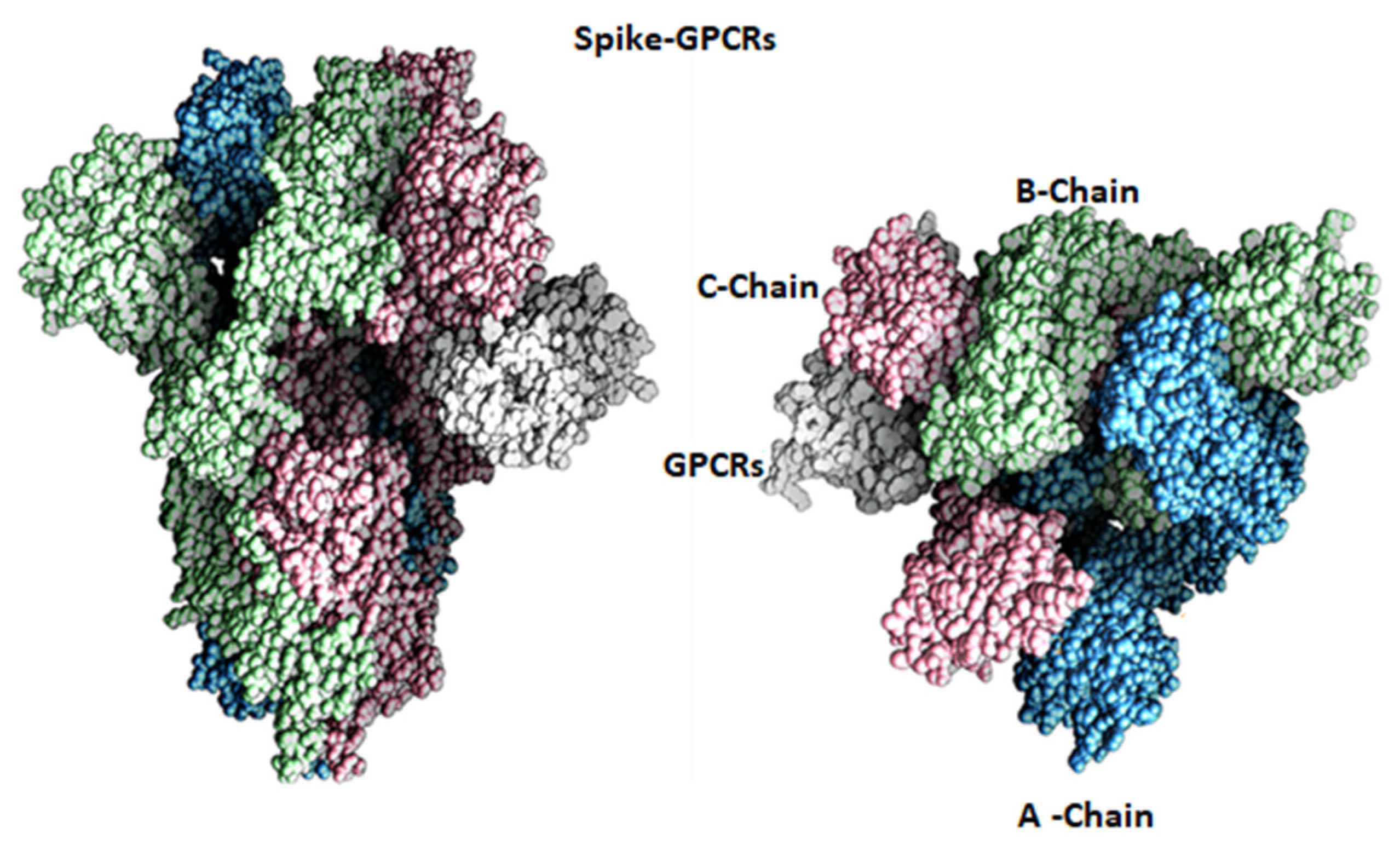
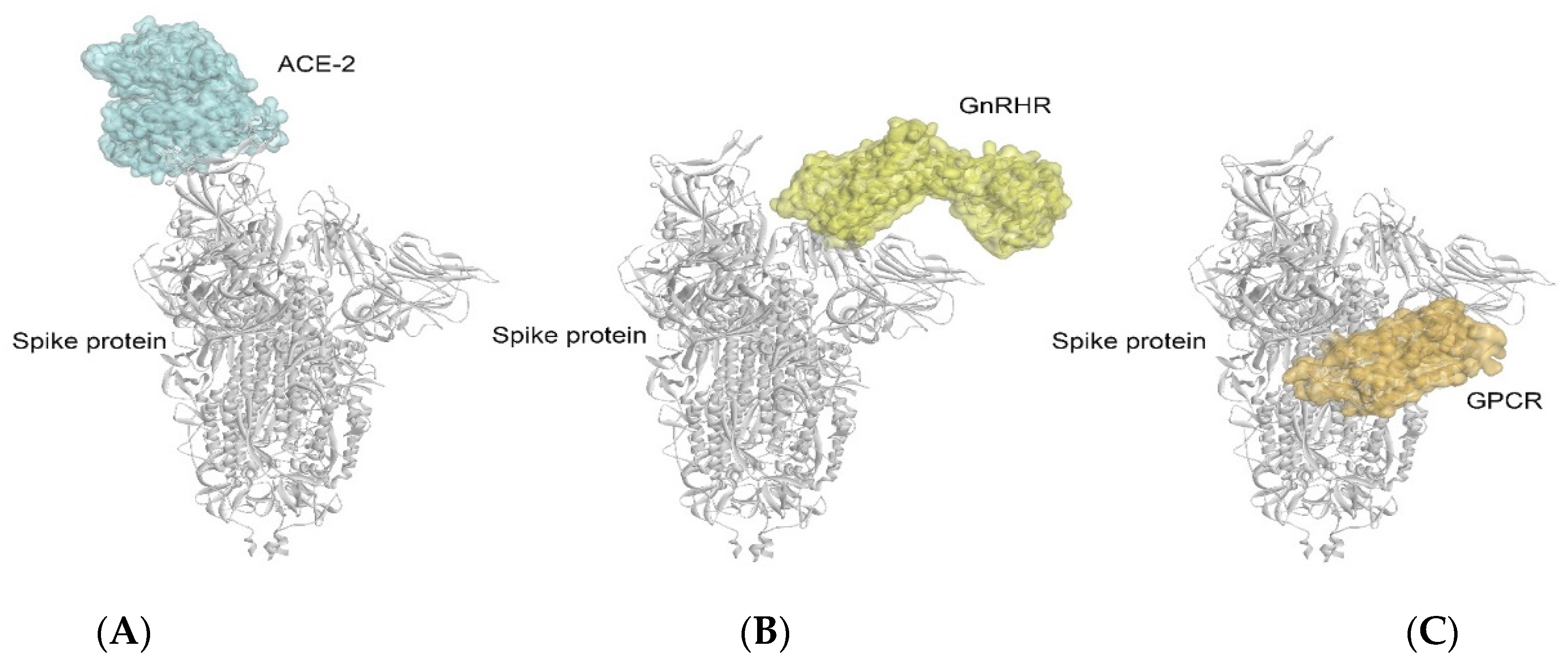
2.1. Docking Analysis
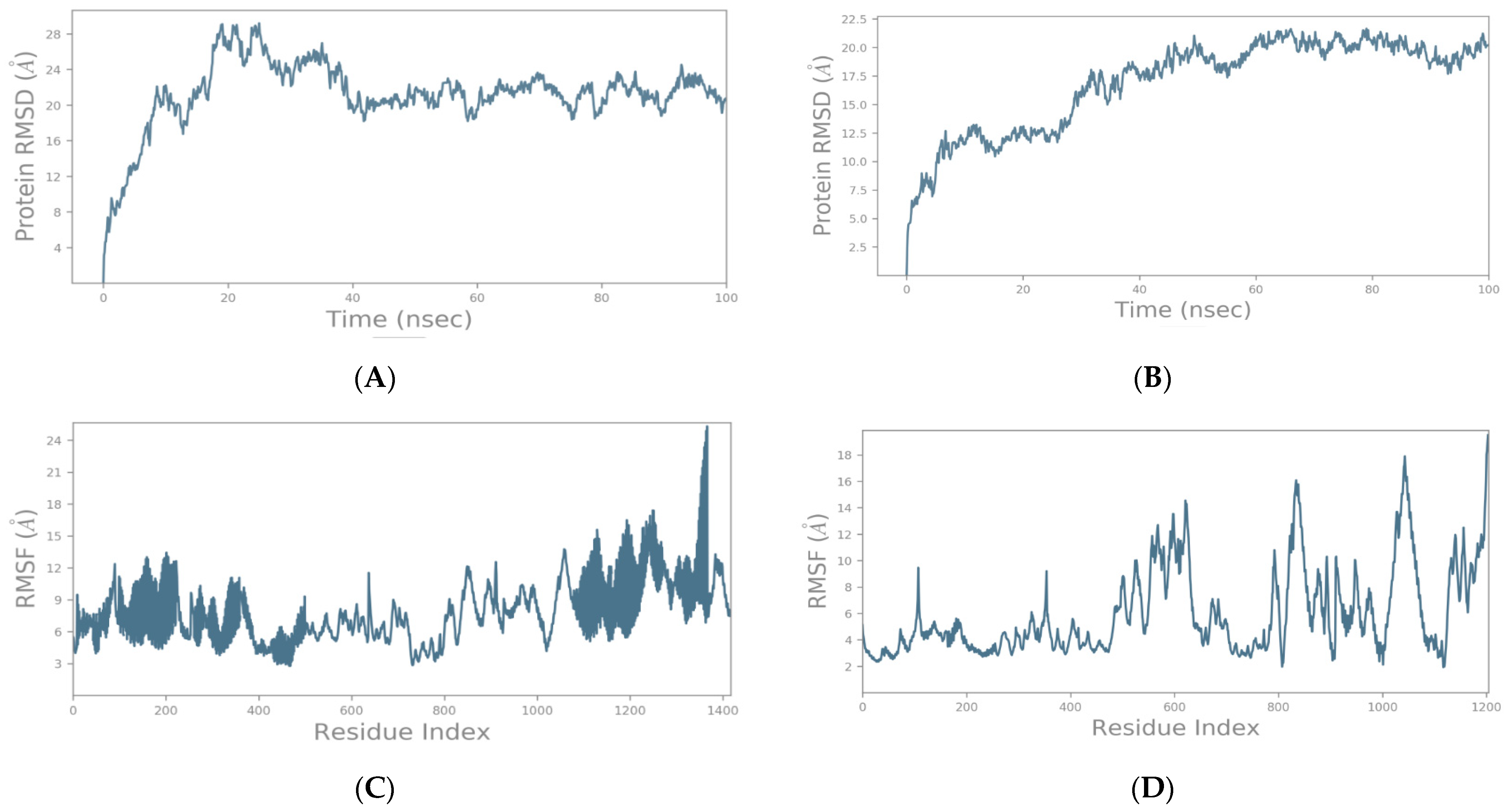
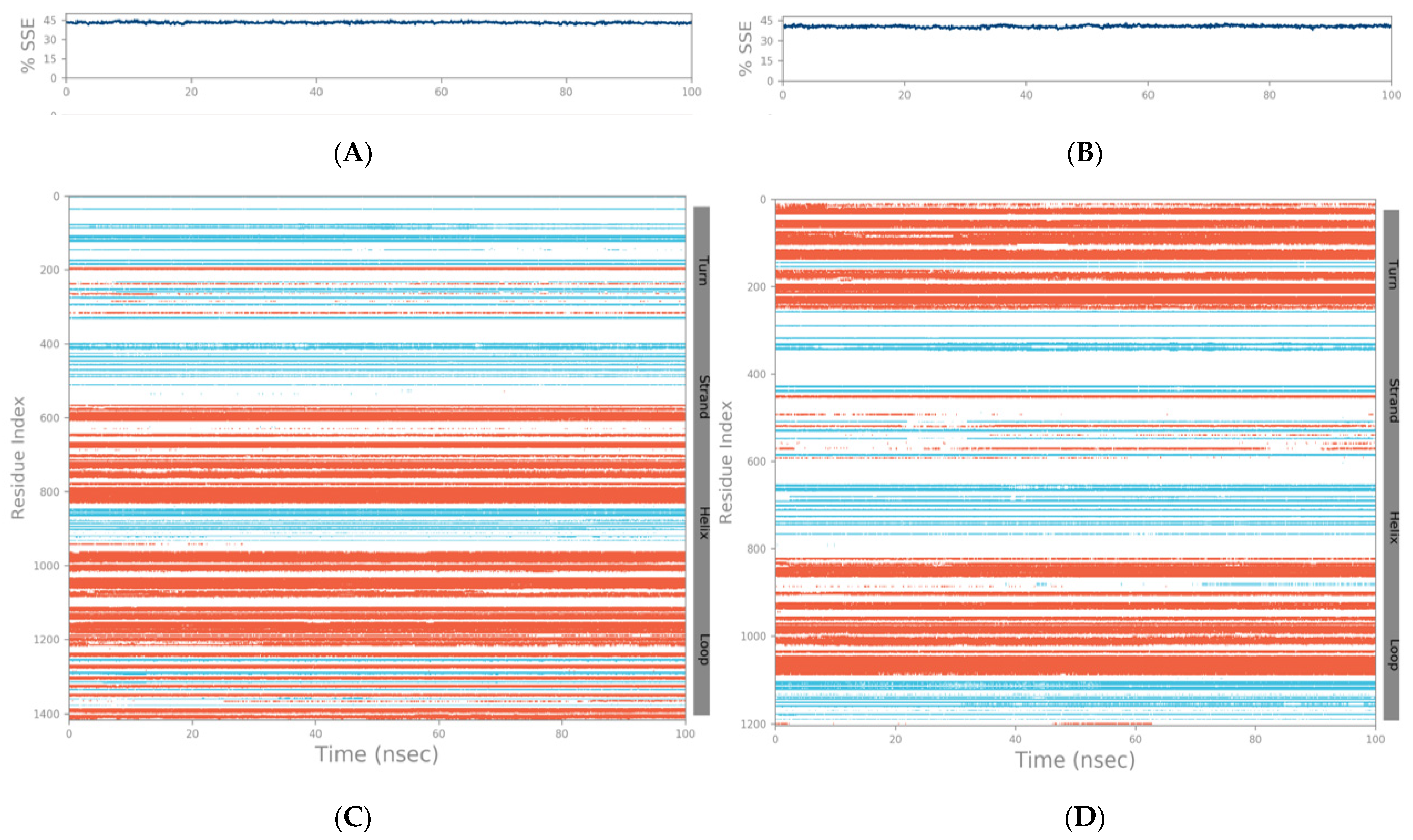
2.2. Molecular Dynamics Simulation Studies of Spike–GnRHR and Spike–GPCR Complexes
3. Materials and Methods
3.1. Proteins Dataset
3.2. Molecular Docking Using PatchDock Program
3.3. Molecular Dynamics (MD) Simulation
3.4. Analysis of Molecular Dynamics Trajectory
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basu, A.; Sarkar, A.; Maulik, U. Molecular docking study of potential phytochemicals and their effects on the complex of SARS-CoV2 spike protein and human ACE2. Sci. Rep. 2020, 10, 17699. [Google Scholar] [CrossRef]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.-J.; Jiang, S. The spike protein of SARS-CoV—a target for vaccine and therapeutic development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef]
- Soni, S.; Jiang, Y.; Tesfaigzi, Y.; Hornick, J.L.; Çataltepe, S. Comparative analysis of ACE2 protein expression in rodent, non-human primate, and human respiratory tract at baseline and after injury: A conundrum for COVID-19 pathogenesis. PLoS ONE 2021, 16, e0247510. [Google Scholar]
- Zeng, M.; Wang, D.-Y.; Mullol, J.; Liu, Z. Chemosensory dysfunction in patients with COVID-19: What do we learn from the global outbreak? Curr. Allergy Asthma Rep. 2021, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.; Hanchapola, I.; Thomas, W.G.; Widdop, R.E.; Smith, A.I.; Perlmutter, P.; Aguilar, M.-I. Structural determinants for binding to angiotensin converting enzyme 2 (ACE2) and angiotensin receptors 1 and 2. Front. Pharmacol. 2015, 6, 5. [Google Scholar] [CrossRef]
- Porrello, E.R.; Delbridge, L.M.; Thomas, W.G. The angiotensin II type 2 (AT2) receptor: An enigmatic seven transmembrane receptor. Front. Biosci.-Landmark 2009, 14, 958–972. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.; Hsieh, F.; Baronas, E.; Godbout, K.; Gosselin, M.; Stagliano, N.; Donovan, M.; Woolf, B.; Robison, K.; Jeyaseelan, R. A novel angiotensin-converting enzyme–related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1-9. Circ. Res. 2000, 87, e1–e9. [Google Scholar] [CrossRef]
- Singh, Y.; Gupta, G.; Satija, S.; Pabreja, K.; Chellappan, D.K.; Dua, K. COVID-19 transmission through host cell directed network of GPCR. Drug Dev. Res. 2020, 81, 647–649. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.; Lely, A.; Navis, G.V.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Bourgonje, A.R.; Abdulle, A.E.; Timens, W.; Hillebrands, J.L.; Navis, G.J.; Gordijn, S.J.; Bolling, M.C.; Dijkstra, G.; Voors, A.A.; Osterhaus, A.D. Angiotensin-converting enzyme 2 (ACE2), SARS-CoV-2 and the pathophysiology of coronavirus disease 2019 (COVID-19). J. Pathol. 2020, 251, 228–248. [Google Scholar] [CrossRef]
- Hopkins, C.; Lechien, J.R.; Saussez, S. More that ACE2? NRP1 may play a central role in the underlying pathophysiological mechanism of olfactory dysfunction in COVID-19 and its association with enhanced survival. Med. Hypotheses 2021, 146, 110406. [Google Scholar] [CrossRef]
- Alhenc-Gelas, F.; Drueke, T.B. Blockade of SARS-CoV-2 infection by recombinant soluble ACE2. Kidney Int. 2020, 97, 1091–1093. [Google Scholar] [CrossRef]
- Brann, D.H.; Tsukahara, T.; Weinreb, C.; Lipovsek, M.; Van den Berge, K.; Gong, B.; Chance, R.; Macaulay, I.C.; Chou, H.-J.; Fletcher, R.B. Non-neuronal expression of SARS-CoV-2 entry genes in the olfactory system suggests mechanisms underlying COVID-19-associated anosmia. Sci. Adv. 2020, 6, eabc5801. [Google Scholar] [CrossRef] [PubMed]
- Penn, R.B.; Benovic, J.L. Regulation of heterotrimeric G protein signaling in airway smooth muscle. Proc. Am. Thorac. Soc. 2008, 5, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Trzaskowski, B.; Latek, D.; Yuan, S.; Ghoshdastider, U.; Debinski, A.; Filipek, S. Action of molecular switches in GPCRs-theoretical and experimental studies. Curr. Med. Chem. 2012, 19, 1090–1109. [Google Scholar] [CrossRef] [PubMed]
- Im, D.-S. Orphan G protein-coupled receptors and beyond. Jpn. J. Pharmacol. 2002, 90, 101–106. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Firestein, S. The good taste of genomics. Nature 2000, 404, 552–553. [Google Scholar] [CrossRef]
- Kerekoppa, R.; Basavaraju, M.; Rao, A.; Geetha, G.R.; Mukund, K.; Sakthivel, J. Single nucleotide polymorphism identification and characterization of GnRHR gene in Bos taurus and Bos indicus cattle. Turk. J. Vet. Anim. Sci. 2015, 39, 10–15. [Google Scholar] [CrossRef]
- Lirón, J.; Prando, A.; Ripoli, M.; Rogberg-Muñoz, A.; Posik, D.; Baldo, A.; Peral-García, P.; Giovambattista, G. Characterization and validation of bovine Gonadotripin releasing hormone receptor (GNRHR) polymorphisms. Res. Vet. Sci. 2011, 91, 391–396. [Google Scholar] [CrossRef]
- Midha, I.K.; Kumar, N.; Kumar, A.; Madan, T. Mega doses of retinol: A possible immunomodulation in Covid-19 illness in resource-limited settings. Rev. Med. Virol. 2021, 31, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vollenberg, R.; Tepasse, P.-R.; Fobker, M.; Hüsing-Kabar, A. Significantly Reduced Retinol Binding Protein 4 (RBP4) Levels in Critically Ill COVID-19 Patients. Nutrients 2022, 14, 2007. [Google Scholar] [CrossRef] [PubMed]
- Fatriansyah, J.F.; Rizqillah, R.K.; Yandi, M.Y.; Sahlan, M. Molecular docking and dynamics studies on propolis sulabiroin-A as a potential inhibitor of SARS-CoV-2. J. King Saud Univ.-Sci. 2022, 34, 101707. [Google Scholar] [CrossRef] [PubMed]
- Perrett, R.M.; McArdle, C.A. Molecular mechanisms of gonadotropin-releasing hormone signaling: Integrating cyclic nucleotides into the network. Front. Endocrinol. 2013, 4, 180. [Google Scholar] [CrossRef]
- Connolly, M.L. Solvent-accessible surfaces of proteins and nucleic acids. Science 1983, 221, 709–713. [Google Scholar] [CrossRef]
- Connolly, M.L. Analytical molecular surface calculation. J. Appl. Crystallogr. 1983, 16, 548–558. [Google Scholar] [CrossRef]
- Zhang, C.; Vasmatzis, G.; Cornette, J.L.; DeLisi, C. Determination of atomic desolvation energies from the structures of crystallized proteins. J. Mol. Biol. 1997, 267, 707–726. [Google Scholar] [CrossRef]
- Priya Doss, C.G.; Chakraborty, C.; Chen, L.; Zhu, H. Integrating in silico prediction methods, molecular docking, and molecular dynamics simulation to predict the impact of ALK missense mutations in structural perspective. BioMed Res. Int. 2014, 2014, 895831. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | GnRHR (Chain:Residue:Atom) | Spike Chain B (Chain:Residue:Atom) | Distance (Å) |
|---|---|---|---|
| 1 | A:GLN204:HE22 | B:GLY413:O | 2.36 |
| 2 | A:THR30:OG1 | B:LYS113:HZ2 | 1.64 |
| 3 | A:TRP205:O | B:LYS378:HZ2 | 1.68 |
| 4 | A:TRP206:O | B:ARG408:HE | 2.04 |
| 5 | A:HIS207:O | B:ARG408:HE | 2.76 |
| 6 | A:HIS207:O | B:ARG408:HH12 | 1.80 |
| 7 | A:SER217:OG | B:TYR508:HH | 2.20 |
| 8 | A:SER217:OG | B:SER375:CB | 3.47 |
| Sr. No. | GPCR (Chain:Residue:Atom) | Spike Chain B (Chain:Residue:Atom) | Distance (Å) |
|---|---|---|---|
| 1 | A:TRP588:HE1 | B:ASN603:O | 2.21 |
| 2 | A:CYS601:SG | B:ASN606:OD1 | 3.50 |
| 3 | A:CYS601:SG | B:ASN606:O | 3.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elkazzaz, M.; Ahmed, A.; Abo-Amer, Y.E.-E.; Hydara, T.; Haikal, A.; Razek, D.N.A.E.; Eltayb, W.A.; Wang, X.; Karpiński, T.M.; Hamza, D.; et al. In Silico Discovery of GPCRs and GnRHRs as Novel Binding Receptors of SARS-CoV-2 Spike Protein Could Explain Neuroendocrine Disorders in COVID-19. Vaccines 2022, 10, 1500. https://doi.org/10.3390/vaccines10091500
Elkazzaz M, Ahmed A, Abo-Amer YE-E, Hydara T, Haikal A, Razek DNAE, Eltayb WA, Wang X, Karpiński TM, Hamza D, et al. In Silico Discovery of GPCRs and GnRHRs as Novel Binding Receptors of SARS-CoV-2 Spike Protein Could Explain Neuroendocrine Disorders in COVID-19. Vaccines. 2022; 10(9):1500. https://doi.org/10.3390/vaccines10091500
Chicago/Turabian StyleElkazzaz, Mahmoud, Amr Ahmed, Yousry Esam-Eldin Abo-Amer, Tamer Hydara, Abdullah Haikal, Dina N. Abd El Razek, Wafa Ali Eltayb, Xiling Wang, Tomasz M. Karpiński, Dalia Hamza, and et al. 2022. "In Silico Discovery of GPCRs and GnRHRs as Novel Binding Receptors of SARS-CoV-2 Spike Protein Could Explain Neuroendocrine Disorders in COVID-19" Vaccines 10, no. 9: 1500. https://doi.org/10.3390/vaccines10091500
APA StyleElkazzaz, M., Ahmed, A., Abo-Amer, Y. E. -E., Hydara, T., Haikal, A., Razek, D. N. A. E., Eltayb, W. A., Wang, X., Karpiński, T. M., Hamza, D., Jabbar, B., & Shamkh, I. M. (2022). In Silico Discovery of GPCRs and GnRHRs as Novel Binding Receptors of SARS-CoV-2 Spike Protein Could Explain Neuroendocrine Disorders in COVID-19. Vaccines, 10(9), 1500. https://doi.org/10.3390/vaccines10091500