
Nucleic Acid Vaccines Encoding Proteins and Virus-like Particles for HIV Prevention

 ,
,
Abstract
:1. Introduction
2. Which Immunogens Should We Use and Why?
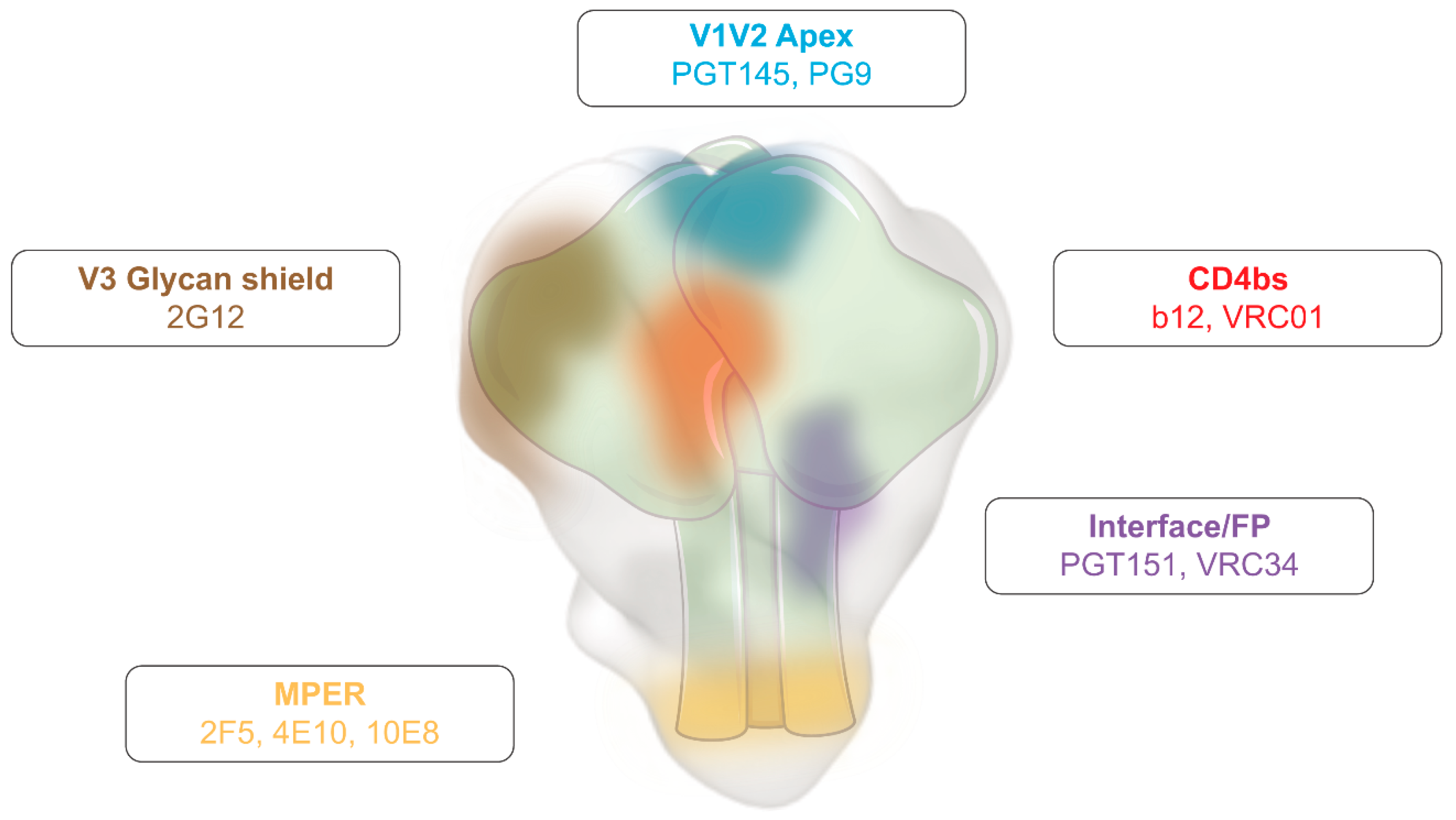
2.1. Elicitation of Protective Humoral Responses
2.2. Elicitation of Protective T Cell Responses
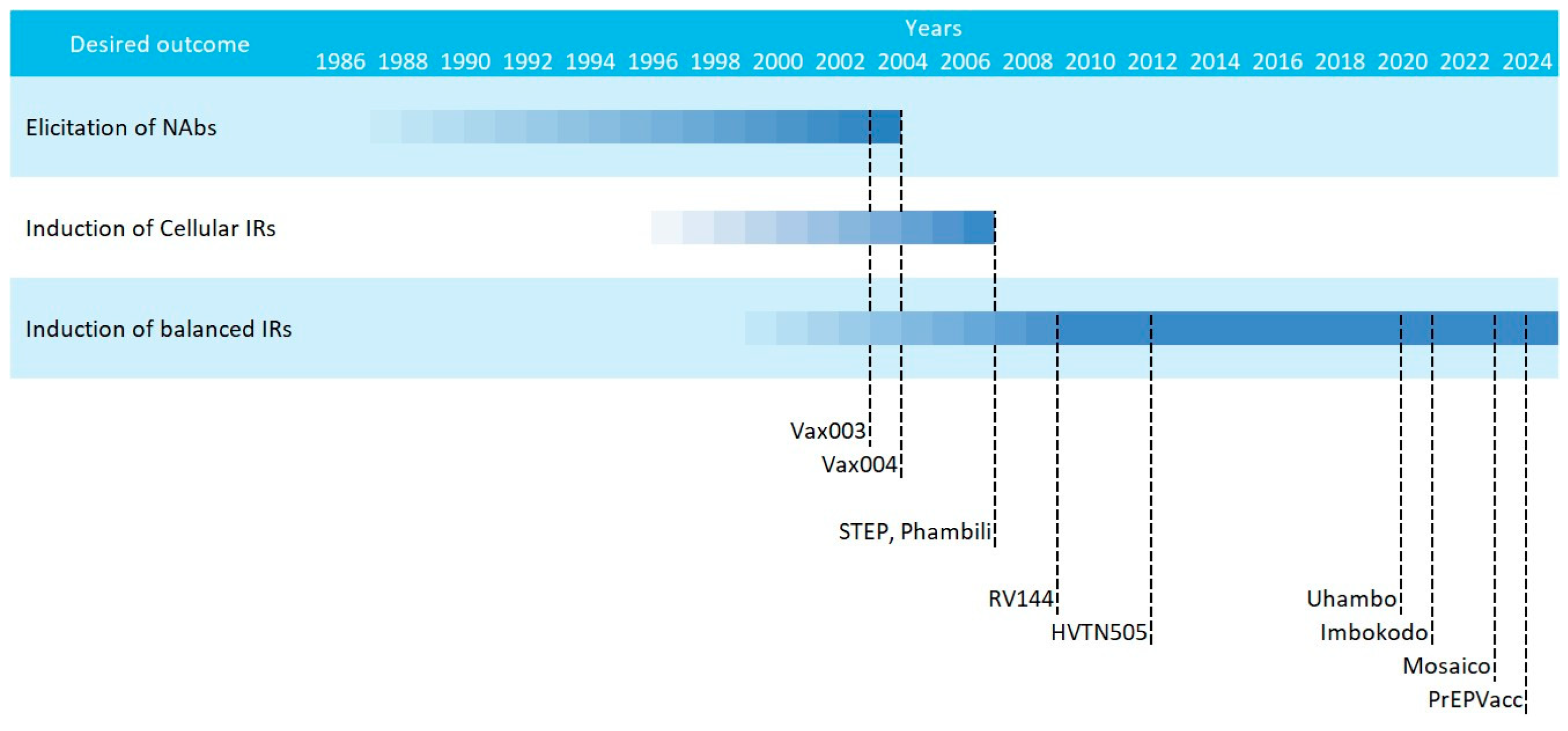
3. HIV Vaccine Efficacy Trials: Past Failures, Future Successes
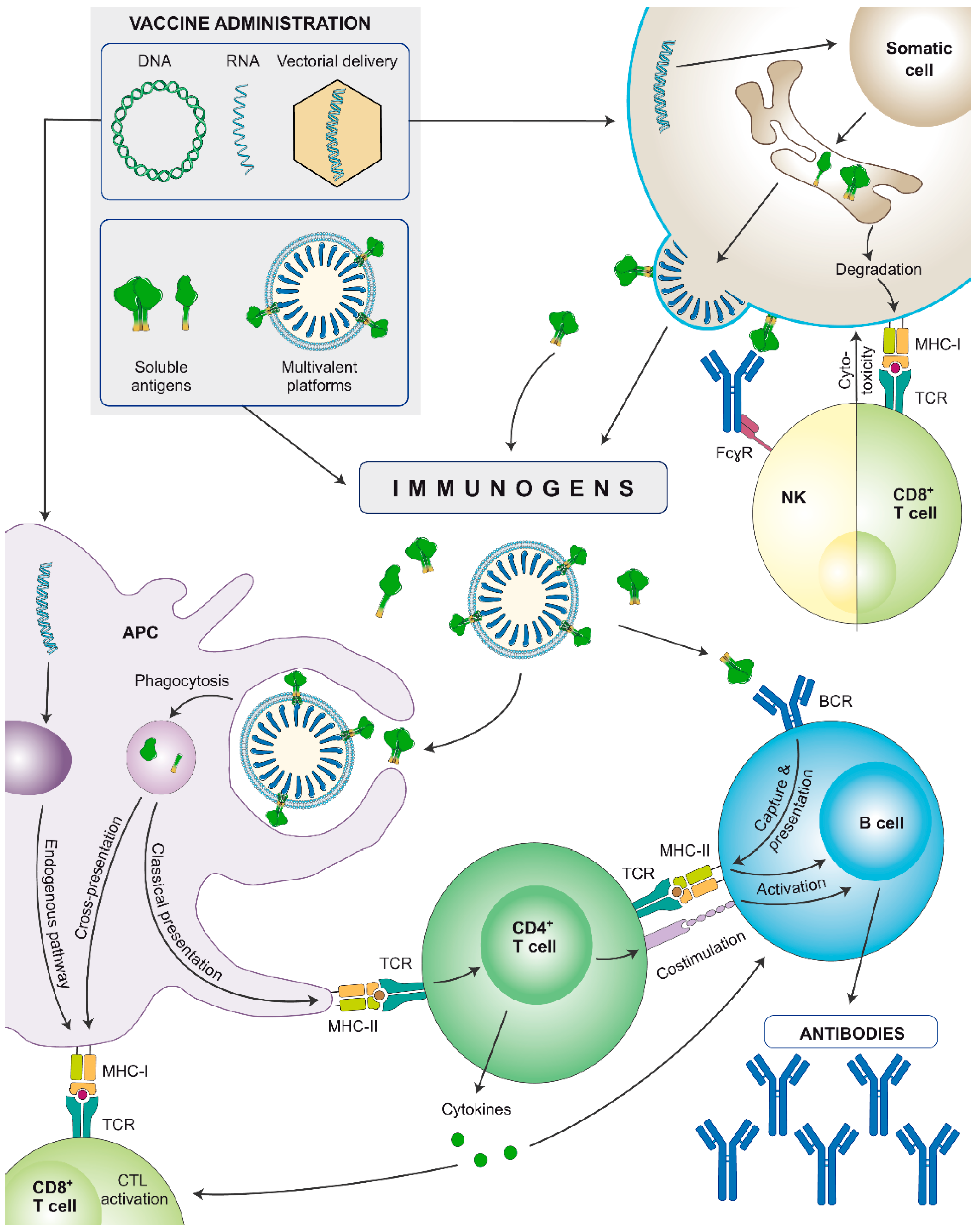
4. Nucleic Acid Vaccines and Their Delivery
4.1. Viral Vectored HIV Vaccines
4.2. DNA and mRNA HIV Vaccines
- -
- Versatility to encode various forms of antigens (no size limitation for encapsulation):
- ∘
- Subunit antigens
- ∘
- Membrane-bound antigens
- ∘
- Multivalent platforms
- -
- Unique production and purification strategies for all immunogens. No need to optimise downstream purification protocols since the host will be the bioreactor.
- -
- Fast sequence modification: easy to adapt to new threats (emerging pathogens or variants), which accelerates screening speed in preclinical trials.
- -
- Less adverse effects than viral vector vaccines (capillary leak syndrome, vector reactivation, anti-vector responses)
- -
- Global production and purification procedures have yet to be easily accessible in developing countries.
- -
- Poor thermostability: requires ultra-freezers for adequate conservation.
- -
- Elevated costs due to intellectual property protection compared to conventional vaccine platforms (viral vectors, inactivated pathogens, subunit proteins).
- -
- Heterogeneous protein production. This limitation is especially relevant for mRNA delivery of proteins in which the therapeutic window is narrower.
4.2.1. Nucleic Acid Vaccines Encoding Viral Particles
4.2.2. Nucleic Acid Vaccines Encoding Virus-like Particles
4.3. Summary and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Rappuoli, R.; Pizza, M.; Del Giudice, G.; De Gregorio, E. Vaccines, New Opportunities for a New Society. Proc. Natl. Acad. Sci. USA 2014, 111, 12288–12293. [Google Scholar] [CrossRef]
- Vanderslott, S.; Dattani, S.; Spooner, F.; Roser, M. Vaccination. Available online: https://ourworldindata.org/vaccination (accessed on 27 January 2024).
- Marani, M.; Katul, G.G.; Pan, W.K.; Parolari, A.J. Intensity and Frequency of Extreme Novel Epidemics. Proc. Natl. Acad. Sci. USA 2021, 118, e2105482118. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Fernández, M.; Melguizo-Rodríguez, L.; Costela-Ruiz, V.J.; González-Acedo, A.; Ramos-Torrecillas, J.; Illescas-Montes, R. The Current Status of COVID-19 Vaccines. A Scoping Review. Drug Discov. Today 2022, 27, 103336. [Google Scholar] [CrossRef]
- Khobragade, A.; Bhate, S.; Ramaiah, V.; Deshpande, S.; Giri, K.; Phophle, H.; Supe, P.; Godara, I.; Revanna, R.; Nagarkar, R.; et al. Efficacy, Safety, and Immunogenicity of the DNA SARS-CoV-2 Vaccine (ZyCoV-D): The Interim Efficacy Results of a Phase 3, Randomised, Double-Blind, Placebo-Controlled Study in India. Lancet 2022, 399, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, Y.; Tong, X.; Kang, J.; Avendaño, M.J.; Serrano, E.F.; García-Salum, T.; Pardo-Roa, C.; Riquelme, A.; Medina, R.A.; Alter, G. Preserved Omicron Spike Specific Antibody Binding and Fc-Recognition across COVID-19 Vaccine Platforms. medRxiv 2021. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA COVID-19 Vaccine. New Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Krienke, C.; Kolb, L.; Diken, E.; Streuber, M.; Kirchhoff, S.; Bukur, T.; Akilli-Öztürk, Ö.; Kranz, L.M.; Berger, H.; Petschenka, J.; et al. A Noninflammatory MRNA Vaccine for Treatment of Experimental Autoimmune Encephalomyelitis. Science 2021, 371, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Kranz, L.M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K.C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; et al. Systemic RNA Delivery to Dendritic Cells Exploits Antiviral Defence for Cancer Immunotherapy. Nature 2016, 534, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; de Souza, M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand. New Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Gray, G.E.; Allen, M.; Moodie, Z.; Churchyard, G.; Bekker, L.-G.; Nchabeleng, M.; Mlisana, K.; Metch, B.; de Bruyn, G.; Latka, M.H.; et al. Safety and Efficacy of the HVTN 503/Phambili Study of a Clade-B-Based HIV-1 Vaccine in South Africa: A Double-Blind, Randomised, Placebo-Controlled Test-of-Concept Phase 2b Study. Lancet Infect. Dis. 2011, 11, 507–515. [Google Scholar] [CrossRef]
- Moodie, Z.; Dintwe, O.; Sawant, S.; Grove, D.; Huang, Y.; Janes, H.; Heptinstall, J.; Omar, F.L.; Cohen, K.; De Rosa, S.C.; et al. Analysis of the HIV Vaccine Trials Network 702 Phase 2b-3 HIV-1 Vaccine Trial in South Africa Assessing RV144 Antibody and T-Cell Correlates of HIV-1 Acquisition Risk. J. Infect. Dis. 2022, 226, 246–257. [Google Scholar] [CrossRef]
- Crabtree Ramírez, B.; González Hernández, L.A.; Cabrera, C.; del Río, C.; González Rodríguez, A.; Sierra Madero, J. Mexican Perspective on the Mosaico HIV Vaccine Trial. Lancet HIV 2023, 10, e426–e427. [Google Scholar] [CrossRef]
- Flynn, N.M.; Forthal, D.N.; Harro, C.D.; Judson, F.N.; Mayer, K.H.; Para, M.F. Placebo-Controlled Phase 3 Trial of a Recombinant Glycoprotein 120 Vaccine to Prevent HIV-1 Infection. J. Infect. Dis. 2005, 191, 654–665. [Google Scholar] [CrossRef]
- Pitisuttithum, P.; Gilbert, P.; Gurwith, M.; Heyward, W.; Martin, M.; van Griensven, F.; Hu, D.; Tappero, J.W.; Choopanya, K. Randomized, Double-Blind, Placebo-Controlled Efficacy Trial of a Bivalent Recombinant Glycoprotein 120 HIV-1 Vaccine among Injection Drug Users in Bangkok, Thailand. J. Infect. Dis. 2006, 194, 1661–1671. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Sanders, R.W.; Moore, J.P. Virus Vaccines: Proteins Prefer Prolines. Cell Host Microbe 2021, 29, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Esteban, I.; Pastor-Quiñones, C.; Usero, L.; Plana, M.; García, F.; Leal, L. In the Era of MRNA Vaccines, Is There Any Hope for HIV Functional Cure? Viruses 2021, 13, 501. [Google Scholar] [CrossRef] [PubMed]
- Reichmuth, A.M.; Oberli, M.A.; Jaklenec, A.; Langer, R.; Blankschtein, D. MRNA Vaccine Delivery Using Lipid Nanoparticles. Ther. Deliv. 2016, 7, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Iketani, S.; Guo, Y.; Chan, J.F.-W.; Wang, M.; Liu, L.; Luo, Y.; Chu, H.; Huang, Y.; Nair, M.S.; et al. Striking Antibody Evasion Manifested by the Omicron Variant of SARS-CoV-2. Nature 2022, 602, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.F.; Wiehe, K.; Borrow, P.; Saunders, K.O.; Korber, B.; Wagh, K.; McMichael, A.J.; Kelsoe, G.; Hahn, B.H.; Alt, F.; et al. Author Correction: Strategies for HIV-1 Vaccines That Induce Broadly Neutralizing Antibodies. Nat. Rev. Immunol. 2023, 23, 265. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Brander, C. HIV T-Cell Vaccines. Adv. Exp. Med. Biol. 2018, 1075, 31–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, B. Molecular Mechanism of HIV-1 Entry. Trends Microbiol. 2019, 27, 878–891. [Google Scholar] [CrossRef]
- Burton, D.R.; Hangartner, L. Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu. Rev. Immunol. 2016, 34, 635–659. [Google Scholar] [CrossRef]
- Kesavardhana, S.; Varadarajan, R. Stabilizing the Native Trimer of HIV-1 Env by Destabilizing the Heterodimeric Interface of the Gp41 Postfusion Six-Helix Bundle. J. Virol. 2014, 88, 9590–9604. [Google Scholar] [CrossRef]
- Masemola, A.; Mashishi, T.; Khoury, G.; Mohube, P.; Mokgotho, P.; Vardas, E.; Colvin, M.; Zijenah, L.; Katzenstein, D.; Musonda, R.; et al. Hierarchical Targeting of Subtype C Human Immunodeficiency Virus Type 1 Proteins by CD8+ T Cells: Correlation with Viral Load. J. Virol. 2004, 78, 3233–3243. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J. The Origin and Diversity of the HIV-1 Pandemic. Trends Mol. Med. 2012, 18, 182–192. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.J.; Scharf, L.; Scheid, J.F.; Klein, F.; Bjorkman, P.J.; Nussenzweig, M.C. Structural Insights on the Role of Antibodies in HIV-1 Vaccine and Therapy. Cell 2014, 156, 633–648. [Google Scholar] [CrossRef]
- Sadanand, S.; Suscovich, T.J.; Alter, G. Broadly Neutralizing Antibodies Against HIV: New Insights to Inform Vaccine Design. Annu. Rev. Med. 2016, 67, 185–200. [Google Scholar] [CrossRef]
- Molinos-Albert, L.M.; Clotet, B.; Blanco, J.; Carrillo, J. Immunologic Insights on the Membrane Proximal External Region: A Major Human Immunodeficiency Virus Type-1 Vaccine Target. Front. Immunol. 2017, 8, 1154. [Google Scholar] [CrossRef]
- Bonsignori, M.; Liao, H.-X.; Gao, F.; Williams, W.B.; Alam, S.M.; Montefiori, D.C.; Haynes, B.F. Antibody-Virus Co-Evolution in HIV Infection: Paths for HIV Vaccine Development. Immunol. Rev. 2017, 275, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Gaebler, C.; Nogueira, L.; Stoffel, E.; Oliveira, T.Y.; Breton, G.; Millard, K.G.; Turroja, M.; Butler, A.; Ramos, V.; Seaman, M.S.; et al. Prolonged Viral Suppression with Anti-HIV-1 Antibody Therapy. Nature 2022, 606, 368–374. [Google Scholar] [CrossRef]
- Niessl, J.; Baxter, A.E.; Mendoza, P.; Jankovic, M.; Cohen, Y.Z.; Butler, A.L.; Lu, C.L.; Dubé, M.; Shimeliovich, I.; Gruell, H.; et al. Combination Anti-HIV-1 Antibody Therapy Is Associated with Increased Virus-Specific T Cell Immunity. Nat. Med. 2020, 26, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, P.; Gruell, H.; Nogueira, L.; Pai, J.A.; Butler, A.L.; Millard, K.; Lehmann, C.; Suárez, I.; Oliveira, T.Y.; Lorenzi, J.C.C.; et al. Combination Therapy with Anti-HIV-1 Antibodies Maintains Viral Suppression. Nature 2018, 561, 479. [Google Scholar] [CrossRef] [PubMed]
- Simek, M.D.; Rida, W.; Priddy, F.H.; Pung, P.; Carrow, E.; Laufer, D.S.; Lehrman, J.K.; Boaz, M.; Tarragona-Fiol, T.; Miiro, G.; et al. Human Immunodeficiency Virus Type 1 Elite Neutralizers: Individuals with Broad and Potent Neutralizing Activity Identified by Using a High-Throughput Neutralization Assay Together with an Analytical Selection Algorithm. J. Virol. 2009, 83, 7337–7348. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, L. Broadly Neutralizing Antibodies and Vaccine Design against HIV-1 Infection. Front. Med. 2020, 14, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Subbaraman, H.; Schanz, M.; Trkola, A. Broadly Neutralizing Antibodies: What Is Needed to Move from a Rare Event in HIV-1 Infection to Vaccine Efficacy? Retrovirology 2018, 15, 52. [Google Scholar] [CrossRef] [PubMed]
- Julg, B.; Liu, P.-T.; Wagh, K.; Fischer, W.M.; Abbink, P.; Mercado, N.B.; Whitney, J.B.; Nkolola, J.P.; McMahan, K.; Tartaglia, L.J.; et al. Protection against a Mixed SHIV Challenge by a Broadly Neutralizing Antibody Cocktail. Sci. Transl. Med. 2017, 9, eaao4235. [Google Scholar] [CrossRef] [PubMed]
- Julg, B.; Barouch, D.H. Neutralizing Antibodies for HIV-1 Prevention. Curr. Opin. HIV AIDS 2019, 14, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Gilbert, P.B.; Juraska, M.; Montefiori, D.C.; Morris, L.; Karuna, S.T.; Edupuganti, S.; Mgodi, N.M.; deCamp, A.C.; Rudnicki, E.; et al. Two Randomized Trials of Neutralizing Antibodies to Prevent HIV-1 Acquisition. New Engl. J. Med. 2021, 384, 1003–1014. [Google Scholar] [CrossRef]
- Rusche, J.R.; Lynn, D.L.; Robert-Guroff, M.; Langlois, A.J.; Lyerly, H.K.; Carson, H.; Krohn, K.; Ranki, A.; Gallo, R.C.; Bolognesi, D.P. Humoral Immune Response to the Entire Human Immunodeficiency Virus Envelope Glycoprotein Made in Insect Cells. Proc. Natl. Acad. Sci. USA 1987, 84, 6924–6928. [Google Scholar] [CrossRef]
- Steimer, K.S.; Scandella, C.J.; Skiles, P.V.; Haigwood, N.L. Neutralization of Divergent HIV-1 Isolates by Conformation-Dependent Human Antibodies to Gp120. Science 1991, 254, 105–108. [Google Scholar] [CrossRef]
- Billich, A. AIDSVAX. VaxGen. Curr. Opin. Investig. Drugs 2001, 2, 1203–1208. [Google Scholar]
- Rerks-Ngarm, S.; Brown, A.E.; Khamboonruang, C.; Thongcharoen, P.; Kunasol, P. HIV/AIDS Preventive Vaccine ‘Prime-Boost’ Phase III Trial: Foundations and Initial Lessons Learned from Thailand. AIDS 2006, 20, 1471–1479. [Google Scholar]
- Jardine, J.G.; Kulp, D.W.; Havenar-Daughton, C.; Sarkar, A.; Briney, B.; Sok, D.; Sesterhenn, F.; Ereño-Orbea, J.; Kalyuzhniy, O.; Deresa, I.; et al. HIV-1 Broadly Neutralizing Antibody Precursor B Cells Revealed by Germline-Targeting Immunogen. Science 2016, 351, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Caillat, C.; Guilligay, D.; Sulbaran, G.; Weissenhorn, W. Neutralizing Antibodies Targeting HIV-1 Gp41. Viruses 2020, 12, 1210. [Google Scholar] [CrossRef]
- Sanders, R.W.; Vesanen, M.; Schuelke, N.; Master, A.; Schiffner, L.; Kalyanaraman, R.; Paluch, M.; Berkhout, B.; Maddon, P.J.; Olson, W.C.; et al. Stabilization of the Soluble, Cleaved, Trimeric Form of the Envelope Glycoprotein Complex of Human Immunodeficiency Virus Type 1. J. Virol. 2002, 76, 8875–8889. [Google Scholar] [CrossRef] [PubMed]
- Pauthner, M.G.; Nkolola, J.P.; Havenar-Daughton, C.; Murrell, B.; Reiss, S.M.; Bastidas, R.; Prévost, J.; Nedellec, R.; von Bredow, B.; Abbink, P.; et al. Vaccine-Induced Protection from Homologous Tier 2 SHIV Challenge in Nonhuman Primates Depends on Serum-Neutralizing Antibody Titers. Immunity 2019, 50, 241–252.e6. [Google Scholar] [CrossRef]
- Sliepen, K.; Han, B.W.; Bontjer, I.; Mooij, P.; Garces, F.; Behrens, A.-J.; Rantalainen, K.; Kumar, S.; Sarkar, A.; Brouwer, P.J.M.; et al. Structure and Immunogenicity of a Stabilized HIV-1 Envelope Trimer Based on a Group-M Consensus Sequence. Nat. Commun. 2019, 10, 2355. [Google Scholar] [CrossRef] [PubMed]
- Dubrovskaya, V.; Tran, K.; Ozorowski, G.; Guenaga, J.; Wilson, R.; Bale, S.; Cottrell, C.A.; Turner, H.L.; Seabright, G.; O’Dell, S.; et al. Vaccination with Glycan-Modified HIV NFL Envelope Trimer-Liposomes Elicits Broadly Neutralizing Antibodies to Multiple Sites of Vulnerability. Immunity 2019, 51, 915–929.e7. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Paynter, J.; Antanasijevic, A.; Allen, J.D.; Eldad, M.; Lee, Y.Z.; Copps, J.; Newby, M.L.; He, L.; Chavez, D.; et al. Single-Component Multilayered Self-Assembling Protein Nanoparticles Presenting Glycan-Trimmed Uncleaved Prefusion Optimized Envelope Trimmers as HIV-1 Vaccine Candidates. Nat. Commun. 2023, 14, 1985. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Excler, J.-L.; Michael, N.L. Lessons from the RV144 Thai Phase III HIV-1 Vaccine Trial and the Search for Correlates of Protection. Annu. Rev. Med. 2015, 66, 423–437. [Google Scholar] [CrossRef]
- Moodie, Z.; Walsh, S.R.; Laher, F.; Maganga, L.; Herce, M.E.; Naidoo, S.; Hosseinipour, M.C.; Innes, C.; Bekker, L.-G.; Grunenberg, N.; et al. Antibody and Cellular Responses to HIV Vaccine Regimens with DNA Plasmid as Compared with ALVAC Priming: An Analysis of Two Randomized Controlled Trials. PLoS Med. 2020, 17, e1003117. [Google Scholar] [CrossRef]
- Gray, G.E.; Bekker, L.-G.; Laher, F.; Malahleha, M.; Allen, M.; Moodie, Z.; Grunenberg, N.; Huang, Y.; Grove, D.; Prigmore, B.; et al. Vaccine Efficacy of ALVAC-HIV and Bivalent Subtype C Gp120-MF59 in Adults. New Engl. J. Med. 2021, 384, 1089–1100. [Google Scholar] [CrossRef]
- Hosseinipour, M.C.; Innes, C.; Naidoo, S.; Mann, P.; Hutter, J.; Ramjee, G.; Sebe, M.; Maganga, L.; Herce, M.E.; deCamp, A.C.; et al. Phase 1 Human Immunodeficiency Virus (HIV) Vaccine Trial to Evaluate the Safety and Immunogenicity of HIV Subtype C DNA and MF59-Adjuvanted Subtype C Envelope Protein. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 72, 50–60. [Google Scholar] [CrossRef]
- Nielsen, C.M.; Ogbe, A.; Pedroza-Pacheco, I.; Doeleman, S.E.; Chen, Y.; Silk, S.E.; Barrett, J.R.; Elias, S.C.; Miura, K.; Diouf, A.; et al. Protein/AS01B Vaccination Elicits Stronger, More Th2-Skewed Antigen-Specific Human T Follicular Helper Cell Responses than Heterologous Viral Vectors. Cell Rep. Med. 2021, 2, 100207. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, M.; Wang, T. Liposomes Used as a Vaccine Adjuvant-Delivery System: From Basics to Clinical Immunization. J. Control. Release 2019, 303, 130–150. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-Correlates Analysis of an HIV-1 Vaccine Efficacy Trial. New Engl. J. Med. 2012, 366, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Wren, L.; Kent, S.J. HIV Vaccine Efficacy Trial: Glimmers of Hope and the Potential Role of Antibody-Dependent Cellular Cytotoxicity. Hum. Vaccin. 2011, 7, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Corey, L.; Gilbert, P.B.; Tomaras, G.D.; Haynes, B.F.; Pantaleo, G.; Fauci, A.S. Immune Correlates of Vaccine Protection against HIV-1 Acquisition: A Review. Sci. Transl. Med. 2015, 7, 310rv7. [Google Scholar] [CrossRef]
- Kracker, S.; Radbruch, A. Immunoglobulin Class Switching: In Vitro Induction and Analysis. Methods Mol. Biol. 2004, 271, 149–159. [Google Scholar] [CrossRef]
- McMichael, A.J.; Rowland-Jones, S.L. Cellular Immune Responses to HIV. Nature 2001, 410, 980–987. [Google Scholar] [CrossRef]
- Gray, E.R.; Bain, R.; Varsaneux, O.; Peeling, R.W.; Stevens, M.M.; McKendry, R.A. P24 Revisited: A Landscape Review of Antigen Detection for Early HIV Diagnosis. AIDS 2018, 32, 2089–2102. [Google Scholar] [CrossRef]
- Coplan, P.M.; Gupta, S.B.; Dubey, S.A.; Pitisuttithum, P.; Nikas, A.; Mbewe, B.; Vardas, E.; Schechter, M.; Kallas, E.G.; Freed, D.C.; et al. Cross-Reactivity of Anti-HIV-1 T Cell Immune Responses among the Major HIV-1 Clades in HIV-1-Positive Individuals from 4 Continents. J. Infect. Dis. 2005, 191, 1427–1434. [Google Scholar] [CrossRef]
- Laher, F.; Ranasinghe, S.; Porichis, F.; Mewalal, N.; Pretorius, K.; Ismail, N.; Buus, S.; Stryhn, A.; Carrington, M.; Walker, B.D.; et al. HIV Controllers Exhibit Enhanced Frequencies of Major Histocompatibility Complex Class II Tetramer(+) Gag-Specific CD4(+) T Cells in Chronic Clade C HIV-1 Infection. J. Virol. 2017, 91, e02477-16. [Google Scholar] [CrossRef]
- Pernas, M.; Tarancón-Diez, L.; Rodríguez-Gallego, E.; Gómez, J.; Prado, J.G.; Casado, C.; Dominguez-Molina, B.; Olivares, I.; Coiras, M.; León, A.; et al. Factors Leading to the Loss of Natural Elite Control of HIV-1 Infection. J. Virol. 2018, 92, e01805-17. [Google Scholar] [CrossRef]
- Payne, R.P.; Kløverpris, H.; Sacha, J.B.; Brumme, Z.; Brumme, C.; Buus, S.; Sims, S.; Hickling, S.; Riddell, L.; Chen, F.; et al. Efficacious Early Antiviral Activity of HIV Gag- and Pol-Specific HLA-B 2705-Restricted CD8+ T Cells. J. Virol. 2010, 84, 10543–10557. [Google Scholar] [CrossRef] [PubMed]
- Sundaramurthi, J.C.; Ashokkumar, M.; Swaminathan, S.; Hanna, L.E. HLA Based Selection of Epitopes Offers a Potential Window of Opportunity for Vaccine Design against HIV. Vaccine 2017, 35, 5568–5575. [Google Scholar] [PubMed]
- Rouphael, N.G.; Morgan, C.; Li, S.S.; Jensen, R.; Sanchez, B.; Karuna, S.; Swann, E.; Sobieszczyk, M.E.; Frank, I.; Wilson, G.J.; et al. DNA Priming and Gp120 Boosting Induces HIV-Specific Antibodies in a Randomized Clinical Trial. J. Clin. Investig. 2019, 129, 4769–4785. [Google Scholar] [CrossRef] [PubMed]
- Sekaly, R.-P. The Failed HIV Merck Vaccine Study: A Step Back or a Launching Point for Future Vaccine Development? J. Exp. Med. 2008, 205, 7–12. [Google Scholar] [CrossRef]
- Msafiri, F.; Joachim, A.; Held, K.; Nadai, Y.; Chissumba, R.M.; Geldmacher, C.; Aboud, S.; Stöhr, W.; Viegas, E.; Kroidl, A.; et al. Frequent Anti-V1V2 Responses Induced by HIV-DNA Followed by HIV-MVA with or without CN54rgp140/GLA-AF in Healthy African Volunteers. Microorganisms 2020, 8, 1722. [Google Scholar] [CrossRef] [PubMed]
- Perdiguero, B.; Pérez, P.; Marcos-Villar, L.; Albericio, G.; Astorgano, D.; Álvarez, E.; Sin, L.; Gómez, C.E.; García-Arriaza, J.; Esteban, M. Highly Attenuated Poxvirus-Based Vaccines Against Emerging Viral Diseases. J. Mol. Biol. 2023, 435, 168173. [Google Scholar] [CrossRef]
- Kajon, A.E.; Weinberg, J.B.; Spindler, K.R. Adenoviruses. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-12-801238-3. [Google Scholar]
- Barouch, D.H.; O’Brien, K.L.; Simmons, N.L.; King, S.L.; Abbink, P.; Maxfield, L.F.; Sun, Y.-H.; La Porte, A.; Riggs, A.M.; Lynch, D.M.; et al. Mosaic HIV-1 Vaccines Expand the Breadth and Depth of Cellular Immune Responses in Rhesus Monkeys. Nat. Med. 2010, 16, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Ondondo, B.; Murakoshi, H.; Clutton, G.; Abdul-Jawad, S.; Wee, E.G.-T.; Gatanaga, H.; Oka, S.; McMichael, A.J.; Takiguchi, M.; Korber, B.; et al. Novel Conserved-Region T-Cell Mosaic Vaccine With High Global HIV-1 Coverage Is Recognized by Protective Responses in Untreated Infection. Mol. Ther. 2016, 24, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Hu, X.; Llano, A.; Rosati, M.; Olvera, A.; Kulkarni, V.; Valentin, A.; Alicea, C.; Pilkington, G.R.; Sardesai, N.Y.; et al. A Human Immune Data-Informed Vaccine Concept Elicits Strong and Broad T-Cell Specificities Associated with HIV-1 Control in Mice and Macaques. J. Transl. Med. 2015, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Laher, F.; Bekker, L.-G.; Garrett, N.; Lazarus, E.M.; Gray, G.E. Review of Preventative HIV Vaccine Clinical Trials in South Africa. Arch. Virol. 2020, 165, 2439–2452. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Crotty, S. HIV Vaccinology: 2021 Update. Semin. Immunol. 2021, 51, 101470. [Google Scholar] [CrossRef]
- Esparza, J. A Brief History of the Global Effort to Develop a Preventive HIV Vaccine. Vaccine 2013, 31, 3502–3518. [Google Scholar] [CrossRef]
- Rhee, S.-Y.; Shafer, R.W. Geographically-Stratified HIV-1 Group M Pol Subtype and Circulating Recombinant Form Sequences. Sci. Data 2018, 5, 180148. [Google Scholar] [CrossRef]
- Gilbert, P.B.; Peterson, M.L.; Follmann, D.; Hudgens, M.G.; Francis, D.P.; Gurwith, M.; Heyward, W.L.; Jobes, D.V.; Popovic, V.; Self, S.G.; et al. Correlation between Immunologic Responses to a Recombinant Glycoprotein 120 Vaccine and Incidence of HIV-1 Infection in a Phase 3 HIV-1 Preventive Vaccine Trial. J. Infect. Dis. 2005, 191, 666–677. [Google Scholar] [CrossRef]
- Harrer, T.; Harrer, E.; Kalams, S.A.; Elbeik, T.; Staprans, S.I.; Feinberg, M.B.; Cao, Y.; Ho, D.D.; Yilma, T.; Caliendo, A.M.; et al. Strong Cytotoxic T Cell and Weak Neutralizing Antibody Responses in a Subset of Persons with Stable Nonprogressing HIV Type 1 Infection. AIDS Res. Hum. Retroviruses 1996, 12, 585–592. [Google Scholar] [CrossRef]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; Del Rio, C.; et al. Efficacy Assessment of a Cell-Mediated Immunity HIV-1 Vaccine (the Step Study): A Double-Blind, Randomised, Placebo-Controlled, Test-of-Concept Trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R.; Desrosiers, R.C.; Doms, R.W.; Feinberg, M.B.; Gallo, R.C.; Hahn, B.; Hoxie, J.A.; Hunter, E.; Korber, B.; Landay, A.; et al. A Sound Rationale Needed for Phase III HIV-1 Vaccine Trials. Science 2004, 303, 316. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.; Franchini, G.; Girard, M.P.; Gotch, F.; Kaleebu, P.; Marthas, M.L.; McChesney, M.B.; McCullough, R.; Mhalu, F.; Salmon-Ceron, D.; et al. Support for the RV144 HIV Vaccine Trial. Science 2004, 305, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Russell, N.D.; Marovich, M.A. Pox-Protein Public Private Partnership Program and Upcoming HIV Vaccine Efficacy Trials. Curr. Opin. HIV AIDS 2016, 11, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Hammer, S.M.; Sobieszczyk, M.E.; Janes, H.; Karuna, S.T.; Mulligan, M.J.; Grove, D.; Koblin, B.A.; Buchbinder, S.P.; Keefer, M.C.; Tomaras, G.D.; et al. Efficacy Trial of a DNA/RAd5 HIV-1 Preventive Vaccine. New Engl. J. Med. 2013, 369, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Janes, H.E.; Cohen, K.W.; Frahm, N.; De Rosa, S.C.; Sanchez, B.; Hural, J.; Magaret, C.A.; Karuna, S.; Bentley, C.; Gottardo, R.; et al. Higher T-Cell Responses Induced by DNA/RAd5 HIV-1 Preventive Vaccine Are Associated with Lower HIV-1 Infection Risk in an Efficacy Trial. J. Infect. Dis. 2017, 215, 1376–1385. [Google Scholar] [CrossRef] [PubMed]
- Bekker, L.-G.; Moodie, Z.; Grunenberg, N.; Laher, F.; Tomaras, G.D.; Cohen, K.W.; Allen, M.; Malahleha, M.; Mngadi, K.; Daniels, B.; et al. Subtype C ALVAC-HIV and Bivalent Subtype C Gp120/MF59 HIV-1 Vaccine in Low-Risk, HIV-Uninfected, South African Adults: A Phase 1/2 Trial. Lancet HIV 2018, 5, e366–e378. [Google Scholar] [CrossRef]
- Baden, L.R.; Stieh, D.J.; Sarnecki, M.; Walsh, S.R.; Tomaras, G.D.; Kublin, J.G.; McElrath, M.J.; Alter, G.; Ferrari, G.; Montefiori, D.; et al. Safety and Immunogenicity of Two Heterologous HIV Vaccine Regimens in Healthy, HIV-Uninfected Adults (TRAVERSE): A Randomised, Parallel-Group, Placebo-Controlled, Double-Blind, Phase 1/2a Study. Lancet HIV 2020, 7, e688–e698. [Google Scholar] [CrossRef]
- Barouch, D.H.; Tomaka, F.L.; Wegmann, F.; Stieh, D.J.; Alter, G.; Robb, M.L.; Michael, N.L.; Peter, L.; Nkolola, J.P.; Borducchi, E.N.; et al. Evaluation of a Mosaic HIV-1 Vaccine in a Multicentre, Randomised, Double-Blind, Placebo-Controlled, Phase 1/2a Clinical Trial (APPROACH) and in Rhesus Monkeys (NHP 13-19). Lancet 2018, 392, 232–243. [Google Scholar] [CrossRef]
- Gray, G. Phase 2b efficacy trial of mosaic HIV-1 vaccine regimen in African women (Imbokodo). In Proceedings of the Conference on Retroviruses and Opportunistic Infections (CROI), Virtual, 12–16 February 2022. [Google Scholar]
- IAVI. IAVI Statement on PrEPVacc Trial. Available online: https://www.iavi.org/features/iavi-statement-on-prepvacc-trial/ (accessed on 28 January 2024).
- Abbasi, J. First MRNA HIV Vaccine Clinical Trial Launches. JAMA 2022, 327, 909. [Google Scholar] [CrossRef]
- Fortner, A.; Bucur, O. MRNA-Based Vaccine Technology for HIV. Discoveries 2022, 10, e150. [Google Scholar] [CrossRef]
- Shepherd, B.O.; Chang, D.; Vasan, S.; Ake, J.; Modjarrad, K. HIV and SARS-CoV-2: Tracing a Path of Vaccine Research and Development. Curr. HIV/AIDS Rep. 2022, 19, 86–93. [Google Scholar] [CrossRef]
- Hobernik, D.; Bros, M. DNA Vaccines-How Far From Clinical Use? Int. J. Mol. Sci. 2018, 19, 3605. [Google Scholar] [CrossRef] [PubMed]
- Ertl, H.C.J. Viral Vectors as Vaccine Carriers. Curr. Opin. Virol. 2016, 21, 1–8. [Google Scholar] [CrossRef]
- Melzi, E.; Willis, J.R.; Ma, K.M.; Lin, Y.C.; Kratochvil, S.; Berndsen, Z.T.; Landais, E.A.; Kalyuzhniy, O.; Nair, U.; Warner, J.; et al. Membrane-Bound MRNA Immunogens Lower the Threshold to Activate HIV Env V2 Apex-Directed Broadly Neutralizing B Cell Precursors in Humanized Mice. Immunity 2022, 55, 2168–2186.e6. [Google Scholar] [CrossRef]
- Cari, L.; Naghavi Alhosseini, M.; Mencacci, A.; Migliorati, G.; Nocentini, G. Differences in the Expression Levels of SARS-CoV-2 Spike Protein in Cells Treated with MRNA-Based COVID-19 Vaccines: A Study on Vaccines from the Real World. Vaccines 2023, 11, 879. [Google Scholar] [CrossRef]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and Challenges in the Delivery of MRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, H.; Meng, L.; Li, F.; Yu, C. Comparison of Immune Responses Elicited by SARS-CoV-2 MRNA and Recombinant Protein Vaccine Candidates. Front. Immunol. 2022, 13, 906457. [Google Scholar] [CrossRef] [PubMed]
- Barry, M. Single-Cycle Adenovirus Vectors in the Current Vaccine Landscape. Expert Rev. Vaccines 2018, 17, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Krüger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron Variant Is Highly Resistant against Antibody-Mediated Neutralization: Implications for Control of the COVID-19 Pandemic. Cell 2022, 185, 447–456.e11. [Google Scholar] [CrossRef] [PubMed]
- Nkolola, J.P.; Barouch, D.H. Prophylactic HIV-1 Vaccine Trials: Past, Present, and Future. Lancet HIV 2023, 11, E117–E124. [Google Scholar] [CrossRef] [PubMed]
- Sasso, E.; D’Alise, A.M.; Zambrano, N.; Scarselli, E.; Folgori, A.; Nicosia, A. New Viral Vectors for Infectious Diseases and Cancer. Semin. Immunol. 2020, 50, 101430. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R. Advancing an HIV Vaccine; Advancing Vaccinology. Nat. Rev. Immunol. 2019, 19, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Atmar, R.L.; Lyke, K.E.; Deming, M.E.; Jackson, L.A.; Branche, A.R.; El Sahly, H.M.; Rostad, C.A.; Martin, J.M.; Johnston, C.; Rupp, R.E.; et al. Homologous and Heterologous COVID-19 Booster Vaccinations. New Engl. J. Med. 2022, 386, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- González, S.; Olszevicki, S.; Gaiano, A.; Salazar, M.; Regairaz, L.; Varela Baino, A.N.; Bartel, E.; Varela, T.; González Martínez, V.V.; Pesci, S.; et al. Protection of Homologous and Heterologous Boosters after Primary Schemes of RAd26-RAd5, ChAdOx1 NCoV-19 and BBIBP-CorV during the Omicron Outbreak in Adults of 50 Years and Older in Argentina: A Test-Negative Case–Control Study. Lancet Reg. Health 2023, 27, 1000607. [Google Scholar] [CrossRef] [PubMed]
- Parks, C.L. Replication-Competent Viral Vectors for Vaccine Delivery. In Human Vaccines: Emerging Technologies in Design and Development; Elsevier: Amsterdam, The Netherlands, 2017; pp. 25–63. [Google Scholar] [CrossRef]
- Gu, J.; Xu, Z.; Liu, Q.; Tang, S.; Zhang, W.; Xie, S.; Chen, X.; Chen, J.; Yong, K.T.; Yang, C.; et al. Building a Better Silver Bullet: Current Status and Perspectives of Non-Viral Vectors for MRNA Vaccines. Adv. Healthc. Mater. 2024, 13, 2302409. [Google Scholar] [CrossRef]
- Scheaffer, S.M.; Lee, D.; Whitener, B.; Ying, B.; Wu, K.; Liang, C.Y.; Jani, H.; Martin, P.; Amato, N.J.; Avena, L.E.; et al. Bivalent SARS-CoV-2 MRNA Vaccines Increase Breadth of Neutralization and Protect against the BA.5 Omicron Variant in Mice. Nat. Med. 2022, 29, 247–257. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. New Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Momin, T.; Kansagra, K.; Patel, H.; Sharma, S.; Sharma, B.; Patel, J.; Mittal, R.; Sanmukhani, J.; Maithal, K.; Dey, A.; et al. Safety and Immunogenicity of a DNA SARS-CoV-2 Vaccine (ZyCoV-D): Results of an Open-Label, Non-Randomized Phase I Part of Phase I/II Clinical Study by Intradermal Route in Healthy Subjects in India. eClinicalMedicine 2021, 38, 101020. [Google Scholar] [CrossRef]
- Hanke, T. New Vector and Vaccine Platforms: MRNA, DNA, Viral Vectors. Curr. Opin. HIV AIDS 2022, 17, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Langer, B.; Renner, M.; Scherer, J.; Schüle, S.; Cichutek, K. Safety Assessment of Biolistic DNA Vaccination. Methods Mol. Biol. 2013, 940, 371–388. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Arun Kumar, S.; Jhan, Y.Y.; Bishop, C.J. Engineering DNA Vaccines against Infectious Diseases. Acta Biomater. 2018, 80, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Eusébio, D.; Neves, A.R.; Costa, D.; Biswas, S.; Alves, G.; Cui, Z.; Sousa, Â. Methods to Improve the Immunogenicity of Plasmid DNA Vaccines. Drug Discov. Today 2021, 26, 2575–2592. [Google Scholar] [CrossRef] [PubMed]
- Elizaga, M.L.; Li, S.S.; Kochar, N.K.; Wilson, G.J.; Allen, M.A.; Tieu, H.V.N.; Frank, I.; Sobieszczyk, M.E.; Cohen, K.W.; Sanchez, B.; et al. Safety and Tolerability of HIV-1 Multiantigen PDNA Vaccine given with IL-12 Plasmid DNA via Electroporation, Boosted with a Recombinant Vesicular Stomatitis Virus HIV Gag Vaccine in Healthy Volunteers in a Randomized, Controlled Clinical Trial. PLoS ONE 2018, 13, e0202753. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, S.P.; Grunenberg, N.A.; Sanchez, B.J.; Seaton, K.E.; Ferrari, G.; Moody, M.A.; Frahm, N.; Montefiori, D.C.; Hay, C.M.; Goepfert, P.A.; et al. Immunogenicity of a Novel Clade B HIV-1 Vaccine Combination: Results of Phase 1 Randomized Placebo Controlled Trial of an HIV-1 GM-CSF-Expressing DNA Prime with a Modified Vaccinia Ankara Vaccine Boost in Healthy HIV-1 Uninfected Adults. PLoS ONE 2017, 12, e0179597. [Google Scholar] [CrossRef] [PubMed]
- Klinman, D.M.; Klaschik, S.; Tross, D.; Shirota, H.; Steinhagen, F. FDA Guidance on Prophylactic DNA Vaccines: Analysis and Recommendations. Vaccine 2010, 28, 2801–2805. [Google Scholar] [CrossRef] [PubMed]
- Jorritsma, S.H.T.; Gowans, E.J.; Grubor-Bauk, B.; Wijesundara, D.K. Delivery Methods to Increase Cellular Uptake and Immunogenicity of DNA Vaccines. Vaccine 2016, 34, 5488–5494. [Google Scholar] [CrossRef]
- Qin, F.; Xia, F.; Chen, H.; Cui, B.; Feng, Y.; Zhang, P.; Chen, J.; Luo, M. A Guide to Nucleic Acid Vaccines in the Prevention and Treatment of Infectious Diseases and Cancers: From Basic Principles to Current Applications. Front. Cell Dev. Biol. 2021, 9, 633776. [Google Scholar] [CrossRef]
- Satkauskas, S.; Bureau, M.F.; Puc, M.; Mahfoudi, A.; Scherman, D.; Miklavcic, D.; Mir, L.M. Mechanisms of In Vivo DNA Electrotransfer: Respective Contributions of Cell Electropermeabilization and DNA Electrophoresis. Mol. Ther. 2002, 5, 133–140. [Google Scholar] [CrossRef]
- Pilkington, E.H.; Suys, E.J.A.; Trevaskis, N.L.; Wheatley, A.K.; Zukancic, D.; Algarni, A.; Al-Wassiti, H.; Davis, T.P.; Pouton, C.W.; Kent, S.J.; et al. From Influenza to COVID-19: Lipid Nanoparticle MRNA Vaccines at the Frontiers of Infectious Diseases. Acta Biomater. 2021, 131, 16–40. [Google Scholar] [CrossRef]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine Into MRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Leal, L.; Guardo, A.C.; Morón-López, S.; Salgado, M.; Mothe, B.; Heirman, C.; Pannus, P.; Vanham, G.; van den Ham, H.J.; Gruters, R.; et al. Phase I Clinical Trial of an Intranodally Administered MRNA-Based Therapeutic Vaccine against HIV-1 Infection. AIDS 2018, 32, 2533–2545. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.D.; Moody, M.A.; Thompson, A.B. Innovations in HIV-1 Vaccine Design. Clin. Ther. 2020, 42, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. How COVID Unlocked the Power of RNA Vaccines. Nature 2021, 589, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Prudden, H.; Tatoud, R.; Slack, C.; Shattock, R.; Anklesaria, P.; Bekker, L.G.; Buchbinder, S. Experimental Medicine for HIV Vaccine Research and Development. Vaccines 2023, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Crommelin, D.J.A.; Anchordoquy, T.J.; Volkin, D.B.; Jiskoot, W.; Mastrobattista, E. Addressing the Cold Reality of MRNA Vaccine Stability. J. Pharm. Sci. 2021, 110, 997–1001. [Google Scholar] [CrossRef]
- Uddin, M.N.; Roni, M.A. Challenges of Storage and Stability of MRNA-Based COVID-19 Vaccines. Vaccines 2021, 9, 1033. [Google Scholar] [CrossRef]
- Saied, A.R.A. Building Africa’s First MRNA Vaccine Facility. Lancet 2023, 402, 287–288. [Google Scholar] [CrossRef]
- Fuenmayor, J.; Gòdia, F.; Cervera, L. Production of Virus-like Particles for Vaccines. N. Biotechnol. 2017, 39, 174–180. [Google Scholar] [CrossRef]
- Thalhauser, S.; Peterhoff, D.; Wagner, R.; Breunig, M. Critical Design Criteria for Engineering a Nanoparticulate HIV-1 Vaccine. J. Control. Release 2020, 317, 322–335. [Google Scholar] [CrossRef]
- Tokatlian, T.; Kulp, D.W.; Mutafyan, A.A.; Jones, C.A.; Menis, S.; Georgeson, E.; Kubitz, M.; Zhang, M.H.; Melo, M.B.; Silva, M.; et al. Enhancing Humoral Responses Against HIV Envelope Trimers via Nanoparticle Delivery with Stabilized Synthetic Liposomes. Sci. Rep. 2018, 8, 16527. [Google Scholar] [CrossRef]
- Cervera, L.; Gòdia, F.; Tarrés-Freixas, F.; Aguilar-Gurrieri, C.; Carrillo, J.; Blanco, J.; Gutiérrez-Granados, S. Production of HIV-1-Based Virus-like Particles for Vaccination: Achievements and Limits. Appl. Microbiol. Biotechnol. 2019, 103, 7367–7384. [Google Scholar] [CrossRef] [PubMed]
- Paliard, X.; Liu, Y.; Wagner, R.; Wolf, H.; Baenziger, J.; Walker, C.M. Priming of Strong, Broad, and Long-Lived HIV Type 1 P55gag-Specific CD8+ Cytotoxic T Cells after Administration of a Virus-Like Particle Vaccine in Rhesus Macaques. AIDS Res. Hum. Retroviruses 2000, 16, 273–282. [Google Scholar] [CrossRef]
- Deml, L.; Speth, C.; Dierich, M.P.; Wolf, H.; Wagner, R. Recombinant HIV-1 Pr55gag Virus-like Particles: Potent Stimulators of Innate and Acquired Immune Responses. Mol. Immunol. 2005, 42, 259–277. [Google Scholar] [CrossRef]
- Veenstra, J.; Williams, I.G.; Colebunders, R.; Dorrell, L.; Tchamouroff, S.E.; Patou, G.; Lange, J.M.A.; Weller, I.V.D.; Goeman, J.; Uthayakumar, S.; et al. Immunization with Recombinant P17/P24:Ty Virus-like Particles in Human Immunodeficiency Virus-Infected Persons. J. Infect. Dis. 1996, 174, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.S.; Cheingsong-Popov, R.; Callow, D.; Foxall, R.; Patou, G.; Hodgkin, K.; Weber, J.N. A Pilot Phase II Study of the Safety and Immunogenicity of HIV P17/P24:VLP (P24-VLP) in Asymptomatic HIV Seropositive Subjects. J. Infect. 1997, 35, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, A.D.; Roggensack, M.; Jaramillo, A.B.; Smith, D.E.; Walker, A.; Gow, I.; McMurchie, M.; Harris, J.; Patou, G.; Cooper, D.A. Safety and Immunogenicity of a Candidate Therapeutic Vaccine, P24 Virus-like Particle, Combined with Zidovudine, in Asymptomatic Subjects. Community HIV Research Network Investigators. AIDS 1998, 12, 175–182. [Google Scholar] [CrossRef]
- Chen, C.W.; Saubi, N.; Joseph-Munné, J. Design Concepts of Virus-Like Particle-Based HIV-1 Vaccines. Front. Immunol. 2020, 11, 573157. [Google Scholar] [CrossRef]
- Klein, J.S.; Bjorkman, P.J. Few and Far between: How HIV May Be Evading Antibody Avidity. PLoS Pathog. 2010, 6, e1000908. [Google Scholar] [CrossRef]
- Deml, L.; Kratochwil, G.; Osterrieder, N.; Knüchel, R.; Wolf, H.; Wagner, R. Increased Incorporation of Chimeric Human Immunodeficiency Virus Type 1 Gp120 Proteins into Pr55gag Virus-Like Particles by an Epstein-Barr Virus Gp220/350-Derived Transmembrane Domain. Virology 1997, 235, 10–25. [Google Scholar] [CrossRef]
- Wang, B.-Z.; Liu, W.; Kang, S.-M.; Alam, M.; Huang, C.; Ye, L.; Sun, Y.; Li, Y.; Kothe, D.L.; Pushko, P.; et al. Incorporation of High Levels of Chimeric Human Immunodeficiency Virus Envelope Glycoproteins into Virus-like Particles. J. Virol. 2007, 81, 10869–10878. [Google Scholar] [CrossRef] [PubMed]
- Stano, A.; Leaman, D.P.; Kim, A.S.; Zhang, L.; Autin, L.; Ingale, J.; Gift, S.K.; Truong, J.; Wyatt, R.T.; Olson, A.J.; et al. Dense Array of Spikes on HIV-1 Virion Particles. J. Virol. 2017, 91, e00415-17. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.; van Diepen, M.; Galant, S.; Kruse, E.; Margolin, E.; Ximba, P.; Hermanus, T.; Moore, P.; Douglass, N.; Williamson, A.-L.; et al. Immunogenicity of HIV-1 Vaccines Expressing Chimeric Envelope Glycoproteins on the Surface of Pr55 Gag Virus-Like Particles. Vaccines 2020, 8, 54. [Google Scholar] [CrossRef]
- Escolano, A.; Gristick, H.B.; Abernathy, M.E.; Merkenschlager, J.; Gautam, R.; Oliveira, T.Y.; Pai, J.; West, A.P.J.; Barnes, C.O.; Cohen, A.A.; et al. Immunization Expands B Cells Specific to HIV-1 V3 Glycan in Mice and Macaques. Nature 2019, 570, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Tarrés-Freixas, F.; Aguilar-Gurrieri, C.; Rodríguez de la Concepción, M.L.; Urrea, V.; Trinité, B.; Ortiz, R.; Pradenas, E.; Blanco, P.; Marfil, S.; Molinos-Albert, L.M.; et al. An Engineered HIV-1 Gag-Based VLP Displaying High Antigen Density Induces Strong Antibody-Dependent Functional Immune Responses. NPJ Vaccines 2023, 8, 51. [Google Scholar] [CrossRef]
- Ortiz, R.; Barajas, A.; Pons-Grífols, A.; Trinité, B.; Tarrés-Freixas, F.; Rovirosa, C.; Urrea, V.; Barreiro, A.; Gonzalez-Tendero, A.; Cardona, M.; et al. Exploring FeLV-Gag-Based VLPs as a New Vaccine Platform-Analysis of Production and Immunogenicity. Int. J. Mol. Sci. 2023, 24, 9025. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, H.; Nabi, G.; McKinstry, W.J.; Khoo, K.K.; Mak, J.; Salazar, A.M.; Tenbusch, M.; Temchura, V.; Überla, K. Intrastructural Help: Harnessing T Helper Cells Induced by Licensed Vaccines for Improvement of HIV Env Antibody Responses to Virus-Like Particle Vaccines. J. Virol. 2018, 92, e00141-18. [Google Scholar] [CrossRef] [PubMed]
- Klessing, S.; Temchura, V.; Tannig, P.; Peter, A.S.; Christensen, D.; Lang, R.; Überla, K. CD4(+) T Cells Induced by Tuberculosis Subunit Vaccine H1 Can Improve the HIV-1 Env Humoral Response by Intrastructural Help. Vaccines 2020, 8, 604. [Google Scholar] [CrossRef]
- Smith, J.M.; Rao Amara, R.; Campbell, D.; Xu, Y.; Patel, M.; Sharma, S.; Butera, S.T.; Ellenberger, D.L.; Yi, H.; Chennareddi, L.; et al. DNA/MVA Vaccine for HIV Type 1: Effects of Codon-Optimization and the Expression of Aggregates or Virus-Like Particles on the Immunogenicity of the DNA Prime. AIDS Res. Hum. Retroviruses 2004, 20, 1335–1347. [Google Scholar] [CrossRef]
- Ellenberger, D.; Wyatt, L.; Li, B.; Buge, S.; Lanier, N.; Rodriguez, I.V.; Sariol, C.A.; Martinez, M.; Monsour, M.; Vogt, J.; et al. Comparative Immunogenicity in Rhesus Monkeys of Multi-Protein HIV-1 (CRF02_AG) DNA/MVA Vaccines Expressing Mature and Immature VLPs. Virology 2005, 340, 21–32. [Google Scholar] [CrossRef]
- Perdiguero, B.; Sánchez-Corzo, C.; Sorzano, C.O.S.; Saiz, L.; Mediavilla, P.; Esteban, M.; Gómez, C.E. A Novel MVA-Based HIV Vaccine Candidate (MVA-Gp145-GPN) Co-Expressing Clade C Membrane-Bound Trimeric Gp145 Env and Gag-Induced Virus-Like Particles (VLPs) Triggered Broad and Multifunctional HIV-1-Specific T Cell and Antibody Responses. Viruses 2019, 11, 160. [Google Scholar] [CrossRef]
- Montefiori, D.C.; Safrit, J.T.; Lydy, S.L.; Barry, A.P.; Miroslawa, B.; Vo, H.T.; Klein, M.; Tartaglia, J.; Robinson, H.L.; Rovinski, B. Induction of Neutralizing Antibodies and Gag-Specific Cellular Immune Responses to an R5 Primary Isolate of Human Immunodeficiency Virus Type 1 in Rhesus Macaques. J. Virol. 2001, 75, 5879–5890. [Google Scholar] [CrossRef] [PubMed]
- Radaelli, A.; Zanotto, C.; Perletti, G.; Elli, V.; Vicenzi, E.; Poli, G.; De Giuli Morghen, C. Comparative Analysis of Immune Responses and Cytokine Profiles Elicited in Rabbits by the Combined Use of Recombinant Fowlpox Viruses, Plasmids and Virus-like Particles in Prime-Boost Vaccination Protocols against SHIV. Vaccine 2003, 21, 2052–2064. [Google Scholar] [CrossRef]
- Radaelli, A.; Bonduelle, O.; Beggio, P.; Mahe, B.; Pozzi, E.; Elli, V.; Paganini, M.; Zanotto, C.; Morghen, C.D.G.; Combadière, B. Prime-Boost Immunization with DNA, Recombinant Fowlpox Virus and VLPSHIV Elicit Both Neutralizing Antibodies and IFNγ-Producing T Cells against the HIV-Envelope Protein in Mice That Control Env-Bearing Tumour Cells. Vaccine 2007, 25, 2128–2138. [Google Scholar] [CrossRef] [PubMed]
- Buonaguro, L.; Devito, C.; Tornesello, M.L.; Schröder, U.; Wahren, B.; Hinkula, J.; Buonaguro, F.M. DNA-VLP Prime-Boost Intra-Nasal Immunization Induces Cellular and Humoral Anti-HIV-1 Systemic and Mucosal Immunity with Cross-Clade Neutralizing Activity. Vaccine 2007, 25, 5968–5977. [Google Scholar] [CrossRef] [PubMed]
- Lifei, Y.; Yufeng, S.; Xiaomin, L.; Xiaoxing, H.; Jingjing, L.; Heng, D.; Ping, Z.; Paul, Z. HIV-1 Virus-Like Particles Produced by Stably Transfected Drosophila S2 Cells: A Desirable Vaccine Component. J. Virol. 2012, 86, 7662–7676. [Google Scholar] [CrossRef]
- Kamdem Toukam, D.; Tenbusch, M.; Stang, A.; Temchura, V.; Storcksdieck genannt Bonsmann, M.; Grewe, B.; Koch, S.; Meyerhans, A.; Nchinda, G.; Kaptue, L.; et al. Targeting Antibody Responses to the Membrane Proximal External Region of the Envelope Glycoprotein of Human Immunodeficiency Virus. PLoS ONE 2012, 7, e38068. [Google Scholar] [CrossRef] [PubMed]
- Nabi, G.; Genannt Bonsmann, M.S.; Tenbusch, M.; Gardt, O.; Barouch, D.H.; Temchura, V.; Überla, K. GagPol-Specific CD4+T-Cells Increase the Antibody Response to Env by Intrastructural Help. Retrovirology 2013, 10, 117. [Google Scholar] [CrossRef]
- Ross, T.M.; Pereira, L.E.; Luckay, A.; McNicholl, J.M.; García-Lerma, J.G.; Heneine, W.; Eugene, H.S.; Pierce-Paul, B.R.; Zhang, J.; Hendry, R.M.; et al. A Polyvalent Clade B Virus-Like Particle HIV Vaccine Combined with Partially Protective Oral Preexposure Prophylaxis Prevents Simian–Human Immunodeficiency Virus Infection in Macaques and Primes for Virus-Amplified Immunity. AIDS Res. Hum. Retroviruses 2014, 30, 1072–1081. [Google Scholar] [CrossRef]
- Benen, T.D.; Tonks, P.; Kliche, A.; Kapzan, R.; Heeney, J.L.; Wagner, R. Development and Immunological Assessment of VLP-Based Immunogens Exposing the Membrane-Proximal Region of the HIV-1 Gp41 Protein. J. Biomed. Sci. 2014, 21, 79. [Google Scholar] [CrossRef]
- Vzorov, A.N.; Wang, L.; Chen, J.; Wang, B.-Z.; Compans, R.W. Effects of Modification of the HIV-1 Env Cytoplasmic Tail on Immunogenicity of VLP Vaccines. Virology 2016, 489, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Gangadhara, S.; Victor, B.; Shen, X.; Chen, X.; Nabi, R.; Kasturi, S.P.; Sabula, M.J.; Labranche, C.C.; Reddy, P.B.J.; et al. Virus-Like Particles Displaying Trimeric Simian Immunodeficiency Virus (SIV) Envelope Gp160 Enhance the Breadth of DNA/Modified Vaccinia Virus Ankara SIV Vaccine-Induced Antibody Responses in Rhesus Macaques. J. Virol. 2016, 90, 8842–8854. [Google Scholar] [CrossRef] [PubMed]
- Storcksdieck genannt Bonsmann, M.; Niezold, T.; Hannaman, D.; Überla, K.; Tenbusch, M. The Improved Antibody Response against HIV-1 after a Vaccination Based on Intrastructural Help Is Complemented by Functional CD8+ T Cell Responses. Vaccine 2016, 34, 1744–1751. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhu, Q.; Huang, X.; Yang, L.; Song, Y.; Zhu, P.; Zhou, P. In Vivo Electroporation in DNA-VLP Prime-Boost Preferentially Enhances HIV-1 Envelope-Specific IgG2a, Neutralizing Antibody and CD8 T Cell Responses. Vaccine 2017, 35, 2042–2051. [Google Scholar] [CrossRef] [PubMed]
- Beltran-Pavez, C.; Ferreira, C.B.; Merino-Mansilla, A.; Fabra-Garcia, A.; Casadella, M.; Noguera-Julian, M.; Paredes, R.; Olvera, A.; Haro, I.; Brander, C.; et al. Guiding the Humoral Response against HIV-1 toward a MPER Adjacent Region by Immunization with a VLP-Formulated Antibody-Selected Envelope Variant. PLoS ONE 2018, 13, e0208345. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Dienger-Stambaugh, K.; Chen, X.; Wei, H.; Phan, S.; Beavis, A.C.; Singh, K.; Adhikary, N.R.D.; Tiwari, P.; Villinger, F.; et al. Parainfluenza Virus 5 Priming Followed by SIV/HIV Virus-Like-Particle Boosting Induces Potent and Durable Immune Responses in Nonhuman Primates. Front. Immunol. 2021, 12, 162. [Google Scholar] [CrossRef]
- Zhang, P.; Narayanan, E.; Liu, Q.; Tsybovsky, Y.; Boswell, K.; Ding, S.; Hu, Z.; Follmann, D.; Lin, Y.; Miao, H.; et al. A Multiclade Env–Gag VLP MRNA Vaccine Elicits Tier-2 HIV-1-Neutralizing Antibodies and Reduces the Risk of Heterologous SHIV Infection in Macaques. Nat. Med. 2021, 27, 2234–2245. [Google Scholar] [CrossRef]
- Beltran-Pavez, C.; Bontjer, I.; Gonzalez, N.; Pernas, M.; Merino-Mansilla, A.; Olvera, A.; Miro, J.M.; Brander, C.; Alcami, J.; Sanders, R.W.; et al. Potent Induction of Envelope-Specific Antibody Responses by Virus-Like Particle Immunogens Based on HIV-1 Envelopes from Patients with Early Broadly Neutralizing Responses. J. Virol. 2022, 96, e0134321. [Google Scholar] [CrossRef]
- Vazquez, T.; Torrieri-Damard, L.; Pitoiset, F.; Levacher, B.; Vigneron, J.; Mayr, L.; Brimaud, F.; Bonnet, B.; Moog, C.; Klatzmann, D.; et al. Particulate Antigens Administrated by Intranasal and Intravaginal Routes in a Prime-Boost Strategy Improve HIV-Specific TFH Generation, High-Quality Antibodies and Long-Lasting Mucosal Immunity. Eur. J. Pharm. Biopharm. 2023, 191, 124–138. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Code | Phase | Immunogen | Formulation | Main Finding | Study Group | Status | Year | Location | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DNA | Poxvirus | AVs | Protein | Adjuvant | |||||||||
| Vaccines that aimed at eliciting neutralising humoral responses | |||||||||||||
| Vax 003 | NCT00006327 | III | Bivalent Clade B&E gp120 | - | - | - | AIDSVAX B/E | Alum | No protection | IDU | Complete | 2003 | Thai |
| Vax 004 | NCT00002441 | III | Bivalent Clade B&B gp120 | - | - | - | AIDSVAX B/B | Alum | No protection. Low Tier1 nAbs | MSM and high risk women | Complete | 2004 | USA Netherlands |
| Vaccines that aimed at inducing protective cellular responses | |||||||||||||
| STEP HVTN502 | NCT00095576 | IIb | Clade B gag, pol & nef | - | - | Ad5 | - | - | Anti-vector Abs may increase risk of infection | High risk population | Stopped | 2007 | America Australia |
| Phambil HVTN503 | NCT00413725 | IIb | Clade B gag, pol & nef | - | - | Ad5 | - | - | South-African population | Stopped | 2007 | South-Africa | |
| Vaccines that aimed at generating balanced adaptive immune responses | |||||||||||||
| RV144 | NCT00223080 | III | Clade B gag-pro, env AE Clade B&E gp120 | - | ALVAC | - | AIDSVAX B/E | Alum | 31.2% protection | Thai population | Complete | 2009 | Thai |
| HVTN505 | NCT00865566 | IIb | Clade B gag, pol & nef Clades A, B, C env | 6 p. | - | rAd5 | - | - | No protection. Anti-gp41 nnAbs | MSM and transgenders | Stopped | 2013 | USA |
| UHAMBO HVTN 702 | NCT02968849 | III | Clade B/C gag-pro & env Clade C gp120 | - | ALVAC | - | Cgp120 C | MF59 | No protection demonstrated | South-African population | Stopped | 2020 | South-Africa |
| IMBOKODO HVTN 705 | NCT03060629 | IIb | Mosaic gag, pol & env Clade C Env trimer | - | - | Ad26 | Clade C gp140 | Alum | No significant protection (25.2%) | Women at risk | Stopped | 2021 | Sub-saharan Africa |
| MOSAICO HVTN 706 | NCT03964415 | III | Mosaic gag, pol & env Clade C + Mosaic gp120 | - | - | Ad26 | TV1.Cgp120 1086.Cgp120 | Alum | No protection demonstrated | MSM and transgenders | Stopped | 2023 | Europe America |
| PrEPVacc | NCT04066881 | IIb | Clade C gag, pol, env, nef Env E Gag-pol Clade A Clade B&E/Clade C Env | 3 p. | MVA | - | CN54gp140 AIDSVAX B/E | MPLA | No protection demonstrated | High risk population | Stopped | 2023 | Uganda |
| Name | Immunogen | Combined Strategy | No. | Adjuvants | Delivery | RoA | Phase | Main Finding | Date | |
|---|---|---|---|---|---|---|---|---|---|---|
| DNA vaccines | ||||||||||
| HIVIS-DNA | Subtypes A, B, C env, gag & B rt/rev | MVA | + | TLR4 agonist | EP | ID | I | Ongoing | - | NCT04301154 |
| Env-C DNA | Subtype C env | Subunit protein | 1 | Alum/ALF43 | - | IM | I | Ongoing | - | NCT04826094 |
| PDPHV201401 | Subtype A, B, C, A/E env & C gag | Subunit protein | 5 | GLA-SE | - | IM | I | Ongoing | - | NCT04927585 |
| DNA-HIV-PT123 | Subtype C env, gag & B/C pol-nef | Subunit/MVA | 3 | Alum | - | IM | IIb | Safe, stopped (no efficacy) | 2023 | NCT04066881 |
| Env/Gag DNA vaccine | Subtypes A, B, C, A/E env & C gag | Subunit protein | 5 | - | - | IM | I | Safe, broad and potent IR | 2021 | NCT03409276 |
| p24CE1/2 & p55_DNA | Conserved elements of p24 & full p55 | - | 2 | IL12 | EP (Ichor) | IM | I | Safe, EP induced higher IR | 2020 | NCT03181789 |
| DNA Nat-B/ Con-S/mosaic | Clade B, Consensus M and Mosaic env | MVA | 3 | - | - | IM | I | N.Y.R | 2020 | NCT02296541 |
| nef/tat/vif, env pDNA | N.D. | MVA | 1 | - | EP | I | Completed | 2019 | NCT02654080 | |
| HIV DNA-C CN54ENV | Subtype C env | Subunit protein | 1 | - | EP (Ichor) | IM/ID | I | - | 2019 | NCT02654080 |
| Pennvax-GP Pennvax-B | Cons. A&C env | - | 1 | IL12 | EP (Cellectra) | ID/IM | I | Robust Ab and cellular IR | 2018 | NCT02431767 |
| GEO-D03 DNA | Subtype B env, gag, pol | MVA | 1 | GM-CSF | - | IM | I | Safe, good for priming | 2017 | NCT01571960 |
| pSG2.HIVconsv | HIV.consv construct | ChAdV or MVA | 1 | - | - | IM | I | Safe, high CTL | 2017 | NCT01151319 |
| MAG-pDNA | Subtype B env, gag, pol, nef, tat & vif | rVSV | 2 | IL-12 | EP (Ichor) | IM | I | Safe, primes CTL | 2016 | NCT01578889 |
| mRNA vaccines | ||||||||||
| V3G CH848 mRNA-Tr2 | Native-like env trimer | Nanoparticle | 1 | Alum | Liposome | IM | I | Recruiting | - | NCT05903339 |
| BG505 trimer | Native-like env trimer | - | 1 | Liposome | IM | I | Ongoing | - | NCT05217641 | |
| mRN1644 | eOD-GT8 & Core-g28v2 | mRNA | 1 | - | Liposome | IM | I | Ongoing | - | NCT05001373 |
| Surface Immunogen | Adjuvant | Aim | Production Platform | Model | Immunisation Regimen | RoA | Major Findings | Ref. |
|---|---|---|---|---|---|---|---|---|
| Nucleic acid HIV-1 Gag VLPs | ||||||||
| Subtype B Env | - | Test DNA/MVA vaccine forming VLPs | In vivo | Macaques | Heterologous (DMM) | IM | Superior Ab response in VLP constructs | [154] |
| CRF02_AG Env | - | Test DNA/MVA CRF02_AG in mVLPs vs VLPs | In vivo | Macaques | Heterologous (DDDM) | IM | Better cellular and humoral response in mVLP | [155] |
| Clade B Env | GM-CSF | Safety & immunogenicity | In vivo | Humans | MVA | IM | Safe, good for priming | [120] |
| Clade C gp145 | - | Compare combine Gag-Pol-Env vs single MVAs | In vivo | BALB/c | MVA | IM | Superior Ab & CTL in the combined MVA | [156] |
| Subtype C gp160 | - | Increase Env density at VLP surface | In vivo | Rabbit | Heterologous (DDMMPP) | IM | No high-density Env in VLPs, Tier 2 autologus nAb | [148] |
| Combined Nucleic acid VLPs and Gag-VLPs | ||||||||
| Subtype B Env | QS21 | Assess immunogenicity | Vero cells In vivo | Macaques | Heterologous (D/V/A) | IM | Autologous nAbs and CTL | [157] |
| Subtype B Env | - | Evaluate VLP as a boost for D/Fpox | Vero cells In vivo | Rabbits | Heterologous (D/Fpox/V) | ID | VLP boosting elicits potent Ab | [158] |
| Subtype B Env | - | Compare heterologous immunisation | Vero cells In vivo | BALB/c mice | Heterologous (3xD/Fpox + VV) | IM/SC | nAbs & CTLs control env-tumour cells | [159] |
| Clade A Gp120 | Eurocine | Compare mucosal immunisation | Insect cells In vivo | BALB/c mice | Homologous (VV) Heterologous (DV) | IN | Superior Ab & CTL with heterologous vaccination | [160] |
| Clade B gp160 and Gp120 | - | Check S2 cell VLP platform | S2 cells In vivo | BALB/c | Heterologous (DDVVV) | SC | Antibody induction and modest neutralisation. | [161] |
| Gp41 variants | - | Induce anti-MPER bNAbs | HEK293T cells In vivo | BALB/c | Heterologous (DDDVVV) | SC | Trimeric MPER increases Ab responses | [162] |
| Clade B gp140 | - | Intrastructural help testing | HEK293T cells In vivo | C57BL/6J | Heterologous (Ad or DNA + VLP) | Footpad | Gag boosts Env via intrastructural help | [163] |
| Clade B Env | Alum or CpG | To test mixing PrEP and vaccine | COS cells In vivo | Rhesus macaque | Heterologous (DDVVV) + PrEP | IM/IN | Combining PrEP + vaccine confers higher protection | [164] |
| Truncated gp41 | Carbopol974 | Induce anti-MPER bNAbs | HEK293F cells In vivo | Rabbit (NZW) | Heterologous (DDVV) | IM | Anti-MPER Ab and low nAbs by gp41-DDVV | [165] |
| Clade B and C Env | - | Test CT on VLP immunogenicity | Insect cells In vivo | Guinea pig | Heterologous (DDVV) | IM | Modified Env CT enhances VLP immunogenicity | [166] |
| SIV239 Env | CD40L, MPLA, R848 | Induce protective IR in rhesus | - | Rhesus macaque | Heterologous (DDMMV) | Hock | Partial protection in DDMMV group | [167] |
| Subtype B gp140 | IL12, IL28 | Induce balanced response by ISH | 293T cells In vivo | BALB/c mice | Homologous (DD) Heterologous (DVV) | IM | Higher Ab & CTL by heterologous ISH-based vaccine | [168] |
| Subtype B gp120 | c-di-GMP | Test in vivo EP in an heterologous regimen | Insect cells In vivo | BALB/c mice | Heterologous (DDVV) | SC | Th1-like response in DDVV w/nAb & CTL | [169] |
| Clade B Env | Adjuplex | Induce bNAbs | HEK293F cells In vivo | Rabbit | Heterologous (DDVVV) | - | Enhanced Ab response by 4E10-selected VLPs | [170] |
| Subtype B Env | R848 | Test PIV vector as a prime | FreeStyle 293 In vivo | Macaques | Heterologous (PIV/VLP) | IN/IM | Potent IR, previous anti-PIV exposition no major effect | [171] |
| Subtype B Env | Adjuplex (prot) | mRNA delivery of Gag VLPs expressing Env | HEK293F cells In vivo | Mice & macaques | Heterologous (mRNA, clades or proteins) | IM | 79% cross-clade risk reduction in macaques | [172] |
| Subtype B SOSIP | Adjuplex | Compare DDPPP vs DDVVV w/ SOSIP Env | FreeStyle 293 In vivo | Rabbits | Heterologous (DDVVV) | - | Strong Ab response DDPPP and DDVVV (lower dose) | [173] |
| Truncated gp41 | - | Genereate high-density VLPs to induce strong Abs | In vivo Expi293 cells | C57BL/6J | Heterologous (DDVV) | SC | ADCC responses & control gp41-tumour cells | [150] |
| Clade B gp140 | - | To induce mucosal IR w/MLV-Gag VLP | In vivo HEK293T cells | BALB/c | Heterologous (DV or DP) | Intravag. | Induction of systemic and mucosal immunity | [174] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarrés-Freixas, F.; Clotet, B.; Carrillo, J.; Blanco, J. Nucleic Acid Vaccines Encoding Proteins and Virus-like Particles for HIV Prevention. Vaccines 2024, 12, 298. https://doi.org/10.3390/vaccines12030298
Tarrés-Freixas F, Clotet B, Carrillo J, Blanco J. Nucleic Acid Vaccines Encoding Proteins and Virus-like Particles for HIV Prevention. Vaccines. 2024; 12(3):298. https://doi.org/10.3390/vaccines12030298
Chicago/Turabian StyleTarrés-Freixas, Ferran, Bonaventura Clotet, Jorge Carrillo, and Julià Blanco. 2024. "Nucleic Acid Vaccines Encoding Proteins and Virus-like Particles for HIV Prevention" Vaccines 12, no. 3: 298. https://doi.org/10.3390/vaccines12030298
APA StyleTarrés-Freixas, F., Clotet, B., Carrillo, J., & Blanco, J. (2024). Nucleic Acid Vaccines Encoding Proteins and Virus-like Particles for HIV Prevention. Vaccines, 12(3), 298. https://doi.org/10.3390/vaccines12030298