
Phytoestrogen β-Sitosterol Exhibits Potent In Vitro Antiviral Activity against Influenza A Viruses

 , ,
, ,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
2.2. Phytochemicals
2.3. Cytotoxicity and Antiviral Assay
2.4. Plaque Reduction Assay (PRA)
2.5. Stage(s) of the Antiviral Action
2.6. Data Collection and Heatmap Construction
2.7. In Silico Docking Studies
2.7.1. Protein Preparation
2.7.2. Ligand Preparation
2.7.3. Docking Setup and Validation of Docking Protocol
2.8. Statistical Analysis
3. Results
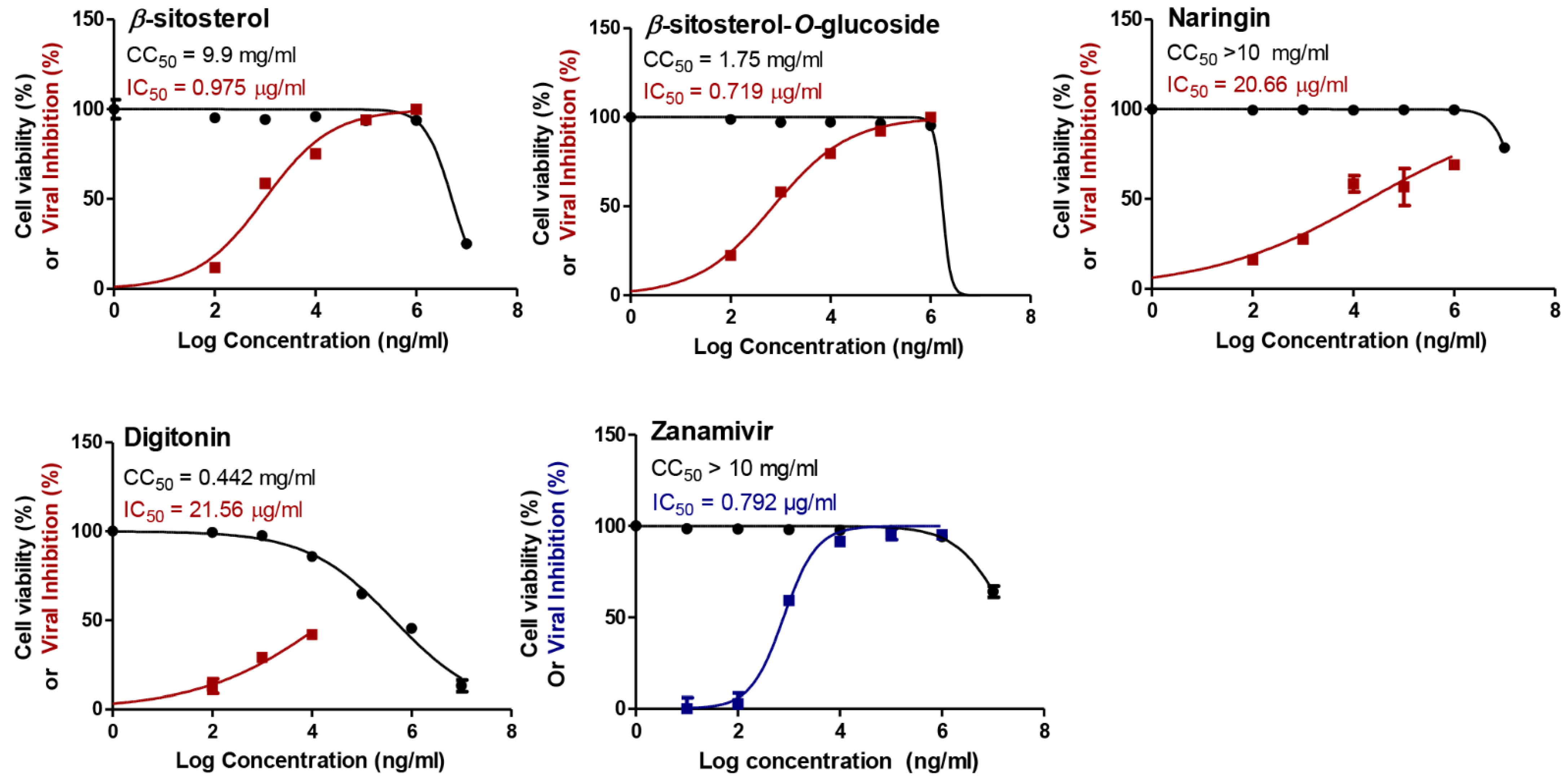
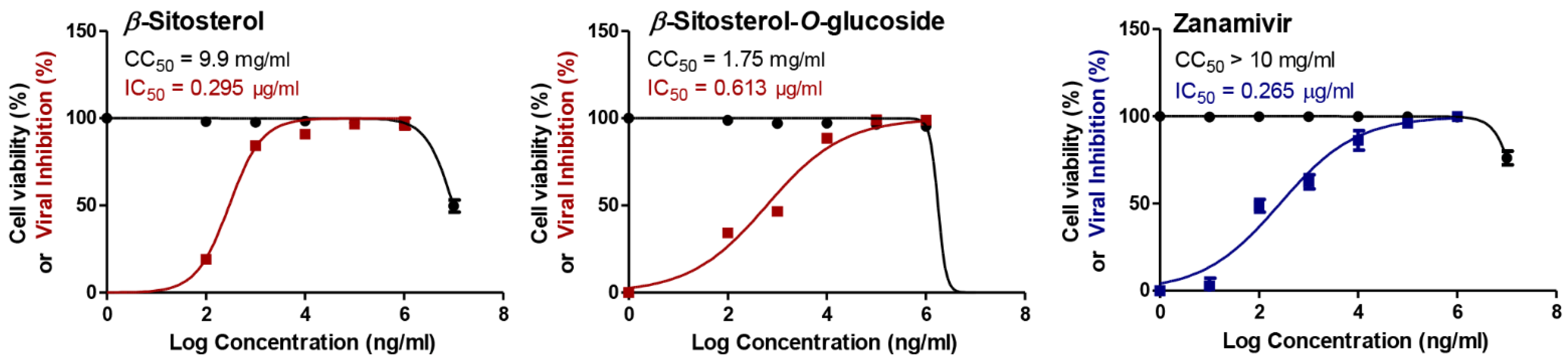
3.1. Cytotoxicity and Antiviral Potential of the Investigated Compounds
3.2. Viral titer Reduction in a Concentration-Dependent Manner
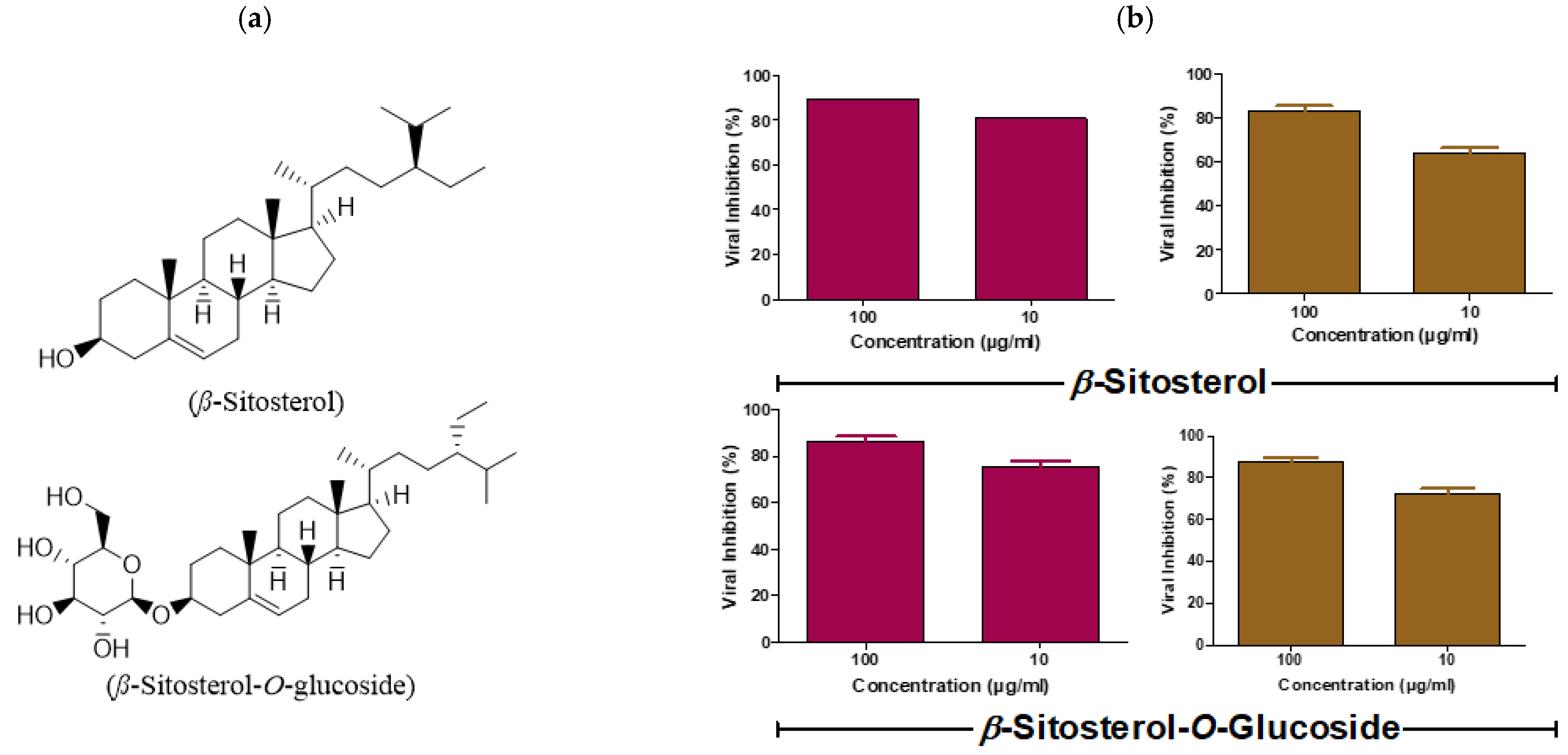
3.3. β-Sitosterol and β-Sitosterol-O-Glucoside affects IAV at Multiple Stages of the Virus Replication Cycle
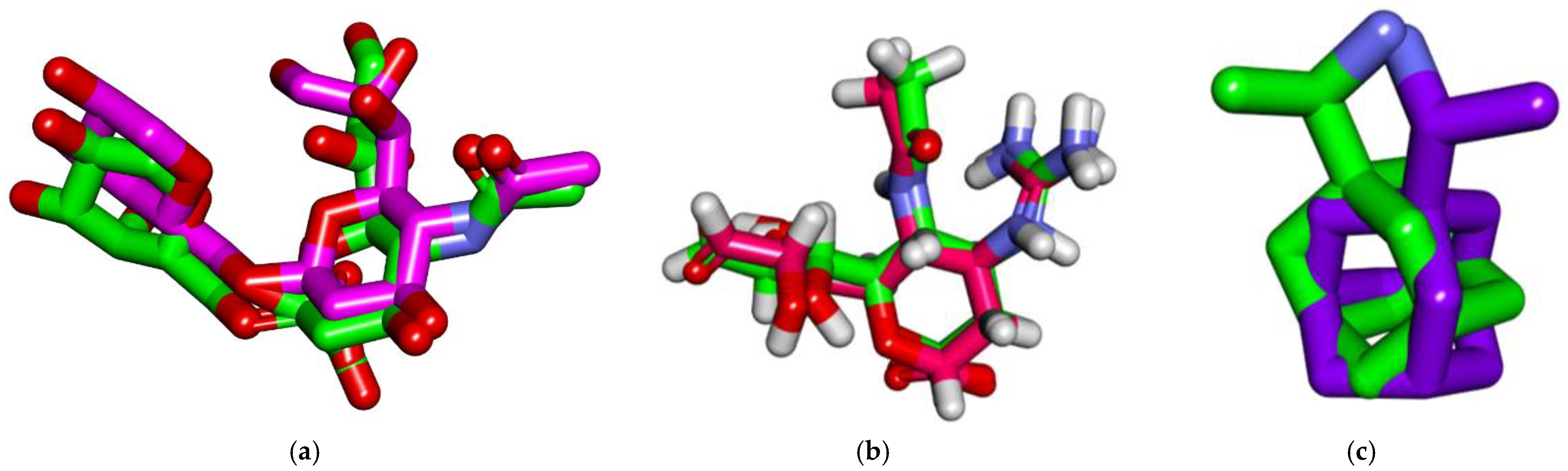
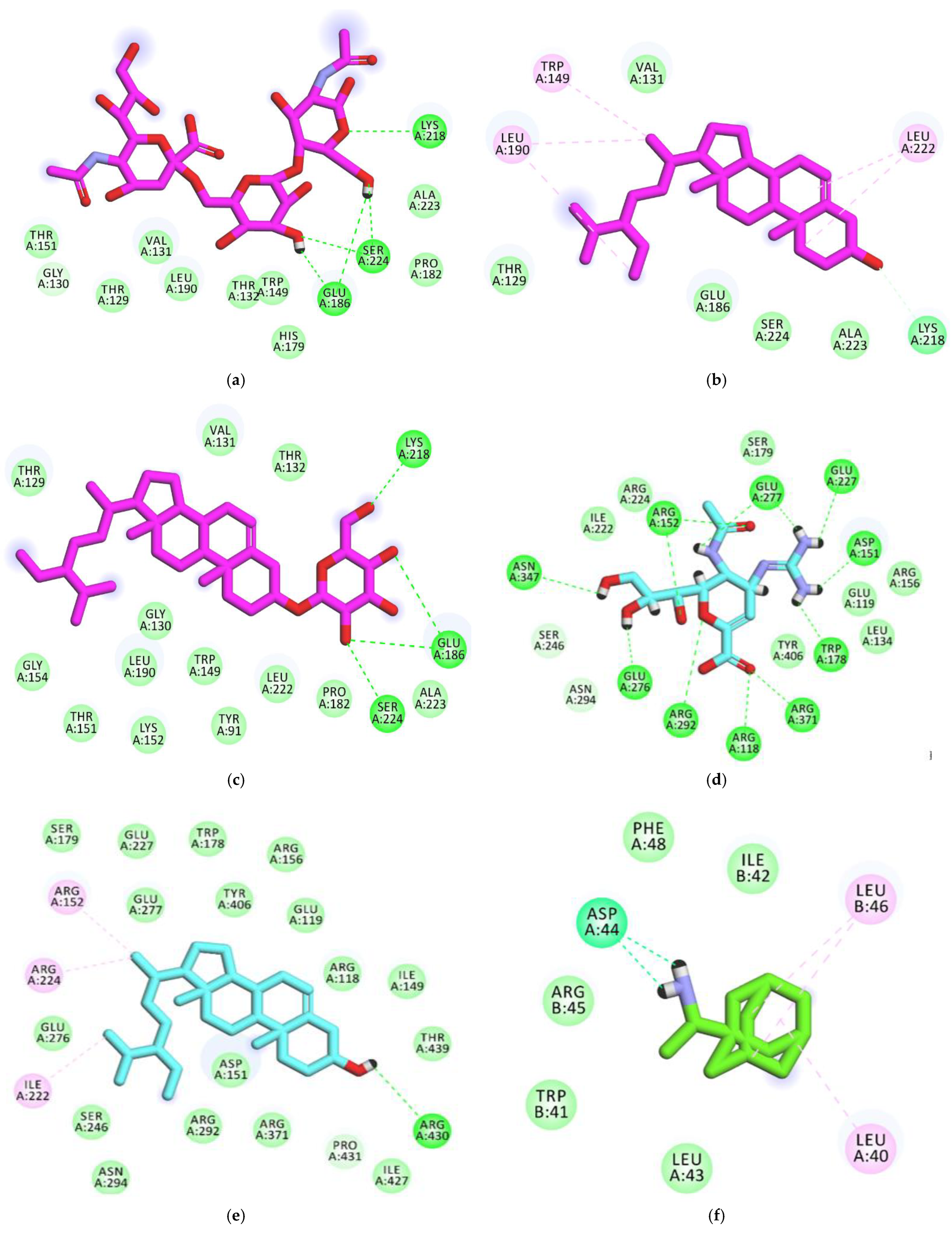
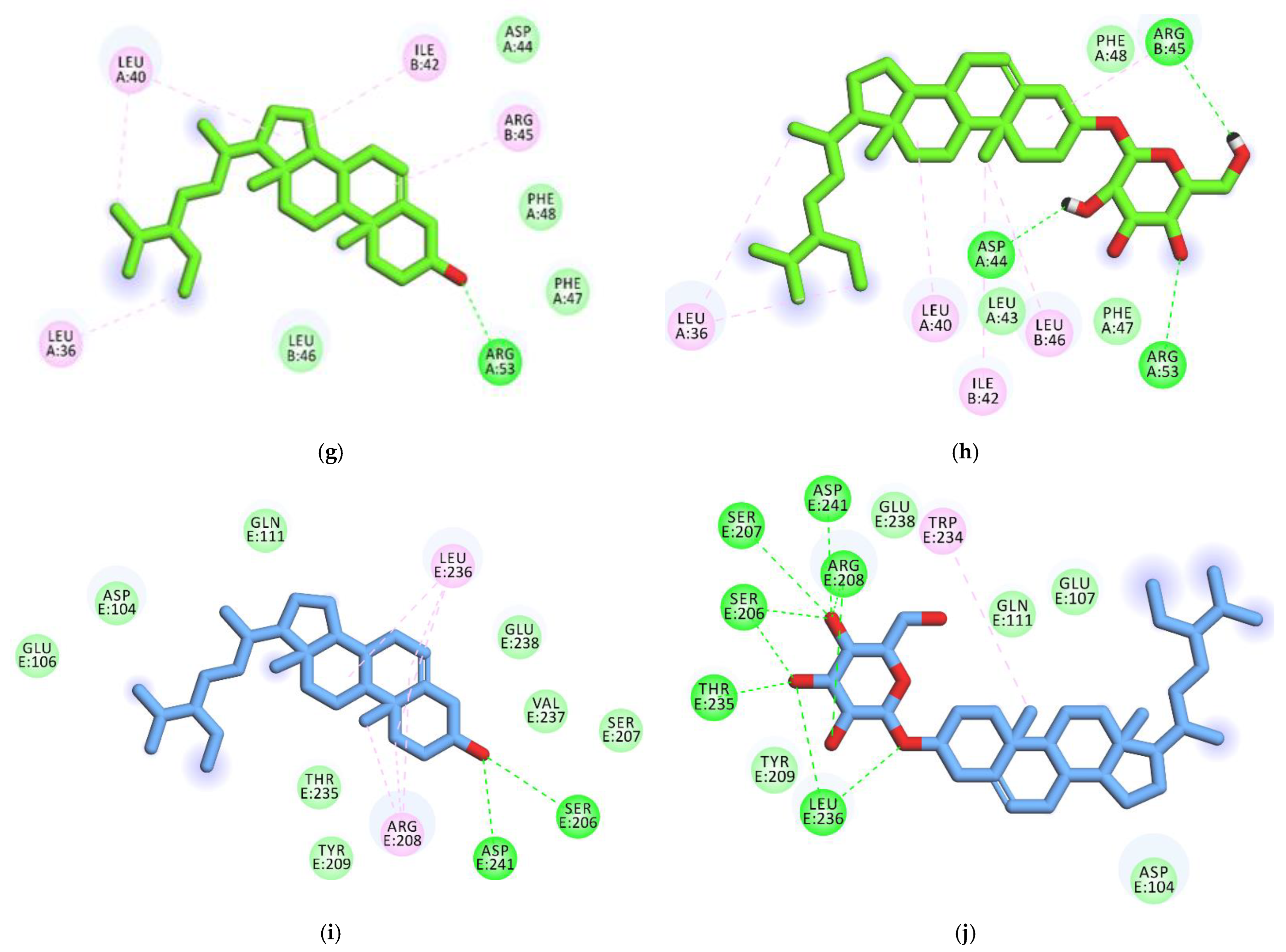
3.4. Docking studies
3.4.1. Validation
3.4.2. Docking Studies against Influenza Hemagglutinin H1 Mutant DH1E
3.4.3. Docking Studies against Influenza A/H1N1 Neuraminidase
3.4.4. Docking Studies against Influenza Proton Channel M2
3.4.5. Docking Studies against Hemagglutinin Head Epitope of Influenza A/H1N1 Virus
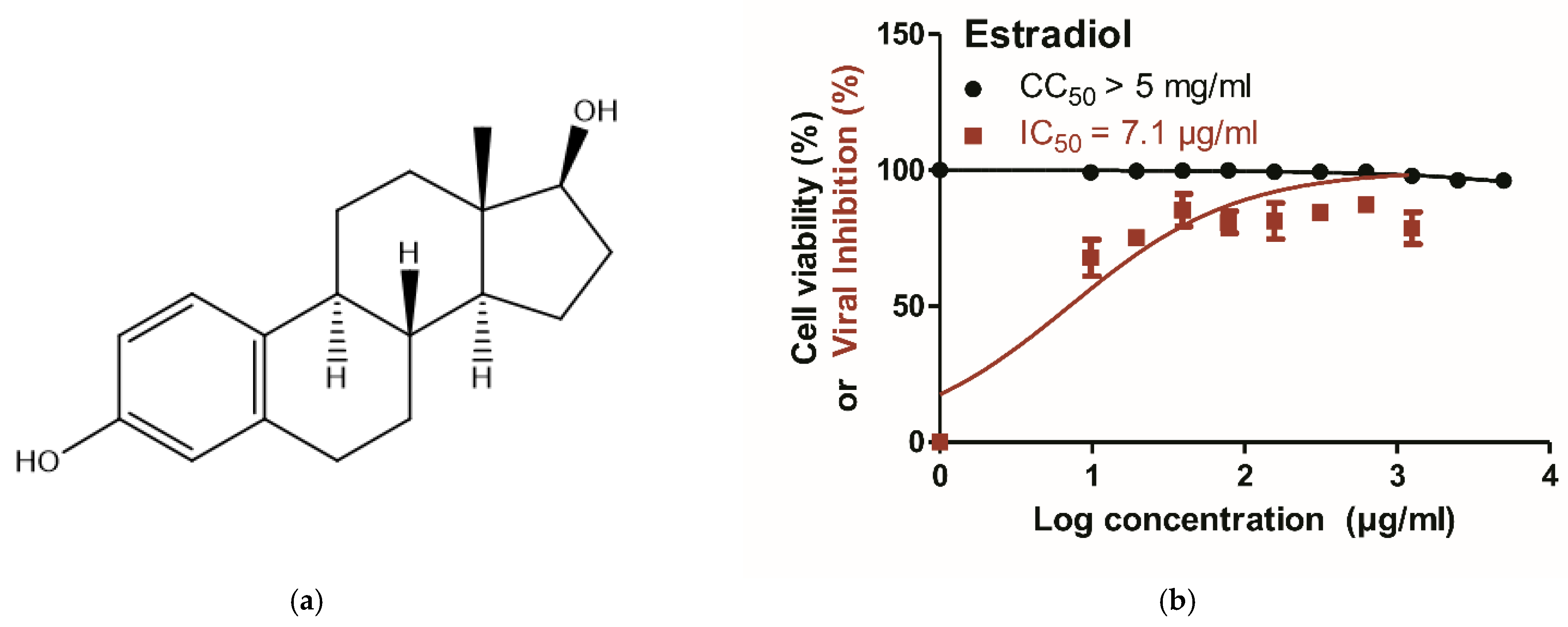
3.5. Phytoestrogen β-Sitosterol Is Likely to Hinder IAV Replication in an Estrogen-Like Mode
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naghavi, M.; Abajobir, A.A.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abera, S.F.; Aboyans, V.; Adetokunboh, O.; Afshin, A.; Agrawal, A. Global, regional, and national age-sex specific mortality for 264 causes of death, 1980–2016: A systematic analysis for the global burden of disease study 2016. Lancet 2017, 390, 1151–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfes, M.A.; Foppa, I.M.; Garg, S.; Flannery, B.; Brammer, L.; Singleton, J.A.; Burns, E.; Jernigan, D.; Olsen, S.J.; Bresee, J. Annual estimates of the burden of seasonal influenza in the united states: A tool for strengthening influenza surveillance and preparedness. Influenza Other Respir. Viruses 2018, 12, 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, L.J.; Im, C.; Dong, H.; Lee, B.E.; Talbot, J.; Meurer, D.P.; Mukhi, S.N.; Drews, S.J.; Yasui, Y. Influenza-like illness-related emergency department visits: Christmas and new year holiday peaks and relationships with laboratory-confirmed respiratory virus detections, edmonton, alberta, 2004–2014. Influenza Other Respir. Viruses 2017, 11, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Kesson, A.M. Respiratory virus infections. Paediatr. Respir. Rev. 2007, 8, 240–248. [Google Scholar] [CrossRef] [PubMed]
- White, D.O.; Brown, L.E. Respiratory viruses. In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 1999; pp. 1488–1496. [Google Scholar] [CrossRef]
- Al-Karmalawy, A.A.; Soltane, R.; Abo Elmaaty, A.; Tantawy, M.A.; Antar, S.A.; Yahya, G.; Chrouda, A.; Pashameah, R.A.; Mustafa, M.; Abu Mraheil, M.; et al. Coronavirus disease (COVID-19) control between drug repurposing and vaccination: A comprehensive overview. Vaccines 2021, 9, 1317. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic potential of influenza a viruses: A comprehensive overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, P.; Neumann, G.; Kawaoka, Y. Orthomyxoviruses. Fields virology. In Fields Virology, 5th ed.; Lippincott-Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 1, pp. 1691–1740. [Google Scholar]
- Taubenberger, J.K.; Morens, D.M. The pathology of influenza virus infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Product of natural evolution (sars, mers, and SARS-CoV-2); deadly diseases, from sars to SARS-CoV-2. Hum. Vaccin. Immunother. 2021, 17, 62–83. [Google Scholar] [CrossRef]
- Putri, W.C.; Muscatello, D.J.; Stockwell, M.S.; Newall, A.T. Economic burden of seasonal influenza in the united states. Vaccine 2018, 36, 3960–3966. [Google Scholar] [CrossRef]
- Okomo, U.; Idoko, O.T.; Kampmann, B. The burden of viral respiratory infections in young children in low-resource settings. Lancet Glob. Health 2020, 8, e454–e455. [Google Scholar] [CrossRef]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Sellers, S.A.; Hagan, R.S.; Hayden, F.G.; Fischer, W.A., 2nd. The hidden burden of influenza: A review of the extra-pulmonary complications of influenza infection. Influenza Other Respir Viruses 2017, 11, 372–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Prim. 2018, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.N.; Fearns, R. Genetic instability of rna viruses. In Genome Stability; Academic Press: Cambridge, MA, USA, 2016; pp. 21–35. [Google Scholar] [CrossRef]
- Wat, D. The common cold: A review of the literature. Eur. J. Intern. Med. 2004, 15, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.F.; Criado, M.F.; Arruda, E. Respiratory viruses. In Encyclopedia of Microbiology; Academic Press: Cambridge, MA, USA, 2009; pp. 500–518. [Google Scholar] [CrossRef]
- Musarra-Pizzo, M.; Pennisi, R.; Ben-Amor, I.; Mandalari, G.; Sciortino, M.T. Antiviral activity exerted by natural products against human viruses. Viruses 2021, 13, 828. [Google Scholar] [CrossRef]
- Hegazy, A.; Mostafa, I.; Elshaier, Y.A.M.M.; Mahmoud, S.H.; Abo Shama, N.M.; Shehata, M.; Yahya, G.; Nasr, N.F.; El-Halawany, A.M.; Ali, M.A.; et al. Robust antiviral activity of santonica flower extract (artemisia cina) against avian and human influenza a viruses: In vitro and chemoinformatic studies. ACS Omega 2022, 7, 41212–41223. [Google Scholar] [CrossRef]
- Lin, L.T.; Hsu, W.C.; Lin, C.C. Antiviral natural products and herbal medicines. J. Tradit. Complement. Med. 2014, 4, 24–35. [Google Scholar] [CrossRef]
- Li, H.; Sun, J.; Xiao, S.; Zhang, L.; Zhou, D. Triterpenoid-mediated inhibition of virus–host interaction: Is now the time for discovering viral entry/release inhibitors from nature? J. Med. Chem. 2020, 63, 15371–15388. [Google Scholar] [CrossRef]
- Mostafa, A.; Mahmoud, S.H.; Shehata, M.; Müller, C.; Kandeil, A.; El-Shesheny, R.; Nooh, H.Z.; Kayali, G.; Ali, M.A.; Pleschka, S. Pa from a recent h9n2 (g1-like) avian influenza a virus (aiv) strain carrying lysine 367 confers altered replication efficiency and pathogenicity to contemporaneous h5n1 in mammalian systems. Viruses 2020, 12, 1046. [Google Scholar] [CrossRef]
- Petersen, H.; Mostafa, A.; Tantawy, M.A.; Iqbal, A.A.; Hoffmann, D.; Tallam, A.; Selvakumar, B.; Pessler, F.; Beer, M.; Rautenschlein, S. Ns segment of a 1918 influenza a virus-descendent enhances replication of h1n1pdm09 and virus-induced cellular immune response in mammalian and avian systems. Front. Microbiol. 2018, 9, 526. [Google Scholar] [CrossRef]
- Sarg, T.M.; El-Domiaty, M.M.; Ateya, A.-M.M.; EI-Dahmy, S.I.; EI-Shazly, A.M. Phytochemical investigation of cen-taurea eryngioides lam. Growing in egypt. Alex. J. Pharm. Sci. 1993, 7, 50–54. [Google Scholar]
- Hamdan, D.; El-Readi, M.Z.; Tahrani, A.; Herrmann, F.; Kaufmann, D.; Farrag, N.; El-Shazly, A.; Wink, M. Chemical composition and biological activity of citrus jambhiri lush. Food Chem. 2011, 127, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Barrera, J.; Martin-Castillo, B.; Buxó, M.; Brunet, J.; Encinar, J.A.; Menendez, J.A. Silibinin and SARS-CoV-2: Dual targeting of host cytokine storm and virus replication machinery for clinical management of COVID-19 patients. J. Clin. Med. 2020, 9, 1770. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.-P.; Wu, L.-Q.; Li, R.; Zhao, X.-F.; Wan, Q.-Y.; Chen, X.-X.; Li, W.-Z.; Wang, G.-F.; Li, K.-S. Identification of 23-(s)-2-amino-3-phenylpropanoyl-silybin as an antiviral agent for influenza a virus infection in vitro and in vivo. Antimicrob. Agents Chemother. 2013, 57, 4433–4443. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Yang, Y.-Z.; Chen, J.-X.; Tang, Y.-Z. Inhibition of pro-inflammatory mediators in raw264. 7 cells by 7-hydroxyflavone and 7, 8-dihydroxyflavone. J. Pharm. Pharmacol. 2017, 69, 865–874. [Google Scholar] [CrossRef]
- Wang, J.; Su, H.; Zhang, T.; Du, J.; Cui, S.; Yang, F.; Jin, Q. Inhibition of enterovirus 71 replication by 7-hydroxyflavone and diisopropyl-flavon7-yl phosphate. PLoS ONE 2014, 9, e92565. [Google Scholar] [CrossRef]
- Lalani, S.; Poh, C.L. Flavonoids as antiviral agents for enterovirus a71 (ev-a71). Viruses 2020, 12, 184. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, P.; Neuner, A.; Nolkemper, S.; Zundel, C.; Nowack, H.; Sensch, K.H.; Reichling, J. Antiviral activity and mode of action of propolis extracts and selected compounds. Phytother. Res. 2010, 24, S20–S28. [Google Scholar] [CrossRef]
- Le Lee, J.; Loe, M.W.C.; Lee, R.C.H.; Chu, J.J.H. Antiviral activity of pinocembrin against zika virus replication. Antivir. Res. 2019, 167, 13–24. [Google Scholar] [CrossRef]
- González-Búrquez, M.d.J.; González-Díaz, F.R.; García-Tovar, C.G.; Carrillo-Miranda, L.; Soto-Zárate, C.I.; Canales-Martínez, M.M.; Penieres-Carrillo, J.G.; Crúz-Sánchez, T.A.; Fonseca-Coronado, S. Comparison between in vitro antiviral effect of mexican propolis and three commercial flavonoids against canine distemper virus. Evid. -Based Complement. Altern. Med. 2018, 2018, 7092416. [Google Scholar] [CrossRef] [Green Version]
- Guler, H.I.; Tatar, G.; Yildiz, O.; Belduz, A.O.; Kolayli, S. Investigation of potential inhibitor properties of ethanolic propolis extracts against ace-ii receptors for COVID-19 treatment by molecular docking study. Arch. Microbiol. 2021, 203, 3557–3564. [Google Scholar] [CrossRef]
- Alalaiwe, A.; Lin, C.-F.; Hsiao, C.-Y.; Chen, E.-L.; Lin, C.-Y.; Lien, W.-C.; Fang, J.-Y. Development of flavanone and its derivatives as topical agents against psoriasis: The prediction of therapeutic efficiency through skin permeation evaluation and cell-based assay. Int. J. Pharm. 2020, 581, 119256. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, G.; Coronel, O.; Trabuco, A.; Bazán, D.; Russo, R.; Alvarenga, N.; Aquino, V. Steroidal saponins from the roots of solanum sisymbriifolium lam.(solanaceae) have inhibitory activity against dengue virus and yellow fever virus. Braz. J. Med. Biol. Res. 2021, 54, e10240. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.H.; Joshi, S.V. Anticancer activity of saponin isolated from albizia lebbeck using various in vitro models. J. Ethnopharmacol. 2019, 231, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.N.T.; Sakoff, J.A.; Vuong, Q.V.; Bowyer, M.C.; Scarlett, C.J. Phytochemical, antioxidant, anti-proliferative and antimicrobial properties of catharanthus roseus root extract, saponin-enriched and aqueous fractions. Mol. Biol. Rep. 2019, 46, 3265–3273. [Google Scholar] [CrossRef]
- Mair, C.E.; Grienke, U.; Wilhelm, A.; Urban, E.; Zehl, M.; Schmidtke, M.; Rollinger, J.M. Anti-influenza triterpene saponins from the bark of burkea africana. J. Nat. Prod. 2018, 81, 515–523. [Google Scholar] [CrossRef]
- Wang, W.-H.; Chuang, H.-Y.; Chen, C.-H.; Chen, W.-K.; Hwang, J.-J. Lupeol acetate ameliorates collagen-induced arthritis and osteoclastogenesis of mice through improvement of microenvironment. Biomed. Pharmacother. 2016, 79, 231–240. [Google Scholar] [CrossRef]
- Pereira Beserra, F.; Sérgio Gushiken, L.F.; Vieira, A.J.; Augusto Bérgamo, D.; Luísa Bérgamo, P.; Oliveira de Souza, M.; Alberto Hussni, C.; Kiomi Takahira, R.; Henrique Nóbrega, R.; Monteiro Martinez, E.R. From inflammation to cutaneous repair: Topical application of lupeol improves skin wound healing in rats by modulating the cytokine levels, nf-κb, ki-67, growth factor expression, and distribution of collagen fibers. Int. J. Mol. Sci. 2020, 21, 4952. [Google Scholar] [CrossRef]
- Smirnova, I.E.; Kazakova, O.B. Structure–anti-influenza type a activity relationship among a series of nitrogen lupane triterpenoids. Nat. Prod. Commun. 2018, 13, 1934578X1801301008. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Leal, J.; Ponce-García, N.; Escalante-Aburto, A. Recent evidence of the beneficial effects associated with glucuronic acid contained in kombucha beverages. Curr. Nutr. Rep. 2020, 9, 163–170. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, Q.; Li, S.; Li, C.; Liao, S.; Yang, X.; Zhou, R.; Zhu, Y.; Teng, L.; Chen, H. Antiviral activity against porcine epidemic diarrhea virus of pogostemon cablin polysaccharide. J. Ethnopharmacol. 2020, 259, 113009. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Si, L.; Ji, S.; Wang, H.; Fang, X.-m.; Yu, L.-y.; Li, R.-y.; Liang, L.-n.; Zhou, D.; Ye, M. Uralsaponins m–y, antiviral triterpenoid saponins from the roots of glycyrrhiza uralensis. J. Nat. Prod. 2014, 77, 1632–1643. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Sun, L.; Zou, S.; Chen, J.; Mao, H.; Zhang, Y.; Liao, N.; Zhang, R. Antiviral effects of houttuynia cordata polysaccharide extract on murine norovirus-1 (mnv-1)—A human norovirus surrogate. Molecules 2019, 24, 1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, W.; Huang, J.; Niu, X.; Shang, H.; Sha, Z.; Miao, Y.; Wang, H.; Chen, R.; Wei, K.; Zhu, R. Screening active fractions from pinus massoniana pollen for inhibiting alv-j replication and their structure activity relationship investigation. Vet. Microbiol. 2021, 252, 108908. [Google Scholar] [CrossRef] [PubMed]
- Chun, B.K.; Schinazi, R.F.; Cheng, Y.-C.; Chu, C.K. Synthesis of 2′, 3′-dideoxy-3′-fluoro-l-ribonucleosides as potential antiviral agents from d-sorbitol. Carbohydr. Res. 2000, 328, 49–59. [Google Scholar] [CrossRef]
- Zhang, H.; Saravanan, K.M.; Yang, Y.; Hossain, M.; Li, J.; Ren, X.; Pan, Y.; Wei, Y. Deep learning based drug screening for novel coronavirus 2019-ncov. Interdiscip. Sci. Comput. Life Sci. 2020, 12, 368–376. [Google Scholar] [CrossRef]
- Maiyoa, F.; Moodley, R.; Singh, M. Phytochemistry, cytotoxicity and apoptosis studies of β-sitosterol-3-oglucoside and β-amyrin from prunus africana. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.-x.; Li, J.; Liang, X.-l.; Pan, X.-p.; Hao, Y.-b.; Xie, P.-f.; Jiang, H.-m.; Yang, Z.-f.; Zhong, N.-s. Β-sitosterol ameliorates influenza a virus-induced proinflammatory response and acute lung injury in mice by disrupting the cross-talk between rig-i and ifn/stat signaling. Acta Pharmacol. Sin. 2020, 41, 1178–1196. [Google Scholar] [CrossRef]
- Chen, C.; Shen, J.-L.; Liang, C.-S.; Sun, Z.-C.; Jiang, H.-F. First discovery of beta-sitosterol as a novel antiviral agent against white spot syndrome virus. Int. J. Mol. Sci. 2022, 23, 10448. [Google Scholar] [CrossRef]
- Khan, S.L.; Siddiqui, F.A. Beta-sitosterol: As immunostimulant, antioxidant and inhibitor of SARS-CoV-2 spike glycoprotein. Arch. Pharmacol. Ther. 2020, 2, 12–16. [Google Scholar]
- Zainab, B.; Ayaz, Z.; Alwahibi, M.S.; Khan, S.; Rizwana, H.; Soliman, D.W.; Alawaad, A.; Abbasi, A.M. In-silico elucidation of moringa oleifera phytochemicals against diabetes mellitus. Saudi J. Biol. Sci. 2020, 27, 2299–2307. [Google Scholar] [CrossRef] [PubMed]
- Behloul, N.; Baha, S.; Guo, Y.; Yang, Z.; Shi, R.; Meng, J. In silico identification of strong binders of the SARS-CoV-2 receptor-binding domain. Eur. J. Pharmacol. 2021, 890, 173701. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Huang, X.; Liao, T.; Li, G.; Yu, X.; You, Y.; Huang, Y. Daucosterol induces autophagic-dependent apoptosis in prostate cancer via jnk activation. Biosci. Trends 2019, 13, 160–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laverdière, I.; Boileau, M.; Neumann, A.L.; Frison, H.; Mitchell, A.; Ng, S.W.; Wang, J.C.; Minden, M.D.; Eppert, K. Leukemic stem cell signatures identify novel therapeutics targeting acute myeloid leukemia. Blood Cancer J. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Yang, C.-W.; Chang, H.-Y.; Lee, Y.-Z.; Hsu, H.-Y.; Lee, S.-J. The cardenolide ouabain suppresses coronaviral replication via augmenting a na+/k+-atpase-dependent pi3k_pdk1 axis signaling. Toxicol. Appl. Pharmacol. 2018, 356, 90–97. [Google Scholar] [CrossRef]
- Cho, J.; Lee, Y.J.; Kim, J.H.; Kim, S.S.; Choi, B.-S.; Choi, J.-H. Antiviral activity of digoxin and ouabain against SARS-CoV-2 infection and its implication for COVID-19. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Katzen, J.; Hurtado, J.; Lecuona, E.; Sznajder, J.I. B32 viral infection of the airway: The cardioactive glycoside ouabain inhibits influenza a viral replication. Am. J. Respir. Crit. Care Med. 2014, 189, 1. [Google Scholar]
- Fan, H.Y.; Heerklotz, H. Digitonin does not flip across cholesterol-poor membranes. J. Colloid Interface Sci. 2017, 504, 283–293. [Google Scholar] [CrossRef]
- Orczyk, M.; Wojciechowski, K.; Brezesinski, G. Disordering effects of digitonin on phospholipid monolayers. Langmuir 2017, 33, 3871–3881. [Google Scholar] [CrossRef]
- Zhang, J.-W.; Wang, H.; Liu, J.; Ma, L.; Hua, R.-H.; Bu, Z.-G. Generation of a stable gfp-reporter zika virus system for high-throughput screening of zika virus inhibitors. Virol. Sin. 2021, 36, 476–489. [Google Scholar] [CrossRef]
- Inoue, Y.; Hasegawa, S.; Yamada, T.; Date, Y.; Mizutani, H.; Nakata, S.; Matsunaga, K.; Akamatsu, H. Analysis of the effects of hydroquinone and arbutin on the differentiation of melanocytes. Biol. Pharm. Bull. 2013, 36, 1722–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baby, K.; Maity, S.; Mehta, C.H.; Suresh, A.; Nayak, U.Y.; Nayak, Y. Targeting SARS-CoV-2 main protease: A computational drug repurposing study. Arch. Med. Res. 2021, 52, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Van Hoof, L.; Totte, J.; Corthout, J.; Pieters, L.; Mertens, F.; Vanden Berghe, D.; Vlietinck, A.; Dommisse, R.; Esmans, E. Plant antiviral agents, vi. Isolation of antiviral phenolic glucosides from populus cultivar beaupre by droplet counter-current chromatography. J. Nat. Prod. 1989, 52, 875–878. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Nishigaya, K.; Takami, K.; Uchikoshi, H.; Chen, I.-S.; Tsai, I.-L. Isolation of salicin derivatives from homalium c ochinchinensis and their antiviral activities. J. Nat. Prod. 2004, 67, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Le, N.P.K.; Herz, C.; Gomes, J.V.D.; Förster, N.; Antoniadou, K.; Mittermeier-Kleßinger, V.K.; Mewis, I.; Dawid, C.; Ulrichs, C.; Lamy, E. Comparative anti-inflammatory effects of salix cortex extracts and acetylsalicylic acid in SARS-CoV-2 peptide and lps-activated human in vitro systems. Int. J. Mol. Sci. 2021, 22, 6766. [Google Scholar] [CrossRef] [PubMed]
- Nzuza, S.; Ndwandwe, D.E.; Owira, P.M. Naringin protects against hiv-1 protease inhibitors-induced pancreatic β-cell dysfunction and apoptosis. Mol. Cell. Endocrinol. 2016, 437, 1–10. [Google Scholar] [CrossRef]
- Chen, R.; Qi, Q.-L.; Wang, M.-T.; Li, Q.-Y. Therapeutic potential of naringin: An overview. Pharm. Biol. 2016, 54, 3203–3210. [Google Scholar] [CrossRef] [Green Version]
- Huseen, N.H.A. Docking study of naringin binding with COVID-19 main protease enzyme. Iraqi J. Pharm. Sci. 2020, 29, 231–238. [Google Scholar] [CrossRef]
- Nzuza, S.; Zondi, S.; Owira, P.M. Naringin prevents hiv-1 protease inhibitors-induced metabolic complications in vivo. PLoS ONE 2017, 12, e0183355. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.-H.; Rao, Y.K.; Tzeng, Y.-M. Inhibitory effects of flavonol glycosides from cinnamomum osmophloeum on inflammatory mediators in lps/ifn-γ-activated murine macrophages. Bioorganic Med. Chem. 2005, 13, 2381–2388. [Google Scholar] [CrossRef]
- Choi, J.-G.; Kim, Y.S.; Kim, J.H.; Chung, H.-S. Antiviral activity of ethanol extract of geranii herba and its components against influenza viruses via neuraminidase inhibition. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Bhardwaj, V.K.; Sharma, J.; Purohit, R.; Kumar, S. In-silico evaluation of bioactive compounds from tea as potential SARS-CoV-2 nonstructural protein 16 inhibitors. J. Tradit. Complement. Med. 2022, 12, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Jorge, A.P.; Horst, H.; de Sousa, E.; Pizzolatti, M.G.; Silva, F.R.M.B. Insulinomimetic effects of kaempferitrin on glycaemia and on 14c-glucose uptake in rat soleus muscle. Chem. -Biol. Interact. 2004, 149, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-L.; He, Y.; Ji, L.-L.; Wang, K.-Y.; Wang, Y.-L.; Chen, D.-F.; Geng, Y.; OuYang, P.; Lai, W.-M. Hepatoprotective potential of isoquercitrin against type 2 diabetes-induced hepatic injury in rats. Oncotarget 2017, 8, 101545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resham, K.; Khare, P.; Bishnoi, M.; Sharma, S.S. Neuroprotective effects of isoquercitrin in diabetic neuropathy via wnt/β-catenin signaling pathway inhibition. Biofactors 2020, 46, 411–420. [Google Scholar] [CrossRef]
- Gao, Z.; Luan, Y.; Yang, P.; Wang, L.; Zhang, H.; Jing, S.; Wang, L.; Wang, T.; Wang, D. Targeting staphylocoagulase with isoquercitrin protects mice from staphylococcus aureus–induced pneumonia. Appl. Microbiol. Biotechnol. 2020, 104, 3909–3919. [Google Scholar] [CrossRef]
- Cho, W.-K.; Yang, H.J.; Ma, J.Y. Lotus (nelumbo nucifera gaertn.) leaf water extracts suppress influenza a viral infection via inhibition of neuraminidase and hemagglutinin. J. Funct. Foods 2022, 91, 105019. [Google Scholar] [CrossRef]
- Ling, L.-j.; Lu, Y.; Zhang, Y.-y.; Zhu, H.-y.; Tu, P.; Li, H.; Chen, D.-f. Flavonoids from houttuynia cordata attenuate h1n1-induced acute lung injury in mice via inhibition of influenza virus and toll-like receptor signalling. Phytomedicine 2020, 67, 153150. [Google Scholar] [CrossRef]
- Semple, S.J.; Pyke, S.M.; Reynolds, G.D.; Flower, R.L. In vitro antiviral activity of the anthraquinone chrysophanic acid against poliovirus. Antivir. Res. 2001, 49, 169–178. [Google Scholar] [CrossRef]
- Narkhede, R.R.; Pise, A.V.; Cheke, R.S.; Shinde, S.D. Recognition of natural products as potential inhibitors of COVID-19 main protease (mpro): In-silico evidences. Nat. Prod. Bioprospecting 2020, 10, 297–306. [Google Scholar] [CrossRef]
- Dong, X.; Zeng, Y.; Liu, Y.; You, L.; Yin, X.; Fu, J.; Ni, J. Aloe-emodin: A review of its pharmacology, toxicity, and pharmacokinetics. Phytother. Res. 2020, 34, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-Y.; Wu, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the sars coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Marasini, B.P.; Rayamajhee, B.; Bhattarai, B.R.; Lamichhane, G.; Khadayat, K.; Adhikari, A.; Khanal, S.; Parajuli, N. Potential roles of medicinal plants for the treatment of viral diseases focusing on covid-19: A review. Phytother. Res. 2021, 35, 1298–1312. [Google Scholar] [CrossRef] [PubMed]
- Robson, B. COVID-19 coronavirus spike protein analysis for synthetic vaccines, a peptidomimetic antagonist, and therapeutic drugs, and analysis of a proposed achilles’ heel conserved region to minimize probability of escape mutations and drug resistance. Comput. Biol. Med. 2020, 121, 103749. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Dong, W. Aloe-emodin induces endoplasmic reticulum stress-dependent apoptosis in colorectal cancer cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 6331. [Google Scholar] [CrossRef]
- Li, S.-W.; Yang, T.-C.; Lai, C.-C.; Huang, S.-H.; Liao, J.-M.; Wan, L.; Lin, Y.-J.; Lin, C.-W. Antiviral activity of aloe-emodin against influenza a virus via galectin-3 up-regulation. Eur. J. Pharmacol. 2014, 738, 125–132. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, N.; Zhong, Y.; Yang, Z.-q. Antiviral effect of emodin from rheum palmatum against coxsakievirus b5 and human respiratory syncytial virus in vitro. J. Huazhong Univ. Sci. Technol. [Med. Sci.] 2015, 35, 916–922. [Google Scholar] [CrossRef]
- Hsu, C.-L.; Yen, G.-C. Effects of flavonoids and phenolic acids on the inhibition of adipogenesis in 3t3-l1 adipocytes. J. Agric. Food Chem. 2007, 55, 8404–8410. [Google Scholar] [CrossRef]
- Santos, P.M.; Vieira, A.J. Antioxidising activity of cinnamic acid derivatives against oxidative stress induced by oxidising radicals. J. Phys. Org. Chem. 2013, 26, 432–439. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, R. Efficacy of trans-2-hydroxycinnamic acid against trichlorfon-induced oxidative stress in wistar rats. Toxicol. Int. 2012, 19, 295. [Google Scholar]
- Enkhtaivan, G.; John, K.M.; Ayyanar, M.; Sekar, T.; Jin, K.-J.; Kim, D.H. Anti-influenza (h1n1) potential of leaf and stem bark extracts of selected medicinal plants of south india. Saudi J. Biol. Sci. 2015, 22, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Saini, D.; Yadav, D. Synthesis and evaluation of vanillin derivatives as antimicrobial agents. Turk. J. Pharm. Sci. 2018, 15, 57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, L.; Zhu, C.; Wu, Z.; Zhang, G.; Gan, X.; Liu, D.; Pan, J.; Hu, D.; Song, B. Facile synthesis of novel vanillin derivatives incorporating a bis (2-hydroxyethyl) dithhioacetal moiety as antiviral agents. J. Agric. Food Chem. 2017, 65, 4582–4588. [Google Scholar] [CrossRef] [PubMed]
- Dhanalakshmi, C.; Janakiraman, U.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M.; Kalandar, A.; Khan, M.A.S.; Guillemin, G.J. Vanillin attenuated behavioural impairments, neurochemical deficts, oxidative stress and apoptosis against rotenone induced rat model of parkinson’s disease. Neurochem. Res. 2016, 41, 1899–1910. [Google Scholar] [CrossRef]
- Hariono, M.; Abdullah, N.; Damodaran, K.; Kamarulzaman, E.E.; Mohamed, N.; Hassan, S.S.; Shamsuddin, S.; Wahab, H.A. Potential new h1n1 neuraminidase inhibitors from ferulic acid and vanillin: Molecular modelling, synthesis and in vitro assay. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Law, W.Y.; Asaruddin, M.R.; Bhawani, S.A.; Mohamad, S. Pharmacophore modelling of vanillin derivatives, favipiravir, chloroquine, hydroxychloroquine, monolaurin and tetrodotoxin as mpro inhibitors of severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). BMC Res. Notes 2020, 13, 1–8. [Google Scholar] [CrossRef]
- Mondal, D. Zanamivir☆. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Mahmoud, A.; Mostafa, A.; Al-Karmalawy, A.A.; Zidan, A.; Abulkhair, H.S.; Mahmoud, S.H.; Shehata, M.; Elhefnawi, M.M.; Ali, M.A. Telaprevir is a potential drug for repurposing against SARS-CoV-2: Computational and in vitro studies. Heliyon 2021, 7, e07962. [Google Scholar] [CrossRef]
- Mostafa, A.; Kandeil, A.; AMM Elshaier, Y.; Kutkat, O.; Moatasim, Y.; Rashad, A.A.; Shehata, M.; Gomaa, M.R.; Mahrous, N.; Mahmoud, S.H. Fda-approved drugs with potent in vitro antiviral activity against severe acute respiratory syndrome coronavirus 2. Pharmaceuticals 2020, 13, 443. [Google Scholar] [CrossRef]
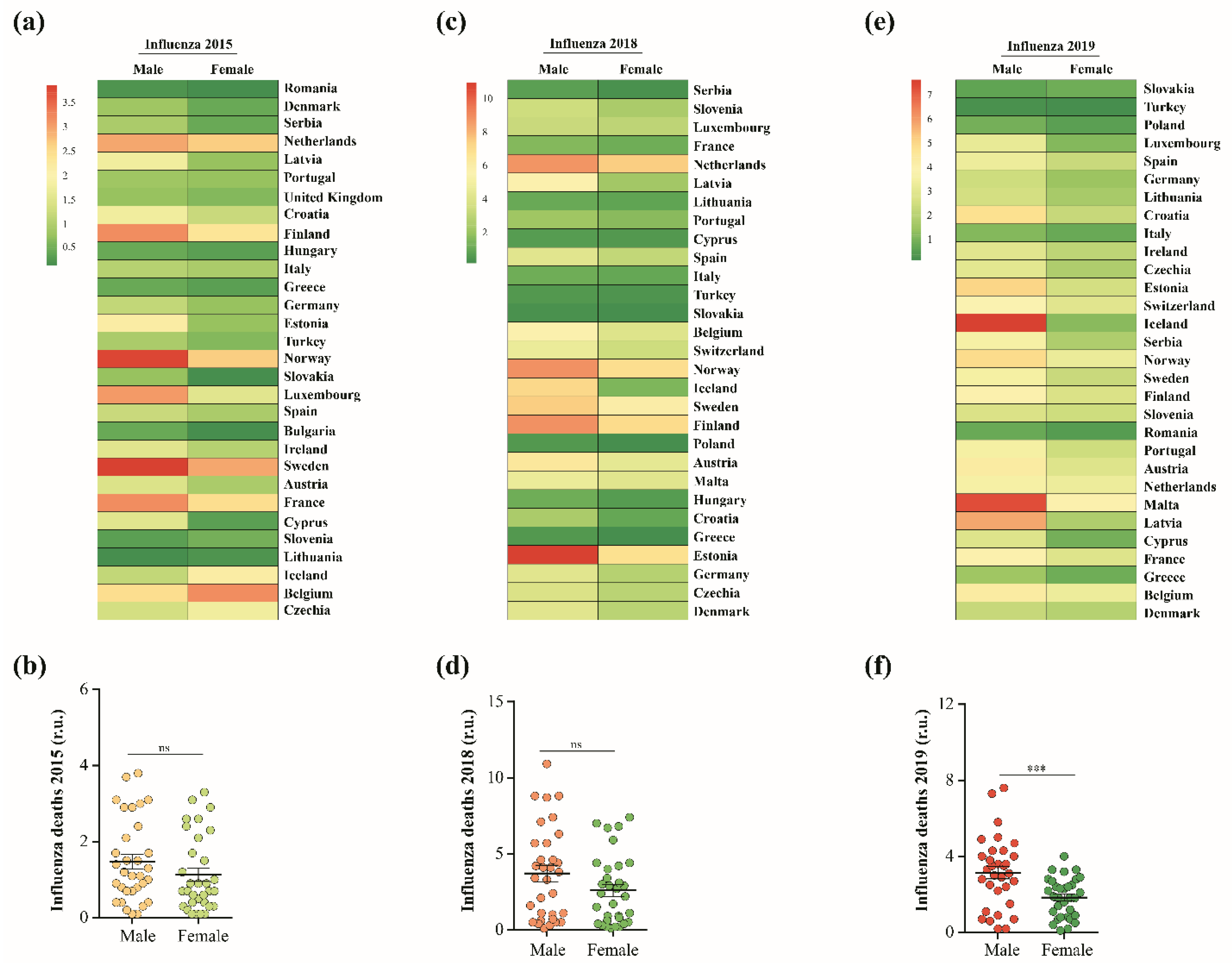
- EUROSTAT. Causes of Death—Standardised Death Rate by Nuts 2 Region of Residence. Available online: https://ec.europa.eu/eurostat/databrowser/view/hlth_cd_asdr2/default/table?lang=en (accessed on 14 January 2023).
- Metsalu, T.; Vilo, J. Clustvis: A web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Alesawy, M.S.; Abdallah, A.E.; Taghour, M.S.; Elkaeed, E.B.H. Eissa, I.; Metwaly, A.M. In silico studies of some isoflavonoids as potential candidates against COVID-19 targeting human ace2 (hace2) and viral main protease (mpro). Molecules 2021, 26, 2806. [Google Scholar] [CrossRef]
- Hagras, M.; El Deeb, M.A.; Elzahabi, H.S.; Elkaeed, E.B.; Mehany, A.B.; Eissa, I.H. Discovery of new quinolines as potent colchicine binding site inhibitors: Design, synthesis, docking studies, and anti-proliferative evaluation. J. Enzym. Inhib. Med. Chem. 2021, 36, 640–658. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, M.M.; Mahdy, H.A.; Alsaif, N.A.; Obaidullah, A.J.; Alkahtani, H.M.; Al-Mehizia, A.A.; Alsubaie, S.M.; Dahab, M.A.; Eissa, I.H. New bis ([1,2,4] triazolo)[4,3-a: 3′,4′-c] quinoxaline derivatives as vegfr-2 inhibitors and apoptosis inducers: Design, synthesis, in silico studies, and anticancer evaluation. Bioorganic Chem. 2021, 112, 104949. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, L.; Liu, Z.; Wang, R. Comparative assessment of scoring functions on an updated benchmark: 2. Evaluation methods and general results. J. Chem. Inf. Model. 2014, 54, 1717–1736. [Google Scholar] [CrossRef]
- Pawar, S.S.; Rohane, S.H. Review on discovery studio: An important tool for molecular docking. Asian J. Res. Chem 2021, 14, 86–88. [Google Scholar] [CrossRef]
- Yousef, R.G.; Sakr, H.M.; Eissa, I.H.; Mehany, A.B.; Metwaly, A.M.; Elhendawy, M.A.; Radwan, M.M.; ElSohly, M.A.; Abulkhair, H.S.; El-Adl, K. New quinoxaline-2 (1 h)-ones as potential vegfr-2 inhibitors: Design, synthesis, molecular docking, admet profile and anti-proliferative evaluations. New J. Chem. 2021, 45, 16949–16964. [Google Scholar] [CrossRef]
- Alesawy, M.S.; Elkaeed, E.B.; Alsfouk, A.A.; Metwaly, A.M.; Eissa, I.H. In silico screening of semi-synthesized compounds as potential inhibitors for SARS-CoV-2 papain-like protease: Pharmacophoric features, molecular docking, admet, toxicity and dft studies. Molecules 2021, 26, 6593. [Google Scholar] [CrossRef]
- Alwani, M.; Yassin, A.; Al-Zoubi, R.M.; Aboumarzouk, O.M.; Nettleship, J.; Kelly, D.; Al-Qudimat, A.R.; Shabsigh, R. Sex-based differences in severity and mortality in COVID-19. Rev. Med. Virol. 2021, 31, e2223. [Google Scholar] [CrossRef]
- Owen, L.; Laird, K.; Shivkumar, M. Antiviral plant-derived natural products to combat rna viruses: Targets throughout the viral life cycle. Lett. Appl. Microbiol. 2022, 75, 476–499. [Google Scholar] [CrossRef]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Tai, W.; Wang, N.; Li, X.; Jiang, S.; Debnath, A.K.; Du, L.; Chen, S. Identification of novel natural products as effective and broad-spectrum anti-zika virus inhibitors. Viruses 2019, 11, 1019. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zhu, Z.; Ding, Y.; Wu, W.; Yang, J.; Liu, S. An oligothiophene compound neutralized influenza a viruses by interfering with hemagglutinin. Biochim. Et Biophys. Acta (BBA) -Biomembr. 2018, 1860, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Luganini, A.; Terlizzi, M.E.; Catucci, G.; Gilardi, G.; Maffei, M.E.; Gribaudo, G. The cranberry extract oximacro® exerts in vitro virucidal activity against influenza virus by interfering with hemagglutinin. Front. Microbiol. 2018, 9, 1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makau, J.N.; Watanabe, K.; Kobayashi, N. Anti-influenza activity of alchemilla mollis extract: Possible virucidal activity against influenza virus particles. Drug Discov. Ther. 2013, 7, 189–195. [Google Scholar] [CrossRef] [Green Version]
- McAuley, J.L.; Gilbertson, B.P.; Trifkovic, S.; Brown, L.E.; McKimm-Breschkin, J.L. Influenza virus neuraminidase structure and functions. Front. Microbiol. 2019, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Lamb, R.A.; Zebedee, S.L.; Richardson, C.D. Influenza virus m2 protein is an integral membrane protein expressed on the infected-cell surface. Cell 1985, 40, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Stevenson, L.M.; Leonard, H.M.; Nieves-Puigdoller, K.; Clotfelter, E.D. Phytoestrogens β -sitosterol and genistein have limited effects on reproductive endpoints in a female fish, betta splendens. BioMed Res. Int. 2014, 2014, 681396. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.H.; Clausen, L.M.; Allred, K.F.; Almada, A.L.; Helferich, W.G. Β-sitosterol, β-sitosterol glucoside, and a mixture of β-sitosterol and β-sitosterol glucoside modulate the growth of estrogen-responsive breast cancer cells in vitro and in ovariectomized athymic mice. J. Nutr. 2004, 134, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Pandey, J.; Dev, K.; Chattopadhyay, S.; Kadan, S.; Sharma, T.; Maurya, R.; Sanyal, S.; Siddiqi, M.I.; Zaid, H.; Tamrakar, A.K. Beta-sitosterol-d-glucopyranoside mimics estrogenic properties and stimulates glucose utilization in skeletal muscle cells. Molecules 2021, 26, 3129. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CAS No. | Class | Reported Biological Activities | Reference |
|---|---|---|---|---|
| Silybin | 22888-70-6 | Flavonoids | Anti-inflammatory and antiviral | [27,28] |
| 7-Hydroxy flavone | 6665-86-7 | Flavonoids | Anti-inflammatory and antiviral | [29,30,31] |
| Pinocembrin | 480-39-7 | Flavonoids | Anti-inflammatory, antiallergic, antioxidant, anticarcinogenic, and antiviral | [32,33,34,35] |
| Flavanone | 487-26-3 | Flavonoids | Anti-inflammatory | [36] |
| Saponin | 8047-15-2 | Triterpene | Antimicrobial, anticancer, antioxidant, antitumor, and antiviral | [37,38,39,40] |
| Lupeol | 545-47-1 | Triterpene | Antioxidant and anti-inflammatory, Antiviral (Lupeol synthetic derivatives) | [41,42,43] |
| Glucuronic acid | 528-16-5 | Sugar acids | Antioxidant, hepatoprotective, and antiviral | [44,45] |
| Galacturonic acid | 9046-38-2 | Sugar acids | Antiviral (As a saponin component) | [45,46,47,48] |
| D-sorbitol | 50-70-4 | Sugar alcohol Carbohydrates | Antiviral and laxative. | [49,50] |
| β-sitosterol | 83-46-5 | Steroids | Antioxidant, anticarcinogenic, anti-inflammatory, and antiviral | [51,52,53,54] |
| β-sitosterol-O-glucoside | 474-58-8 | Steroids | Antidiabetic, anticancer and antiviral | [55,56,57] |
| Ouabain | 630-60-4 | Steroid cardiac glycosides | Anticancer and antiviral | [58,59,60,61] |
| Digitonin | 11024-24-1 | Steroid saponin glycosides | Lipid solubilizing and antiviral | [62,63,64] |
| Arbutin | 497-76-7 | Phenolic glycosides | Antimelanogenesis, antidiuretic, and antiviral | [65,66] |
| D- (-) salicin | 138-52-3 | Phenolic glycosides | Antiviral and anti-inflammatory. | [67,68,69] |
| Naringin | 10236-47-2 | Flavonoid glycosides | Anti-inflammatory, anticancer, and antiviral | [70,71,72,73] |
| Kaempferitrin | 482-38-2 | Flavonoid glycosides | Hypoglycemic, anti-inflammatory, and antiviral | [74,75,76,77] |
| Isoquercitrin | 482-35-9 | Flavonoid glycosides | Antioxidant, antipruritic, neuroprotective, antibacterial, hepatoprotective, anti-inflammatory, and antiviral | [78,79,80,81,82] |
| Chrysophanic acid | 481-74-3 | Anthraquinones | Antiviral | [83,84] |
| Aloe emodin | 481-72-1 | Anthraquinones | Antiviral, anticancer, anti-inflammatory, and antibacterial. | [85,86,87,88,89,90,91] |
| O-Coumaric acid | 614-60-8 | Phenols | Antiadipogenesis, antioxidant, and antiviral (as a component of a plant, indirectly) | [92,93,94,95] |
| Vanillin | 121-33-5 | Phenols | Antiviral, antimicrobial, anti-inflammatory, antiapoptotic, neuroprotective, and antioxidant | [96,97,98,99,100] |
| Zanamivir | 139110-80-8 | NAIs | Anti-influenza | [101] |
| Compound | Virus | CC50 (mg/mL) | IC50 (mg/mL) | SI |
|---|---|---|---|---|
| Silybin | H1N1 | 9.48 | N/A | ND |
| H5N1 | N/A | ND | ||
| 7-Hydroxy flavone | H1N1 | 5.83 | 0.360 | 16.194 |
| H5N1 | N/A | ND | ||
| Naringin | H1N1 | >10 | 0.0206 | >485.43 |
| H5N1 | N/A | ND | ||
| Pinocembrin | H1N1 | >10 | >10 | >1 |
| H5N1 | N/A | ND | ||
| Kaempferitrin | H1N1 | >10 | 0.0478 | >209.20 |
| H5N1 | N/A | ND | ||
| Flavanone | H1N1 | 0.45 | N/A | ND |
| H5N1 | N/A | ND | ||
| Isoquercitrin | H1N1 | 0.71 | 0.167 | 4.25 |
| H5N1 | N/A | ND | ||
| Saponin | H1N1 | >10 | 0.326 | >30.674 |
| H5N1 | N/A | ND | ||
| Lupeol | H1N1 | 0.56 | 0.0936 | 5.98 |
| H5N1 | N/A | ND | ||
| D-Glucuronic acid | H1N1 | 10.26 | N/A | ND |
| H5N1 | N/A | ND | ||
| D-Galacturonic acid | H1N1 | 9.61 | N/A | ND |
| H5N1 | N/A | ND | ||
| β-sitosterol | H1N1 | 9.9 | 0.000975 | 10,154 |
| H5N1 | 0.000295 | 33,559 | ||
| β-sitosterol-O-glucoside | H1N1 | 1.75 | 0.000719 | 2434 |
| H5N1 | 0.000613 | 2855 | ||
| Ouabain | H1N1 | 0.176 | N/A | ND |
| H5N1 | N/A | ND | ||
| Digitonin | H1N1 | 0.442 | 0.0215 | 20.56 |
| H5N1 | N/A | ND | ||
| Chrysophanic acid | H1N1 | 0.0461 | N/A | ND |
| H5N1 | N/A | ND | ||
| Aloe emodin | H1N1 | 2.28 | 0.729 | 3.127 |
| H5N1 | N/A | ND | ||
| Arbutin | H1N1 | >10 | 0.764 | >13.09 |
| H5N1 | N/A | ND | ||
| O-Coumaric acid | H1N1 | 9.172 | NA | ND |
| H5N1 | N/A | ND | ||
| Vanillin | H1N1 | 0.242 | N/A | ND |
| H5N1 | N/A | ND | ||
| D-sorbitol | H1N1 | >10 | 3.11 | >3.215 |
| H5N1 | N/A | ND | ||
| D-(-) salicin | H1N1 | >10 | N/A | ND |
| H5N1 | N/A | ND | ||
| Zanamivir | H1N1 | >10 | 0.000792 | >12,626 |
| H5N1 | 0.000265 | >37,736 |
| Steroid | Concentration (µg/mL) | Stage of Antiviral Action | ||
|---|---|---|---|---|
| Viral Replication | Viricidal | Viral Adsorption | ||
| β-sitosterol | 1 | 31.8% | 84% | 28% |
| 10 | 41.2% | 98.5% | 37.5% | |
| 100 | 52.9% | 99% | 52.5% | |
| β-sitosterol-O-glucoside | 1 | 28.7% | 83.6% | 35.8% |
| 10 | 37.7% | 93.3% | 48.3% | |
| 100 | 52.8% | 97.1% | 56.9% | |
| Compound | Hemagglutinin | Neuraminidase | M2 | Hemagglutinin Head Epitope |
|---|---|---|---|---|
| β-sitosterol | −6.40 | −29.40 | −10.97 | −10.90 |
| β-sitosterol-O-glucoside | −6.78 | −30.13 | −10.75 | −8.97 |
| 6′-Sialyl-N-acetyllactosamine | −5.66 | - | - | - |
| Zanamivir | - | −19.28 | - | - |
| Rimantadine | - | - | −9.968 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shokry, S.; Hegazy, A.; Abbas, A.M.; Mostafa, I.; Eissa, I.H.; Metwaly, A.M.; Yahya, G.; El-Shazly, A.M.; Aboshanab, K.M.; Mostafa, A. Phytoestrogen β-Sitosterol Exhibits Potent In Vitro Antiviral Activity against Influenza A Viruses. Vaccines 2023, 11, 228. https://doi.org/10.3390/vaccines11020228
Shokry S, Hegazy A, Abbas AM, Mostafa I, Eissa IH, Metwaly AM, Yahya G, El-Shazly AM, Aboshanab KM, Mostafa A. Phytoestrogen β-Sitosterol Exhibits Potent In Vitro Antiviral Activity against Influenza A Viruses. Vaccines. 2023; 11(2):228. https://doi.org/10.3390/vaccines11020228
Chicago/Turabian StyleShokry, Sara, Akram Hegazy, Ahmad M. Abbas, Islam Mostafa, Ibrahim H. Eissa, Ahmed M. Metwaly, Galal Yahya, Assem M. El-Shazly, Khaled M. Aboshanab, and Ahmed Mostafa. 2023. "Phytoestrogen β-Sitosterol Exhibits Potent In Vitro Antiviral Activity against Influenza A Viruses" Vaccines 11, no. 2: 228. https://doi.org/10.3390/vaccines11020228
APA StyleShokry, S., Hegazy, A., Abbas, A. M., Mostafa, I., Eissa, I. H., Metwaly, A. M., Yahya, G., El-Shazly, A. M., Aboshanab, K. M., & Mostafa, A. (2023). Phytoestrogen β-Sitosterol Exhibits Potent In Vitro Antiviral Activity against Influenza A Viruses. Vaccines, 11(2), 228. https://doi.org/10.3390/vaccines11020228