
Harnessing Pentameric Scaffold of Cholera Toxin B (CTB) for Design of Subvirion Recombinant Dengue Virus Vaccine


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression Vector and Mutagenesis
2.2. Protein Expression and Purification
2.3. GM1 Binding Assay
2.4. Immunization and Sera Collection
2.5. Virus and Serum Antibody Analysis
2.6. Neutralization Assay
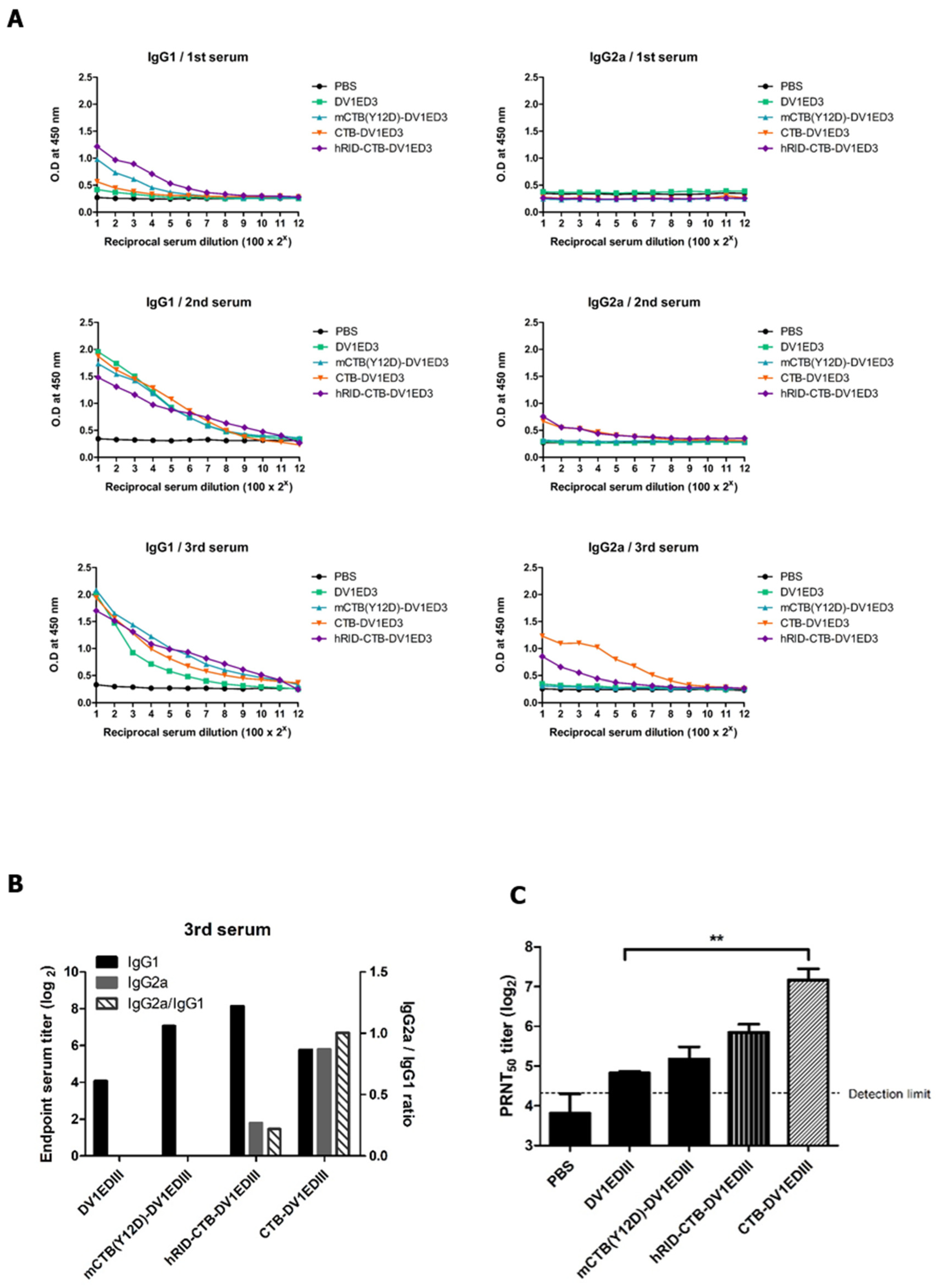
3. Results
3.1. Development of Recombinant Proteins
3.2. Protein Purification and Pentamer Analysis
3.3. Immunogenicity Study of the CTB Fusion Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; WHO: Geneva, Switzerland, 2009.
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.; Hay, S.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.; et al. The global burden of dengue: An analysis from the Global Burden of Disease Study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef]
- Murray, N.E.A.; Quam, M.B.; Wilder-Smith, A. Epidemiology of dengue: Past, present and future prospects. Clin. Epidemiol. 2013, 5, 299–309. [Google Scholar] [PubMed]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef]
- Martina, B.E.E.; Koraka, P.; Osterhaus, A.D.M.E. Dengue Virus Pathogenesis: An Integrated View. Clin. Microbiol. Rev. 2009, 22, 564–581. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The Impact of the Newly Licensed Dengue Vaccine in Endemic Countries. PLoS Neglected Trop. Dis. 2016, 10, e5179. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B.; Russell, P.K. Protective and immunological behavior of chimeric yellow fever dengue vaccine. Vaccine 2016, 34, 1643–1647. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Dengvaxia sensitizes seronegatives to vaccine enhanced disease regardless of age. Vaccine 2017, 35, 6355–6358. [Google Scholar] [CrossRef]
- Schmitz, J.; Roehrig, J.; Barrett, A.; Hombach, J. Next generation dengue vaccines: A review of candidates in preclinical development. Vaccine 2011, 29, 7276–7284. [Google Scholar] [CrossRef]
- Prompetchara, E.; Ketloy, C.; Thomas, S.J.; Ruxrungtham, K. Dengue vaccine: Global development update. Asian Pac. J. Allergy Immunol. 2019, 38, 178–185. [Google Scholar]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A continuing global threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef]
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of Dengue Virus: Implications for Flavivirus Organization, Maturation, and Fusion. Cell 2002, 108, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Beltramello, M.; Williams, K.L.; Simmons, C.P.; Macagno, A.; Simonelli, L.; Quyen, N.T.H.; Sukupolvi-Petty, S.; Navarro-Sanchez, E.; Young, P.R.; de Silva, A.M.; et al. The Human Immune Response to Dengue Virus Is Dominated by Highly Cross-Reactive Antibodies Endowed with Neutralizing and Enhancing Activity. Cell Host Microbe 2010, 8, 271–283. [Google Scholar] [CrossRef]
- Flipse, J.; Wilschut, J.; Smit, J.M. Molecular Mechanisms Involved in Antibody-Dependent Enhancement of Dengue Virus Infection in Humans. Traffic 2012, 14, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kostyuchenko, V.A.; Zhang, Q.; Tan, J.L.; Ng, T.-S.; Lok, S.-M. Immature and Mature Dengue Serotype 1 Virus Structures Provide Insight into the Maturation Process. J. Virol. 2013, 87, 7700–7707. [Google Scholar] [CrossRef]
- Zhang, X.; Ge, P.; Yu, X.; Brannan, J.M.; Bi, G.; Zhang, Q.; Schein, S.; Zhou, Z.H. Cryo-EM structure of the mature dengue virus at 3.5-Å resolution. Nat. Struct. Mol. Biol. 2013, 20, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.K.; Shrivastava, A. Recent Developments in Recombinant Protein–Based Dengue Vaccines. Front. Immunol. 2018, 9, 1919. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Seong, B.L. Exploiting virus-like particles as innovative vaccines against emerging viral infections. J. Microbiol. 2017, 55, 220–230. [Google Scholar] [CrossRef]
- Jennings, G.T.; Bachmann, M.F. The coming of age of virus-like particle vaccines. Biol. Chem. 2008, 389, 521–536. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Zabel, F.; Kündig, T.M.; Bachmann, M.F. Virus-induced humoral immunity: On how B cell responses are initiated. Curr. Opin. Virol. 2013, 3, 357–362. [Google Scholar] [CrossRef]
- Baldauf, K.J.; Royal, J.M.; Hamorsky, K.T.; Matoba, N. Cholera Toxin B: One Subunit with Many Pharmaceutical Applications. Toxins 2015, 7, 974–996. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, T. Cholera Toxin Subunit B as Adjuvant—An Accelerator in Protective Immunity and a Break in Autoimmunity. Vaccines 2015, 3, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, Y.; Harakuni, T.; Yamaguchi, R.; Miyata, T.; Arakawa, T. Cholera toxin B subunit pentamer reassembled from Escherichia coli inclusion bodies for use in vaccination. Vaccine 2016, 34, 1268–1274. [Google Scholar] [CrossRef]
- Harakuni, T.; Sugawa, H.; Komesu, A.; Tadano, M.; Arakawa, T. Heteropentameric Cholera Toxin B Subunit Chimeric Molecules Genetically Fused to a Vaccine Antigen Induce Systemic and Mucosal Immune Responses: A Potential New Strategy to Target Recombinant Vaccine Antigens to Mucosal Immune Systems. Infect. Immun. 2005, 73, 5654–5665. [Google Scholar] [CrossRef]
- Murrell, S.; Wu, S.-C.; Butler, M. Review of dengue virus and the development of a vaccine. Biotechnol. Adv. 2011, 29, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging technologies for low-cost, rapid vaccine manufacture. Biotechnol. J. 2019, 14, 1800376. [Google Scholar] [CrossRef]
- Choi, S.I.; Ryu, K.; Seong, B.L. RNA-mediated chaperone type for de novo protein folding. RNA Biol. 2009, 6, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-S.; Kim, Y.S.; Son, A.; Kim, J.; Kwon, S.B.; Kim, M.H.; Kim, P.; Kim, J.; Byun, Y.H.; Sung, J.; et al. Chaperna-mediated assembly of ferritin-based Middle East respiratory syndrome-coronavirus nanoparticles. Front. Immunol. 2018, 9, 1093. [Google Scholar] [CrossRef]
- Jobling, M.G.; Holmes, R.K. Mutational Analysis of Ganglioside GM1-Binding Ability, Pentamer Formation, and Epitopes of Cholera Toxin B (CTB) Subunits and CTB/Heat-Labile Enterotoxin B Subunit Chimeras. Infect. Immun. 2002, 70, 1260–1271. [Google Scholar] [CrossRef]
- Spangler, B.D. Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol. Rev. 1992, 56, 622–647. [Google Scholar] [CrossRef]
- Hwang, B.J.; Jang, Y.; Bin Kwon, S.; Yu, J.E.; Lim, J.; Roh, Y.H.; Seong, B.L. RNA-assisted self-assembly of monomeric antigens into virus-like particles as a recombinant vaccine platform. Biomaterials 2021, 269, 120650. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.B.; Yu, J.E.; Park, C.; Lee, J.; Seong, B.L. Nucleic acid-dependent structural transition of the intrinsically disordered N-terminal appended domain of human Lysyl-tRNA synthetase. Int. J. Mol. Sci. 2018, 19, 3016. [Google Scholar] [CrossRef]
- Ulmer, J.B.; Valley, U.; Rappuoli, R. Vaccine manufacturing: Challenges and solutions. Nat. Biotechnol. 2006, 24, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; O’Hagan, D. Advances in vaccine adjuvants. Nat. Biotechnol. 1999, 17, 1075–1081. [Google Scholar] [CrossRef]
- Black, M.; Trent, A.; Tirrell, M.; Olive, C. Advances in the design and delivery of peptide subunit vaccines with a focus on Toll-like receptor agonists. Expert Rev. Vaccines 2010, 9, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Burnette, W.N. Recombinant subunit vaccines. Curr. Opin. Biotechnol. 1991, 2, 882–892. [Google Scholar] [CrossRef]
- Valenzuela, P.; Medina, A.; Rutter, W.J.; Ammerer, G.; Hall, B.D. Synthesis and assembly of hepatitis B virus surface antigen particles in yeast. Nature 1982, 298, 347–350. [Google Scholar] [CrossRef]
- Sanchez, J.; Svennerholm, A.M.; Holmgren, J. Genetic fusion of a non-toxic heat-stable enterotoxin-related decapeptide antigen to cholera toxin B-subunit. FEBS Lett. 1988, 241, 110–114. [Google Scholar] [CrossRef]
- Shiloach, J.; Fass, R. Growing E. coli to high cell density—A historical perspective on method development. Biotechnol. Adv. 2005, 23, 345–357. [Google Scholar] [CrossRef]
- Coutelier, J.P.; van der Logt, J.T.; Heessen, F.W.; Warnier, G.; Van Snick, J. IgG2a restriction of murine antibodies elicited by viral infections. J. Exp. Med. 1987, 165, 64–69. [Google Scholar] [CrossRef]
- Simmons, M.; Hayes, C.G.; Kochel, T.; Raviprakash, K.; Murphy, G.S. Characterization of antibody responses to combinations of a dengue-2 DNA and dengue-2 recombinant subunit vaccine. Am. J. Trop. Med. Hyg. 2001, 65, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.C.; He, J.; Wu, C.-Y.; Kelsall, B.L. Cholera Toxin Suppresses Interleukin (IL)-12 Production and IL-12 Receptor β1 and β2 Chain Expression. J. Exp. Med. 1999, 189, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Boirivant, M.; Fuss, I.J.; Ferroni, L.; De Pascale, M.; Strober, W. Oral Administration of Recombinant Cholera Toxin Subunit B Inhibits IL-12-Mediated Murine Experimental (Trinitrobenzene Sulfonic Acid) Colitis. J. Immunol. 2001, 166, 3522–3532. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.-Q.; Yang, X.; Wei, Y.; Chen, J.-T.; Wang, X.-J.; Peng, H.-J. A Review on Dengue Vaccine Development. Vaccines 2020, 8, 63. [Google Scholar] [CrossRef]
- Fibriansah, G.; Tan, J.L.; Smith, S.A.; de Alwis, R.; Ng, T.-S.; Kostyuchenko, V.A.; Jadi, R.S.; Kukkaro, P.; de Silva, A.M.; Crowe, J.E.; et al. A highly potent human antibody neutralizes dengue virus serotype 3 by binding across three surface proteins. Nat. Commun. 2015, 6, 6341. [Google Scholar] [CrossRef]
- Halstead, S.; O’Rourke, E. Dengue viruses and mononuclear phagocytes. I. Infection enhancement by non-neutralizing antibody. J. Exp. Med. 1977, 146, 201–217. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sung, J.; Cheong, Y.; Kim, Y.-S.; Ahn, J.; Sohn, M.H.; Byun, S.; Seong, B.-L. Harnessing Pentameric Scaffold of Cholera Toxin B (CTB) for Design of Subvirion Recombinant Dengue Virus Vaccine. Vaccines 2024, 12, 92. https://doi.org/10.3390/vaccines12010092
Sung J, Cheong Y, Kim Y-S, Ahn J, Sohn MH, Byun S, Seong B-L. Harnessing Pentameric Scaffold of Cholera Toxin B (CTB) for Design of Subvirion Recombinant Dengue Virus Vaccine. Vaccines. 2024; 12(1):92. https://doi.org/10.3390/vaccines12010092
Chicago/Turabian StyleSung, Jemin, Yucheol Cheong, Young-Seok Kim, Jina Ahn, Myung Hyun Sohn, Sanguine Byun, and Baik-Lin Seong. 2024. "Harnessing Pentameric Scaffold of Cholera Toxin B (CTB) for Design of Subvirion Recombinant Dengue Virus Vaccine" Vaccines 12, no. 1: 92. https://doi.org/10.3390/vaccines12010092
APA StyleSung, J., Cheong, Y., Kim, Y. -S., Ahn, J., Sohn, M. H., Byun, S., & Seong, B. -L. (2024). Harnessing Pentameric Scaffold of Cholera Toxin B (CTB) for Design of Subvirion Recombinant Dengue Virus Vaccine. Vaccines, 12(1), 92. https://doi.org/10.3390/vaccines12010092