Vaccine Efficacy on the Novel Reassortant H9N2 Virus in Indonesia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation and Propagation
2.2. Extraction of Viral RNA, RT-PCR
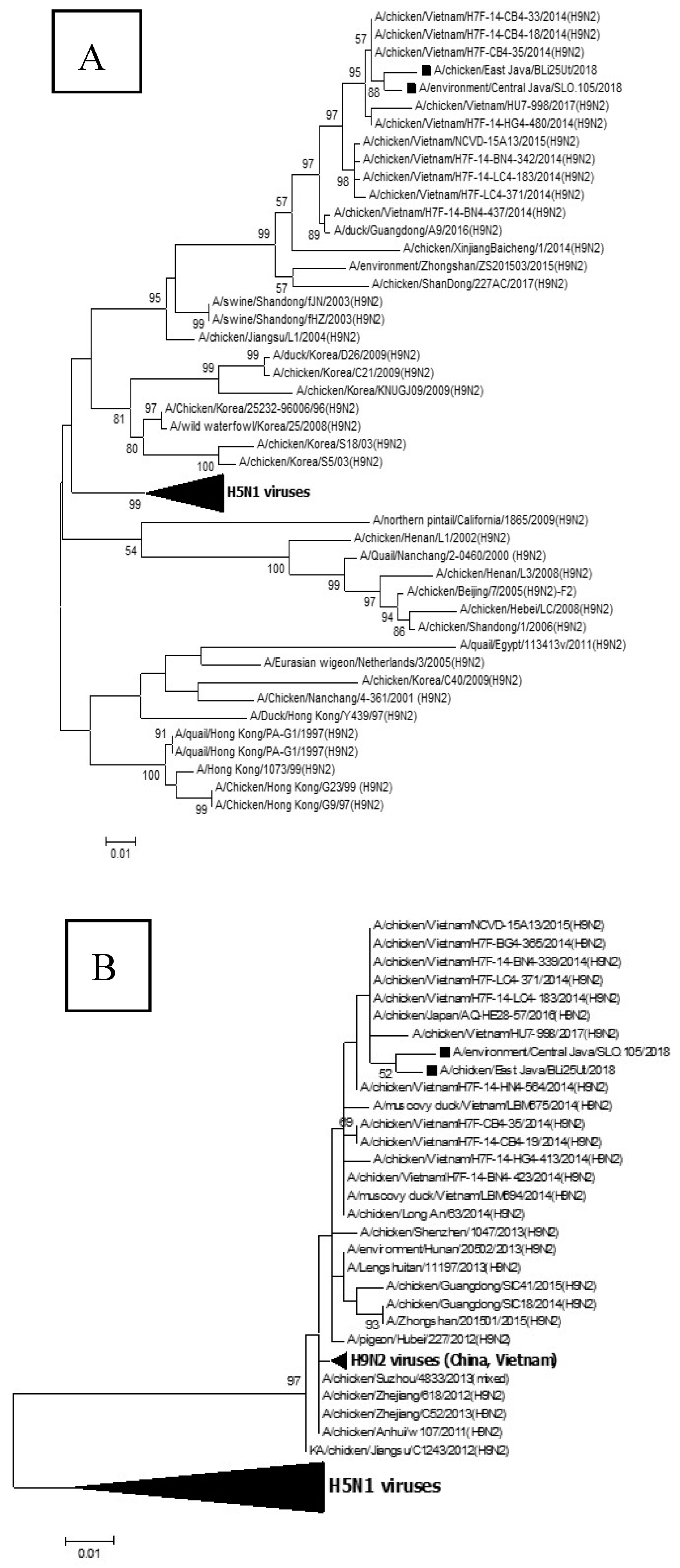
2.3. DNA Sequencing, Virus Characterization, and Phylogenetic Tree
2.4. Pathogenicity Study of AI H9N2 Virus (BLi25Ut/18)
2.5. Vaccine Efficacy Test
2.6. Statistical Analysis
3. Results
3.1. Reassortant Virus Analysis
3.2. Pathogenicity Study of BLi25Ut/18 Virus
3.3. Vaccine Efficacy against BLi25Ut/18 Virus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swayne, D.E.; Suarez, D.L. Highly pathogenic avian influenza. Rev. Sci. Tech. l’OIE 2000, 19, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Li, X.; Goraya, M.U.; Wang, S.; Chen, J.L. Evolution of influenza a virus by mutation and re-assortment. Int. J. Mol. Sci. 2017, 18, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, J.C.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E. Influenza virus: A master of metamorphosis. J. Infect. 2000, 40, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.; Lowen, A.C. Influenza A virus reassortment. In Influenza Pathogenesis and Control-Volume I; Compans, R., Oldstone, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 377–401. ISBN 978-3-319-11154-4. [Google Scholar]
- Ludwig, S.; Schultz, U.; Mandler, J.; Fitch, W.M.; Scholtissek, C. Phylogenetic relationship of the nonstructural (NS) genes of influenza A viruses. Virology 1991, 183, 566–577. [Google Scholar] [CrossRef]
- Dharmayanti, N.L.P.I.; Ibrahim, F.E.R.A.; Soebandrio, A.M.I.N. Influenza H5N1 Virus of Birds Surrounding H5N1 Human Cases Have Specific Characteristics on the Matrix Protein. HAYATI J. Biosci. 2011. [Google Scholar] [CrossRef] [Green Version]
- Dharmayanti, N.L.P.I.; Thor, S.W.; Zanders, N.; Hartawan, R.; Ratnawati, A.; Jang, Y.; Rodriguez, M.; Suarez, D.L.; Samaan, G.; Pudjiatmoko; et al. Attenuation of highly pathogenic avian influenza A(H5N1) viruses in Indonesia following the reassortment and acquisition of genes from low pathogenicity avian influenza A virus progenitors. Emerg. Microbes Infect. 2018, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dharmayanti, N.L.P.I.; Hewajuli, D.A.; Ratnawati, A.; Hartawan, R. Genetic diversity of the H5N1 viruses in live bird markets, Indonesia. J. Vet. Sci. 2020, 21. [Google Scholar] [CrossRef]
- WHO. WHO Regional Office for Europe Overview on Avian Influenza for Public Health Professionals; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Homme, P.J.; Easterday, B.C. Avian influenza virus infections. I. Characteristics of influenza A-turkey-Wisconsin-1966 virus. Avian Dis. 1970, 14, 66–74. [Google Scholar] [CrossRef]
- Peiris, M.; Yuen, K.Y.; Leung, C.W.; Chan, K.H.; Ip, P.L.S.; Lai, R.W.M.; Orr, W.K.; Shortridge, K.F. Human infection with influenza H9N2. Lancet 1999, 354, 916–917. [Google Scholar] [CrossRef]
- Ma, C.; Cui, S.; Sun, Y.; Zhao, J.; Zhang, D.; Zhang, L.; Zhang, Y.; Pan, Y.; Wu, S.; Duan, W.; et al. Avian influenza A (H9N2) virus infections among poultry workers, swine workers, and the general population in Beijing, China, 2013-2016: A serological cohort study. Influenza Other Respi. Viruses 2019, 13, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Jonas, M.; Sahesti, A.; Murwijati, T.; Lestariningsih, C.L.; Irine, I.; Ayesda, C.S.; Prihartini, W.; Mahardika, G.N. Identification of avian influenza virus subtype H9N2 in chicken farms in Indonesia. Prev. Vet. Med. 2018, 159, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Novianti, A.N.; Rahardjo, K.; Prasetya, R.R.; Nastri, A.M.; Dewantari, J.R.; Rahardjo, A.P.; Estoepangestie, A.T.S.; Shimizu, Y.K.; Poetranto, E.D.; Soegiarto, G.; et al. Whole-Genome Sequence of an Avian Influenza A/H9N2 Virus Isolated from an Apparently Healthy Chicken at a Live-Poultry Market in Indonesia. Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Zhang, Z.; Chen, W. The study of avian influenza: I. The isolation and preliminary serological identification of avian influenza virus in chicken. Chin. J. Vet. Med. 1994, 20, 3–5. [Google Scholar]
- Bonfante, F.; Mazzetto, E.; Zanardello, C.; Fortin, A.; Gobbo, F.; Maniero, S.; Bigolaro, M.; Davidson, I.; Haddas, R.; Cattoli, G.; et al. A G1-lineage H9N2 virus with oviduct tropism causes chronic pathological changes in the infundibulum and a long-lasting drop in egg production. Vet. Res. 2018, 49, 83. [Google Scholar] [CrossRef] [Green Version]
- Wiyono, A.; Indriani, R.; Dharmayanti, N.; Damayant, R.; Parede, L.; Syafriati, T. Darminto Isolasi Dan Karakterisasi Virus Highly Pathogenic Avian Influenza Subtipe H5 Dari Ayam Asal Wabah Di Indonesia ( Isolation And Characterization Of Virus Of Highly Pathogenic Avian Influenza H5 Subtype Of Chicken From Outbreaks In Indonesia ). J. Ilmu Ternak dan Vet. 2004, 9, 2004. [Google Scholar]
- Zhu, W.; Dong, J.; Zhang, Y.; Yang, L.; Li, X.; Chen, T.; Zhao, X.; Wei, H.; Bo, H.; Zeng, X.; et al. A Gene Constellation in Avian Influenza A (H7N9) Viruses May Have Facilitated the Fifth Wave Outbreak in China. Cell Rep. 2018, 23, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Arai, Y.; Ibrahim, M.S.; Elgendy, E.M.; Daidoji, T.; Ono, T.; Suzuki, Y.; Nakaya, T.; Matsumoto, K.; Watanabe, Y. Genetic Compatibility of Reassortants between Avian H5N1 and H9N2 Influenza Viruses with Higher Pathogenicity in Mammals. J. Virol. 2018, 93. [Google Scholar] [CrossRef] [Green Version]
- OIE. Avian Influenza (Infection With Avian Influenza Viruses). In OIE Terrestrial Manual; OIE: Paris, France, 2018; pp. 821–843. [Google Scholar]
- Dharmayanti, N.L.P.I.; Hartawan, R.; Pudjiatmoko, H.W.; Hardiman, A.B.; Donis, R.; Davis, C.T.; Samaan, G. Genetic characterization of clade 2.3.2.1 avian influenza A(H5N1) viruses, Indonesia, 2012. Emerg. Infect. Dis. 2014, 20, 671–674. [Google Scholar] [CrossRef]
- Indriani, R.; Dharmayanti, N.; Syakir, M. Vaksin Kombinasi Avian Influenza Hpai Dan Lpai, [Patent]. 2019, pp. 1–13. Available online: https://pdki-indonesia.dgip.go.id/index.php/paten/eDdOQjRNVFhHZDB5NnRkM1YzbkNuQT09?q=vaksin+kombinasi&type=1 (accessed on 10 June 2020).
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938. [Google Scholar] [CrossRef]
- Monne, I.; Yamage, M.; Dauphin, G.; Claes, F.; Ahmed, G.; Giasuddin, M.; Salviato, A.; Ormelli, S.; Bonfante, F.; Schivo, A.; et al. Reassortant Avian Influenza A(H5N1) Viruses with H9N2-PB1 Gene in Poultry, Bangladesh. Emerg. Infect. Dis. 2013, 19, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Marinova-Petkova, A.; Feeroz, M.M.; Rabiul Alam, S.; Kamrul Hasan, M.; Akhtar, S.; Jones-Engel, L.; Walker, D.; McClenaghan, L.; Rubrum, A.; Franks, J.; et al. Multiple introductions of highly pathogenic avian influenza H5N1 viruses into Bangladesh. Emerg. Microbes Infect. 2014, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nishi, T.; Okamatsu, M.; Sakurai, K.; Chu, H.D.; Thanh, L.P.; van Nguyen, L.; van Hoang, N.; Thi, D.N.; Sakoda, Y.; Kida, H. Genetic Analysis of an H5N2 Highly Pathogenic Avian Influenza Virus Isolated from a Chicken in a Live Bird Market in Northern Vietnam in 2012. J. Vet. Med. Sci. 2014, 76, 85–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Meng, F.; Huang, D.; Sheng, X.; Wang, Y.; Zhang, W.; Chang, W.; Wang, L.; Qin, Z. Genomic and Phylogenetic Characterization of Novel, Recombinant H5N2 Avian Influenza Virus Strains Isolated from Vaccinated Chickens with Clinical Symptoms in China. Viruses 2015, 7, 887–898. [Google Scholar] [CrossRef]
- Naguib, M.M.; Kinne, J.; Chen, H.; Chan, K.-H.; Joseph, S.; Wong, P.-C.; Woo, P.C.Y.; Wernery, R.; Beer, M.; Wernery, U.; et al. Outbreaks of highly pathogenic avian influenza H5N1 clade 2.3.2.1c in hunting falcons and kept wild birds in Dubai implicate intercontinental virus spread. J. Gen. Virol. 2015, 96, 3212–3222. [Google Scholar] [CrossRef]
- Tosh, C.; Nagarajan, S.; Kumar, M.; Murugkar, H.V.; Venkatesh, G.; Shukla, S.; Mishra, A.; Mishra, P.; Agarwal, S.; Singh, B.; et al. Multiple introductions of a reassortant H5N1 avian influenza virus of clade 2.3.2.1c with PB2 gene of H9N2 subtype into Indian poultry. Infect. Genet. Evol. 2016, 43, 173–178. [Google Scholar] [CrossRef]
- Bi, Y.; Zhang, Z.; Liu, W.; Yin, Y.; Hong, J.; Li, X.; Wang, H.; Wong, G.; Chen, J.; Li, Y.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Struck Migratory Birds in China in 2015. Sci. Rep. 2015, 5, 12986. [Google Scholar] [CrossRef] [Green Version]
- Monne, I.; Meseko, C.; Joannis, T.; Shittu, I.; Ahmed, M.; Tassoni, L.; Fusaro, A.; Cattoli, G. Highly Pathogenic Avian Influenza A(H5N1) Virus in Poultry, Nigeria, 2015. Emerg. Infect. Dis. 2015, 21, 1275–1277. [Google Scholar] [CrossRef]
- Suttie, A.; Karlsson, E.A.; Deng, Y.-M.; Horm, S.V.; Yann, S.; Tok, S.; Sorn, S.; Holl, D.; Tum, S.; Hurt, A.C.; et al. Influenza A(H5N1) viruses with A(H9N2) single gene (matrix or PB1) reassortment isolated from Cambodian live bird markets. Virology 2018, 523, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Deng, Y.-M. Influenza virus antigenic variation, host antibody production and new approach to control epidemics. Virol. J. 2009, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.C. Reassortment of influenza A virus genes linked to PB1 polymerase gene. Int. Congr. Ser. 2004, 1263, 714–718. [Google Scholar] [CrossRef]
- Urbaniak, K.; Markowska-Daniel, I.; Kowalczyk, A.; Kwit, K.; Pomorska-Mól, M.; Fracek, B.; Pejsak, Z. Reassortment process after co-infection of pigs with avian H1N1 and swine H3N2 influenza viruses. BMC Vet. Res. 2017, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Cao, B.; Hu, Y.; Feng, Z.; Wang, D.; Hu, W.; Chen, J.; Jie, Z.; Qiu, H.; Xu, K.; et al. Human Infection with a Novel Avian-Origin Influenza A (H7N9) Virus. N. Engl. J. Med. 2013, 368, 1888–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, C.; Liu, Q.; Bawa, B.; Shen, H.; Qi, W.; Chen, Y.; Mok, C.K.P.; García-Sastre, A.; Richt, J.A.; Ma, W. Pathogenicity and transmissibility of reassortant H9 influenza viruses with genes from pandemic H1N1 virus. J. Gen. Virol. 2012, 93, 2337–2345. [Google Scholar] [CrossRef]
- Kim, S.-H. Challenge for One Health: Co-Circulation of Zoonotic H5N1 and H9N2 Avian Influenza Viruses in Egypt. Viruses 2018, 10, 121. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-K.; Seiler, P.; Forrest, H.L.; Khalenkov, A.M.; Franks, J.; Kumar, M.; Karesh, W.B.; Gilbert, M.; Sodnomdarjaa, R.; Douangngeun, B.; et al. Pathogenicity and Vaccine Efficacy of Different Clades of Asian H5N1 Avian Influenza A Viruses in Domestic Ducks. J. Virol. 2008, 82, 11374–11382. [Google Scholar] [CrossRef] [Green Version]
- Capua, I.; Marangon, S. Control of avian influenza in poultry. Emerg. Infect. Dis. 2006, 12, 1319–1324. [Google Scholar] [CrossRef]
- Sims, L.; Tripodi, A.; Gardner, E.; Aguanno, R. Rational Use of Vaccination for Prevention and Control of H5 Highly Pathogenic Avian Influenza; The Food and Agriculture Organization (FAO): Rome, Italy, 2016; Volume 10, pp. 1–12. [Google Scholar]
- Gomaa, M.R.; Khalil, A.A.; Kandeil, A.; Sabir, J.S.M.; Kayed, A.; Moatasim, Y.; El saied, M.F.; El-safty, M.M.; Kayali, G.; Ali, M.A. Development of an effective contemporary trivalent avian influenza vaccine against circulating H5N1, H5N8, and H9N2 in Egypt. Poult. Sci. 2019, 98, 6289–6295. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antisera | BLi25Ut/18 |
|---|---|
| H9N2/2017 | 10 log2 |
| H5N1 Clade 2.1.3 | Negative |
| H5N1 Clade 2.3.2 | Negative |
| ND genotype VII | Negative |
| ND genotype VI | Negative |
| ND RIVs genotype 2 | Negative |
| Post-Infection (Days) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| Infected | h:10 | h:10 | h:10 | s:10d | s:10a,d | s: 9d dt:1 | s:8d h:1 | s:6d h:3 | s:6d h:3 | s:6d h:3 | s:6d h:3 | s:6d h:3 | s:6d h:3 | s:6d h:3 |
| Control | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 | h:10 |
| Challenge Group | Virus Shedding in SPF Chickens (Day Post-Challenge) | ||
|---|---|---|---|
| Days 4 | Days 8 | Days 14 | |
| Group 1. Inactivated bivalent vaccine | 3/10 | 2/10 | 0/10 |
| Group 2. Inactivated monovalent H9N2 reassortant vaccine | 2/10 | 1/10 | 0/10 |
| Group 3. Unvaccinated SPF chickens | 9/10 | 4/9 * | 2/9 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dharmayanti, N.L.P.I.; Indriani, R.; Nurjanah, D. Vaccine Efficacy on the Novel Reassortant H9N2 Virus in Indonesia. Vaccines 2020, 8, 449. https://doi.org/10.3390/vaccines8030449
Dharmayanti NLPI, Indriani R, Nurjanah D. Vaccine Efficacy on the Novel Reassortant H9N2 Virus in Indonesia. Vaccines. 2020; 8(3):449. https://doi.org/10.3390/vaccines8030449
Chicago/Turabian StyleDharmayanti, Ni Luh Putu Indi, Risa Indriani, and Diana Nurjanah. 2020. "Vaccine Efficacy on the Novel Reassortant H9N2 Virus in Indonesia" Vaccines 8, no. 3: 449. https://doi.org/10.3390/vaccines8030449
APA StyleDharmayanti, N. L. P. I., Indriani, R., & Nurjanah, D. (2020). Vaccine Efficacy on the Novel Reassortant H9N2 Virus in Indonesia. Vaccines, 8(3), 449. https://doi.org/10.3390/vaccines8030449