
COVID-19: The Ethno-Geographic Perspective of Differential Immunity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
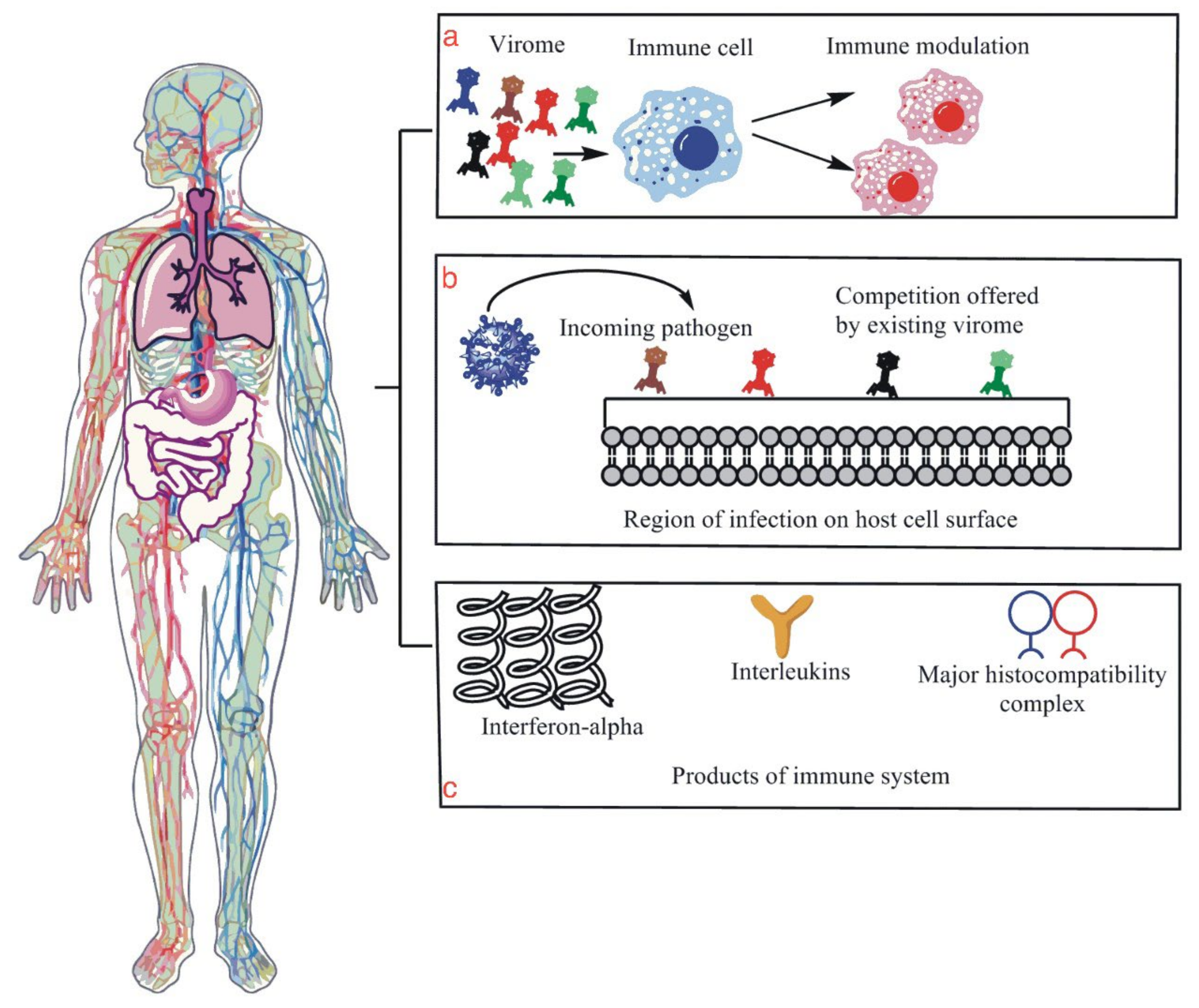
2. Human Microbiota
- Digestion and the ability to harvest nutrients [11].
- Modulation of the appetite signal of the body [11].
- The production of important vitamins for the body (including vitamin K) [12].
- The regulation of epithelial growth and development [13].
- The metabolization of various xenobiotics in the body [13].
- Modulation of the host immune system [14].
- Modulating the human endogenous retrovirus (HERV) expression plays a role in modulating future antiviral immunity [15].
- The bacteriophages in our body help us fight against bacterial infections by directly targeting the virus particle [17].
- The most notable is how it fights off other incoming viruses for competition in the region while also immune priming the body in preparation for infection by external pathogenic viruses [16].
- The microbiota heavily influence innate lymphoid cells’ (ILCs) development, maintenance, and function [18]. The ILCs, as well as possessing all the characteristics of an innate immune cell, also have the characteristics of adaptive T cells to release inflammatory agents, but they lack antigen-specific receptors [19]. Furthermore, ILCs are diverse as they have five subsets: type 1, 2, and 3 ILCs, lymphoid tissue inducer cells, and natural killer cells [20]. Several studies have shown that ILC subsets are interconvertible if they receive the appropriate cytokine stimuli, demonstrating these cells’ high degree of plasticity [21]. They also have particularly unique roles in immunity that normal adaptive immune cells or innate immune cells do not possess on their own [22].
2.1. Human Microbiota and COVID-19
2.2. Hygiene Hypothesis of Human Microbiota and COVID-19
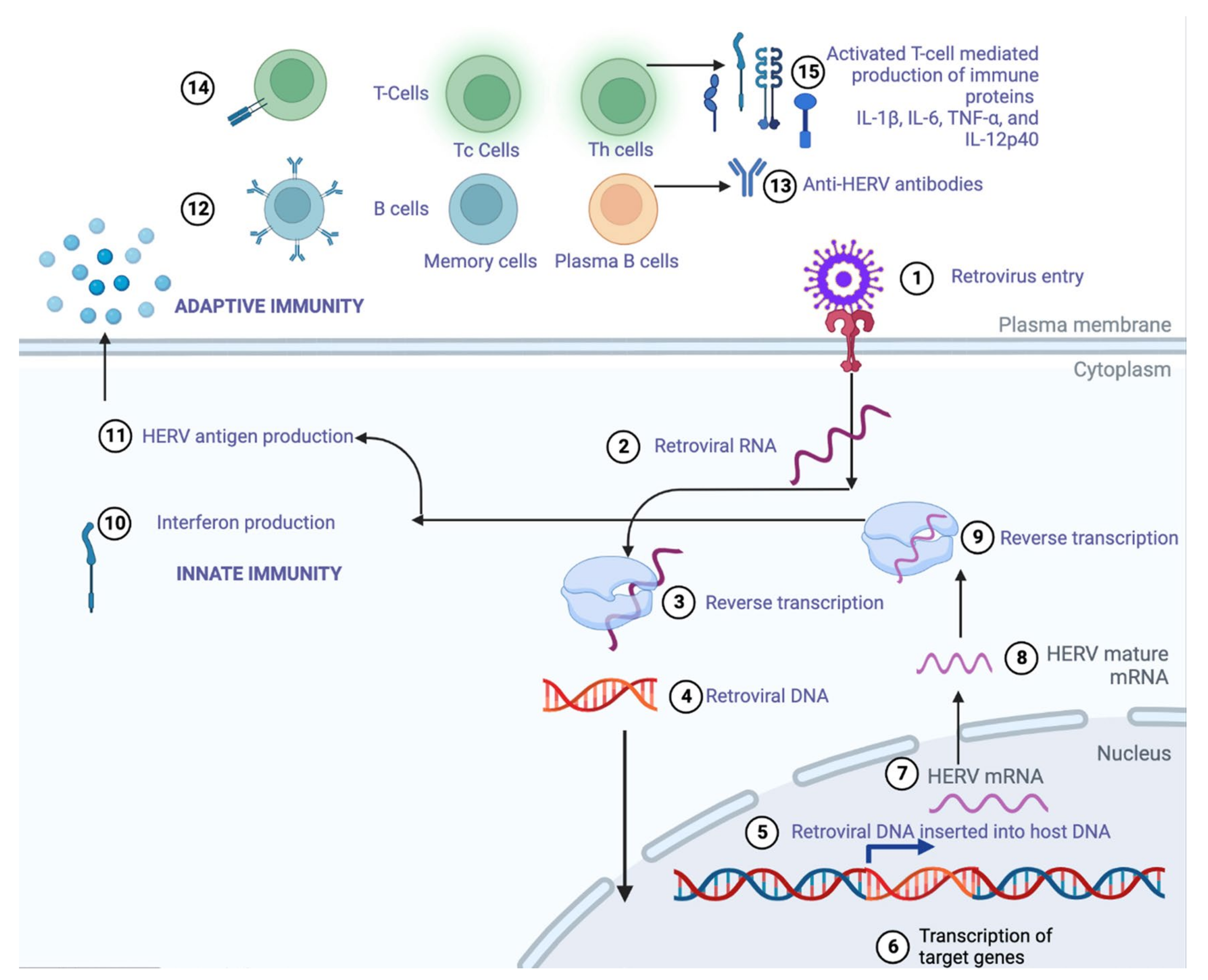
3. Human Endogenous Retroviruses (HERVs)
3.1. Activation of Innate Immune System
3.2. Production of Long Non-Coding RNAs (lncRNAs)
3.3. Modulation of the Immune System by ERV-Based Proteins
3.4. Receptor Interference
3.5. Complementing in a Negative Manner
3.6. Interference in the Replicative Cycle
3.7. Increase in the Diversity and Plasticity of Immune Genes
3.8. LTRs-Based Enhancement of Antiviral Genes
3.9. Genetic Memory
3.10. Innate Immune System Triggering
4. Previous Unrecorded Local Epidemics
5. Copy Number Variation
6. Potential Cell–Gene-Based Immunotherapy Human-Gene-Transfer-Based Genetic Engineering, Gene Drive Technologies
7. Conclusions
8. Future Work Directions
- The collection of strategically randomized samples of DNAs, i.e., setting up a broad spectrum of subjects from major demographic regions of the world. Important things to keep in mind while choosing subjects would be that they should be genetically diverse from each other and not just from different countries (for example, the Eastern Punjab region of Pakistan and the Western Punjab region of India have people with similar gene pools. Therefore, the results of this sample might not be so prominent). It is important to at least have subjects from major parts of Europe, North and South America, and Asia (Southeast Asian Island nations and South Asian countries included).
- For the virome and microbiome hypothesis, metagenomic analysis and classification and then a comparison of subjects’ microbiomes with each other’s, and finding the potentially helpful groups of bacteria, viruses and bacteriophages, may allow us to move towards unleashing the therapeutic arenas of the microbiome and virome, which include the modulation of a person’s microbiome, phage therapy, pre-, and pro-biotic therapies, microbiome transplantation, etc.
- HERVs will require genomic analysis and genome sequences. If we can figure out the region of HERV that is potentially boosting the immune system of a subject, we may be able to replicate it and find its molecular basis and biological causative agents involved, thus allowing us to obtain a better understanding of the disease host interaction role of ERV in our genome, all of which, in turn, may allow us to utilize the possible therapeutic benefits.
- To test the ENV hypothesis, an extensive genomic analysis (NextGen) would be required to identify potential genes and find their respective roles in epigenetics and immune modulation, etc., which would allow us to understand the functions of the mechanisms better.
- A study on the population of Pakistan and Bangladesh for hypothesis 3 can also be performed to investigate whether such a case exists or not. This will require a lot of data analysis, bioinformatics, and big data comparative analysis, but if such a case is indeed found, then we may be able to exploit the same phenomenon to potentiate relative immunity against a future pathogen with the help of the region’s previously encountered and less dangerous pathogens.
- Lastly, having all of this analytical big data will allow us to use AI and various algorithms to predict the effect of future SARS-CoV* strains in a certain region or a geographical location based on their biological capabilities and not just the genetic backbone, which will give the scientific and medical communities a huge edge in refocusing their limited resources.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coronavirus Disease (COVID-19)—World Health Organization. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 19 March 2022).
- Kim, D.; Lee, J.-Y.; Yang, J.-S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef] [PubMed]
- Yuki, K.; Fujiogi, M.; Koutsogiannaki, S. COVID-19 Pathophysiology: A Review. Clin. Immunol. 2020, 215, 108427. [Google Scholar] [CrossRef]
- Shoaib, N.; Noureen, N.; Munir, R.; Shah, F.A.; Ishtiaq, N.; Jamil, N.; Batool, R.; Khalid, M.; Khan, I.; Iqbal, N.; et al. COVID-19 Severity: Studying the Clinical and Demographic Risk Factors for Adverse Outcomes. PLoS ONE 2021, 16, e0255999. [Google Scholar] [CrossRef] [PubMed]
- Is the Severe COVID-19 Over in Europe? Available online: https://healthmanagement.org/c/icu/issuearticle/is-the-severe-covid-19-over-in-europe (accessed on 20 March 2022).
- Statement—Risk Remains High in Eastern Europe and Central Asia with Arrival of COVID-19 Omicron Variant. Available online: https://www.euro.who.int/en/media-centre/sections/statements/2022/statement-risk-remains-high-in-eastern-europe-and-central-asia-with-arrival-of-covid-19-omicron-variant (accessed on 20 March 2022).
- Konstantinoudis, G.; Cameletti, M.; Gómez-Rubio, V.; Gómez, I.L.; Pirani, M.; Baio, G.; Larrauri, A.; Riou, J.; Egger, M.; Vineis, P.; et al. Regional Excess Mortality during the 2020 COVID-19 Pandemic in Five European Countries. Nat. Commun. 2022, 13, 482. [Google Scholar] [CrossRef] [PubMed]
- Population of Europe (2022)—Worldometer. Available online: https://www.worldometers.info/world-population/europe-population/ (accessed on 20 March 2022).
- Unleashing the Potential of a Young Pakistan | Human Development Reports. Available online: https://hdr.undp.org/en/content/unleashing-potential-young-pakistan (accessed on 28 April 2022).
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raoult, D. Obesity Pandemics and the Modification of Digestive Bacterial Flora. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.G.; Laiño, J.E.; del Valle, M.J.; Vannini, V.; van Sinderen, D.; Taranto, M.P.; de Valdez, G.F.; de Giori, G.S.; Sesma, F. B-Group Vitamin Production by Lactic Acid Bacteria—Current Knowledge and Potential Applications. J. Appl. Microbiol. 2011, 111, 1297–1309. [Google Scholar] [CrossRef]
- Clarke, G.; Sandhu, K.V.; Griffin, B.T.; Dinan, T.G.; Cryan, J.F.; Hyland, N.P. Gut Reactions: Breaking Down Xenobiotic–Microbiome Interactions. Pharmacol. Rev. 2019, 71, 198–224. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.; Lübben, W.; Slomka, H.; Gebler, J.; Konert, M.; Cai, C.; Neubrandt, L.; Prazeres da Costa, O.; Paul, S.; Dehnert, S.; et al. Nucleic Acid-Sensing Toll-like Receptors Are Essential for the Control of Endogenous Retrovirus Viremia and ERV-Induced Tumors. Immunity 2012, 37, 867–879. [Google Scholar] [CrossRef]
- Breitbart, M.; Hewson, I.; Felts, B.; Mahaffy, J.M.; Nulton, J.; Salamon, P.; Rohwer, F. Metagenomic Analyses of an Uncultured Viral Community from Human Feces. J. Bacteriol. 2003, 185, 6220–6223. [Google Scholar] [CrossRef] [Green Version]
- Young, J.C.; Chehoud, C.; Bittinger, K.; Bailey, A.; Diamond, J.M.; Cantu, E.; Haas, A.R.; Abbas, A.; Frye, L.; Christie, J.D.; et al. Viral Metagenomics Reveal Blooms of Anelloviruses in the Respiratory Tract of Lung Transplant Recipients. Am. J. Transplant. 2015, 15, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Ganal-Vonarburg, S.C.; Duerr, C.U. The Interaction of Intestinal Microbiota and Innate Lymphoid Cells in Health and Disease throughout Life. Immunology 2020, 159, 39–51. [Google Scholar] [CrossRef]
- Innate Lymphoid Cells: A New Paradigm in Immunology Science. Available online: https://www.science.org/doi/10.1126/science.aaa6566 (accessed on 20 January 2023).
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- DuPage, M.; Bluestone, J.A. Harnessing the Plasticity of CD4(+) T Cells to Treat Immune-Mediated Disease. Nat. Rev. Immunol. 2016, 16, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Sawa, S.; Cherrier, M.; Lochner, M.; Satoh-Takayama, N.; Fehling, H.J.; Langa, F.; Di Santo, J.P.; Eberl, G. Lineage Relationship Analysis of RORgammat+ Innate Lymphoid Cells. Science 2010, 330, 665–669. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Weber-Dąbrowska, B.; Żaczek, M.; Borysowski, J.; Górski, A. The Presence of Bacteriophages in the Human Body: Good, Bad or Neutral? Microorganisms 2020, 8, 2012. [Google Scholar] [CrossRef]
- Merad, M.; Subramanian, A.; Wang, T.T. An Aberrant Inflammatory Response in Severe COVID-19. Cell Host Microbe 2021, 29, 1043–1047. [Google Scholar] [CrossRef]
- Combes, A.J.; Courau, T.; Kuhn, N.F.; Hu, K.H.; Ray, A.; Chen, W.S.; Chew, N.W.; Cleary, S.J.; Kushnoor, D.; Reeder, G.C.; et al. Global Absence and Targeting of Protective Immune States in Severe COVID-19. Nature 2021, 591, 124–130. [Google Scholar] [CrossRef]
- Pairo-Castineira, E.; Clohisey, S.; Klaric, L.; Bretherick, A.D.; Rawlik, K.; Pasko, D.; Walker, S.; Parkinson, N.; Fourman, M.H.; Russell, C.D.; et al. Genetic Mechanisms of Critical Illness in COVID-19. Nature 2021, 591, 92–98. [Google Scholar] [CrossRef]
- Cash, H.L.; Whitham, C.V.; Behrendt, C.L.; Hooper, L.V. Symbiotic Bacteria Direct Expression of an Intestinal Bactericidal Lectin. Science 2006, 313, 1126–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surana, N.K.; Kasper, D.L. Moving beyond microbiome-wide associations to causal microbe identification. Nature 2018, 554, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, S.; Weiss, S.T.; Liu, Y.-Y. Dissecting the Role of the Human Microbiome in COVID-19 via Metagenome-Assembled Genomes. Nat. Commun. 2022, 13, 5235. [Google Scholar] [CrossRef] [PubMed]
- Mining the Human Gut Microbiota for Immunomodulatory Organisms—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/28215708/ (accessed on 20 January 2023).
- Zuo, T.; Liu, Q.; Zhang, F.; Lui, G.C.-Y.; Tso, E.Y.; Yeoh, Y.K.; Chen, Z.; Boon, S.S.; Chan, F.K.; Chan, P.K.; et al. Depicting SARS-CoV-2 Faecal Viral Activity in Association with Gut Microbiota Composition in Patients with COVID-19. Gut 2021, 70, 276–284. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [Green Version]
- Vanaja, S.K.; Rathinam, V.A.K.; Fitzgerald, K.A. Mechanisms of Inflammasome Activation: Recent Advances and Novel Insights. Trends Cell Biol. 2015, 25, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, D.; Guo, Y.; Kamada, N. Interaction between the Inflammasome and Commensal Microorganisms in Gastrointestinal Health and Disease. EMBO Mol. Med. 2021, 13, e13452. [Google Scholar] [CrossRef]
- Aymonnier, K.; Ng, J.; Fredenburgh, L.E.; Zambrano-Vera, K.; Münzer, P.; Gutch, S.; Fukui, S.; Desjardins, M.; Subramaniam, M.; Baron, R.M.; et al. Inflammasome Activation in Neutrophils of Patients with Severe COVID-19. Blood Adv. 2022, 6, 2001–2013. [Google Scholar] [CrossRef]
- An Enteric Virus Can Replace the Beneficial Function of Commensal Bacteria—PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4257755/ (accessed on 20 April 2022).
- Rampelli, S.; Turroni, S.; Schnorr, S.L.; Soverini, M.; Quercia, S.; Barone, M.; Castagnetti, A.; Biagi, E.; Gallinella, G.; Brigidi, P.; et al. Characterization of the Human DNA Gut Virome across Populations with Different Subsistence Strategies and Geographical Origin. Environ. Microbiol. 2017, 19, 4728–4735. [Google Scholar] [CrossRef]
- Zuo, T.; Sun, Y.; Wan, Y.; Yeoh, Y.K.; Zhang, F.; Cheung, C.P.; Chen, N.; Luo, J.; Wang, W.; Sung, J.J.Y.; et al. Human-Gut-DNA Virome Variations across Geography, Ethnicity, and Urbanization. Cell Host Microbe 2020, 28, 741–751.e4. [Google Scholar] [CrossRef]
- Finlay, B.B.; Amato, K.R.; Azad, M.; Blaser, M.J.; Bosch, T.C.G.; Chu, H.; Dominguez-Bello, M.G.; Ehrlich, S.D.; Elinav, E.; Geva-Zatorsky, N.; et al. The hygiene hypothesis, the COVID pandemic, and consequences for the human microbiome. Proc. Natl. Acad. Sci. USA 2021, 118, e2010217118. [Google Scholar] [CrossRef]
- McPherson, J.D.; Marra, M.; Hillier, L.; Waterston, R.H.; Chinwalla, A.; Wallis, J.; Sekhon, M.; Wylie, K.; Mardis, E.R.; Wilson, R.K.; et al. A Physical Map of the Human Genome. Nature 2001, 409, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Qiu, Y.; Liu, H.; Liang, B.; Fan, B.; Zhou, X.; Liu, D. Transcription Profile of Human Endogenous Retroviruses in Response to Dengue Virus Serotype 2 Infection. Virology 2020, 544, 21–30. [Google Scholar] [CrossRef]
- Vargiu, L.; Rodriguez-Tomé, P.; Sperber, G.O.; Cadeddu, M.; Grandi, N.; Blikstad, V.; Tramontano, E.; Blomberg, J. Classification and Characterization of Human Endogenous Retroviruses; Mosaic Forms Are Common. Retrovirology 2016, 13, 7. [Google Scholar] [CrossRef] [Green Version]
- Esposito, F.; Tramontano, E. Past and Future. Current Drugs Targeting HIV-1 Integrase and Reverse Transcriptase-Associated Ribonuclease H Activity: Single and Dual Active Site Inhibitors. Antivir. Chem. Chemother. 2014, 23, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Bannert, N.; Kurth, R. The Evolutionary Dynamics of Human Endogenous Retroviral Families. Annu. Rev. Genom. Hum. Genet. 2006, 7, 149–173. [Google Scholar] [CrossRef]
- Zhou, Z.; Ren, L.; Zhang, L.; Zhong, J.; Xiao, Y.; Jia, Z.; Guo, L.; Yang, J.; Wang, C.; Jiang, S.; et al. Heightened Innate Immune Responses in the Respiratory Tract of COVID-19 Patients. Cell Host Microbe 2020, 27, 883–890.e2. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, Y.; Cao, L.; Wang, D.; Guo, M.; Jiang, A.; Guo, D.; Hu, W.; Yang, J.; Tang, Z.; et al. Transcriptomic Characteristics of Bronchoalveolar Lavage Fluid and Peripheral Blood Mononuclear Cells in COVID-19 Patients. Emerg. Microbes Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef]
- Kitsou, K.; Kotanidou, A.; Paraskevis, D.; Karamitros, T.; Katzourakis, A.; Tedder, R.; Hurst, T.; Sapounas, S.; Kotsinas, A.; Gorgoulis, V.; et al. Upregulation of Human Endogenous Retroviruses in Bronchoalveolar Lavage Fluid of COVID-19 Patients. Microbiol. Spectr. 2021, 9, e01260-21. [Google Scholar] [CrossRef]
- Srinivasachar Badarinarayan, S.; Sauter, D. Switching Sides: How Endogenous Retroviruses Protect Us from Viral Infections. J. Virol. 2021, 95, e02299-20. [Google Scholar] [CrossRef]
- Dopkins, N.; O’Mara, M.M.; Singh, B.; Marston, J.L.; Bendall, M.L.; Nixon, D.F. How human endogenous retroviruses interact with the microbiota in health and disease. Trends Microbiol. 2022, 30, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA Helicase RIG-I Has an Essential Function in Double-Stranded RNA-Induced Innate Antiviral Responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Qi, F.; Wu, F.; Nie, H.; Song, Y.; Shao, L.; Han, J.; Wu, Z.; Saiyin, H.; Wei, G.; et al. Endogenous Retrovirus-Derived Long Noncoding RNA Enhances Innate Immune Responses via Derepressing RELA Expression. mBio 2019, 10, e00937-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NeST, a Long Noncoding RNA, Controls Microbial Susceptibility and Epigenetic Activation of the Ifng Locus—PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3577098/ (accessed on 20 April 2022).
- Human Endogenous Retrovirus Type W Envelope Protein Inhibits Oligodendroglial Precursor Cell Differentiation—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/23836485/ (accessed on 20 March 2022).
- Vogt, P.K.; Ishizaki, R. Patterns of Viral Interference in the Avian Leukosis and Sarcoma Complex. Virology 1966, 30, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Role of Angiotensin-Converting Enzyme 2 (ACE2) in COVID-19. Critical Care. Available online: https://ccforum.biomedcentral.com/articles/10.1186/s13054-020-03120-0 (accessed on 28 May 2022).
- A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/32015507/ (accessed on 28 May 2022).
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-Converting Enzyme 2 Is a Functional Receptor for the SARS Coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Abbasi, A.Z.; Kiyani, D.A.; Hamid, S.M.; Saalim, M.; Fahim, A.; Jalal, N. Spiking Dependence of SARS-CoV-2 Pathogenicity on TMPRSS2. J. Med. Virol. 2021, 93, 4205–4218. [Google Scholar] [CrossRef]
- Steck, F.T.; Rubin, H. The Mechanism of Interference between an Avian Leukosis Virus and Rous Sarcoma Virus. II. Early Steps of Infection by RSV of Cells under Conditions of Interference. Virology 1966, 29, 642–653. [Google Scholar] [CrossRef]
- Taylor, G.M.; Gao, Y.; Sanders, D.A. Fv-4: Identification of the Defect in Env and the Mechanism of Resistance to Ecotropic Murine Leukemia Virus. J. Virol. 2001, 75, 11244–11248. [Google Scholar] [CrossRef] [Green Version]
- Brinzevich, D.; Young, G.R.; Sebra, R.; Ayllon, J.; Maio, S.M.; Deikus, G.; Chen, B.K.; Fernandez-Sesma, A.; Simon, V.; Mulder, L.C.F. HIV-1 Interacts with Human Endogenous Retrovirus K (HML-2) Envelopes Derived from Human Primary Lymphocytes. J. Virol. 2014, 88, 6213–6223. [Google Scholar] [CrossRef] [Green Version]
- Boso, G.; Buckler-White, A.; Kozak, C.A. Ancient Evolutionary Origin and Positive Selection of the Retroviral Restriction Factor Fv1 in Muroid Rodents. J. Virol. 2018, 92, e00850-18. [Google Scholar] [CrossRef]
- Yan, Y.; Buckler-White, A.; Wollenberg, K.; Kozak, C.A. Origin, Antiviral Function and Evidence for Positive Selection of the Gammaretrovirus Restriction Gene Fv1 in the Genus Mus. Proc. Natl. Acad. Sci. USA 2009, 106, 3259–3263. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Skaletsky, H.; Rozen, S.; Gromoll, J.; Nieschlag, E.; Oates, R.; Page, D.C. Deletion of Azoospermia Factor a (AZFa) Region of Human Y Chromosome Caused by Recombination between HERV15 Proviruses. Hum. Mol. Genet. 2000, 9, 2291–2296. [Google Scholar] [CrossRef] [Green Version]
- Campbell, I.M.; Gambin, T.; Dittwald, P.; Beck, C.R.; Shuvarikov, A.; Hixson, P.; Patel, A.; Gambin, A.; Shaw, C.A.; Rosenfeld, J.A.; et al. Human Endogenous Retroviral Elements Promote Genome Instability via Non-Allelic Homologous Recombination. BMC Biol. 2014, 12, 74. [Google Scholar] [CrossRef] [Green Version]
- Feschotte, C.; Gilbert, C. Endogenous Viruses: Insights into Viral Evolution and Impact on Host Biology. Nat. Rev. Genet. 2012, 13, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.F.; Coffin, J.M. Evidence for Genomic Rearrangements Mediated by Human Endogenous Retroviruses during Primate Evolution. Nat. Genet. 2001, 29, 487–489. [Google Scholar] [CrossRef]
- Doxiadis, G.G.M.; de Groot, N.; Bontrop, R.E. Impact of Endogenous Intronic Retroviruses on Major Histocompatibility Complex Class II Diversity and Stability. J. Virol. 2008, 82, 6667–6677. [Google Scholar] [CrossRef] [Green Version]
- Kulski, J.K.; Gaudieri, S.; Martin, A.; Dawkins, R.L. Coevolution of PERB11 (MIC) and HLA Class I Genes with HERV-16 and Retroelements by Extended Genomic Duplication. J. Mol. Evol. 1999, 49, 84–97. [Google Scholar] [CrossRef]
- Ito, J.; Sugimoto, R.; Nakaoka, H.; Yamada, S.; Kimura, T.; Hayano, T.; Inoue, I. Systematic Identification and Characterization of Regulatory Elements Derived from Human Endogenous Retroviruses. PLoS Genet. 2017, 13, e1006883. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, V.; Cheng, Y.; Ma, Z.; Li, D.; Xing, X.; Edge, P.; Snyder, M.P.; Wang, T. Widespread Contribution of Transposable Elements to the Innovation of Gene Regulatory Networks. Genome Res. 2014, 24, 1963–1976. [Google Scholar] [CrossRef] [Green Version]
- Wildschutte, J.H.; Williams, Z.H.; Montesion, M.; Subramanian, R.P.; Kidd, J.M.; Coffin, J.M. Discovery of Unfixed Endogenous Retrovirus Insertions in Diverse Human Populations. Proc. Natl. Acad. Sci. USA 2016, 113, E2326–E2334. [Google Scholar] [CrossRef]
- Hurwitz, J.L.; Jones, B.G.; Charpentier, E.; Woodland, D.L. Hypothesis: RNA and DNA Viral Sequence Integration into the Mammalian Host Genome Supports Long-Term B Cell and T Cell Adaptive Immunity. Viral Immunol. 2017, 30, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Bourque, G.; Leong, B.; Vega, V.B.; Chen, X.; Lee, Y.L.; Srinivasan, K.G.; Chew, J.-L.; Ruan, Y.; Wei, C.-L.; Ng, H.H.; et al. Evolution of the Mammalian Transcription Factor Binding Repertoire via Transposable Elements. Genome Res. 2008, 18, 1752–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Lagemaat, L.N.; Landry, J.-R.; Mager, D.L.; Medstrand, P. Transposable Elements in Mammals Promote Regulatory Variation and Diversification of Genes with Specialized Functions. Trends Genet. 2003, 19, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C. Transposable Elements and the Evolution of Regulatory Networks. Nat. Rev. Genet. 2008, 9, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbalat, R.; Ewald, S.E.; Mouchess, M.L.; Barton, G.M. Nucleic Acid Recognition by the Innate Immune System. Annu. Rev. Immunol. 2011, 29, 185–214. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Control of Adaptive Immunity by the Innate Immune System. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-Specific Recognition of Single-Stranded RNA via Toll-like Receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- McCarroll, S.A.; Altshuler, D.M. Copy-Number Variation and Association Studies of Human Disease. Nat. Genet. 2007, 39, S37–S42. [Google Scholar] [CrossRef]
- Machado, L.R.; Ottolini, B. An Evolutionary History of Defensins: A Role for Copy Number Variation in Maximizing Host Innate and Adaptive Immune Responses. Front. Immunol. 2015, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- Perry, G.H.; Dominy, N.J.; Claw, K.G.; Lee, A.S.; Fiegler, H.; Redon, R.; Werner, J.; Villanea, F.A.; Mountain, J.L.; Misra, R.; et al. Diet and the Evolution of Human Amylase Gene Copy Number Variation. Nat. Genet. 2007, 39, 1256–1260. [Google Scholar] [CrossRef]
- Macdonald, M. A Novel Gene Containing a Trinucleotide Repeat That Is Expanded and Unstable on Huntington’s Disease Chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Myers, R.H. Huntington’s Disease Genetics. NeuroRx 2004, 1, 255–262. [Google Scholar] [CrossRef]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef]
- Genetic Engineering of T Cells for Immunotherapy|Nature Reviews Genetics. Available online: https://www.nature.com/articles/s41576-021-00329-9 (accessed on 12 December 2022).
- mdabrow1 Gene Transfer Research. Available online: https://www.hopkinsmedicine.org/institutional_review_board/guidelines_policies/guidelines/gene_transfer.html (accessed on 12 December 2022).
- Bier, E. Gene Drives Gaining Speed. Nat. Rev. Genet. 2022, 23, 5–22. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullah, U.; Saleh, N.; Shaw, P.; Jalal, N. COVID-19: The Ethno-Geographic Perspective of Differential Immunity. Vaccines 2023, 11, 319. https://doi.org/10.3390/vaccines11020319
Abdullah U, Saleh N, Shaw P, Jalal N. COVID-19: The Ethno-Geographic Perspective of Differential Immunity. Vaccines. 2023; 11(2):319. https://doi.org/10.3390/vaccines11020319
Chicago/Turabian StyleAbdullah, Usman, Ned Saleh, Peter Shaw, and Nasir Jalal. 2023. "COVID-19: The Ethno-Geographic Perspective of Differential Immunity" Vaccines 11, no. 2: 319. https://doi.org/10.3390/vaccines11020319
APA StyleAbdullah, U., Saleh, N., Shaw, P., & Jalal, N. (2023). COVID-19: The Ethno-Geographic Perspective of Differential Immunity. Vaccines, 11(2), 319. https://doi.org/10.3390/vaccines11020319