A Novel Rhoptry Protein as Candidate Vaccine against Eimeria tenella Infection

{kind=link}
{kind=link}
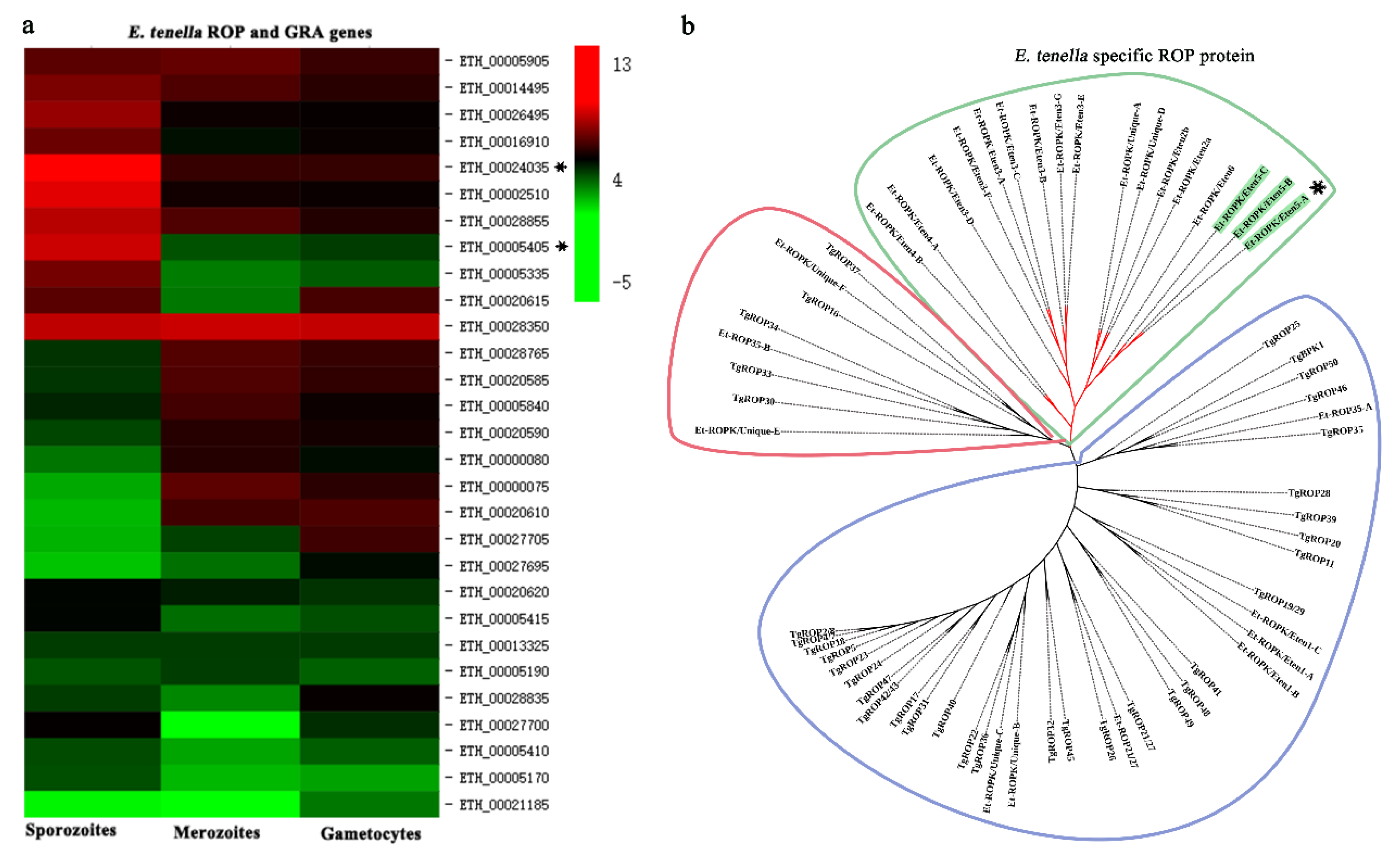{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Parasites and Animals
2.3. Bioinformatic Analysis
2.4. Cloning, Expression and Purification of Four Sporozoite Antigens
2.5. Sera
2.6. Immunoblotting and Immunofluorescence Assay
2.7. The Protective Efficacy Comparison of Four Recombinant Sporozoite Proteins
2.8. Statistical Analysis
3. Results
3.1. Screening and Characterization of Four Sporozoite Antigens
3.2. Cloning and Expression of the Four Sporozoite Proteins
3.3. Immunoblot Analysis of the Four Recombinant Sporozoite Proteins
3.4. Immunolocalizations of Endogenous Et-ROPK-Eten5-A and Et-GRA12
3.5. Recombinant Et-ROPK-Eten5-A Induces Effective Protection against E. tenella
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Paul, G.C.; Friend, D.G. Clostridial enterotoxemia and coccidiosis in weanling cottontail rabbits (Sylvilagus audubonii, Sylvilagus floridanus, Sylvilagus nuttallii) from Colorado, USA. J. Wildl. Dis. 2019, 55, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.D.; Barta, J.R.; Blake, D.; Gruber, A.; Jenkins, M.; Smith, N.C.; Suo, X.; Tomley, F.M. A selective review of advances in coccidiosis research, in Advances in parasitology. Adv. Parasitol. 2013, 83, 93–171. [Google Scholar] [PubMed]
- Blake, D.P.; Tomley, F.M. Securing poultry production from the ever-present Eimeria challenge. Trends Parasitol. 2014, 30, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Witcombe, D.M.; Smith, N.C. Strategies for anti-coccidial prophylaxis. Parasitology 2014, 141, 1379–1389. [Google Scholar] [CrossRef]
- Bussière, F.I.; Niepceron, A.; Sausset, A.; Esnault, E.; Silvestre, A.; Walker, R.A.; Smith, N.C.; Quéré, P.; Laurent, F. Establishment of an in vitro chicken epithelial cell line model to investigate Eimeria tenella gamete development. Parasit. Vectors 2018, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odden, A.; Denwood, M.J.; Stuen, S.; Robertson, L.J.; Ruiz, A.; Hamnes, I.S.; Hektoen, L.; Enemark, H.L. Field evaluation of anticoccidial efficacy: A novel approach demonstrates reduced efficacy of toltrazuril against ovine Eimeria spp. in Norway. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 304–311. [Google Scholar] [CrossRef]
- Venkatas, J.; Adeleke, M. A review of Eimeria antigen identification for the development of novel anticoccidial vaccines. Parasitol. Res. 2019, 118, 1701–1710. [Google Scholar] [CrossRef]
- Song, X.; Ren, Z.; Yan, R.; Xu, L.; Li, X. Induction of protective immunity against Eimeria tenella, Eimeria necatrix, Eimeria maxima and Eimeria acervulina infections using multivalent epitope DNA vaccines. Vaccine 2015, 33, 2764–2770. [Google Scholar] [CrossRef]
- Zhao, P.; Li, Y.; Zhou, Y.; Zhao, J.; Fang, R. In vivo immunoprotective comparison between recombinant protein and DNA vaccine of Eimeria tenella surface antigen 4. Vet. Parasitol. 2020, 278, 109032. [Google Scholar] [CrossRef]
- Chapman, H.D.; Jeffers, T.K. Vaccination of chickens against coccidiosis ameliorates drug resistance in commercial poultry production. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Ziomko, I.; Karamon, J.; Cencek, T.; Gornowicz, E.; Ashash, U. Prevention of broiler chick coccidiosis using the inactivated subunit vaccine CoxAbic®. Bull. Vet. Inst. Pulawy. 2005, 49, 299–302. [Google Scholar]
- Lebrun, M.; Carruthers, V.B.; Cesbron-Delauw, M.-F. Toxoplasma secretory proteins and their roles in cell invasion and intracellular survival. In Toxoplasma Gondii; Elsevier: Amsterdam, The Netherlands, 2014; pp. 389–453. [Google Scholar]
- Bradley, P.J.; Ward, C.; Cheng, S.J.; Alexander, D.L.; Coller, S.; Coombs, G.H.; Dunn, J.D.; Ferguson, D.J.; Sanderson, S.J.; Wastling, J.M.; et al. Proteomic Analysis of Rhoptry Organelles Reveals Many Novel Constituents for Host-Parasite Interactions in Toxoplasma gondii. J. Biol. Chem. 2005, 280, 34245–34258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajj, H.E.; Demey, E.; Poncet, J.; Lebrun, M.; Wu, B.; Galéotti, N.; Fourmaux, M.N.; Mercereau-Puijalon, O.; Vial, H.; Labesse, G.; et al. The ROP2 family of Toxoplasma gondii rhoptry proteins: Proteomic and genomic characterization and molecular modeling. Proteomics 2006, 6, 5773–5784. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, L.; Chen, F.; Harb, O.S.; Davis, P.H.; Beiting, D.P.; Brownback, C.S.; Ouloguem, D.; Roos, D.S. Integrative Genomic Approaches Highlight a Family of Parasite-Specific Kinases that Regulate Host Responses. Cell Host Microbe 2010, 8, 208–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafiqi, S.I.; Garg, R.; Reena, K.K.; Ram, H.; Singh, M.; Banerjee, P.S. Immune response and protective efficacy of Eimeria tenella recombinant refractile body protein, EtSO7, in chickens. Vet. Parasitol. 2018, 258, 108–113. [Google Scholar] [CrossRef]
- Subramanian, B.M.; Sriraman, R.; Rao, N.H.; Raghul, J.; Thiagarajan, D.; Srinivasan, V.A. Cloning, expression and evaluation of the efficacy of a recombinant Eimeria tenella sporozoite antigen in birds. Vaccine 2008, 26, 3489–3496. [Google Scholar] [CrossRef]
- Li, J.; Zheng, J.; Gong, P.; Zhang, X. Efficacy of Eimeria tenella rhomboid-like protein as a subunit vaccine in protective immunity against homologous challenge. Parasitol. Res. 2012, 110, 1139–1145. [Google Scholar] [CrossRef]
- Jahn, D.; Matros, A.; Bakulina, A.Y.; Tiedemann, J.; Schubert, U.; Giersberg, M.; Haehnel, S.; Zoufal, K.; Mock, H.P.; Kipriyanov, S.M. Model structure of the immunodominant surface antigen of Eimeria tenella identified as a target for sporozoite-neutralizing monoclonal antibody. Parasitol. Res. 2009, 105, 655. [Google Scholar] [CrossRef]
- Song, X.; Gao, Y.; Xu, L.; Yan, R.; Li, X. Partial protection against four species of chicken coccidia induced by multivalent subunit vaccine. Vet. Parasitol. 2015, 212, 80–85. [Google Scholar] [CrossRef]
- Blake, D.P.; Pastor-Fernández, I.; Nolan, M.J.; Tomley, F.M. Recombinant anticoccidial vaccines-a cup half full? Infect. Genet. Evol. 2017, 55, 358–365. [Google Scholar] [CrossRef]
- Jenkins, M.; Fetterer, R.; Miska, K.; Tuo, W.; Kwok, O.; Dubey, J.P. Characterization of the Eimeria maxima sporozoite surface protein IMP1. Vet. Parasitol. 2015, 211, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Lin, Q.; Qiu, J.; Qin, M.; Tang, X.; Suo, X.; Huang, Z.; Liu, X. Immunogenicity and protective efficacy of an Eimeria vaccine candidate based on Eimeria tenella immune mapped protein 1 and chicken CD40 ligand. Vet. Parasitol. 2015, 210, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Lin, J.; Han, H.; Zhao, Q.; Dong, H.; Zhu, S.; Huang, B. Identification and partial characterization of a serine protease inhibitor (serpin) of Eimeria tenella. Parasitol. Res. 2012, 110, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Long, P.L.; Millard, B.J.; Joyner, L.P.; Norton, C.C. A guide to laboratory techniques used in the study and diagnosis of avian coccidiosis. Folia Vet. Lat. 1976, 6, 201–217. [Google Scholar]
- Walker, R.A.; Sharman, P.A.; Miller, C.M.; Lippuner, C.; Okoniewski, M.; Eichenberger, R.M.; Ramakrishnan, C.; Brossier, F.; Deplazes, P.; Hehl, A.B. RNA Seq analysis of the Eimeria tenella gametocyte transcriptome reveals clues about the molecular basis for sexual reproduction and oocyst biogenesis. BMC Genom. 2015, 16, 94. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, H.; Liu, J.; Hao, P.; Ma, L.; Liu, Q. The Apoptotic Role of Metacaspase in Toxoplasma gondii. Front. Microbiol. 2015, 6, 1560. [Google Scholar] [CrossRef] [Green Version]
- Schmatz, D.M.; Crane, M.S.; Murray, P.K. Purification of Eimeria sporozoites by DE-52 anion exchange chromatography. J. Eukaryot. Microbiol. 2010, 31, 181–183. [Google Scholar]
- Hu, D.; Wang, C.; Wang, S.; Tang, X.; Duan, C.; Zhang, S.; Suo, J.; Deng, M.; Lv, Y.; Suo, X. Comparative transcriptome analysis of Eimeria maxima (Apicomplexa: Eimeriidae) suggests DNA replication activities correlating with its fecundity. BMC Genom. 2018, 19, 1–10. [Google Scholar] [CrossRef]
- Johnson, J.; Reid, W.M. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef]
- Reid, A.J.; Blake, D.P.; Ansari, H.R.; Billington, K.; Browne, H.P.; Bryant, J.; Dunn, M.; Hung, S.S.; Kawahara, F.; Miranda-Saavedra, D. Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Res. 2014, 24, 1676–1685. [Google Scholar] [CrossRef] [Green Version]
- Talevich, E.; Kannan, N. Structural and evolutionary adaptation of rhoptry kinases and pseudokinases, a family of coccidian virulence factors. BMC Evol. Biol. 2013, 13, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carruthers, V.; Boothroyd, J.C. Pulling together: An integrated model of Toxoplasma cell invasion. Curr. Opin. Microbiol. 2007, 10, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.A.; Sausset, A.; Gnahoui-David, A.; Silva, A.; Brionne, A.; Le Vern, Y.; Bussière, F.I.; Tottey, J.; Lacroix-Lamandé, S.; Laurent, F. Eimeria tenella ROP kinase EtROP1 induces G0/G1 cell cycle arrest and inhibits host cell apoptosis. Cell Microbiol. 2019, 21, e13027. [Google Scholar] [CrossRef] [Green Version]
- Rick, B.; Dubremetz, J.-F.; Entzeroth, R. A merozoite-specific 22-kDa rhoptry protein of the coccidium Eimeria nieschulzi (Sporozoa, Coccidia) is exocytosed in the parasitophorous vacuole upon host cell invasion. Parasitol. Res. 1998, 84, 291–296. [Google Scholar] [CrossRef]
- Yang, C.; Liu, J.; Ma, L.; Zhang, X.; Zhang, X.; Zhou, B.; Zhu, X.; Liu, Q. NcGRA17 is an important regulator of parasitophorous vacuole morphology and pathogenicity of Neospora caninum. Vet. Parasitol. 2018, 264, 26–34. [Google Scholar] [CrossRef]
- Song, X.; Zhao, X.; Xu, L.; Yan, R.; Li, X. Immune protection duration and efficacy stability of DNA vaccine encoding Eimeria tenella TA4 and chicken IL-2 against coccidiosis. Res. Vet. Sci. 2017, 111, 31–35. [Google Scholar] [CrossRef]
- Dong, H.; Yang, S.; Zhao, Q.; Han, H.; Zhu, S.; Zhu, X.; Li, C.; Wang, Z.; Xia, W.; Men, Q. Molecular characterization and protective efficacy of silent information regulator 2A from Eimeria tenella. Parasit. Vectors 2016, 9, 602. [Google Scholar] [CrossRef] [Green Version]
- Williams, R. Anticoccidial vaccines for broiler chickens: Pathways to success. Avian. Pathol. 2002, 31, 317–353. [Google Scholar] [CrossRef]
- Lin, R.-Q.; Lillehoj, H.S.; Lee, S.K.; Oh, S.; Panebra, A.; Lillehoj, E.P. Vaccination with Eimeria tenella elongation factor-1α recombinant protein induces protective immunity against E. tenella and E. maxima infections. Vet. Parasitol. 2017, 243, 79–84. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Yang, X.; Zhang, T.; Liu, J.; Liu, Q. A Novel Rhoptry Protein as Candidate Vaccine against Eimeria tenella Infection. Vaccines 2020, 8, 452. https://doi.org/10.3390/vaccines8030452
Song X, Yang X, Zhang T, Liu J, Liu Q. A Novel Rhoptry Protein as Candidate Vaccine against Eimeria tenella Infection. Vaccines. 2020; 8(3):452. https://doi.org/10.3390/vaccines8030452
Chicago/Turabian StyleSong, Xingju, Xu Yang, Taotao Zhang, Jing Liu, and Qun Liu. 2020. "A Novel Rhoptry Protein as Candidate Vaccine against Eimeria tenella Infection" Vaccines 8, no. 3: 452. https://doi.org/10.3390/vaccines8030452
APA StyleSong, X., Yang, X., Zhang, T., Liu, J., & Liu, Q. (2020). A Novel Rhoptry Protein as Candidate Vaccine against Eimeria tenella Infection. Vaccines, 8(3), 452. https://doi.org/10.3390/vaccines8030452