Baseline Levels of Influenza-Specific B Cells and T Cell Responses Modulate Human Immune Responses to Swine Variant Influenza A/H3N2 Vaccine

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Viruses and Antigens
2.3. Peripheral Blood Mononuclear Cell Isolation
2.4. Serum Hemagglutination Inhibition (HAI) and Neutralization (Neut) Antibody Assays
2.5. Antibody-Secreting Cells (Plasmablasts) Assay
2.6. H3N2 Specific CD4+ T Cells by Intracellular Cytokine and CD154 Staining Assay
2.7. Memory B Cell (MBC) Assay
2.8. Statistical Methods
3. Results
3.1. Demographics
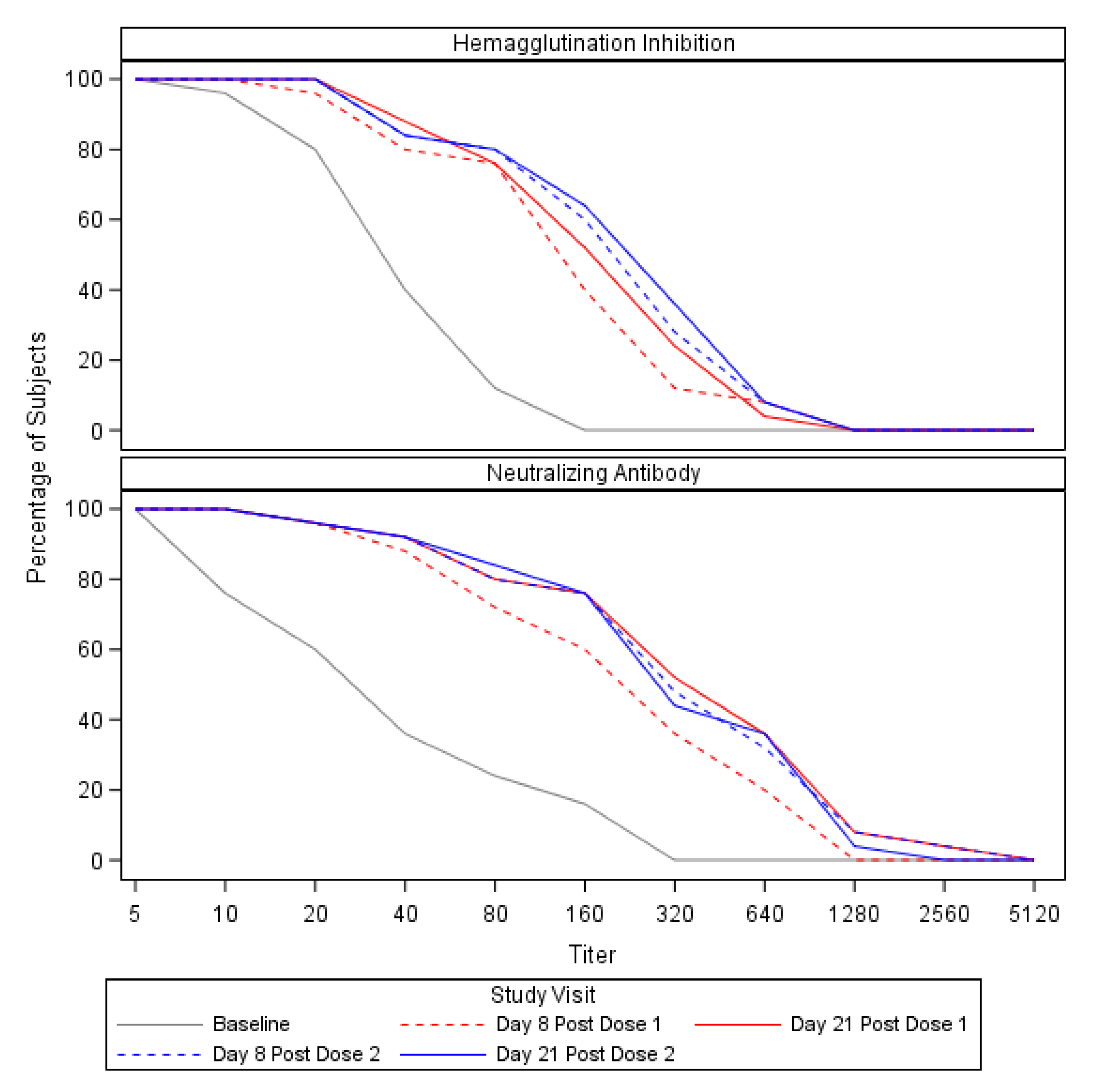
3.2. Preexisting Cross-Reactive Antibody and Expansion of the Antibody Response
3.3. Plasmablast Responses Correlated with HAI and Neut Titers and Were Detected against Seasonal H3N2 Antigens
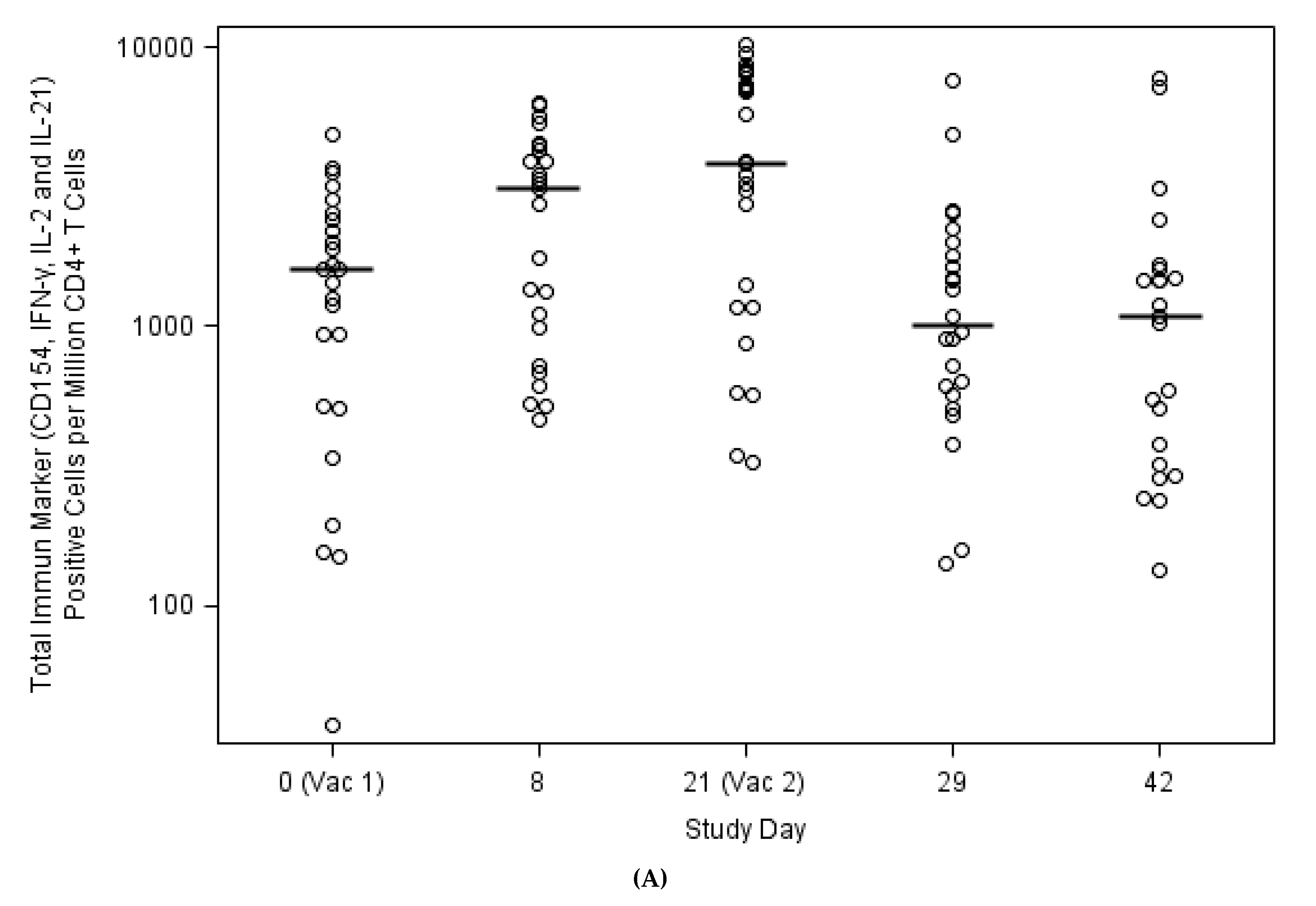
3.4. Expansion of Circulating Antiviral CD4+ T Cells against H3N2v Correlated with Antibody Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Case Count: Detected U.S. Human Infections with H3N2v by State since August 2011|Swine/Variant Influenza (Flu). Available online: https://www.cdc.gov/flu/swineflu/h3n2v-case-count.htm (accessed on 2 July 2019).
- Olsen, C.W.; Karasin, A.I.; Carman, S.; Li, Y.; Bastien, N.; Ojkic, D.; Alves, D.; Charbonneau, G.; Henning, B.M.; Low, D.E.; et al. Triple Reassortant H3N2 Influenza A Viruses, Canada, 2005. Emerg. Infect. Dis. 2006, 12, 1132–1135. [Google Scholar] [CrossRef]
- Blanton, L.; Wentworth, D.E.; Alabi, N.; Azziz-Baumgartner, E.; Barnes, J.; Brammer, L.; Burns, E.; Davis, C.T.; Dugan, V.G.; Fry, A.M.; et al. Update: Influenza Activity—United States and Worldwide, May 21–September 23, 2017. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 1043–1051. [Google Scholar] [CrossRef] [Green Version]
- Epperson, S.; Jhung, M.; Richards, S.; Quinlisk, P.; Ball, L.; Moll, M.; Boulton, R.; Haddy, L.; Biggerstaff, M.; Brammer, L.; et al. Human Infections with Influenza A(H3N2) Variant Virus in the United States, 2011–2012. Clin. Infect. Dis. 2013, 57, S4–S11. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.N.; Senne, D.A.; Landgraf, J.S.; Swenson, S.L.; Erickson, G.; Rossow, K.; Liu, L.; Yoon, K.; Krauss, S.; Webster, R.G. Genetic Reassortment of Avian, Swine, and Human Influenza A Viruses in American Pigs. J. Virol. 1999, 73, 8851. [Google Scholar] [CrossRef] [Green Version]
- Keitel, W.A.; Jackson, L.A.; Edupuganti, S.; Winokur, P.L.; Mulligan, M.J.; Thornburg, N.J.; Patel, S.M.; Rouphael, N.G.; Lai, L.; Bangaru, S.; et al. Safety and Immunogenicity of a Subvirion Monovalent Unadjuvanted Inactivated Influenza A(H3N2) Variant Vaccine in Healthy Persons ≥ 18 Years Old. J. Infect. Dis. 2015, 212, 552–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Reagent Resource. Available online: https://www.internationalreagentresource.org/ (accessed on 11 March 2020).
- BEI Resources. Available online: http://www.beiresources.org (accessed on 21 July 2019).
- Rowe, T.; Abernathy, R.A.; Hu-Primmer, J.; Thompson, W.W.; Lu, X.; Lim, W.; Fukuda, K.; Cox, N.J.; Katz, J.M. Detection of Antibody to Avian Influenza A (H5N1) Virus in Human Serum by Using a Combination of Serologic Assays. J. Clin. Microbiol. 1999, 37, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotty, S.; Aubert, R.D.; Glidewell, J.; Ahmed, R. Tracking human antigen-specific memory B cells: A sensitive and generalized ELISPOT system. J. Immunol. Methods 2004, 286, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Spensieri, F.; Borgogni, E.; Zedda, L.; Bardelli, M.; Buricchi, F.; Volpini, G.; Fragapane, E.; Tavarini, S.; Finco, O.; Rappuoli, R.; et al. Human Circulating influenza-CD4+ ICOS1+IL-21+ T Cells Expand after Vaccination, Exert Helper Function, and Predict Antibody Responses. Proc. Natl. Acad. Sci. USA 2013, 110, 14330–14335. [Google Scholar] [CrossRef] [Green Version]
- Houser, K.V.; Katz, J.M.; Tumpey, T.M. Seasonal Trivalent Inactivated Influenza Vaccine Does Not Protect against Newly Emerging Variants of Influenza A (H3N2v) Virus in Ferrets. J. Virol. 2013, 87, 1261–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waalen, K.; Kilander, A.; Dudman, S.G.; Ramos-Ocao, R.; Hungnes, O. Age-dependent prevalence of antibodies cross-reactive to the influenza A(H3N2) variant virus in sera collected in Norway in 2011. Eurosurveillance 2012, 17, 20170. [Google Scholar] [PubMed]
- Munoz, F.M.; Anderson, E.J.; Bernstein, D.I.; Harrison, C.J.; Pahud, B.; Anderson, E.; Creech, C.B.; Berry, A.A.; Kotloff, K.L.; Walter, E.B.; et al. Safety and immunogenicity of unadjuvanted subvirion monovalent inactivated influenza H3N2 variant (H3N2v) vaccine in children and adolescents. Vaccine 2019, 37, 5161–5170. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Janjua, N.Z.; De Serres, G.; Purych, D.; Gilca, V.; Scheifele, D.W.; Dionne, M.; Sabaiduc, S.; Gardy, J.L.; Li, G.; et al. Cross-reactive and Vaccine-Induced Antibody to an Emerging Swine-Origin Variant of Influenza A Virus Subtype H3N2 (H3N2v). J. Infect. Dis. 2012, 206, 1852–1861. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; He, X.-S.; Holmes, T.H.; Dekker, C.L.; Kemble, G.W.; Arvin, A.M.; Greenberg, H.B. Influence of Prior Influenza Vaccination on Antibody and B-Cell Responses. PLoS ONE 2008, 3, e2975. [Google Scholar] [CrossRef] [Green Version]
- Sacadura-Leite, E.; Sousa-Uva, A.; Rebelo-de-Andrade, H. Antibody response to the influenza vaccine in healthcare workers. Vaccine 2012, 30, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Halliley, J.L.; Kyu, S.; Kobie, J.J.; Walsh, E.E.; Falsey, A.R.; Randall, T.D.; Treanor, J.; Feng, C.; Sanz, I.; Lee, F.E.-H. Peak frequencies of circulating human influenza-specific antibody secreting cells correlate with serum antibody response after immunization. Vaccine 2010, 28, 3582–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, R.J.; Brokstad, K.A.; Zuckerman, M.A.; Wood, J.M.; Haaheim, L.R.; Oxford, J.S. An early humoral immune response in peripheral blood following parenteral inactivated influenza vaccination. Vaccine 1994, 12, 993–999. [Google Scholar] [CrossRef]
- El-Madhun, A.S.; Cox, R.J.; Søreide, A.; Olofsson, J.; Haaheim, L.R. Systemic and Mucosal Immune Responses in Young Children and Adults after Parenteral Influenza Vaccination. J. Infect. Dis. 1998, 178, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; Jaimes, M.C.; Holmes, T.H.; Dekker, C.L.; Mahmood, K.; Kemble, G.W.; Arvin, A.M.; Greenberg, H.B. Comparison of the Influenza Virus-Specific Effector and Memory B-Cell Responses to Immunization of Children and Adults with Live Attenuated or Inactivated Influenza Virus Vaccines. J. Virol. 2007, 81, 215–228. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, T.M.; Li, C.K.F.; Chui, C.S.C.; Huang, A.K.Y.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef]
- He, X.-S.; Holmes, T.H.; Sasaki, S.; Jaimes, M.C.; Kemble, G.W.; Dekker, C.L.; Arvin, A.M.; Greenberg, H.B. Baseline Levels of Influenza-Specific CD4 Memory T-Cells Affect T-Cell Responses to Influenza Vaccines. PLoS ONE 2008, 3, e2574. [Google Scholar] [CrossRef]
- Nayak, J.L.; Fitzgerald, T.F.; Richards, K.A.; Yang, H.; Treanor, J.J.; Sant, A.J. CD4+ T-Cell Expansion Predicts Neutralizing Antibody Responses to Monovalent, Inactivated 2009 Pandemic Influenza A(H1N1) Virus Subtype H1N1 Vaccine. J. Infect. Dis. 2013, 207, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Bangaru, S.; Nieusma, T.; Kose, N.; Thornburg, N.J.; Finn, J.A.; Kaplan, B.S.; King, H.G.; Singh, V.; Lampley, R.M.; Sapparapu, G.; et al. Recognition of influenza H3N2 variant virus by human neutralizing antibodies. JCI Insight 2016, 1, e86673. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Mishina, M.; Chung, J.R.; Cole, K.S.; Nowalk, M.P.; Martin, J.M.; Spencer, S.; Flannery, B.; Zimmerman, R.K.; Sambhara, S. Cell-Mediated Immunity Against Antigenically Drifted Influenza A(H3N2) Viruses in Children During a Vaccine Mismatch Season. J. Infect. Dis. 2016, 214, 1030–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Values (Sample Size n = 25) |
|---|---|
| Age | |
| Mean (SD) | 38.4 (14.4) |
| Median [Min, Max] | 30.0 [22.0, 63.0] |
| Gender—n (%) | |
| Female | 11 (44) |
| Male | 14 (56) |
| Race—n (%) | |
| Asian | 4 (16) |
| Black/African American | 8 (32) |
| Multi-Racial | 1 (4) |
| White | 12 (48) |
| Prior IIV 1 Receipt—n (%) | |
| None | 5 (20) |
| 2012/13 IIV only | 1 (4) |
| 2011/12 IIV only | 7 (28) |
| Both 2011/12 + 2012/13 IIV | 11 (44) |
| Unknown prior IIV | 1 (4) |
| Antibody Titers | Measured Metric | Day 0 (Pre-Vaccination) | Day 8 (Post Vaccination 1) | Day 21 (Post Vaccination 1, Immediately Pre-Vaccination 2) | Day 29 (Day 8 Post Vaccination 2) | Day 42 (Day 21 Post Vaccination 2) |
|---|---|---|---|---|---|---|
| Hemagglutination Inhibition (HAI) | Geometric Mean Titer (95% CI) | 27.5 (19.8, 38.3) | 91.9 (59.1, 142.8) | 117.9 (79.8, 174.2) | 126.4 (81.7, 195.6) | 135.5 (86.2, 213.0) |
| Seroconversion 1—n (%) | NA | 13 (52.0) | 14 (56.0) | 13 (52.0) | 13 (52.0) | |
| Neutralizing Antibody (Neut) | Geometric Mean Titer (95% CI) | 28.5 (16.0, 50.7) | 178.0 (103.6, 306.0) | 285.5 (162.2, 502.5) | 276.9 (157.8, 486.1) | 258.3 (149.7, 445.6) |
| Seroconversion 1—n (%) | NA | 12 (48.0) | 13 (52.0) | 13 (52.0) | 13 (52.0) |
| Measured Metric | IgG | IgA | ||||
|---|---|---|---|---|---|---|
| Strains | H3N2v Minnesota/2010 | H3N2 Perth/2009 | H3N2 Victoria/2011 | H3N2v Minnesota/2010 | H3N2 Perth/2009 | H3N2 Victoria/2011 |
| Median (min, max) | 162 (0, 9477) | 9.0 (0, 1377) | 9.0 (0, 2187) | 30.0 (0, 509) | 0.0 (0, 126) | 0.0 (0, 99) |
| Geometric Mean (95% CI) | 204.7 (104.2, 402.2) | 58.0 (20.4,164.7) | 84.0 (32.1, 219.9) | 56.1 (30.8, 101.8) | 23.7 (12.9, 43.4) | 21.2 (9.1, 49.3) |
| Number of Subjects with a Positive Response (%) | 22 (88) | 14 (56) | 13 (52) | 20 (80) | 9 (36) | 7 (28) |
| Plasmablasts | Hemagglutination Inhibition Antibody (p-Value) | Neutralizing Antibody (p-Value) |
|---|---|---|
| IgG | 0.469 (0.018) | 0.417 (0.038) |
| IgA | 0.647 (<0.001) | 0.457 (0.021) |
| Measured Metrics | Hemagglutination Inhibition Titer (p-Value) | Neutralizing Antibody Titer (p-Value) | ||||||
|---|---|---|---|---|---|---|---|---|
| Days | Day 8 | Day 21 | Day 29 | Day 42 | Day 8 | Day 21 | Day 29 | Day 42 |
| CD154, IFN-γ, IL-2 or IL-21 expression CD4+ T-Cells on Day 0 | −0.293 (0.155) | −0.307 (0.135) | −0.290 (0.159) | −0.207 (0.321) | 0.169 (0.421) | −0.216 (0.300) | −0.239 (0.249) | −0.267 (0.197) |
| Measured Metrics | Hemagglutination Inhibition Titer (p-Value) | Neutralizing Antibody Titer (p-Value) | ||||||
|---|---|---|---|---|---|---|---|---|
| CD154, IFN-γ, IL-2 or IL-21 expression CD4+ T Cells change from baseline (Day 0) | Day 8 | Day 21 | Day 29 | Day 42 | Day 8 | Day 21 | Day 29 | Day 42 |
| Day 8–Day 0 | 0.582 (0.002) | 0.649 (<0.001) | 0.526 (0.007) | 0.265 (0.200) | 0.621 (<0.001) | 0.859 (<0.001) | 0.870 (<0.001) | 0.602 (<0.001) |
| Day 21–Day 0 | 0.667 (<0.001) | 0.542 (0.005) | 0.295 (0.153) | 0.865 (<0.001) | 0.874 (<0.001) | 0.618 (<0.001) | ||
| Day 29–Day 0 | 0.525 (0.008) | 0.279 (0.187) | 0.909 (<0.001) | 0.687 (<0.001) | ||||
| Day 42–Day 0 | 0.280 (0.196) | 0.679 (<0.001) | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, L.; Rouphael, N.; Xu, Y.; Sherman, A.C.; Edupuganti, S.; Anderson, E.J.; Lankford-Turner, P.; Wang, D.; Keitel, W.; McNeal, M.M.; et al. Baseline Levels of Influenza-Specific B Cells and T Cell Responses Modulate Human Immune Responses to Swine Variant Influenza A/H3N2 Vaccine. Vaccines 2020, 8, 126. https://doi.org/10.3390/vaccines8010126
Lai L, Rouphael N, Xu Y, Sherman AC, Edupuganti S, Anderson EJ, Lankford-Turner P, Wang D, Keitel W, McNeal MM, et al. Baseline Levels of Influenza-Specific B Cells and T Cell Responses Modulate Human Immune Responses to Swine Variant Influenza A/H3N2 Vaccine. Vaccines. 2020; 8(1):126. https://doi.org/10.3390/vaccines8010126
Chicago/Turabian StyleLai, Lilin, Nadine Rouphael, Yongxian Xu, Amy C. Sherman, Srilatha Edupuganti, Evan J. Anderson, Pamela Lankford-Turner, Dongli Wang, Wendy Keitel, Monica M. McNeal, and et al. 2020. "Baseline Levels of Influenza-Specific B Cells and T Cell Responses Modulate Human Immune Responses to Swine Variant Influenza A/H3N2 Vaccine" Vaccines 8, no. 1: 126. https://doi.org/10.3390/vaccines8010126
APA StyleLai, L., Rouphael, N., Xu, Y., Sherman, A. C., Edupuganti, S., Anderson, E. J., Lankford-Turner, P., Wang, D., Keitel, W., McNeal, M. M., Cross, K., Hill, H., Bellamy, A. R., & Mulligan, M. J. (2020). Baseline Levels of Influenza-Specific B Cells and T Cell Responses Modulate Human Immune Responses to Swine Variant Influenza A/H3N2 Vaccine. Vaccines, 8(1), 126. https://doi.org/10.3390/vaccines8010126