1. Introduction
Equine influenza (EI), caused by equine influenza virus (EIV), is highly contagious and is one of the major viral respiratory diseases in horses [
1]. Equine influenza outbreaks occur worldwide (except in Iceland, New Zealand, and Australia) with high morbidity and potentially huge economic consequences [
2,
3]. EIV infection produces a considerable spectrum of clinical disease, with characteristic clinical signs seen in naïve horses and less severe disease generally seen in vaccinated horses, with subclinical or mild and transient clinical signs. There is evidence that infection with different EIV strains (genotypes) results in different clinical outcomes (disease-causing phenotypes). Certain strains appear more likely to produce primary viral pneumonia and others are more likely to be followed by secondary bacterial infection, while the remainder produce less severe disease.
The most effective method of limiting EIV infection and outbreaks is vaccination [
4]. Several EIV vaccines are available commercially and these can be divided into three groups based on vaccine type: 1―adjuvanted, whole or subunit, inactivated virus vaccines; 2―adjuvanted, modified viral vector vaccine; 3―live attenuated virus vaccine [
4]. An adjuvant is included to stimulate the host immune response to the target antigen(s) in the whole inactivated or subunit virus vaccines and modified viral vector vaccine. The adjuvanted vaccines are administered by deep intramuscular injection and the live attenuated vaccine by intranasal application.
Population (herd) vaccination coverage is needed to control EI. However, many horse owners are unaware that vaccination does not protect an individual horse from infection and disease but helps to reduce the prevalence and severity of disease. Moreover, a small percentage of the population fails to mount and/or maintain an adequate immune response after appropriate immunisation and, therefore, remains susceptible to EI despite having been vaccinated [
5,
6,
7]. Although the reasons for this remain unclear, age appears to be a significant factor [
8]. There is also likely to be a strong host effect, with genotype and previous immune experience also playing roles [
9,
10]. The timing of vaccination is important. A primary vaccination course usually consists of two vaccinations (V1 and V2) followed by an initial revaccination (V3) 5–6 months after the primary vaccination course. In fact, EIV-specific antibody titres can decline significantly between V2 and V3, meaning that some horses may be protected insufficiently for several weeks [
11,
12]. This so-called immunity gap poses a risk to all EI vaccines [
13]. Horses infected during the immunity gap may show little or no clinical signs but may shed sufficient virus to infect naïve, unvaccinated horses, as demonstrated in an experimental study using sentinels [
5,
14].
It is believed that vaccine efficacy can be compromised by a mismatch between the vaccine and field virus strain(s). The antigenic evolution of EIV is closely monitored by the OIE (World Organisation for Animal Health) and associated laboratories and institutes. For this purpose, EIV sequences are analysed for antigenic variation using predictive tools and models (e.g., antigenic cartography, differentiation of viruses using haemagglutination inhibition (HI) with ferret sera). The OIE expert surveillance panel reviews the laboratory and epidemiological data on the circulation of EIV worldwide and, based on this, anticipates the impact of antigenic shift on vaccine efficacy and makes annual recommendations on the antigenic composition of EIV vaccines [
4,
15]. Nowadays, EIV strains belonging to the Florida Clade (FC) 1 and 2 sublineages predominate, and these have been included in the OIE recommendations for EIV vaccines since 2010, along with the recommendation that a European lineage is no longer required [
15]. Since 2010, there have been no changes in the expert surveillance panel recommendations, whereas human seasonal influenza vaccines are updated annually. This is in line with previous findings that EIVs are genetically more stable than human influenza viruses [
16,
17].
The method of analysis of EIV antigenic evolution, on which these recommendations are based, and the criteria for updating EIV vaccines are identical to those used for human seasonal influenza vaccines [
18,
19]. Haemagglutination inhibition (HI) assays with ferret antisera are used, amongst other techniques, to analyse the antigenic changes in the haemagglutinin (HA) viral surface protein. Ferrets are known to be the most sensitive species for influenza virus antigenic variation and mount very strain-specific responses [
18,
20]. This means that ferrets can discriminate minor antigenic differences from major antigenic differences. However, these apparently large antigenic differences in EIV may not be biologically significant in horses [
20,
21]. This has already been observed for avian influenza where the vaccine is capable of providing sufficient protection against infection with new field strains although the vaccine virus strain differs from field virus [
22,
23]. It is possible that the vaccine type may play a more important role in determining EIV vaccine efficacy than minor antigenic differences between vaccine and field strains that may not be biologically relevant in the horse.
The process of updating an EIV vaccine can take years and is costly, and there is strict guidance on the requirements for demonstrating vaccine efficacy, safety, and quality that go beyond the OIE expert surveillance panel recommendations for EIV strains [
24]. To date, there is only one commercially available vaccine that is able to comply with all of the current OIE expert surveillance panel recommendations. This is relatively simple for the modified vector vaccine, where the HA in the replicative defective canarypox (ALVAC) vector expressing the HA of the EIV strain in question is replaced, with HA expression being confirmed by Western blot using a monoclonal anti-EIV HA antibody, and the final vaccine construct is titrated in chicken embryo fibroblast cell culture using HA-specific immunofluorescence [
25]. However, for other EIV vaccines, this is more challenging because in addition to replacing the whole EI virus or subunit, it has to be possible to differentiate the new virus strain(s) from the existing strain in the vaccine using the originally registered tests.
This study evaluated the duration of immunity at 4.5 months after the primary vaccination course (V1 and V2) of the adjuvanted, modified vector vaccine (expressing the HA gene of a FC1 and a FC2 strain) and a ISCOMatrix adjuvanted, whole inactivated virus vaccine (containing a FC1 and a European lineage strain) based on experimental infection with a recent Florida Clade 2 field strain. The aim was to provide insight into the impact of vaccine type and minor antigenic differences between vaccine and field strains on vaccine efficacy, after challenge during the most susceptible period (the immunity gap). The study showed that the two vaccines were very similar based on induced protective antibody titres (single radial haemolysis (SRH), haemagglutination inhibition (HI) and virus neutralisation (VN)) to FC1 and FC2 strains and in reducing clinical signs and viral shedding of EIV, and they provide equal protection to EIV strains currently circulating in the horse population.
2. Materials and Methods
2.1. Animals
This study was performed in 19 Norwegian Fjord horses (mares and geldings) aged between 4 and 4.5 years of age. The horses were obtained from two premises (UK and The Netherlands) and transported to a single study site (UK) where they arrived 3 weeks before EIV vaccination. Prior to transport, all horses were vaccinated against tetanus with Equilis Te (MSD Animal Health, The Netherlands) according to manufacturer’s instructions. All horses were confirmed to be seronegative for EIV based on screening of blood samples at the premises of origin and during acclimatisation at the study site. Upon arrival at the study site, horses were allocated to three rooms based on previous housing to ease mixing of horses and prevent unnecessary stress. The horses in each room were allocated randomly to a treatment group by the study investigator to ensure evenly distributed treatment groups in each of the three rooms. On day 31, the horses were moved into one room incorporating three pens to increase the space per animal, without changing the mix of treatment groups. Sample size (7 horses per vaccinated group and 5 control horses) was based on the European Pharmacopoeia criteria for EIV vaccine (inactivated), which requires at least 6 horses in the vaccinated group and at least 4 horses in the control group [
26].
2.2. Vaccines
Two commercially available vaccines were used: (i) Equilis Prequenza (MSD Animal Health; lots A118A01 and A115A02), a whole inactivated virus vaccine containing EIV strains A/equine/Newmarket/2/93 (European lineage) and A/equine/South Africa/4/03 (FC1 sublineage), adjuvanted with ISCOMatrix (purified saponin, cholesterol and phosphatidylcholine); (ii) ProteqFlu (Boehringer Ingelheim (formerly Merial)); lots L435907 and L443492), a modified vector vaccine containing two modified recombinant canarypox vectors expressing the HA gene of EIV strains A/equine/Ohio/03 (FC1 sublineage) or A/equine/Richmond/07 (FC2 sublineage), adjuvanted with carbomer. The investigational veterinary products (IVP) were used within the stated shelf-life of the product.
2.3. Viruses
Virus was grown in embryonated hens’ eggs, purified, and titrated as previously described [
27]. Briefly, 10–11 days old embryonated eggs were inoculated with virus and incubated for 3 days at 37 °C. Virus was harvested from the allantoic fluid, concentrated, and purified. To titrate the virus, a 10-fold serial dilution was made and used to inoculate 10–11 days old embryonated eggs. The allantoic fluid was harvested after incubation for 3 days at 37 °C and the EIV titre measured by haemagglutination assay (HA) using chicken red blood cells. The virus titre was calculated based on the number of EIV-positive and -negative eggs for each sample dilution and expressed as log
10 EID50/mL.
EIV H3N8 A/equi-2/Wexford/14 (FC2 sublineage) was used for the experimental infection (challenge) and kindly provided by the Irish Equine Centre (Ireland). EIV H3N8 A/equine/South Africa/4/03 (FC1 sublineage) and A/equine/Meath/07 (FC2 sublineage) were used as single radial haemolysis (SRH) antigens. EIV H3N8 strains A/equine/Ohio/03 (FC1 sublineage), A/equine/South Africa/4/03 (FC1 sublineage), A/equine/Shropshire/10 (FC2 sublineage), and A/equine/Richmond/1/07 (FC2 sublineage) were used as HI and virus-neutralising (VN) antigens.
2.4. Vaccination and Challenge Protocol
Horses were allocated randomly into three treatment groups. Group 1 (n = 5) was the unvaccinated control group, group 2 (n = 7) received Equilis Prequenza and group 3 (n = 7) received ProteqFlu. Each horse from groups 2 and 3 received 1 mL (single dose) of vaccine by deep intramuscular injection (21G × 1, 0.8 × 40 mm needle) in the left neck on day 0 (V1) and in the right neck on day 28 (V2). On day 148, all horses were challenged with EIV H3N8 A/equi-2/Wexford/14 (FC2 sublineage). Each horse received 2 mL of phosphate-buffered saline-diluted allantoic fluid containing a target titre of 107 egg infectious dose 50 (EID50) challenge virus administered intranasally by individual nebulisation using the Flexineb® portable equine nebuliser (Nortev, Ireland). All these procedures were conducted at Drayton Animal Health, Stratford-upon-Avon, UK.
To ensure complete blinding, the vaccines were labelled as A and B and were not identified by their trade names. The unblinding information was only available to one individual from MSD Animal Health, who was blinded to the allocation of the animals to treatment groups. The preparation, documentation, and administration of vaccinations was performed by the study investigator who was not involved in clinical observations nor laboratory or data analysis. All of the personnel involved in clinical observations or laboratory or data analysis were blinded to the allocation of the animals to treatment groups. Unblinding took place after all of the samples and data had been analysed so that each of the treatment groups could be linked to the vaccine administered and individual animals to their treatment group.
This study was performed under the supervision of the investigator in compliance with the authorised study protocol (and amendments), standard operating procedures (SOPs), and Home Office License according to the Animal (Scientific Procedures) Act 1986.
2.5. Collection of Samples
Blood samples were collected from all animals on days −13/−12 (during acclimatisation), 0 (prior to vaccination), 9, 28 (prior to vaccination), 51, 86, 121, 148 (prior to challenge), 155, and 162. Blood samples were transported to the laboratory for separation of serum and stored at −15°C or below until testing. No blood sample could be collected from horse #2914166 on days 0 and 9 and horse #2904506 on day 9.
Nasal swab samples were collected from all horses on days 0 and 28 (prior to vaccination), day 147 (prior to challenge) and daily from day 149 until day 162. Swabs were taken from only one nostril per day and nostrils alternated between each sampling occasion. Transport medium (phosphate-buffered saline with foetal calf serum and antibiotics/antifungals) was added to each nasal swab and swab samples were stored at ≤−70 °C until testing. No nasal swab could be collected from horse #2910058 on days 157, 159, and 160.
2.6. Serology
Serum was analysed at the Irish Equine Centre (Ireland) for EIV antibodies to FC1 and FC2 using an SRH assay as previously described [
28,
29,
30]. Serum was analysed at MSD Animal Health (The Netherlands) for EIV antibodies to FC1 and FC2 using a HI assay and for EIV neutralising antibodies using a VN assay as previously described [
30,
31].
For SRH, a significant rise in antibody titre was defined as an increase in the SRH of 25 mm
2 or 50%, whichever was smaller, between paired serum samples. A poor responder was defined as a horse that did not mount a mean H3N8 SRH antibody response of >25 mm
2 post-vaccination. HI titres were expressed as log
2 values of the reciprocal of the highest serum dilution that gave complete inhibition of haemagglutination. A titre of >4 was regarded as significant and a two-fold increase in the titre indicative of seroconversion. A titre of <4 was converted to 3 for statistical analysis and graphical presentation. VN titres were expressed as log
2 values of the reciprocal of the highest serum dilution that gave complete virus neutralisation. A titre of <2 was converted to 1 for statistical analysis and graphical presentation. Raw data are provided in
Tables S1–S3.
2.7. Clinical Observations
Clinical observations, including rectal temperature, cough, nasal discharge, ocular discharge, anorexia, depression, and dyspnoea, were evaluated, recorded, and analysed using the scoring system listed in
Table 1, and conducted by trained personnel who observed the horses once daily from day 146 to 162. On day 148, clinical observations were carried out prior to challenge. The horses were observed together for 20 min for cough (first coughing score), depression, and dyspnoea. After 20 min, the horses were led through a race one by one. In the race, laryngeal palpation was performed to assess cough (second coughing score), nasal and ocular discharges were assessed, and anorexia was assessed by offering concentrate feed in a bucket. A digital thermometer was used to measure rectal temperature. Rectal temperature could not be measured from horse #2907628 on day 0. Raw data are provided in
Table S4A, Table S4B, Table S4C, Table S4D and Table S4E.
2.8. Viral Shedding
Nasal swab samples were analysed at MSD Animal Health (The Netherlands) to determine shedding of live EIV after challenge using a virus titration assay in embryonated SPF hens’ eggs [
28]. Firstly, all swab samples were screened for presence of live virus and, thereafter, the amount of live virus was quantified only in the virus-positive samples.
Furthermore, nasal swab samples were analysed at the Irish Equine Centre (Ireland) to detect EIV genomic material after challenge by analysing viral RNA expression using EIV qPCR, as previously described [
32]. Nasal swabs with a cycle threshold (Ct) value ≤40 were classified as positive for viral RNA.
2.9. Data and Statistical Analysis
Vaccine efficacy was assessed based on the Equine Influenza Vaccine (Inactivated) 0249 Monograph [
26]. According to this Monograph, the assessment of efficacy is based on vaccinated horses showing no more than mild clinical signs and control horses showing characteristic clinical signs of EIV infection. The average number of days on which the virus is shed, and the respective virus titres should be significantly lower in the vaccinated horses than in the control horses.
For the statistical analysis, descriptive statistics (i.e., arithmetic mean, plotting the data, etc.) were used to summarise trends, but no significance testing (i.e., p-values) was conducted. In addition, inferential statistics (i.e., regression modelling)―a statistical method where a small but representative sample is used to describe the characteristics of a larger population―was used to show whether there were significant differences between vaccinated and control animals after challenge (i.e., to evaluate vaccine efficacy to EIV challenge) using the parameters described below.
Antibody titres after vaccination were plotted, and the trends summarised using descriptive analysis. Subsequently, antibody titres after challenge were compared between groups using a linear mixed model analysis of variance (ANOVA) for repeated measures (SAS procedure proc mixed) using treatment group, days post-challenge (dpc), and their interaction as explanatory variables. In addition, the seroconversion trend in antibody titres after challenge were investigated for all three groups and compared between groups. The first step was fitting a linear model of the log2 antibody titre as response variable and time (i.e., dpc), treatment group, and their interaction as explanatory variables (SAS procedure proc glm). Subsequently, it was investigated whether it could be assumed that the slope was common to the vaccinated and control groups (i.e., the antibody titre had a similar seroconversion trend per group) or were treatment-specific (i.e., the antibody titre had a different seroconversion trend per group) by looking at whether the interaction effect was significant (treatment-specific slope) or not (common slope).
Total clinical score data (the overall of the clinical score plus the rectal temperature score) and clinical score data over time after challenge were plotted and analysed using generalised estimating equations (GEE) using a cumulative logit model, accounting for the correlation in the repeated measures of a horse (SAS procedure proc genmod with repeated statement). The response variable was included as an ordinal response in the GEE model using a multinomial distribution with a cumulative logic link function.
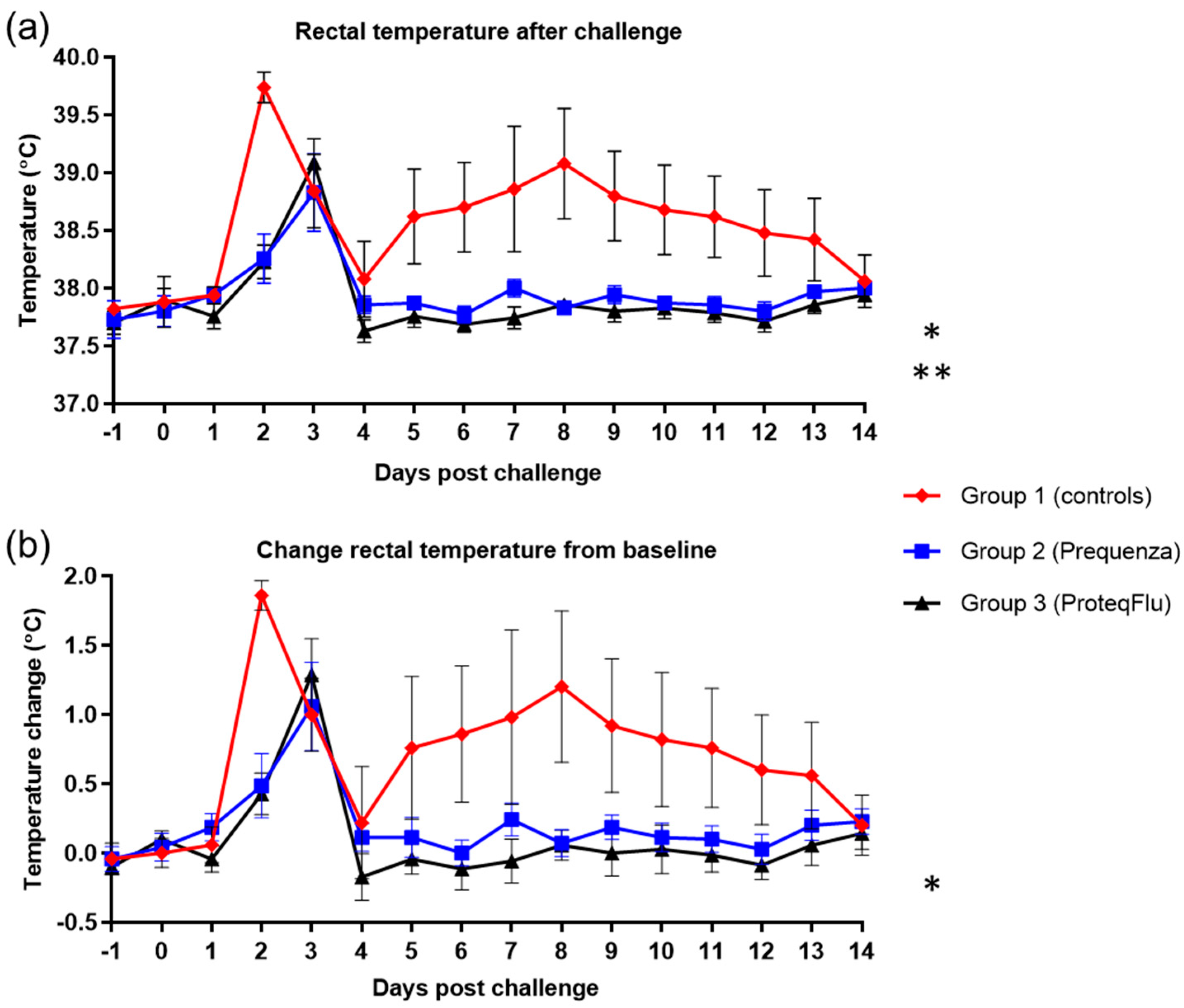
Rectal temperature data after challenge were plotted and evaluated separately from the total clinical score, and for this, the mean of the pre-challenge values was taken as baseline. Temperature data over time were evaluated using a linear mixed statistical model for repeated measures, including baseline as covariate (SAS procedure proc mixed). In addition, the difference in peak rectal temperature from baseline was evaluated using ANOVA.
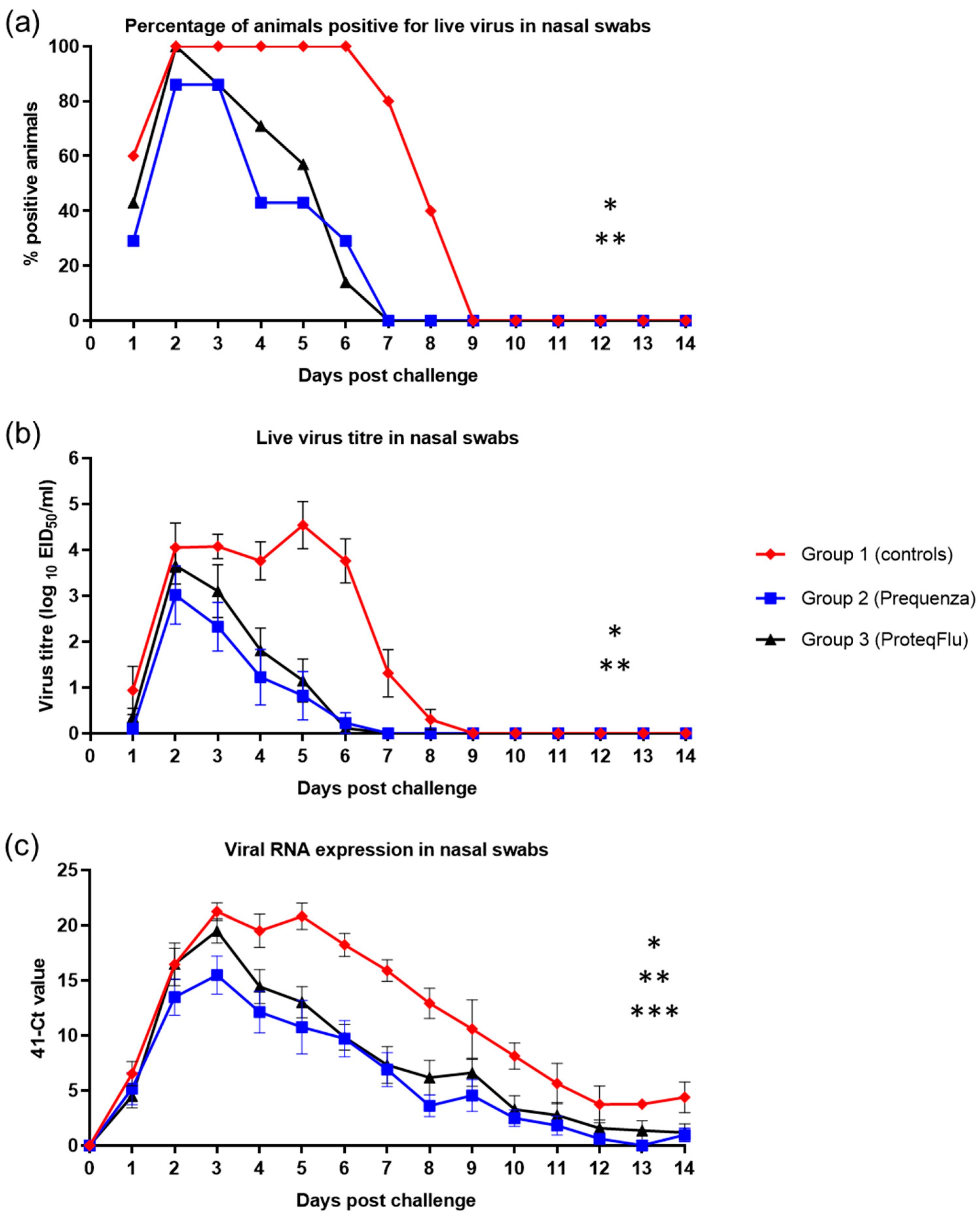
Virus isolation data were plotted and then categorised as positive or negative and analysed as a repeated measure (pos/neg) using GEE methodology. Secondly, the viral shedding (titre in nasal swabs after challenge) was analysed by a linear mixed model ANOVA for repeated measures (SAS procedure proc mixed). Furthermore, the duration of virus shedding in days was counted and analysed by ANOVA. Nasal swab virus qPCR results for viral shedding was plotted and analysed by a linear mixed model ANOVA for repeated measures (SAS procedure proc mixed).
For all inferential statistical analyses, the level of significance α was set at 0.05 and tests were two-sided. A statistical trend approaching significance was detected when the p-values were between 0.05 and 0.1. Statistical software package SAS V9.4 (SAS Institute Inc., Cary, NC, USA) was used.
4. Discussion
The present study evaluated the efficacy of two commercially available EIV vaccines, an ISCOMatrix-adjuvanted, whole inactivated virus vaccine (Equilis Prequenza, group 2) or a carbomer-adjuvanted, modified vector vaccine (ProteqFlu, group 3). Serological responses, clinical signs, and viral shedding were analysed after experimental infection with a recent FC2 EI field strain at 4.5 months after the primary vaccination course (V1 and V2) and compared with unvaccinated control horses. The aim was to provide insight into the impact of vaccine type and minor antigenic differences between vaccine and field strains on vaccine efficacy, after challenge during the most susceptible period (the immunity gap). EI vaccines reduce clinical signs and viral shedding after EIV infection. In horses that have been vaccinated or infected previously, clinical signs are usually mild or inapparent. After vaccination, high antibody titres at least initially are often enough to provide protection, even where the vaccine strain differs from the current field strain. However, there have also been reports of some appropriately vaccinated horses that have developed clinical signs of influenza similar in magnitude and duration to those described for non-vaccinated horses (vaccination breakdown) following the appearance of a new field strain that differs antigenically (antigenic drift) from the EI strain(s) present in the commercially available vaccines [
32,
33,
34,
35].
It was hypothesised that due to the antigenic differences between the FC2 EI challenge strain and the virus strain(s) in the vaccine used for group 2 (Equilis Prequenza), a lower level of protection (more marked fever, more pronounced clinical signs, more marked and prolonged viral shedding, and more marked seroconversion following experimental infection) might be seen. However, following experimental infection, there were no significant differences in rectal temperature and clinical signs between the vaccinated groups and these were significantly lower than in unvaccinated controls. In addition, viral shedding was significantly reduced in both vaccinated groups compared to the unvaccinated controls. While there were differences between the two vaccinated groups in the number of horses that shed virus and the amount of viral RNA detected, there were no significant differences in viral shedding between these groups. Viable influenza virus was isolated (using embryonated hens’ eggs) from six out of seven of the horses in group 2 (Equilis Prequenza, which contains a FC1 and an European lineage strain and thus complies partly with the current OIE expert surveillance panel recommendations), and from all seven of the horses in group 3 (ProteqFlu, which contains FC1 and 2 lineage strains and thus complies fully with those recommendations). In addition, the clinical score and viral shedding pattern tended to be lower in group 2 than in group 3. There were also differences between the three groups in the excretion of EIV genomic material (PCR): in groups 1 (controls) and 3 (ProteqFlu), horses shed EIV genomic material up to 30 dpc, while in group 2 (Equilis Prequenza) horses no longer shed EIV genomic material by 14 dpc. Moreover, significantly less EIV material was shed by group 2 than by group 3. This was unexpected since group 2 was vaccinated with a vaccine that contains a FC1 and a European lineage strain and no FC2 lineage strain.
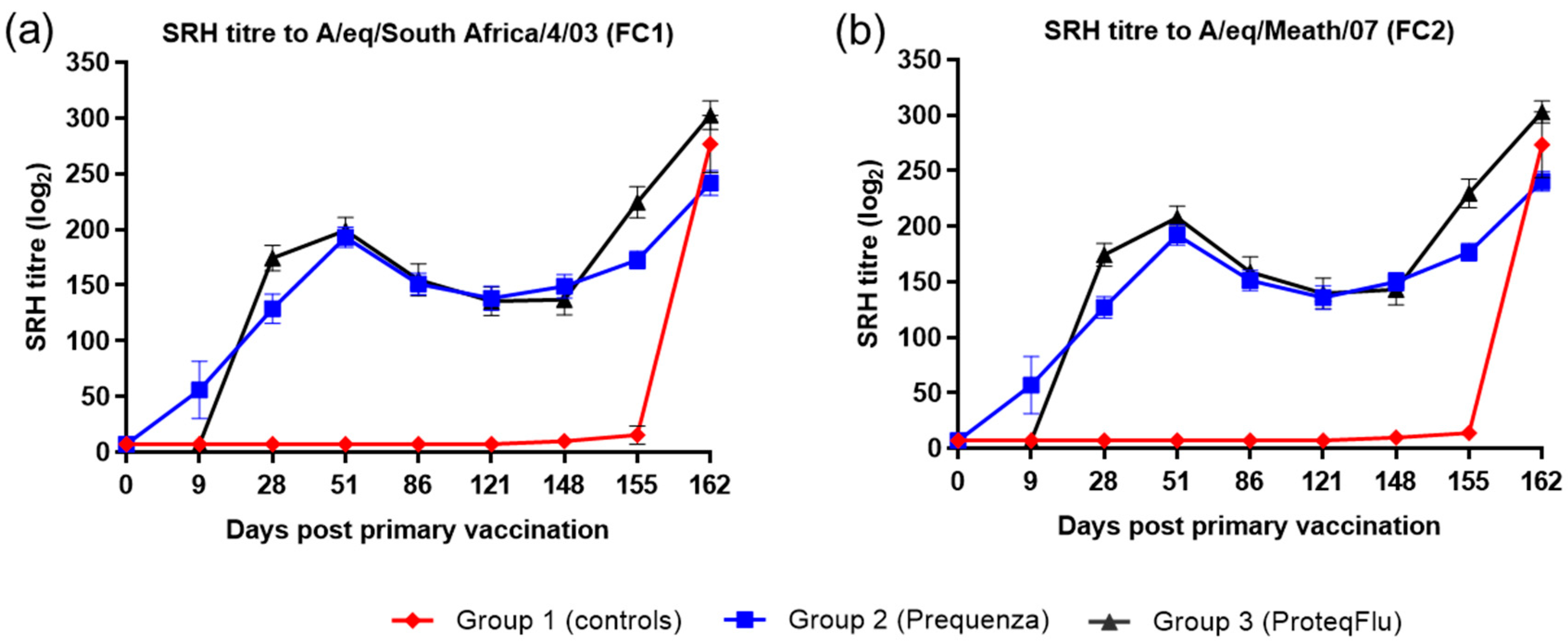
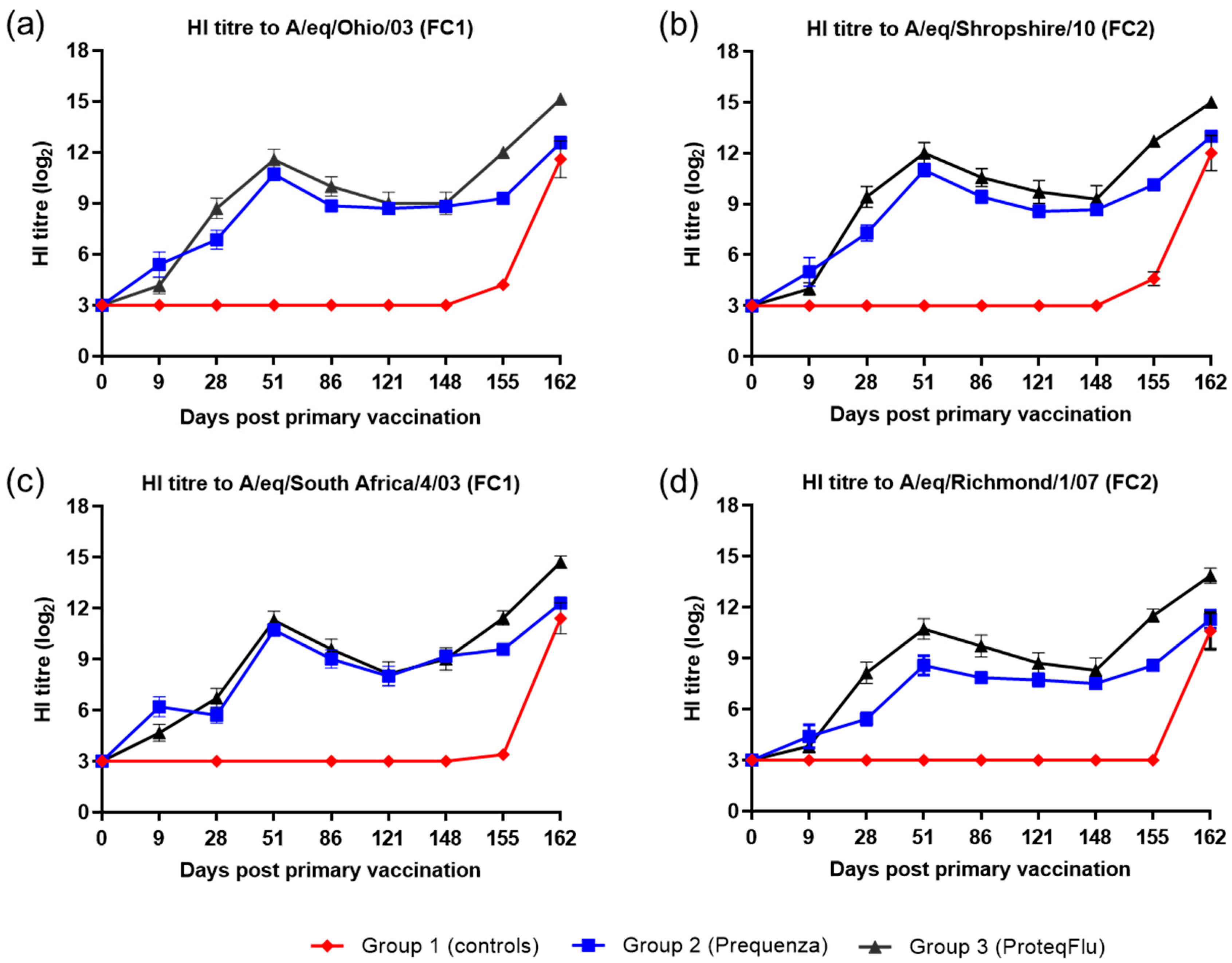
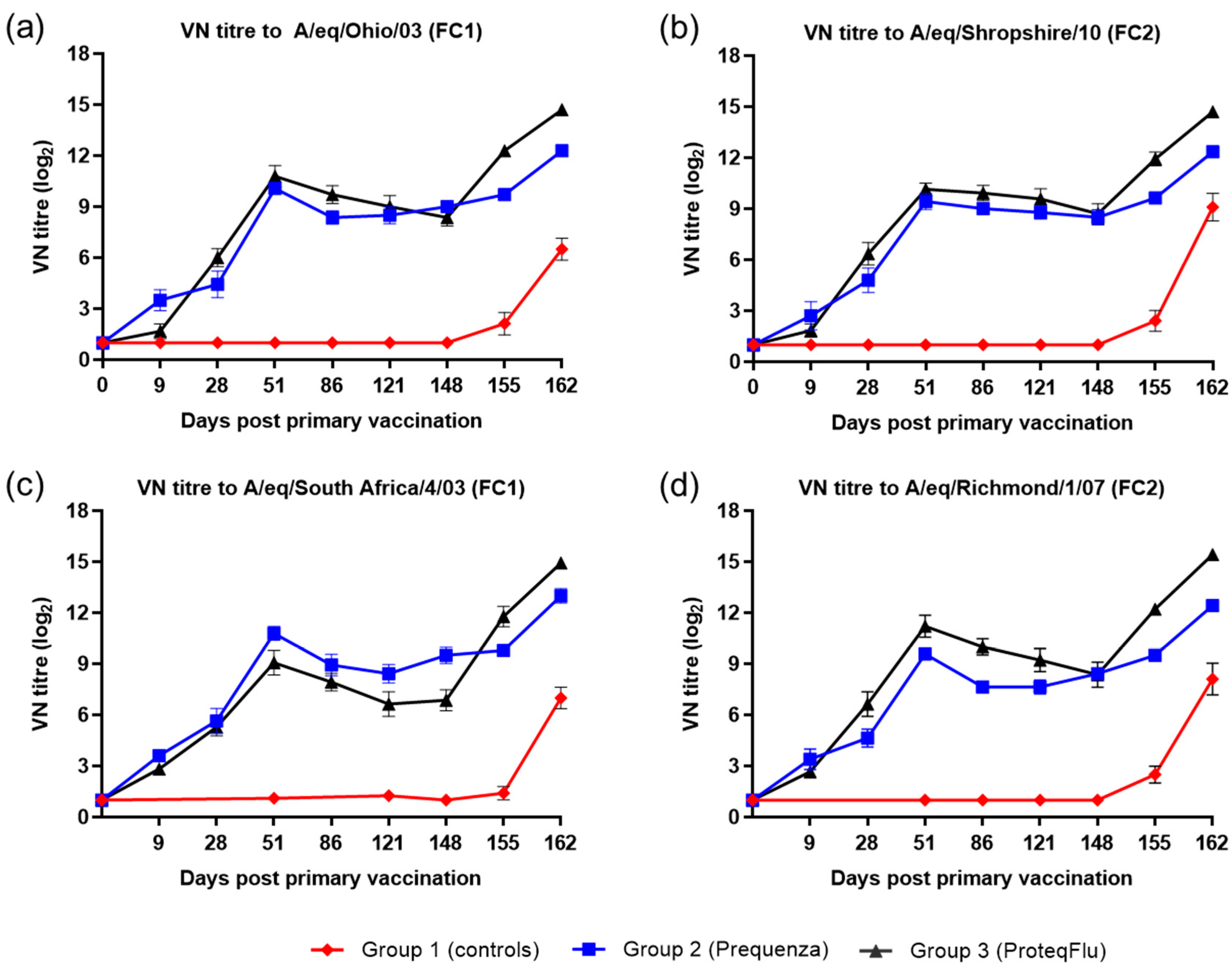
The serological response of all horses after vaccination and virus challenge was monitored since antibody titres measured by SRH and HI assays provide a correlate of protection against EI infection [
4]. In addition, a VN assay, which identifies antibodies that inhibit the entry of virus into cells and virus replication, was used and has also been shown to be suitable for monitoring EIV serology after vaccination and/or infection [
36,
37]. These three serological assays are based on different principles but yielded comparable results, in line with previous findings [
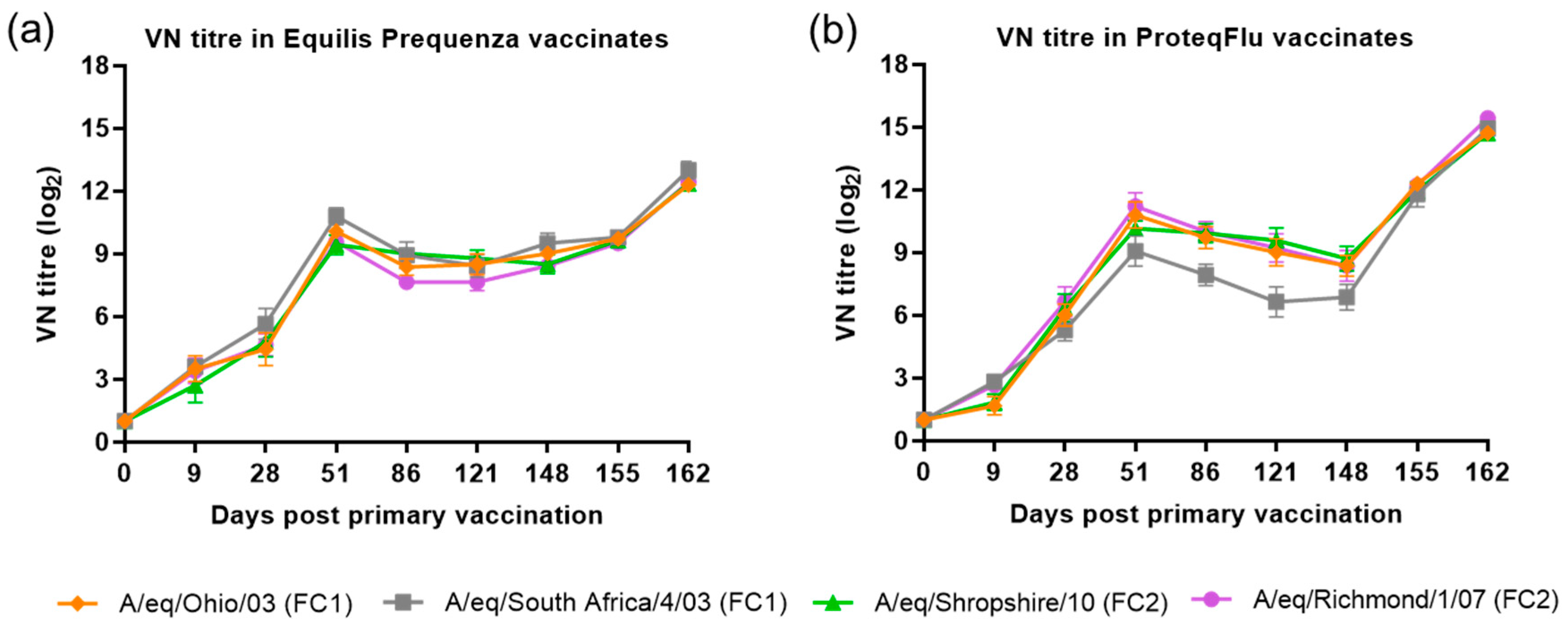
38]. Before challenge, both the serological profile and titres of the SRH and HI assay were comparable in the two vaccinated groups, irrespective of which FC sublineage strain was used in the assay. The serological profile of the VN titres in both vaccinated groups was also comparable to the SRH and HI serological profiles with two exceptions. VN titres to A/equine/Richmond/07 (FC2) on days 28–121 were lower in group 2 (Equilis Prequenza) than in group 3 (ProteqFlu) and VN titres to A/equine/South Africa/4/03 (FC1) on days 51–148 were higher in group 2 than in group 3. It has been reported previously that strain-specific antibodies give higher titres in a VN assay [
39]. This likely explains the differences in VN titre that were observed, since A/equine/Richmond/07 is present in ProteqFlu (group 3) while A/equine/South Africa/4/03 is present in Equilis Prequenza (group 2).
The level of seroconversion after challenge indicates the amount of virus present, with a higher amount of virus triggering a more marked antibody response. After experimental infection, the seroconversion measured in the SRH, HI and VN assays was significantly higher in group 3 (ProteqFlu) than in group 2 (Equilis Prequenza). The increase in antibody titres (slope) after challenge was similar to the control group in group 3 but significantly lower than in the control group in group 2. Moreover, antibody titres after challenge (average of 155 and 162 dpv; 7 and 14 dpc) were significantly lower in group 2 than in group 3. This may suggest that the response to vaccination and/or immune response was likely better in group 2 than in group 3 since the same amount of virus was used to challenge each horse but there was a less pronounced serological response to challenge in group 2 than in group 3. Another unexpected finding was that VN results for group 2 were similar for all FC1 and FC2 strains, although no FC2 sublineage strain is present in the vaccine (Equilis Prequenza) used. In contrast, the vaccine used for group 3 (ProteqFlu) contains both FC1 and FC2 sublineage strains, and while similar VN results were seen for FC2 sublineage strains and FC1 A/equine/Ohio/03, this was not the case for A/equine/South Africa/4/03 (FC1). This may be due to the difference in composition of the two vaccines: one is a whole inactivated virus vaccine containing the whole virus including the conserved internal viral proteins, while the other is a vectored vaccine containing only the HA protein, which is the most immunogenic but also the most variable viral protein. A vaccine that contains only a viral subunit will be more vulnerable to antigenic changes than a whole virus vaccine. The role of antigenic changes and how these relate to vaccine efficacy and cross-reactivity will be investigated in future studies.
Humoral (utilising circulating antibodies) and cell-mediated (utilising white blood cells) immunity are the two key components of the immune system. Clinical signs of EIV infection are related to the duration and level of host chemotactic, proinflammatory and antiviral cytokines rather than virus replication. Most EIV antibodies (induced by either natural infection or vaccination) are directed towards the major viral surface structural proteins haemagglutinin (HA) and neuraminidase (NA), neutralise virus before infection occurs and inhibit the release of virus from cells after replication, thus preventing clinical signs and virus transmission. Cell-mediated immunity is also important in the control of virus infection as this is important for elimination of virus-infected cells. Cell-mediated immunity has been reported previously in EI-vaccinated horses [
12,
27,
40]. The OIE uses a number of different antigenic analyses as a basis for its vaccine update recommendations, including the detection of changes in HA based on HI tests carried out using ferret antisera. Antisera raised in ferrets is used for this test because this species mounts a strain-specific response to influenza that picks up even minor antigenic variations. We have demonstrated that antisera raised in horses against several EIV strains cross react to a far greater extent than those raised in ferrets. In other words, the horse is able to cope with some minor antigenic variation of its own influenza virus, whereas the ferret recognises this change as significant. Our data show that even where HA from a field virus differs antigenically (antigenic drift) from that of the virus strain(s) present in a vaccine, this does not necessarily affect vaccine efficacy. Since both humoral and cell-mediated immunity play a role in protection against EIV infection, there is a need for additional methods to assess vaccine efficacy against new field strains in horses. For example, when the OIE expert surveillance panel recommends updating an EIV vaccine strain or strains, as an alternative to simply updating the strains, the existing commercially available vaccines could be evaluated against these field strains in horses to demonstrate that efficacy―including reduction of clinical signs and viral shedding―is maintained. This evaluation should be performed during the most sensitive period, the so-called immunity gap that occurs a few months after the primary vaccination course (V1 and V2).
Several different approaches to EIV vaccine development have been taken and various EIV strains have been used in different EIV vaccines over the years. Currently, three EIV vaccines are on the market in Europe: an ISCOMatrix-adjuvanted, whole inactivated virus vaccine, an ISCOM-adjuvanted inactivated virus subunit (specific viral proteins) vaccine, and a carbomer-adjuvanted modified viral vector vaccine. It is conceivable that differences in the composition of these vaccines and the mechanism in which they induce immunity could easily lead to differences in vaccine efficacy that are far more significant than the differences from simply updating the strain in a different type of vaccine. The whole inactivated virus vaccine contains all viral proteins while the modified viral vector vaccine contains only the HA gene. A viral vector vaccine triggers the immune response in a similar fashion to a single cycle of a live influenza virus infection while a whole inactivated virus vaccine needs adjuvant to trigger the immune response. However, Geeraedts et al. (2008) [
41] found that the quality and quantity of the immune response triggered by a whole inactivated influenza virus vaccine and a live influenza virus was similar, suggesting that the integrity of the virus particle structure is more important for induction of an effective immune response than presence or absence of virus replication. While antibody titre is a correlate of protection against influenza virus, cell-mediated immunity also plays a role in protection, especially in cross-protection against heterologous virus strains. In mice, whole inactivated influenza virus vaccines have been shown to induce higher HI and VN titres than subunit vaccines, stimulate dendritic cells, and induce a Th1 response, providing better protection against infection [
41]. Stimulation of murine dendritic cells with a non-adjuvanted, whole inactivated influenza virus vaccine in vitro resulted in a response similar to stimulation with live virus, but a non-adjuvanted, inactivated virus subunit vaccine did not [
42]. In addition, whole inactivated influenza virus vaccines, but not subunit vaccines, have been shown to protect mice against lethal heterologous virus challenge through cell-mediated immunity to conserved internal influenza virus proteins [
43].
Many variables play a role in the efficacy of different whole inactivated influenza virus vaccines and it is therefore important that the efficacy and safety of these vaccines be compared. For example, the method used to inactivate a whole virus can also affect vaccine efficacy due to the alterations it causes in the structure of the virus. For example, the qualitative immune response is less following inactivation with formalin than inactivation with beta-propiolactone in mice [
43]. In addition, the choice of the adjuvant for a whole inactivated influenza virus vaccine is crucial because the induction of an appropriate immune response is important and differs for each adjuvant [
4,
44,
45]. While a highly reactogenic adjuvant can lead to good vaccine efficacy, this might compromise vaccine safety and the opposite may be true for a poorly reactogenic adjuvant.

 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}