Enhancement of Tetravalent Immune Responses to Highly Conserved Epitopes of a Dengue Peptide Vaccine Conjugated to Polystyrene Nanoparticles


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Viruses
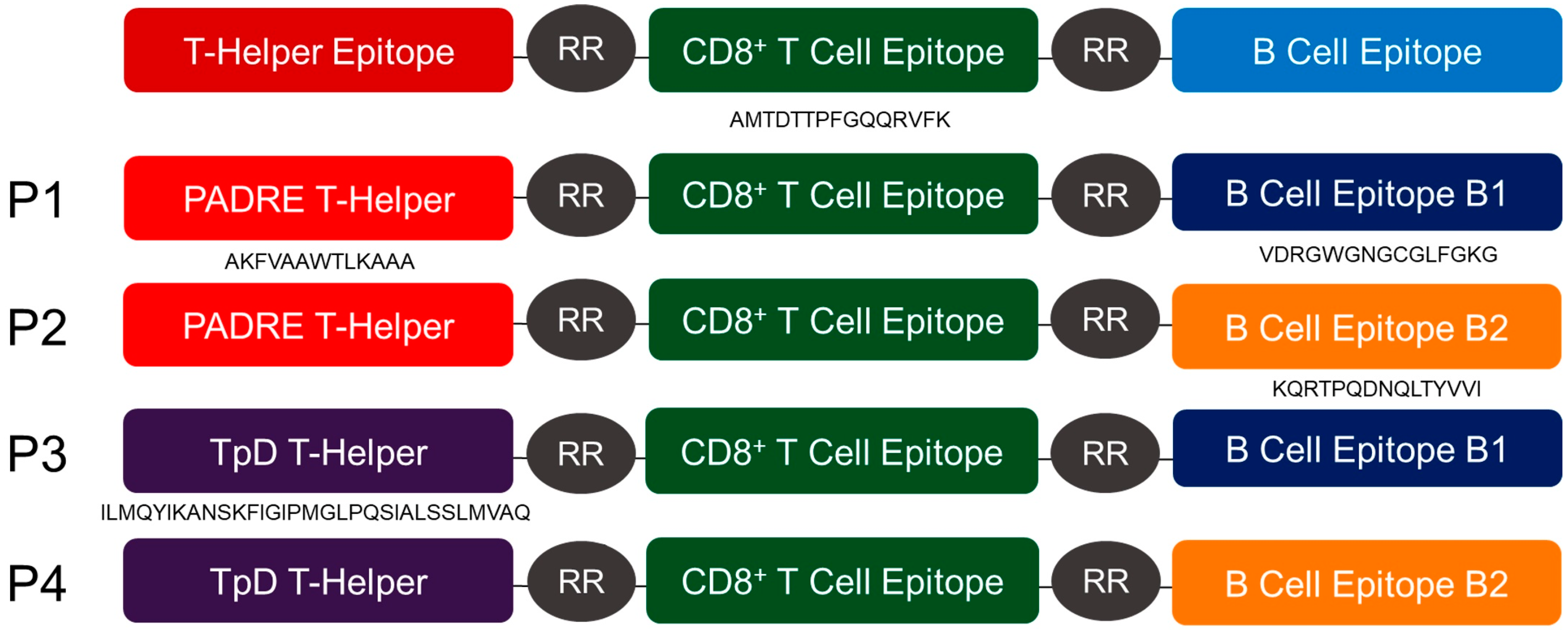
2.2. Design and Synthesis of Peptides
2.3. Conjugation of Synthetic Peptides to Carboxylated PSNPs
2.4. Determination of the Conjugation Efficiency
2.5. Determination of the Size of Nanovaccines
2.6. Determination of Nanoparticle Recovery
2.7. Transmission Electron Microscopy (TEM) Analysis
2.8. Mice and Immunization
2.9. Enzyme Linked Immunosorbent Assay (ELISA)
2.10. Focus Reduction Neutralization Test (FRNT)
2.11. IFN-γ Enzyme-Linked Immunospot (ELISPOT) Assay
2.12. Statistical Analysis
3. Results
3.1. Polystyrene-Based Nanovaccine Formulations
3.2. Humoral Immune Responses
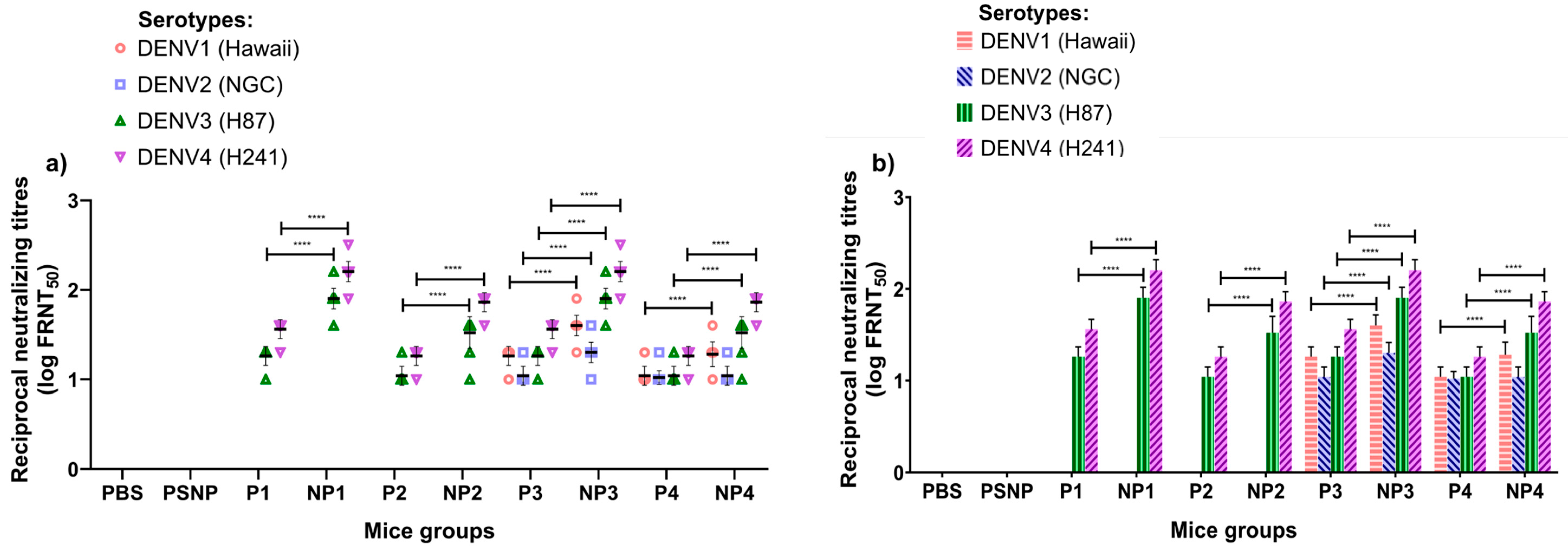
3.3. Neutralizing Antibody Responses
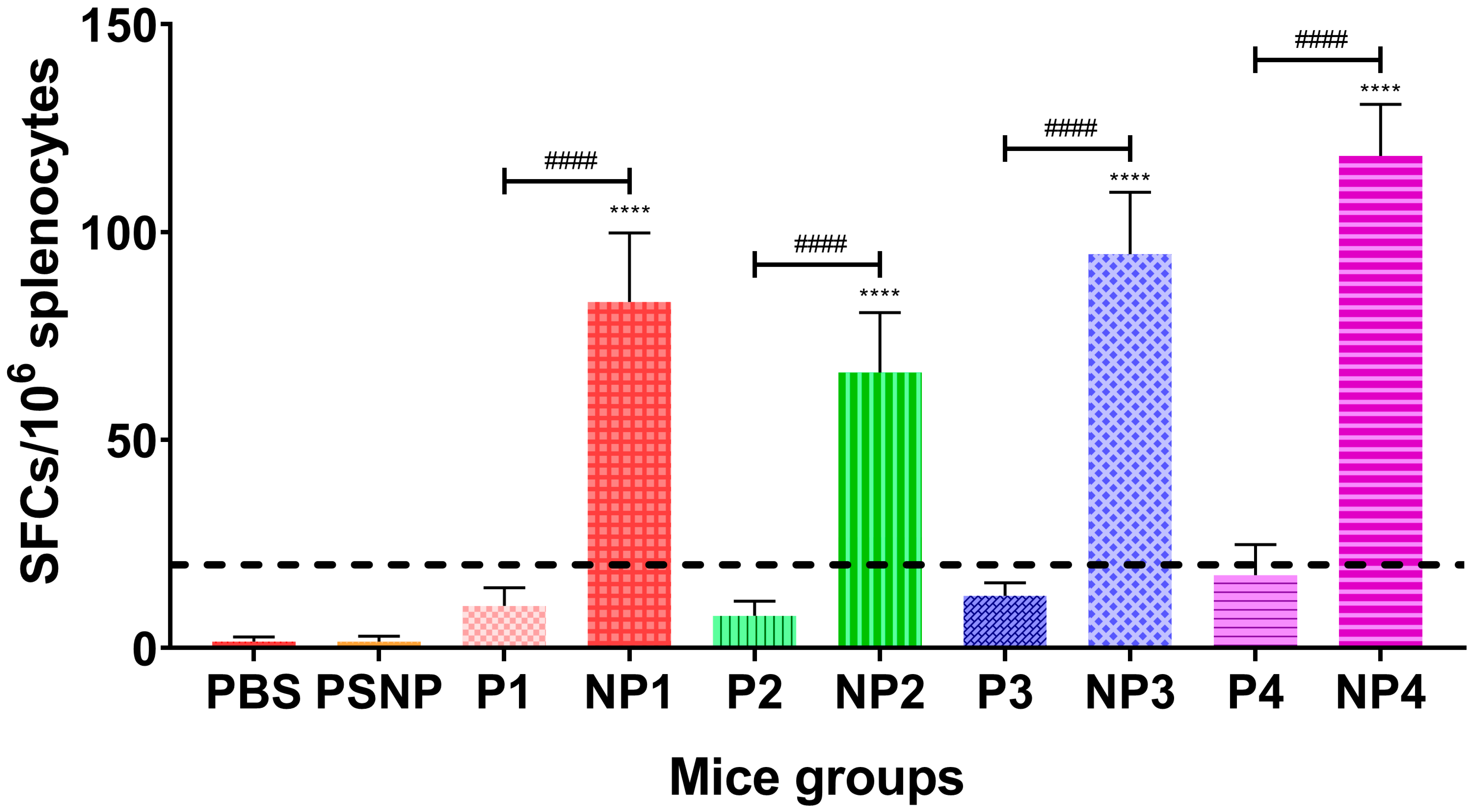
3.4. IFN-γ Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Thomas, S.J.; Rothman, A.L. Trials and tribulations on the path to developing a dengue vaccine. Vaccine 2015, 33 (Suppl. 4), D24–D31. [Google Scholar] [CrossRef]
- Sarathy, V.V.; White, M.; Li, L.; Kaiser, J.A.; Campbell, G.A.; Milligan, G.N.; Bourne, N.; Barrett, A.D.T. Characterization of a murine model of non-lethal, symptomatic dengue virus infection. Sci. Rep. 2018, 8, 4900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.R.; Zhang, S.L.; Tan, H.C.; Chan, Y.K.; Chow, A.; Lim, A.P.; Vasudevan, S.G.; Hanson, B.J.; Ooi, E.E. Ligation of Fc gamma receptor IIB inhibits antibody-dependent enhancement of dengue virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 12479–12484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejnirattisai, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-reacting antibodies enhance dengue virus infection in humans. Science (N.Y.) 2010, 328, 745–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongkolsapaya, J.; Dejnirattisai, W.; Xu, X.N.; Vasanawathana, S.; Tangthawornchaikul, N.; Chairunsri, A.; Sawasdivorn, S.; Duangchinda, T.; Dong, T.; Rowland-Jones, S.; et al. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nat. Med. 2003, 9, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Bashyam, H.S.; Green, S.; Rothman, A.L. Dengue virus-reactive CD8+ T cells display quantitative and qualitative differences in their response to variant epitopes of heterologous viral serotypes. J. Immunol. 2006, 176, 2817–2824. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, B.D.; Whitehead, S.S.; Pierce, K.K.; Tibery, C.M.; Grier, P.L.; Hynes, N.A.; Larsson, C.J.; Sabundayo, B.P.; Talaat, K.R.; Janiak, A.; et al. The live attenuated dengue vaccine TV003 elicits complete protection against dengue in a human challenge model. Sci. Transl. Med. 2016, 8, 330ra336. [Google Scholar] [CrossRef] [Green Version]
- Biswal, S.; Reynales, H.; Saez-Llorens, X.; Lopez, P.; Borja-Tabora, C.; Kosalaraksa, P.; Sirivichayakul, C.; Watanaveeradej, V.; Rivera, L.; Espinoza, F.; et al. Efficacy of a Tetravalent dengue vaccine in healthy children and adolescents. N. Engl. J. Med. 2019, 381, 2009–2019. [Google Scholar] [CrossRef]
- Diaz, C.; Lin, L.; Martinez, L.J.; Eckels, K.H.; Campos, M.; Jarman, R.G.; De La Barrera, R.; Lepine, E.; Toussaint, J.F.; Febo, I.; et al. Phase I randomized study of a tetravalent dengue purified inactivated vaccine in healthy adults from puerto rico. Am. J. Trop. Med. Hyg. 2018, 98, 1435–1443. [Google Scholar] [CrossRef]
- Boigard, H.; Cimica, V.; Galarza, J.M. Dengue-2 virus-like particle (VLP) based vaccine elicits the highest titers of neutralizing antibodies when produced at reduced temperature. Vaccine 2018, 36, 7728–7736. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Pan, C.H.; Chen, M.Y.; Hsieh, C.H.; Tsai, J.P.; Liu, H.H.; Liu, S.J.; Chong, P.; Leng, C.H.; Chen, H.W. Immunogenicity of a novel tetravalent vaccine formulation with four recombinant lipidated dengue envelope protein domain IIIs in mice. Sci. Rep. 2016, 6, 30648. [Google Scholar] [CrossRef] [PubMed]
- Flasche, S.; Jit, M.; Rodriguez-Barraquer, I.; Coudeville, L.; Recker, M.; Koelle, K.; Milne, G.; Hladish, T.J.; Perkins, T.A.; Cummings, D.A.; et al. The Long-Term Safety, Public health impact, and cost-effectiveness of routine vaccination with a recombinant, live-attenuated dengue vaccine (Dengvaxia): A model comparison study. PLoS Med. 2016, 13, e1002181. [Google Scholar] [CrossRef] [Green Version]
- Halstead, S.B. Achieving safe, effective, and durable Zika virus vaccines: Lessons from dengue. Lancet Infect. Dis. 2017, 17, e378–e382. [Google Scholar] [CrossRef] [Green Version]
- Reginald, K.; Chan, Y.; Plebanski, M.; Poh, C.L. Development of peptide vaccines in dengue. Curr. Pharm. Des. 2018, 24, 1157–1173. [Google Scholar] [CrossRef] [Green Version]
- Hammerling, G.J.; Vogt, A.B.; Kropshofer, H. Antigen processing and presentation--towards the millennium. Immunol. Rev. 1999, 172, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Jazayeri, S.D.; Poh, C.L. Development of universal influenza vaccines targeting conserved viral proteins. Vaccines 2019, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, S.R.; van Beek, J.; de Jonge, J.; Luytjes, W.; van Baarle, D. T cell responses to viral infections—Opportunities for peptide vaccination. Front. Immunol. 2014, 5, 171. [Google Scholar] [CrossRef]
- Correia-Pinto, J.F.; Csaba, N.; Schiller, J.T.; Alonso, M.J. Chitosan-poly (I:C)-PADRE based nanoparticles as delivery vehicles for synthetic peptide vaccines. Vaccines 2015, 3, 730–750. [Google Scholar] [CrossRef]
- Fraser, C.C.; Altreuter, D.H.; Ilyinskii, P.; Pittet, L.; LaMothe, R.A.; Keegan, M.; Johnston, L.; Kishimoto, T.K. Generation of a universal CD4 memory T cell recall peptide effective in humans, mice and non-human primates. Vaccine 2014, 32, 2896–2903. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Peng, L.; Zhao, W.; Zhong, H.; Zhang, F.; Yan, Z.; Cao, H. Synthetic peptides containing B- and T-cell epitope of dengue virus-2 E domain III provoked B- and T-cell responses. Vaccine 2011, 29, 3695–3702. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.D.; Gao, Q.; Wilson, K.L.; Heyerick, A.; Plebanski, M. A nanoparticle based Sp17 peptide vaccine exposes new immuno-dominant and species cross-reactive B cell epitopes. Vaccines 2015, 3, 875–893. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.L.; Xiang, S.D.; Plebanski, M. Montanide, poly I:C and nanoparticle based vaccines promote differential suppressor and effector cell expansion: A study of induction of CD8 T cells to a minimal Plasmodium berghei epitope. Front. Microbiol. 2015, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, K.; Gopinath, K.; Nandhini, D. Computational prediction of immunodominant antigenic regions & potential protective epitopes for dengue vaccination. Indian J. Med Res. 2016, 144, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Bhatnagar, S.; Kumari, K.; Mittal, N.; Sukhralia, S.; Gopirajan At, S.; Dhanaraj, P.S.; Lal, R. Highly conserved epitopes of DENV structural and non-structural proteins: Candidates for universal vaccine targets. Gene 2019, 695, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Sidney, J.; Southwood, S.; Ruppert, J.; Oseroff, C.; Maewal, A.; Snoke, K.; Serra, H.M.; Kubo, R.T.; Sette, A.; et al. Development of high potency universal DR-restricted helper epitopes by modification of high affinity DR-blocking peptides. Immunity 1994, 1, 751–761. [Google Scholar] [CrossRef]
- Shi, J.; Sun, J.; Wu, M.; Hu, N.; Li, J.; Li, Y.; Wang, H.; Hu, Y. Inferring protective CD8+ T-cell epitopes for NS5 protein of four serotypes of dengue virus chinese isolates based on HLA-A, -B and -C allelic distribution: Implications for epitope-based universal vaccine design. PLoS ONE 2015, 10, e0138729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaffari-Nazari, H.; Tavakkol-Afshari, J.; Jaafari, M.R.; Tahaghoghi-Hajghorbani, S.; Masoumi, E.; Jalali, S.A. Improving multi-epitope long peptide vaccine potency by using a strategy that enhances CD4+ T help in BALB/c mice. PLoS ONE 2015, 10, e0142563. [Google Scholar] [CrossRef]
- Leng, C.H.; Liu, S.J.; Tsai, J.P.; Li, Y.S.; Chen, M.Y.; Liu, H.H.; Lien, S.P.; Yueh, A.; Hsiao, K.N.; Lai, L.W.; et al. A novel dengue vaccine candidate that induces cross-neutralizing antibodies and memory immunity. Microbes Infect. 2009, 11, 288–295. [Google Scholar] [CrossRef]
- Dunn, M.D.; Rossi, S.L.; Carter, D.M.; Vogt, M.R.; Mehlhop, E.; Diamond, M.S.; Ross, T.M. Enhancement of anti-DIII antibodies by the C3d derivative P28 results in lower viral titers and augments protection in mice. Virol. J. 2010, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, B.; Poh, C.L.; Kirk, K.; McBride, W.J.; Aaskov, J.; Grollo, L. Synthetic B-cell epitopes eliciting cross-neutralizing antibodies: Strategies for future dengue vaccine. PLoS ONE 2016, 11, e0155900. [Google Scholar] [CrossRef] [Green Version]
- Yauch, L.E.; Zellweger, R.M.; Kotturi, M.F.; Qutubuddin, A.; Sidney, J.; Peters, B.; Prestwood, T.R.; Sette, A.; Shresta, S. A protective role for dengue virus-specific CD8+ T cells. J. Immunol. 2009, 182, 4865–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivino, L. T cell immunity to dengue virus and implications for vaccine design. Expert Rev. Vaccines 2016, 15, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Machabert, T.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A.; et al. Effect of dengue serostatus on dengue vaccine safety and efficacy. N. Engl. J. Med. 2018, 379, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Dengvaxia sensitizes seronegatives to vaccine enhanced disease regardless of age. Vaccine 2017, 35, 6355–6358. [Google Scholar] [CrossRef]
- Ali, M.; Pandey, R.K.; Khatoon, N.; Narula, A.; Mishra, A.; Prajapati, V.K. Exploring dengue genome to construct a multi-epitope based subunit vaccine by utilizing immunoinformatics approach to battle against dengue infection. Sci. Rep. 2017, 7, 9232. [Google Scholar] [CrossRef]
- Sirskyj, D.; Diaz-Mitoma, F.; Golshani, A.; Kumar, A.; Azizi, A. Innovative bioinformatic approaches for developing peptide-based vaccines against hypervariable viruses. Immunol. Cell Biol. 2011, 89, 81–89. [Google Scholar] [CrossRef]
- Khan, A.M.; Miotto, O.; Nascimento, E.J.; Srinivasan, K.N.; Heiny, A.T.; Zhang, G.L.; Marques, E.T.; Tan, T.W.; Brusic, V.; Salmon, J.; et al. Conservation and variability of dengue virus proteins: Implications for vaccine design. PLoS Negl. Trop. Dis. 2008, 2, e272. [Google Scholar] [CrossRef]
- Rathore, A.S.; Sarker, A.; Gupta, R.D. Designing antibody against highly conserved region of dengue envelope protein by in silico screening of scFv mutant library. PLoS ONE 2019, 14, e0209576. [Google Scholar] [CrossRef]
- Anandarao, R.; Swaminathan, S.; Khanna, N. The identification of immunodominant linear epitopes of dengue type 2 virus capsid and NS4a proteins using pin-bound peptides. Virus Res. 2005, 112, 60–68. [Google Scholar] [CrossRef]
- Murphy, B.R.; Whitehead, S.S. Immune response to dengue virus and prospects for a vaccine. Annu. Rev. Immunol. 2011, 29, 587–619. [Google Scholar] [CrossRef]
- Li, L.; Meng, W.; Horton, M.; DiStefano, D.R.; Thoryk, E.A.; Pfaff, J.M.; Wang, Q.; Salazar, G.T.; Barnes, T.; Doranz, B.J.; et al. Potent neutralizing antibodies elicited by dengue vaccine in rhesus macaque target diverse epitopes. PLoS Pathog. 2019, 15, e1007716. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, R.; Hu, Z.Z.; Vinayaka, C.R.; Sagripanti, J.L.; Frost, S.D.; Kosakovsky Pond, S.L.; Wu, C.H. Computational analysis and identification of amino acid sites in dengue E proteins relevant to development of diagnostics and vaccines. Virus Genes 2007, 35, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Rocha, R.P.; Livonesi, M.C.; Fumagalli, M.J.; Rodrigues, N.F.; da Costa, L.C.; Dos Santos, M.C.; de Oliveira Rocha, E.S.; Kroon, E.G.; Malaquias, L.C.; Coelho, L.F. Evaluation of tetravalent and conserved synthetic peptides vaccines derived from dengue virus envelope domain I and II. Virus Res. 2014, 188, 122–127. [Google Scholar] [CrossRef]
- Eunju, O.; Lee, Y.T.; Ko, E.J.; Kim, K.H.; Lee, Y.N.; Song, J.M.; Kwon, Y.M.; Kim, M.C.; Perez, D.R.; Kang, S.M. Roles of major histocompatibility complex class II in inducing protective immune responses to influenza vaccination. J. Virol. 2014, 88, 7764–7775. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.; Fikes, J.; Hoffman, S.; Franke, E.; Sacci, J.; Appella, E.; Chisari, F.V.; Guidotti, L.G.; Chesnut, R.W.; Livingston, B.; et al. The optimization of helper T lymphocyte (HTL) function in vaccine development. Immunol. Res. 1998, 18, 79–92. [Google Scholar] [CrossRef]
- Agadjanyan, M.G.; Ghochikyan, A.; Petrushina, I.; Vasilevko, V.; Movsesyan, N.; Mkrtichyan, M.; Saing, T.; Cribbs, D.H. Prototype Alzheimer’s disease vaccine using the immunodominant B cell epitope from beta-amyloid and promiscuous T cell epitope pan HLA DR-binding peptide. J. Immunol. 2005, 174, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, S.; Lei, J.; Zhu, Y.; Zhou, X.; Xiao, J.; Xiang, T. Prophylactic herpes simplex virus type 2 vaccine adjuvanted with a universal CD4 T cell helper peptide induces long-term protective immunity against lethal challenge in mice. Int. Immunopharmacol. 2018, 61, 100–108. [Google Scholar] [CrossRef]
- Xiang, S.D.; Wilson, K.; Day, S.; Fuchsberger, M.; Plebanski, M. Methods of effective conjugation of antigens to nanoparticles as non-inflammatory vaccine carriers. Methods (Calif.) 2013, 60, 232–241. [Google Scholar] [CrossRef]
- Fifis, T.; Gamvrellis, A.; Crimeen-Irwin, B.; Pietersz, G.A.; Li, J.; Mottram, P.L.; McKenzie, I.F.; Plebanski, M. Size-dependent immunogenicity: Therapeutic and protective properties of nano-vaccines against tumors. J. Immunol. 2004, 173, 3148–3154. [Google Scholar] [CrossRef] [Green Version]
- Mottram, P.L.; Leong, D.; Crimeen-Irwin, B.; Gloster, S.; Xiang, S.D.; Meanger, J.; Ghildyal, R.; Vardaxis, N.; Plebanski, M. Type 1 and 2 immunity following vaccination is influenced by nanoparticle size: Formulation of a model vaccine for respiratory syncytial virus. Mol. Pharm. 2007, 4, 73–84. [Google Scholar] [CrossRef]
- Fifis, T.; Mottram, P.; Bogdanoska, V.; Hanley, J.; Plebanski, M. Short peptide sequences containing MHC class I and/or class II epitopes linked to nano-beads induce strong immunity and inhibition of growth of antigen-specific tumour challenge in mice. Vaccine 2004, 23, 258–266. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science (N.Y.) 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Titta, A.; Ballester, M.; Julier, Z.; Nembrini, C.; Jeanbart, L.; van der Vlies, A.J.; Swartz, M.A.; Hubbell, J.A. Nanoparticle conjugation of CpG enhances adjuvancy for cellular immunity and memory recall at low dose. Proc. Natl. Acad. Sci. USA 2013, 110, 19902–19907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanagata, N. CpG oligodeoxynucleotide nanomedicines for the prophylaxis or treatment of cancers, infectious diseases, and allergies. Int. J. Nanomed. 2017, 12, 515–531. [Google Scholar] [CrossRef] [Green Version]
- Kelly, H.G.; Kent, S.J.; Wheatley, A.K. Immunological basis for enhanced immunity of nanoparticle vaccines. Expert Rev. Vaccines 2019, 18, 269–280. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Nanovaccines a | Conjugation pH b | Conjugation Efficiency (%) c | Recovery of Nanoparticles (%) | Size (nm) |
|---|---|---|---|---|
| NP1 | 6.5 | 76.0 ± 2.6 | 88.3 ± 1.5 | 98.1 ± 1.9 |
| NP2 | 6.5 | 76.7 ± 2.1 | 87.0 ± 1.1 | 100.8 ± 2.9 |
| NP3 | 7.5 | 75.3 ± 2.2 | 90.3 ± 1.5 | 63.1 ± 1.1 |
| NP4 | 7.5 | 73.0 ± 2.4 | 88.0 ± 1.6 | 63.4 ± 1.4 |
| Mice Groups | Mean Reciprocal IgG Antibody Titer | |||
|---|---|---|---|---|
| Day 0 | Day 14 | Day 28 | Day 42 | |
| PBS | <10 | <10 | <10 | <10 |
| PSNP | <10 | <20 | <40 | <40 |
| P1 | <10 | <20 | <40 | <80 |
| NP1 | <10 | <80 | <2560 | <10,240 |
| P2 | <10 | <20 | <40 | <80 |
| NP2 | <10 | <80 | <1280 | <5120 |
| P3 | <10 | <20 | <40 | <80 |
| NP3 | <10 | <40 | <320 | <2560 |
| P4 | <10 | <20 | <40 | <80 |
| NP4 | <10 | <40 | <160 | <1280 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, Y.; Jazayeri, S.D.; Ramanathan, B.; Poh, C.L. Enhancement of Tetravalent Immune Responses to Highly Conserved Epitopes of a Dengue Peptide Vaccine Conjugated to Polystyrene Nanoparticles. Vaccines 2020, 8, 417. https://doi.org/10.3390/vaccines8030417
Chan Y, Jazayeri SD, Ramanathan B, Poh CL. Enhancement of Tetravalent Immune Responses to Highly Conserved Epitopes of a Dengue Peptide Vaccine Conjugated to Polystyrene Nanoparticles. Vaccines. 2020; 8(3):417. https://doi.org/10.3390/vaccines8030417
Chicago/Turabian StyleChan, Yanqi, Seyed Davoud Jazayeri, Babu Ramanathan, and Chit Laa Poh. 2020. "Enhancement of Tetravalent Immune Responses to Highly Conserved Epitopes of a Dengue Peptide Vaccine Conjugated to Polystyrene Nanoparticles" Vaccines 8, no. 3: 417. https://doi.org/10.3390/vaccines8030417
APA StyleChan, Y., Jazayeri, S. D., Ramanathan, B., & Poh, C. L. (2020). Enhancement of Tetravalent Immune Responses to Highly Conserved Epitopes of a Dengue Peptide Vaccine Conjugated to Polystyrene Nanoparticles. Vaccines, 8(3), 417. https://doi.org/10.3390/vaccines8030417