

Population-Based Analysis of the Immunoglobulin G Response to Different COVID-19 Vaccines in Brazil
 ,
,  and
and 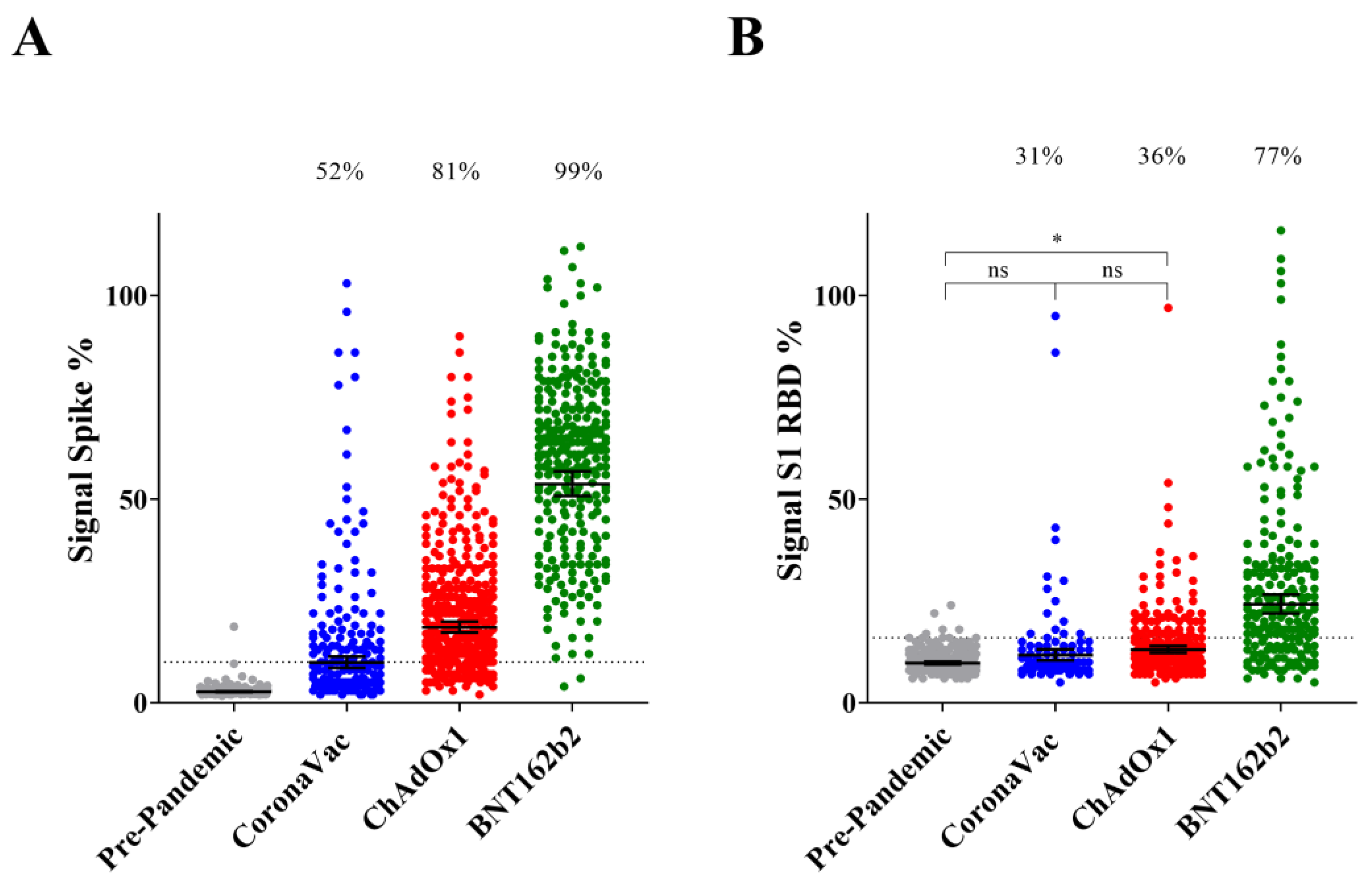{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Sampling
2.2. Serological Analysis
2.3. Statistical Analysis
3. Results
3.1. Seroconversion Rates for Nucleocapsid, Spike Protein and S1 RBD
3.2. IgG Reactive to Spike Protein and S1 RBD upon Completion of Primary Vaccination
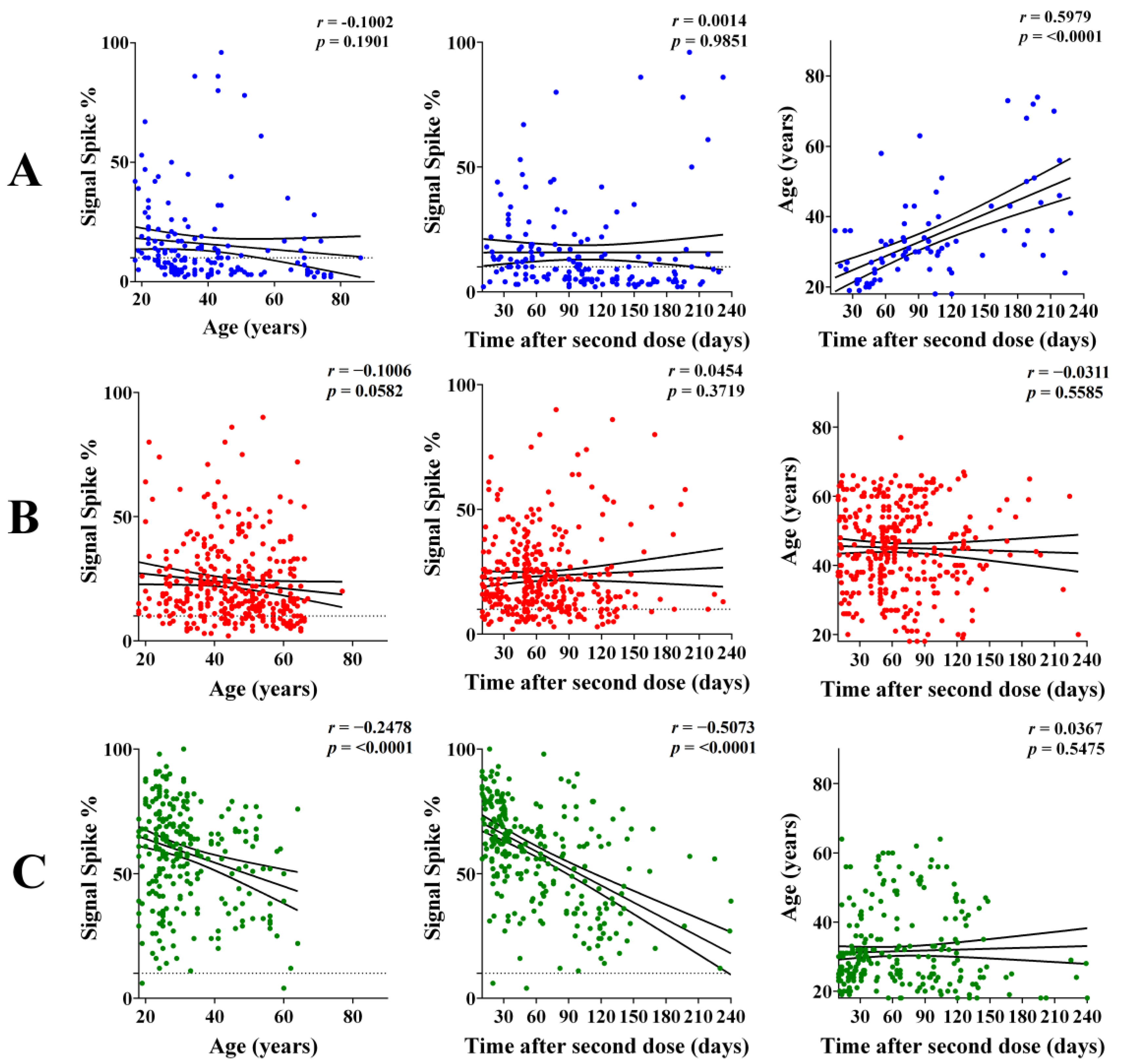
3.3. IgG Response According to Age and Interval after the Second Dose
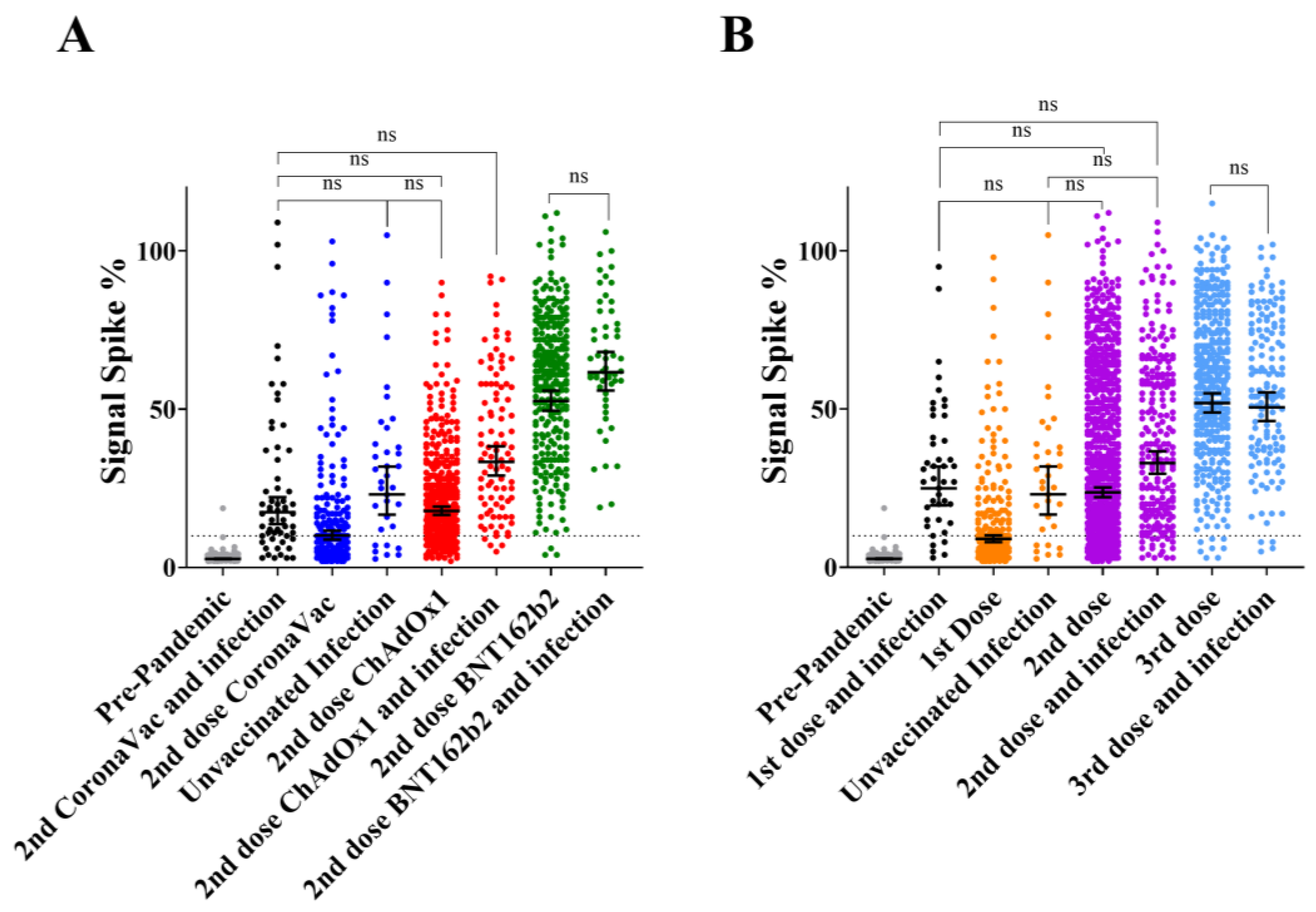
3.4. Comparison of Primary Vaccination Concurring with Natural Infection Accordingly to Vaccine Type
3.5. IgG Response to Infection vs. Multiple Dose Vaccination
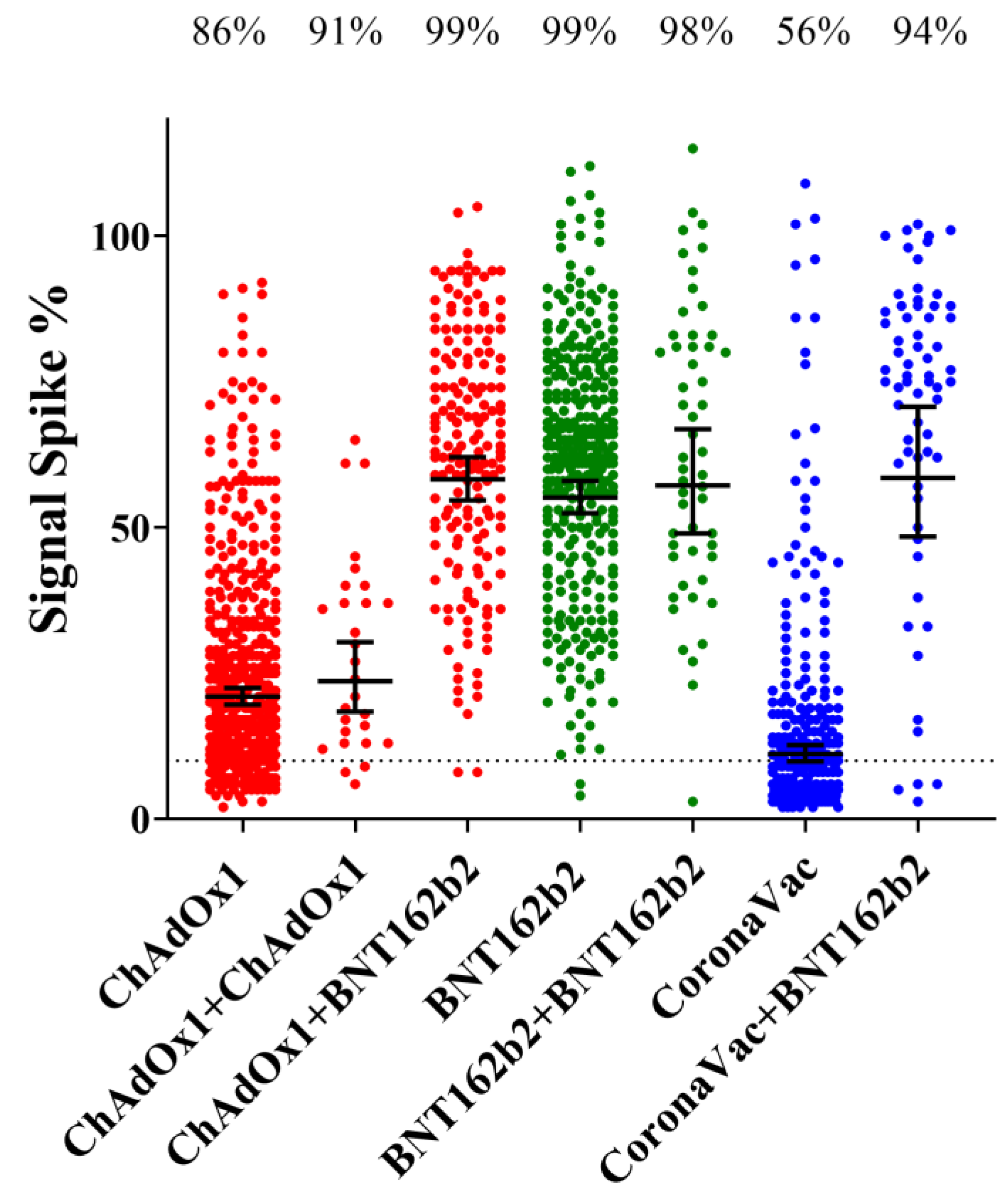
3.6. Effect of the Booster Dose (3rd Dose) According to the Vaccine Type
3.7. Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortega, N.; Ribes, M.; Vidal, M.; Rubio, R.; Aguilar, R.; Williams, S.; Dobaño, C. Seven-month kinetics of SARS-CoV-2 antibodies and role of pre-existing antibodies to human coronaviruses. Nat. Commun. 2021, 12, 4740. [Google Scholar] [CrossRef]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Sadarangani, M.; Marchant, A.; Kollmann, T.R. Immunological mechanisms of vaccine-induced protection against COVID-19 in humans. Nat. Rev. Immunol. 2021, 21, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Tanriover, M.D.; Doğanay, H.L.; Akova, M.; Güner, H.R.; Azap, A.; Akhan, S.; Köse, Ş.; Erdinç, F.Ş.; Akalın, E.H.; Tabak, Ö.F.; et al. Efficacy and safety of an inactivated whole-virion SARS-CoV-2 vaccine (CoronaVac): Interim results of a double-blind, randomised, placebo-controlled, phase 3 trial in Turkey. Lancet 2021, 398, 213–222. [Google Scholar] [CrossRef]
- Cerqueira-Silva, T.; Katikireddi, S.V.; Oliveira, V.D.A.; Flores-Ortiz, R.; Júnior, J.B.; Paixão, E.S.; Robertson, C.; Penna, G.O.; Werneck, G.L.; Barreto, M.L.; et al. Vaccine effectiveness of heterologous CoronaVac plus BNT162b2 in Brazil. Nat. Med. 2022, 28, 838–843. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Gilbert, P.B.; Montefiori, D.C.; McDermott, A.B.; Fong, Y.; Benkeser, D.; Deng, W.; Zhou, H.; Houchens, C.R.; Martins, K.; Jayashankar, L.; et al. Immune correlates analysis of the mRNA- 1273 COVID-19 vaccine efficacy clinical trial. Science 2022, 375, 43–50. [Google Scholar] [CrossRef]
- Feng, S.; Phillips, D.J.; White, T.; Sayal, H.; Aley, P.K.; Bibi, S.; Voysey, M. Correlates of protection against symptomatic and asymptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 2032–2040. [Google Scholar] [CrossRef]
- Medeiros-Ribeiro, A.C.; Aikawa, N.E.; Saad, C.G.S.; Yuki, E.F.N.; Pedrosa, T.; Fusco, S.R.G.; Rojo, P.T.; Pereira, R.M.R.; Shinjo, S.K.; Andrade, D.C.O.; et al. Immunogenicity and safety of an inactivated virus vaccine against SARS-Cov-2 in patients with autoimmune rheumatic diseases. Arthritis Rheumatol. 2021, 73, 1744–1751. [Google Scholar] [CrossRef]
- Bochnia-Bueno, L.; De Almeida, S.M.; Raboni, S.M.; Adamoski, D.; Amadeu LL, M.; Carstensen, S.; Nogueira, M.B. SARS-CoV-2 Vaccination with CoronaVac: Seroconversion Rate in Healthcare Workers After 40 Days. SSRN Electron. J. 2021. [Google Scholar] [CrossRef]
- Grenfell, R.F.; Almeida, N.B.; Filgueiras, P.S.; Corsini, C.A.; Gomes, S.V.; de Miranda, D.A.; Campi-Azevedo, A.C. Immunogenicity, Effectiveness, and Safety of Inactivated Virus (CoronaVac) Vaccine in a Two-Dose Primary Protocol and BNT162b2 Heterologous Booster in Brazil (Immunita-001): A One Year Period Follow Up Phase 4 Study. Front. Immunol. 2022, 13, 918896. [Google Scholar] [CrossRef] [PubMed]
- Huergo, L.F.; Paula, N.M.; Gonçalves, A.C.A.; Kluge, C.H.S.; Marins, P.H.S.A.; Camargo, H.S.C.; Sant’Ana, T.P.; Farias, L.R.P.; Aldrighi, J.D.; Lima, ÊS.; et al. SARS-CoV-2 Seroconversion in Response to Infection and Vaccination: A Time Series Local Study in Brazil. Microbiol. Spectr. 2022, 10, e01026-22. [Google Scholar] [CrossRef] [PubMed]
- Conzentino, M.S.; Gonçalves, A.C.A.; Paula, N.M.; Rego, F.G.M.; Zanette, D.L.; Aoki, M.N.; Nardin, J.M.; Huergo, L.F. A magnetic bead immunoassay to detect high affinity human IgG reactive to SARS-CoV-2 Spike S1 RBD produced in Escherichia coli. Braz. J. Microbiol. 2022, 53, 1263–1269. [Google Scholar] [CrossRef]
- Huergo, L.F.; Selim, K.A.; Conzentino, M.S.; Gerhardt, E.C.M.; Santos, A.R.S.; Wagner, B.; Alford, J.T.; Deobald, N.; Pedrosa, F.O.; de Souza, E.M.; et al. Magnetic Bead-Based Immunoassay Allows Rapid, Inexpensive, and Quantitative Detection of Human SARS-CoV-2 Antibodies. ACS Sens. 2021, 3, 703–708. [Google Scholar] [CrossRef]
- Conzentino, M.S.; Forchhammer, K.; Souza, E.M.; Pedrosa, F.O.; Nogueira, M.B.; Raboni, S.M.; Rego, F.G.M.; Zanette, D.L.; Aoki, M.N.; Nardin, J.M.; et al. Antigen production and development of an indirect ELISA based on the nucleocapsid protein to detect human SARS-CoV-2 seroconversion. Braz. J. Microbiol. 2021, 52, 2069–2073. [Google Scholar] [CrossRef]
- Conzentino, M.S.; Santos, T.P.; Selim, K.A.; Wagner, B.; Alford, J.T.; Deobald, N.; Paula, N.M.; Rego, F.G.; Zanette, D.L.; Aoki, M.N.; et al. Ultra-fast, high throughput and inexpensive detection of SARS-CoV-2 seroconversion using Ni2+ magnetic beads. Anal. Biochem. 2021, 631, 114360. [Google Scholar] [CrossRef]
- Alvim, R.G.; Lima, T.M.; Rodrigues, D.A.; Marsili, F.F.; Bozza, V.B.; Higa, L.M.; Monteiro, F.L.; Abreu, D.P.; Leitão, I.C.; Carvalho, R.S.; et al. From a recombinant key antigen to an accurate, affordable serological test: Lessons learnt from COVID-19 for future pandemics. Biochem. Eng. J. 2022, 186, 108537. [Google Scholar] [CrossRef]
- Adamoski, D.; Baura VA, D.; Rodrigues, A.C.; Royer, C.A.; Aoki, M.N.; Tschá, M.K.; Carvalho de Oliveira, J. SARS-CoV-2 Delta and Omicron Variants Surge in Curitiba, Southern Brazil, and Its Impact on Overall COVID-19 Lethality. Viruses 2022, 14, 809. [Google Scholar] [CrossRef]
- Carreño, J.M.; Alshammary, H.; Tcheou, J.; Singh, G.; Raskin, A.J.; Kawabata, H.; Sominsky, L.A.; Clark, J.J.; Adelsberg, D.C.; Bielak, D.A.; et al. Activity of convalescent and vaccine serum against SARS-CoV-2 Omicron. Nature 2022, 602, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Saunders, N.; Maes, P.; Guivel-Benhassine, F.; Planchais, C.; Buchrieser, J.; Bolland, W.H.; Porrot, F.; Staropoli, I.; Lemoine, F.; et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 2021, 602, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.P.Y.; Ho, J.C.W.; Cheung, M.-C.; Ng, K.-C.; Ching, R.H.H.; Lai, K.-L.; Kam, T.T.; Gu, H.; Sit, K.-Y.; Hsin, M.K.Y.; et al. SARS-CoV-2 Omicron variant replication in human bronchus and lung ex vivo. Nature 2022, 603, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Romero-Pinedo, S.; Quesada, M.; Horndler, L.; Álvarez-Fernández, S.; Olmo, A.; Abia, D.; Alarcón, B.; Delgado, P. Vaccine Type-, Age- and Past Infection- Dependence of the Humoral Response to SARS-CoV-2 Spike S Protein. Front. Immunol. 2022, 13, 809285. [Google Scholar] [CrossRef] [PubMed]
- Barin, B.; Kasap, U.; Selçuk, F.; Volkan, E.; Uluçkan, Ö. Comparison of SARS-CoV-2 anti-spike receptor binding domain IgG antibody responses after CoronaVac, BNT162b2, ChAdOx1 COVID-19 vaccines, and a single booster dose: A prospective, longitudinal population-based study. Lancet Microbe 2022, 3, e274–e283. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.A.C.; Weckx, L.; Clemens, R.; Mendes, A.V.A.; Souza, A.R.; Silveira, M.B.V.; da Guarda, S.N.F.; de Nobrega, M.M.; Pinto, M.I.D.M.; Gonzalez, I.G.S.; et al. Heterologous versus homologous COVID-19 booster vaccination in previous recipients of two doses of CoronaVac COVID-19 vaccine in Brazil (RHH-001): A phase 4, non-inferiority, single blind, randomised study. Lancet 2022, 399, 521–529. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paula, N.M.; Conzentino, M.S.; Gonçalves, A.C.A.; da Silva, R.; Weissheimer, K.V.; Kluge, C.H.S.; Marins, P.H.S.A.; Camargo, H.S.C.; Farias, L.R.P.; Sant’Ana, T.P.; et al. Population-Based Analysis of the Immunoglobulin G Response to Different COVID-19 Vaccines in Brazil. Vaccines 2023, 11, 21. https://doi.org/10.3390/vaccines11010021
Paula NM, Conzentino MS, Gonçalves ACA, da Silva R, Weissheimer KV, Kluge CHS, Marins PHSA, Camargo HSC, Farias LRP, Sant’Ana TP, et al. Population-Based Analysis of the Immunoglobulin G Response to Different COVID-19 Vaccines in Brazil. Vaccines. 2023; 11(1):21. https://doi.org/10.3390/vaccines11010021
Chicago/Turabian StylePaula, Nigella M., Marcelo S. Conzentino, Ana C. A. Gonçalves, Renata da Silva, Karin V. Weissheimer, Carlos H. S. Kluge, Paulo H. S. A. Marins, Haxley S. C. Camargo, Lucas R. P. Farias, Thamyres P. Sant’Ana, and et al. 2023. "Population-Based Analysis of the Immunoglobulin G Response to Different COVID-19 Vaccines in Brazil" Vaccines 11, no. 1: 21. https://doi.org/10.3390/vaccines11010021
APA StylePaula, N. M., Conzentino, M. S., Gonçalves, A. C. A., da Silva, R., Weissheimer, K. V., Kluge, C. H. S., Marins, P. H. S. A., Camargo, H. S. C., Farias, L. R. P., Sant’Ana, T. P., Vargas, L. R., Aldrighi, J. D., Lima, Ê. S., Jacotenski, G. T., Pedrosa, F. O., Gonçalves, A. G., Joucoski, E., & Huergo, L. F. (2023). Population-Based Analysis of the Immunoglobulin G Response to Different COVID-19 Vaccines in Brazil. Vaccines, 11(1), 21. https://doi.org/10.3390/vaccines11010021