Contrasting Immunomodulatory Effects of Probiotic and Pathogenic Bacteria on Eastern Oyster, Crassostrea Virginica, Larvae



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures
2.2. Oyster Larvae
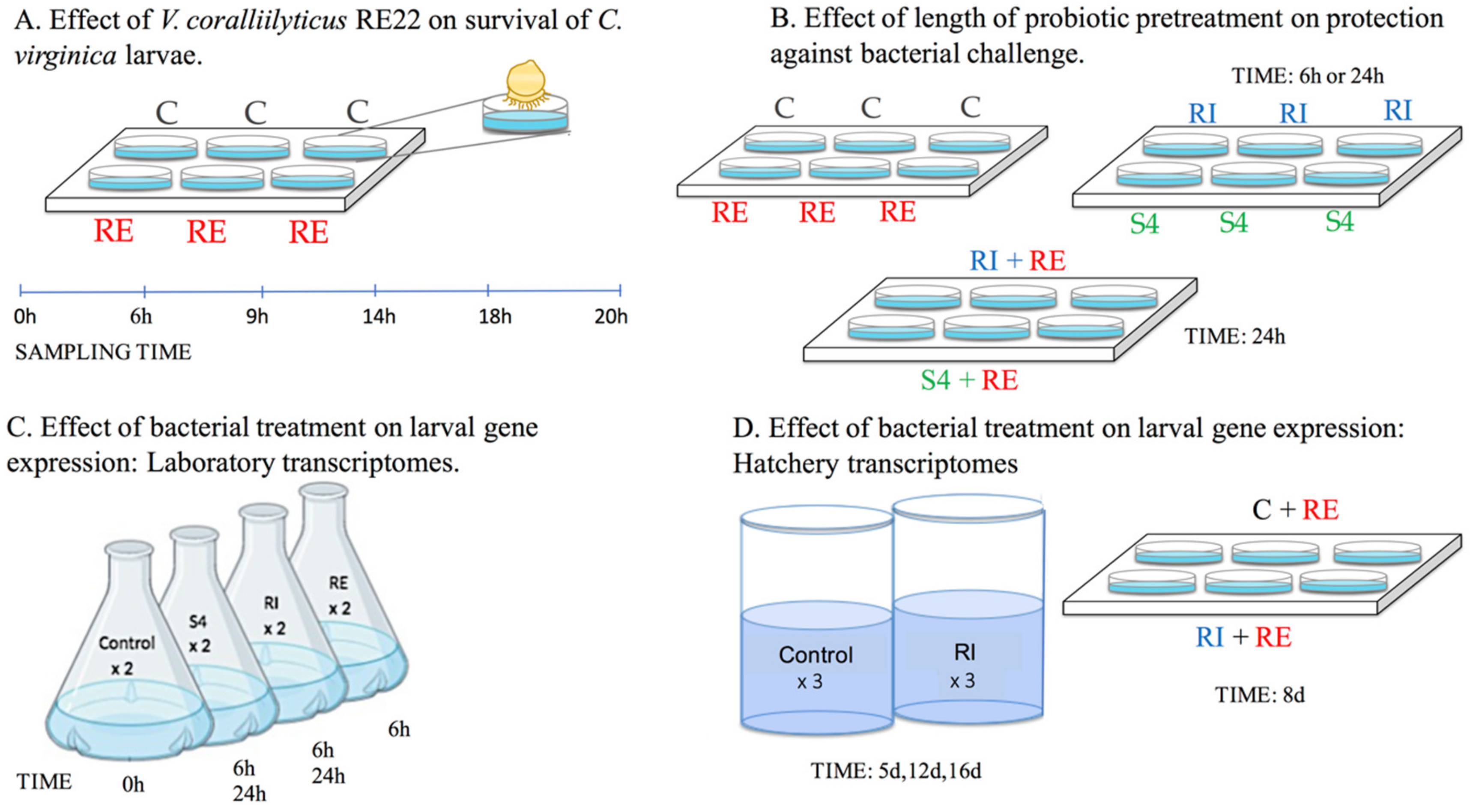
2.3. Effect of V. coralliilyticus RE22 on Survival of C. virginica Larvae
2.4. Effect of Length of Probiotic Pretreatment on Protection against Pathogen Challenge
2.5. Effect of Bacterial Treatment on Larval Gene Expression: Laboratory Transcriptomes
2.6. Effect of Bacterial Treatment on Larval Gene Expression: Hatchery Transcriptomes
2.7. RNA Extraction, cDNA Prep, and Sequencing
2.8. Assembly, Annotation, and Differential Expression Analysis
3. Results
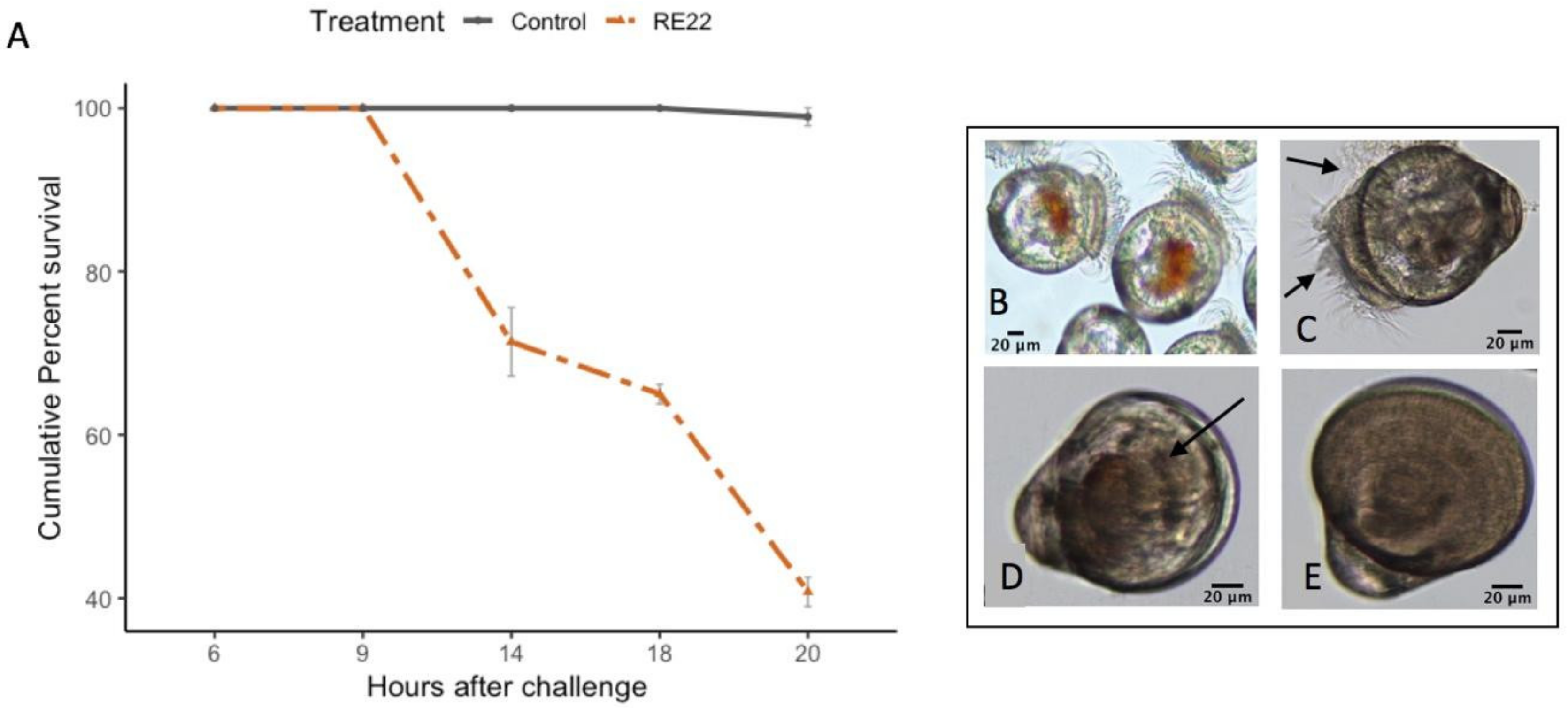
3.1. Rapid Mortality in C. virginica Larvae after Challenge with Pathogen V. coralliilyticus RE22
3.2. Pretreatment of Larvae with Probionts RI and S4 for 6 or 24 h before Challenge Conferred Protection against Pathogen RE22
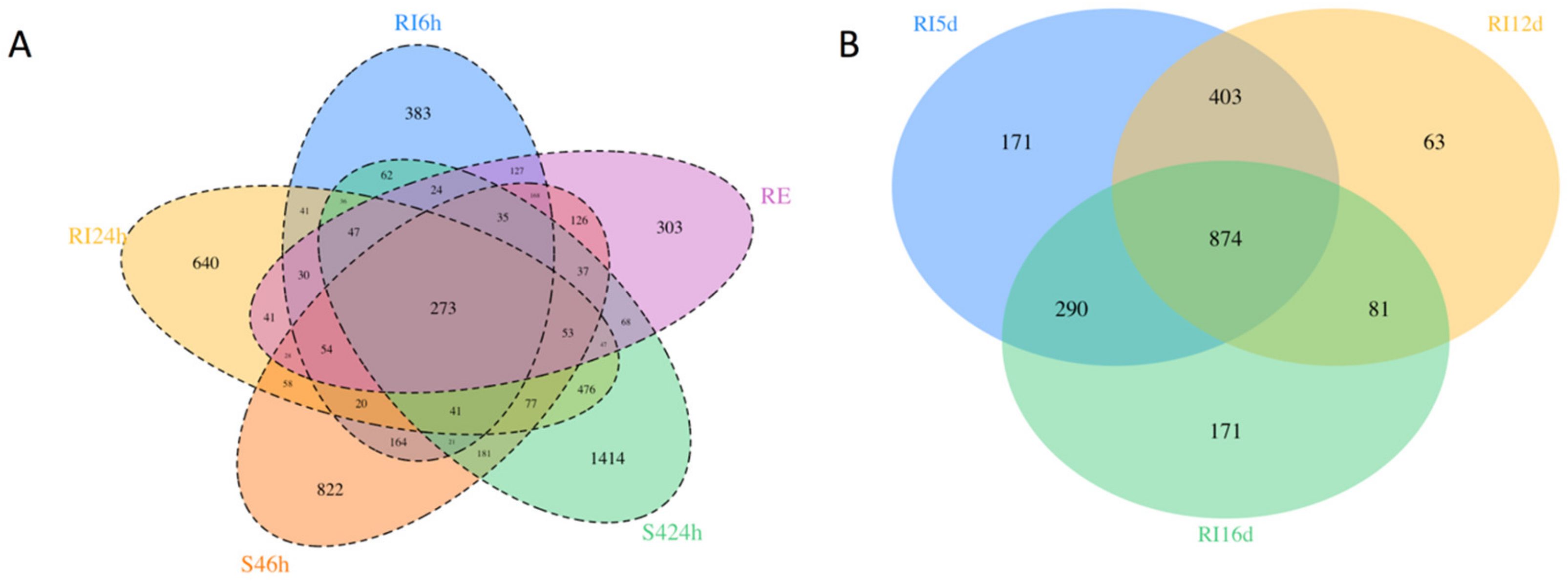
3.3. Transcriptome and Differential Gene Expression Analysis Statistics
3.4. Gene Ontology Enrichment Analysis Showed an Enrichment in Genes Involved in Metabolism in Larvae Treated with RE22, but Not in Probiotic Treated Larvae
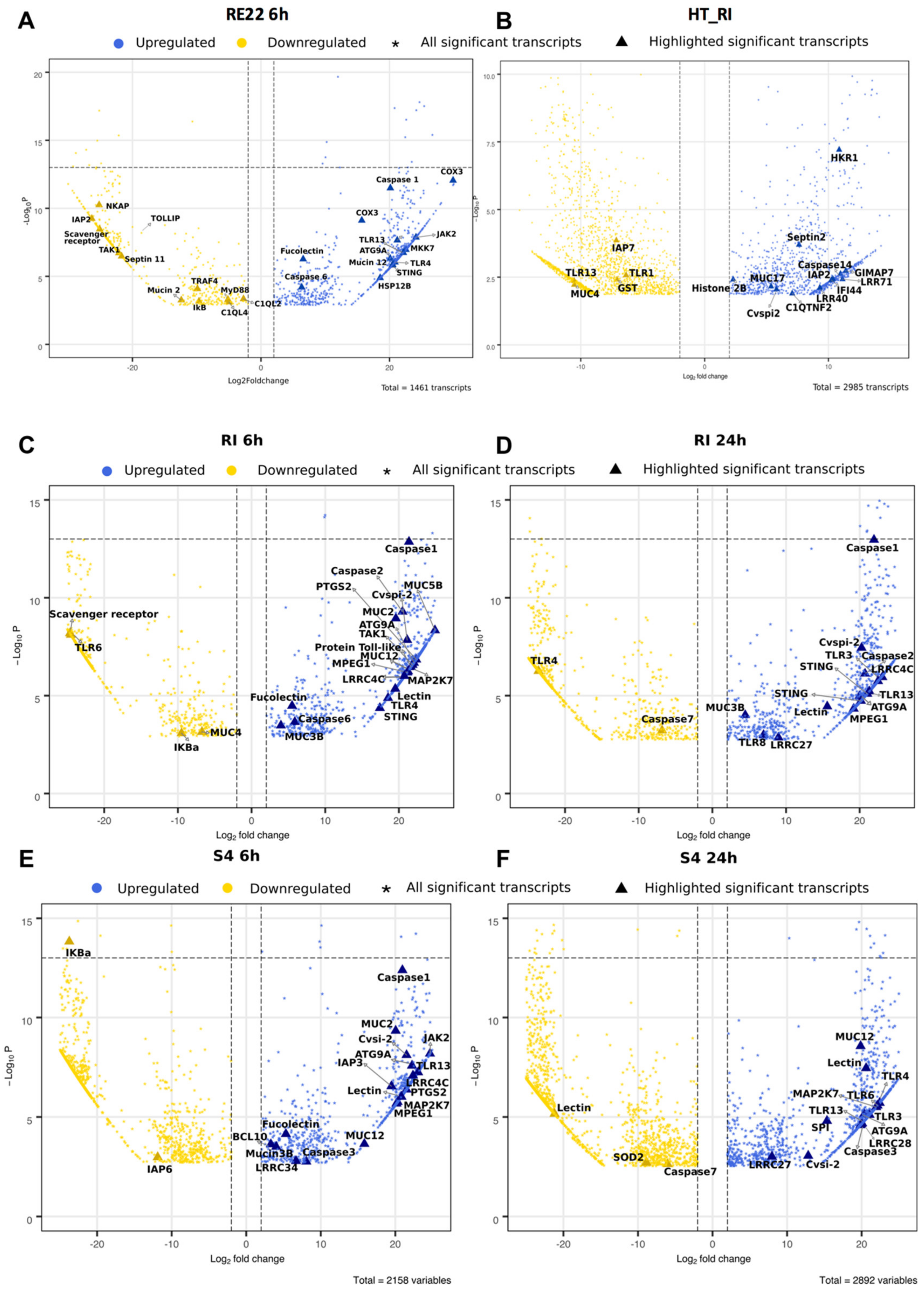
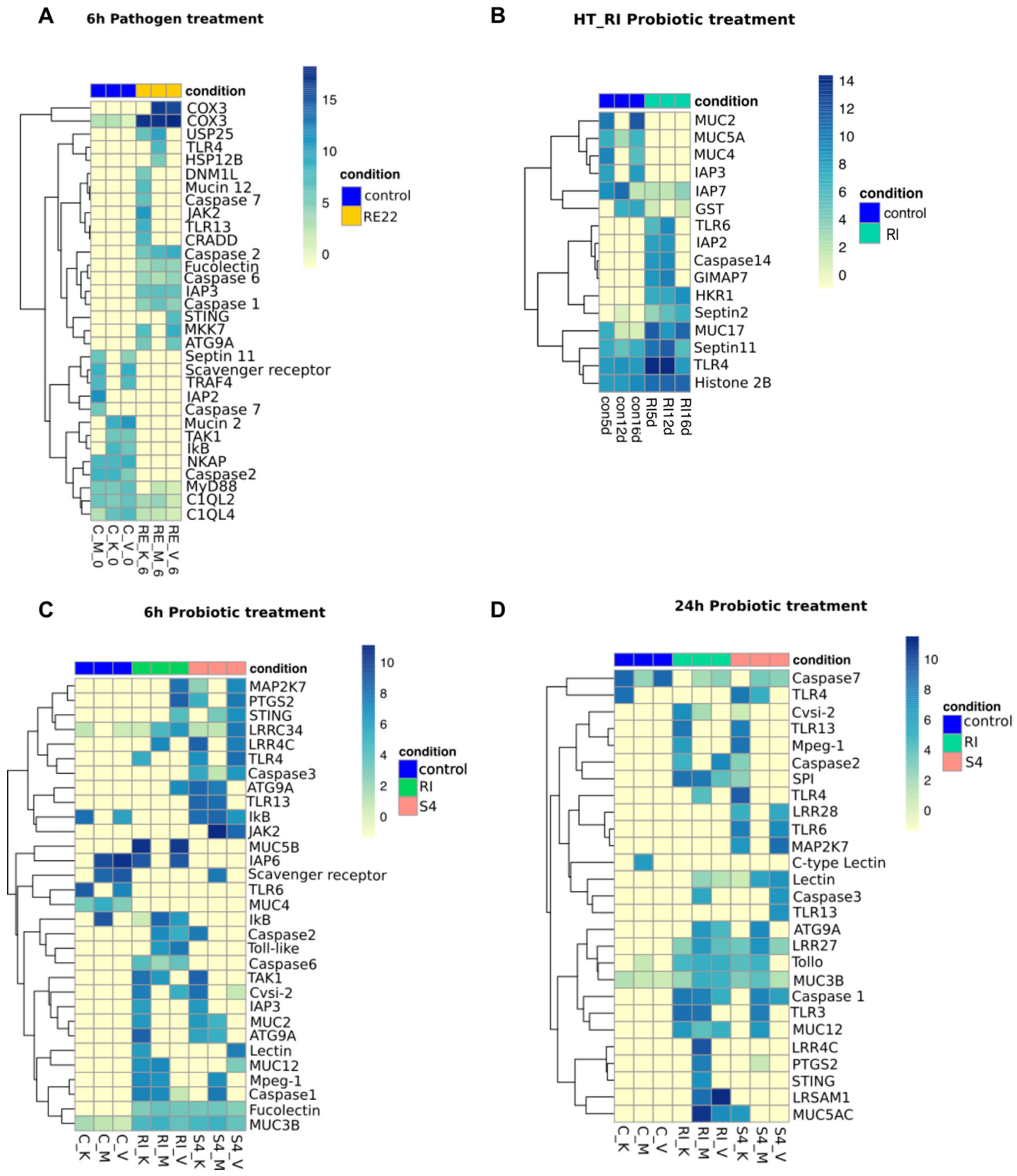
3.5. Contrasting Effects of Treatment with Probionts RI and S4 and Pathogen RE22 on Larval Immunity
3.5.1. Pattern Recognition Receptors (PRRs)
3.5.2. Immune Signaling Pathways
3.5.3. Immune Effectors
3.5.4. Other Immune Molecules
3.5.5. Gene Expression Patterns Show Signs of Metabolic Demand and Stress in RE22 Exposed Larvae
4. Discussion
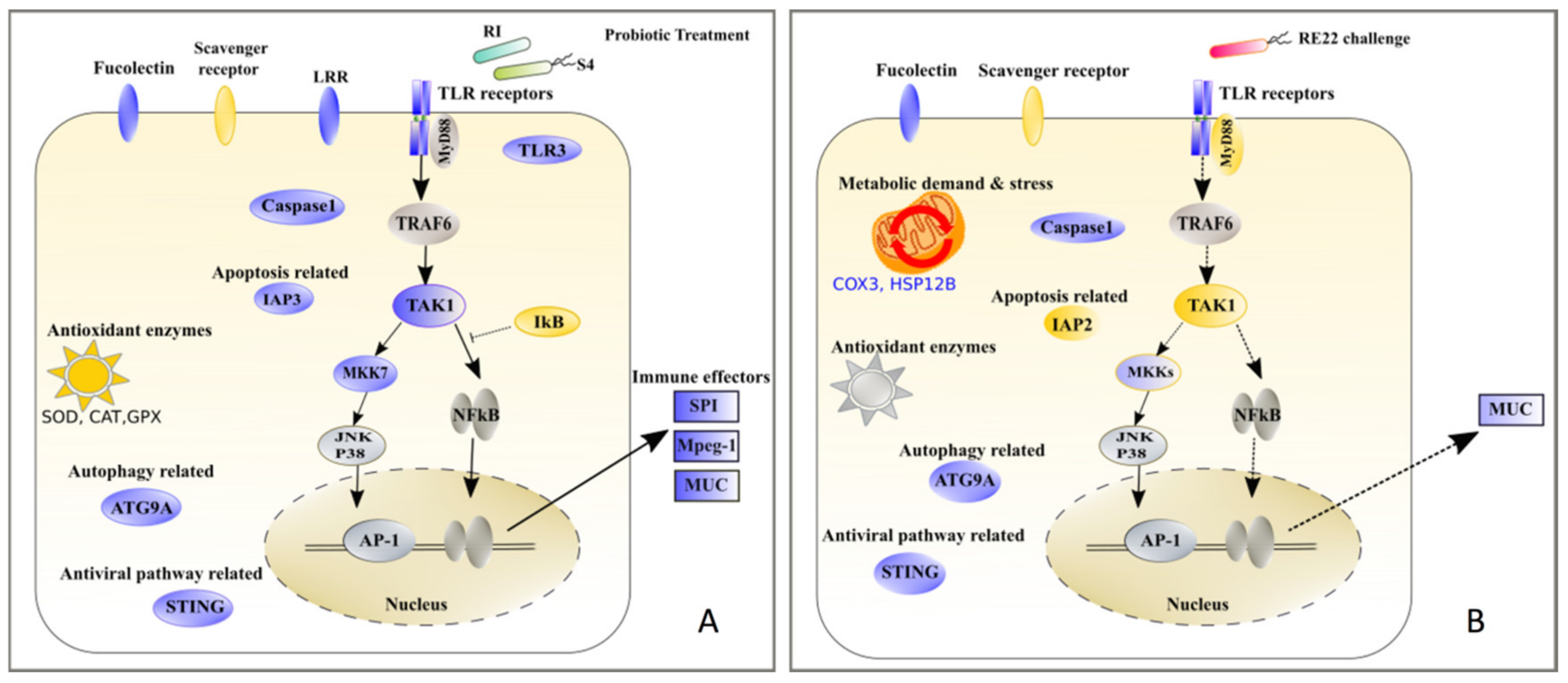
4.1. Pathogenesis of RE22 Vibriosis in Eastern Oyster Larvae: Role of Immunosuppression and Metabolic Stress
4.2. Protection against RE22 Challenge in Probiotic-Treated Eastern Oyster Larvae: Role of Immunomodulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer International Publishing: Cham, Switzerland, 2016; ISBN 9783319326733. [Google Scholar]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Primers 2018, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Roux, F.; Wegner, K.; Polz, M. Oysters and vibrios as a model for disease dynamics in wild animals. Trends Microbiol. 2016, 24, 568–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafferty, K.D. Marine infectious disease ecology. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 473–496. [Google Scholar] [CrossRef] [Green Version]
- Mohamad, N.; Amal, M.N.A.; Yasin, I.S.; Zamri Saad, M.; Nasruddin, N.S.; Al-saari, N.; Mino, S.; Sawabe, T. Vibriosis in cultured marine fishes: A review. Aquaculture 2019, 512, 734289. [Google Scholar] [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.; Hélaouët, P.; Edwards, M.; Höfle, M.; Brettar, I.; Colwell, R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef] [Green Version]
- Dubert, J.; Barja, J.L.; Romalde, J.L. New insights into pathogenic Vibrios affecting bivalves in hatcheries: Present and future prospects. Front. Microbiol. 2017, 08, 762. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2018: Meeting the Sustainable Development Goals; Food and Agriculture Organization: Rome, Italy, 2018; ISBN 9789251305621. [Google Scholar]
- Gradoville, M.R.; Crump, B.C.; Häse, C.C.; White, A.E. Environmental controls of oyster-pathogenic Vibrio spp. in Oregon estuaries and a shellfish hatchery. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Roux, F.; Wegner, K.; Craig, B.-A.; Vezzulli, L.; Osorio, C.; Amaro, C.; Ritchie, J.; Defoirdt, T.; Delphine, D.-G.; Blokesch, M.; et al. The emergence of Vibrio pathogens in Europe: Ecology, evolution, and pathogenesis (Paris, 11–12th March 2015). Front. Microbiol. 2015, 6, 830. [Google Scholar] [CrossRef] [Green Version]
- Travers, M.-A.; Boettcher Miller, K.; Roque, A.; Friedman, C.S. Bacterial diseases in marine bivalves. J. Invertebr. Pathol. 2015, 131, 11–31. [Google Scholar] [CrossRef] [Green Version]
- Elston, R.A.; Hasegawa, H.; Humphrey, K.L.; Polyak, I.K.; Häse, C.C. Re-emergence of Vibrio tubiashii in bivalve shellfish aquaculture: Severity, environmental drivers, geographic extent and management. Dis. Aquat. Organ. 2008, 82, 119–134. [Google Scholar] [CrossRef]
- Tubiash, H.S.; Colwell, R.R.; Sakazaki, R. Marine vibrios associated with bacillary necrosis, a disease of larval and juvenile bivalve mollusks. J. Bacteriol. 1970, 103, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-León, J.; Villamill, L.; Salger, S.A.; Sallum, R.H.; Remacha-Triviño, A.; Leavitt, D.F.; Gómez-Chiarri, M. Survival of eastern oysters Crassostrea virginica from three lines following experimental challenge with bacterial pathogens. Dis. Aquat. Organ. 2008, 79, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Lind, E.J.; Boin, M.A.; Hase, C.C. The Extracellular Metalloprotease of Vibrio tubiashii Is a Major Virulence Factor for Pacific Oyster (Crassostrea gigas) Larvae. Appl. Environ. Microbiol. 2008, 74, 4101–4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, G.P.; Watson, M.A.; Needleman, D.S.; Church, K.M.; Häse, C.C. Mortalities of Eastern and Pacific Oyster Larvae Caused by the Pathogens Vibrio coralliilyticus and Vibrio tubiashii. Appl. Environ. Microbiol. 2015, 81, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubert, J.; Nelson, D.R.; Spinard, E.J.; Kessner, L.; Marta, G.-C.; da Costa, F.; Prado, S.; Barja, J.L. Following the infection process of vibriosis in Manila clam (Ruditapes philippinarum) larvae through GFP-tagged pathogenic Vibrio species. J. Invertebr. Pathol. 2016, 133, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balbi, T.; Auguste, M.; Cortese, K.; Montagna, M.; Borello, A.; Pruzzo, C.; Vezzulli, L.; Canesi, L. Responses of Mytilus galloprovincialis to challenge with the emerging marine pathogen Vibrio coralliilyticus. Fish Shellfish Immunol. 2019, 84, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Destoumieux-Garzón, D.; Canesi, L.; Oyanedel, D.; Travers, M.-A.; Charrière, G.M.; Pruzzo, C.; Vezzulli, L. Vibrio-bivalve interactions in health and disease. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Rubio, T.; Oyanedel, D.; Labreuche, Y.; Toulza, E.; Luo, X.; Bruto, M.; Chaparro, C.; Torres, M.; de Lorgeril, J.; Haffner, P.; et al. Species-specific mechanisms of cytotoxicity toward immune cells determine the successful outcome of Vibrio infections. Proc. Natl. Acad. Sci. USA 2019, 116, 14238–14247. [Google Scholar] [CrossRef] [Green Version]
- Prado, S.; Romalde, J.L.; Barja, J.L. Review of probiotics for use in bivalve hatcheries. Vet. Microbiol. 2010, 145, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Antimicrobial resistance in bacteria isolated from aquaculture sources in Australia. J. Appl. Microbiol. 2006, 100, 1103–1113. [Google Scholar] [CrossRef]
- Ho, S.P.; Hsu, T.Y.; Chen, M.H.; Wang, W.S. Antibacterial effect of chloramphenicol, thiamphenicol and florfenicol against aquatic animal bacteria. J. Vet. Med. Sci. 2000, 62, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, H.; Skubel, S.A.; Patel, H.; Cai Shi, D.; Bushek, D.; Chikindas, M.L. From Farm to Fingers: An Exploration of Probiotics for Oysters, from Production to Human Consumption. Probiotics Antimicrob. Proteins 2020, 12, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO). 2006. Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 1 September 2020).
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, prebiotics and immunomodulation of gut mucosal defences: Homeostasis and immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Gueimonde, M.; Peña, A.S.; Bernardo, D. Intestinal microbiota as modulators of the immune system. J. Immunol. Res. 2015, 2015, 159094. [Google Scholar] [CrossRef] [PubMed]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringø, E. Probiotics in shellfish aquaculture. Aquac. Fish. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Douillet, P.A.; Langdon, C.J. Use of a probiotic for the culture of larvae of the Pacific oyster (Crassostrea gigas Thunberg). Aquaculture 1994, 119, 25–40. [Google Scholar] [CrossRef]
- Kesarcodi-Watson, A.; Miner, P.; Nicolas, J.-L.; Robert, R. Protective effect of four potential probiotics against pathogen-challenge of the larvae of three bivalves: Pacific oyster (Crassostrea gigas), flat oyster (Ostrea edulis) and scallop (Pecten maximus). Aquaculture 2012, 344–349, 29–34. [Google Scholar] [CrossRef]
- Khouadja, S.; Haddaji, N.; Hanchi, M.; Bakhrouf, A. Selection of lactic acid bacteria as candidate probiotics for Vibrio parahaemolyticus depuration in pacific oysters (Crassostrea gigas). Aquac. Res. 2017, 48, 1885–1894. [Google Scholar] [CrossRef]
- Karim, M.; Zhao, W.; Rowley, D.; Nelson, D.; Marta, G.-C. Probiotic Strains for Shellfish Aquaculture: Protection of Eastern Oyster, Crassostrea virginica, Larvae and Juveniles Againsl Bacterial Challenge. J. Shellfish Res. 2013, 33, 401–408. [Google Scholar] [CrossRef]
- Sohn, S.; Lundgren, K.; Tammi, K.; Karim, M.; Smolowitz, R.; Nelson, D.R.; Rowley, D.C.; Marta, G.-C. Probiotic strains for disease management in hatchery larviculture of the eastern oyster Crassostrea virginica. J. Shellfish Res. 2016, 35, 307–317. [Google Scholar] [CrossRef]
- Beyersmann, P.G.; Tomasch, J.; Son, K.; Stocker, R.; Göker, M.; Wagner-Döbler, I.; Simon, M.; Brinkhoff, T. Dual function of tropodithietic acid as antibiotic and signaling molecule in global gene regulation of the probiotic bacterium Phaeobacter inhibens. Sci. Rep. 2017, 7, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, M.J.; D’alvise, P.W.; Rygaard, A.M.; Gram, L. Biofilm formation is not a prerequisite for production of the antibacterial compound tropodithietic acid in Phaeobacter inhibens DSM17395. J. Appl. Microbiol. 2014, 117, 1592–1600. [Google Scholar] [CrossRef] [PubMed]
- Grotkjær, T.; Bentzon-Tilia, M.; D’Alvise, P.; Dierckens, K.; Bossier, P.; Gram, L. Phaeobacter inhibens as probiotic bacteria in non-axenic Artemia and algae cultures. Aquaculture 2016, 462, 64–69. [Google Scholar] [CrossRef]
- Zhao, W.; Dao, C.; Karim, M.; Marta, G.-C.; Rowley, D.; Nelson, D. Contributions of tropodithietic acid and biofilm formation to the probiotic activity of Phaeobacter inhibens. BMC Microbiol. 2016, 16, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Yuan, T.; Piva, C.; Spinard, E.J.; Schuttert, C.W.; Rowley, D.C.; Nelson, D.R. The probiotic bacterium Phaeobacter inhibens downregulates virulence factor transcription in the shellfish pathogen Vibrio coralliilyticus by N-Acyl Homoserine Lactone Production. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newaj-Fyzul, A.; Austin, B. Probiotics, immunostimulants, plant products and oral vaccines, and their role as feed supplements in the control of bacterial fish diseases. J. Fish Dis. 2015, 38, 937–955. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High throughout Sequence Data. 2014. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 1 May 2018).
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Daley, T.; Smith, A.D. Predicting the molecular complexity of sequencing libraries. Nat. Methods 2013, 10, 325–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; Juan, G.-G.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Alexa, A.; Rahnenführer, J.; Lengauer, T. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 2006, 22, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Gerdol, M.; Marta, G.-C.; Castillo, M.G.; Figueras, A.; Fiorito, G.; Moreira, R.; Novoa, B.; Pallavicini, A.; Ponte, G.; Roumbedakis, K.; et al. Immunity in molluscs: Recognition and effector mechanisms, with a focus on Bivalvia. In Advances in Comparative Immunology; Springer International Publishing: Cham, Switzerland, 2018; ISBN 9783319767673. [Google Scholar]
- Estes, R.M.; Friedman, C.S.; Elston, R.A.; Herwig, R.P. Pathogenicity testing of shellfish hatchery bacterial isolates on Pacific oyster Crassostrea gigas larvae. Dis. Aquat. Organ. 2004, 58, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Decker, S.; Saulnier, D. Vibriosis induced by experimental cohabitation in Crassostrea gigas: Evidence of early infection and down-expression of immune-related genes. Fish Shellfish Immunol. 2011, 30, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Genard, B.; Miner, P.; Nicolas, J.-L.L.; Moraga, D.; Boudry, P.; Pernet, F.; Tremblay, R. Integrative study of physiological changes associated with bacterial infection in Pacific oyster larvae. PLoS ONE 2013, 8, e64534. [Google Scholar] [CrossRef] [Green Version]
- Bouallegui, Y. Immunity in mussels: An overview of molecular components and mechanisms with a focus on the functional defenses. Fish Shellfish Immunol. 2019, 89, 158–169. [Google Scholar] [CrossRef]
- Toubiana, M.; Rosani, U.; Giambelluca, S.; Cammarata, M.; Gerdol, M.; Pallavicini, A.; Venier, P.; Roch, P. Toll signal transduction pathway in bivalves: Complete cds of intermediate elements and related gene transcription levels in hemocytes of immune stimulated Mytilus galloprovincialis. Dev. Comp. Immunol. 2014, 45, 300–312. [Google Scholar] [CrossRef] [Green Version]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB Signaling in macrophages: Dynamics, crosstalk and signal integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Mu, C.; Yu, Y.; Zhao, J.; Wang, L.; Song, X.; Zhang, H.; Qiu, L.; Song, L. An inhibitor kappaB homologue from bay scallop Argopecten irradians. Fish Shellfish Immunol. 2010, 28, 687–694. [Google Scholar] [CrossRef]
- Zhang, G.; Ghosh, S. Negative regulation of toll-like receptor-mediated signaling by Tollip. J. Biol. Chem. 2002, 277, 7059–7065. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Li, R.; Wang, J.; Wang, S.; Zhang, M.; Hu, X.; Zhang, L.; Wang, S.; Wang, R.; Bao, Z. Identification, characterization and expression profiling of the Tollip gene in Yesso scallop (Patinopecten yessoensis). Genes Genet. Syst. 2015, 90, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shaghdali, K.; Durante, B.; Hayward, C.; Beal, J.; Foey, A. Macrophage subsets exhibit distinct E. coli-LPS tolerisable cytokines associated with the negative regulators, IRAK-M and Tollip. PLoS ONE 2019, 14, e0214681. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Ding, D.; Pan, B.; Bu, W. The TLR13-MyD88-NF-κB signalling pathway of Cyclina sinensis plays vital roles in innate immune responses. Fish Shellfish Immunol. 2017, 70, 720–730. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jouaux, A.; Ford, S.E.; Lelong, C.; Sourdaine, P.; Mathieu, M.; Guo, X. Transcriptome analysis reveals strong and complex antiviral response in a mollusc. Fish Shellfish Immunol 2015, 46, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Gerdol, M. Immune-related genes in gastropods and bivalves: A comparative overview. Invertebr. Surviv. J. 2017, 14, 103–118. [Google Scholar] [CrossRef]
- Vanhove, A.S.; Rubio, T.P.; Nguyen, A.N.; Lemire, A.; Roche, D.; Nicod, J.; Vergnes, A.; Poirier, A.C.; Disconzi, E.; Bachère, E.; et al. Copper homeostasis at the host vibrio interface: Lessons from intracellular vibrio transcriptomics. Environ. Microbiol. 2016, 18, 875–888. [Google Scholar] [CrossRef]
- Vidal-Dupiol, J.; Ladrière, O.; Destoumieux-Garzón, D.; Sautière, P.-E.; Meistertzheim, A.-L.; Tambutté, E.; Tambutté, S.; Duval, D.; Fouré, L.; Adjeroud, M.; et al. Innate immune responses of a scleractinian coral to vibriosis. J. Biol. Chem. 2011, 286, 22688–22698. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Dupiol, J.; Dheilly, N.M.; Rondon, R.; Grunau, C.; Cosseau, C.; Smith, K.M.; Freitag, M.; Adjeroud, M.; Mitta, G. Thermal stress triggers broad Pocillopora damicornis transcriptomic remodeling, while Vibrio coralliilyticus infection induces a more targeted immuno-suppression response. PLoS ONE 2014, 9, e107672. [Google Scholar] [CrossRef] [Green Version]
- Rey-Campos, M.; Moreira, R.; Valenzuela-Muñoz, V.; Gallardo-Escárate, C.; Novoa, B.; Figueras, A. High individual variability in the transcriptomic response of Mediterranean mussels to Vibrio reveals the involvement of myticins in tissue injury. Sci. Rep. 2019, 9, 3569. [Google Scholar] [CrossRef]
- Muñoz, K.; Flores-Herrera, P.; Gonçalves, A.T.; Rojas, C.; Yáñez, C.; Mercado, L.; Brokordt, K.; Schmitt, P. The immune response of the scallop Argopecten purpuratus is associated with changes in the host microbiota structure and diversity. Fish Shellfish Immunol. 2019, 91, 241–250. [Google Scholar] [CrossRef]
- Xu, D.; Zhao, Z.; Zhou, Z.; Lin, Y.; Zhang, X.; Zhang, Y.; Zhang, Y.; Li, J.; Mao, F.; Xiao, S.; et al. Mechanistic molecular responses of the giant clam Tridacna crocea to Vibrio coralliilyticus challenge. PLoS ONE 2020, 15, e0231399. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Song, X.; Song, L. The oyster immunity. Dev. Comp. Immunol. 2018, 80, 99–118. [Google Scholar] [CrossRef]
- Cao, R.; Liu, Y.; Wang, Q.; Yang, D.; Liu, H.; Ran, W.; Qu, Y.; Zhao, J. Seawater acidification reduced the resistance of Crassostrea gigas to Vibrio splendidus challenge: An energy metabolism perspective. Front. Physiol. 2018, 9, 880. [Google Scholar] [CrossRef]
- McDowell, I.C.; Nikapitiya, C.; Aguiar, D.; Lane, C.E.; Istrail, S.; Gomez-Chiarri, M. Transcriptome of American oysters, Crassostrea virginica, in response to bacterial challenge: Insights into potential mechanisms of disease resistance. PLoS ONE 2014, 9, e105097. [Google Scholar] [CrossRef] [Green Version]
- Richard, G.; Guérard, F.; Corporeau, C.; Lambert, C.; Paillard, C.; Pernet, F. Metabolic responses of clam Ruditapes philippinarum exposed to its pathogen Vibrio tapetis in relation to diet. Dev. Comp. Immunol. 2016, 60, 96–107. [Google Scholar] [CrossRef] [Green Version]
- Adeshina, I.; Abubakar, M.I.-O.; Ajala, B.E. Dietary supplementation with Lactobacillus acidophilus enhanced the growth, gut morphometry, antioxidant capacity, and the immune response in juveniles of the common carp, Cyprinus carpio. Fish Physiol. Biochem. 2020, 46, 1375–1385. [Google Scholar] [CrossRef]
- Chien, C.-C.; Lin, T.-Y.; Chi, C.-C.; Liu, C.-H. Probiotic, Bacillus subtilis E20 alters the immunity of white shrimp, Litopenaeus vannamei via glutamine metabolism and hexosamine biosynthetic pathway. Fish Shellfish Immunol. 2020, 98, 176–185. [Google Scholar] [CrossRef]
- Llewellyn, A.; Foey, A. Probiotic Modulation of Innate Cell Pathogen Sensing and Signaling Events. Nutrients 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- López-Carvallo, J.A.; Mazón-Suástegui, J.M.; Hernández-Oñate, M.Á.; Tovar-Ramírez, D.; Abasolo-Pacheco, F.; Morelos-Castro, R.M.; Arcos-Ortega, G.F. Transcriptome analysis of Catarina scallop (Argopecten ventricosus) juveniles treated with highly-diluted immunomodulatory compounds reveals activation of non-self-recognition system. PLoS ONE 2020, 15, e0233064. [Google Scholar] [CrossRef]
- Van Baarlen, P.; Troost, F.; van der Meer, C.; Hooiveld, G.; Boekschoten, M.; Brummer, R.J.M.; Kleerebezem, M. Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 4562–4569. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sánchez, T.; Ruiz-Zarzuela, I.; de Blas, I.; Balcázar, J.L. Probiotics in aquaculture: A current assessment. Rev. Aquacult. 2014, 6, 133–146. [Google Scholar] [CrossRef]
- McCormack, R.; Podack, E.R. Perforin-2/Mpeg1 and other pore-forming proteins throughout evolution. J. Leukoc. Biol. 2015, 98, 761–768. [Google Scholar] [CrossRef]
- He, X.; Zhang, Y.; Yu, Z. An Mpeg (macrophage expressed gene) from the Pacific oyster Crassostrea gigas: Molecular characterization and gene expression. Fish Shellfish Immunol. 2011, 30, 870–876. [Google Scholar] [CrossRef]
- Wiens, M.; Korzhev, M.; Krasko, A.; Thakur, N.L.; Sanja, P.-O.; Breter, H.J.; Ushijima, H.; Bärbel, D.-S.; Müller, I.M.; Müller, W.E. Innate immune defense of the sponge Suberites domuncula against bacteria involves a MyD88-dependent signaling pathway. Induction of a perforin-like molecule. J. Biol. Chem. 2005, 280, 27949–27959. [Google Scholar] [CrossRef] [Green Version]
- Bathige, S.D.N.K.; Umasuthan, N.; Whang, I.; Lim, B.-S.; Won, S.H.; Lee, J. Antibacterial activity and immune responses of a molluscan macrophage expressed gene-1 from disk abalone, Haliotis discus discus. Fish Shellfish Immunol. 2014, 39, 263–272. [Google Scholar] [CrossRef]
- Allam, B.; Espinosa, E. Bivalve immunity and response to infections: Are we looking at the right place? Fish Shellfish Immunol 2016, 53, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, E.; Koller, A.; Allam, B. Proteomic characterization of mucosal secretions in the eastern oyster, Crassostrea virginica. J. Proteom. 2016, 132, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Su, J.; Wang, Z.; Yan, X.; Yu, R.; Ma, P.; Li, Y.; Du, J. Transcriptomic analysis of Crassostrea sikamea × Crassostrea angulata hybrids in response to low salinity stress. PLoS ONE 2017, 12, e0171483. [Google Scholar] [CrossRef] [Green Version]
- Tuomola, E.M.; Ouwehand, A.C.; Salminen, S.J. The effect of probiotic bacteria on the adhesion of pathogens to human intestinal mucus. FEMS Immunol. Med. Microbiol. 1999, 26, 137–142. [Google Scholar] [CrossRef]
- Yu, H.; He, Y.; Wang, X.; Zhang, Q.; Bao, Z.; Guo, X. Polymorphism in a serine protease inhibitor gene and its association with disease resistance in the eastern oyster (Crassostrea virginica Gmelin). Fish Shellfish Immunol 2011, 30, 757–762. [Google Scholar] [CrossRef]
- Nikapitiya, C.; McDowel, I.C.; Villamil, L.; Muñoz, P.; Sohn, S.; Marta, G.-C. Identification of potential general markers of disease resistance in American oysters, Crassostrea virginica through gene expression studies. Fish Shellfish Immunol. 2014, 41, 27–36. [Google Scholar] [CrossRef]
- Kawasaki, H.; Koyama, T.; Conlon, J.; Yamakura, F.; Iwamuro, S. Antimicrobial action of histone H2B in Escherichia coli: Evidence for membrane translocation and DNA-binding of a histone H2B fragment after proteolytic cleavage by outer membrane proteinase T. Biochimie 2008, 90, 1693–1702. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dorrington, T.; Marta, G.-C. The role of histones in the immune responses of aquatic invertebrates. Invertebr. Surviv. J. 2013, 10, 94–101. [Google Scholar]
- Poirier, A.C.; Schmitt, P.; Rosa, R.D.; Vanhove, A.S.; Sylvie, K.-J.; Rubio, T.P.; Charrière, G.M.; Delphine, D.-G. Antimicrobial Histones and DNA Traps in Invertebrate Immunity EVIDENCES IN CRASSOSTREA GIGAS. J. Biol. Chem. 2014, 289, 24821–24831. [Google Scholar] [CrossRef] [Green Version]
- Nitta, T.; Takahama, Y. The lymphocyte guard-IANs: Regulation of lymphocyte survival by IAN/GIMAP family proteins. Trends Immunol. 2007, 28, 58–65. [Google Scholar] [CrossRef]
- McDowell, I.C.; Modak, T.H.; Lane, C.E.; Gomez-Chiarri, M. Multi-species protein similarity clustering reveals novel expanded immune gene families in the eastern oyster Crassostrea virginica. Fish Shellfish Immunol. 2016, 53, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Torraca, V.; Mostowy, S. Septins and Bacterial Infection. Front. Cell Dev. Biol. 2016, 4, 127. [Google Scholar] [CrossRef] [Green Version]
- Mostowy, S.; Cossart, P. Septins as key regulators of actin based processes in bacterial infection. Biol. Chem. 2011, 392, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Manzanares, M.; Sánchez-Madrid, F. Role of the cytoskeleton during leukocyte responses. Nat. Rev. Immunol. 2004, 4, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Shenoy, A.R. The cytoskeleton in cell-autonomous immunity: Structural determinants of host defence. Nat. Rev. Immunol. 2015, 15, 559–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | RPS (Average +/− SD) | ||
|---|---|---|---|
| 6 h | 24 h | 7 days | |
| S4 + RE22 | 37 ± 26 | 41 ± 2 | - |
| RI + RE22 | 30 ± 39 | 45 ± 5 | 28 ± 6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modak, T.H.; Gomez-Chiarri, M. Contrasting Immunomodulatory Effects of Probiotic and Pathogenic Bacteria on Eastern Oyster, Crassostrea Virginica, Larvae. Vaccines 2020, 8, 588. https://doi.org/10.3390/vaccines8040588
Modak TH, Gomez-Chiarri M. Contrasting Immunomodulatory Effects of Probiotic and Pathogenic Bacteria on Eastern Oyster, Crassostrea Virginica, Larvae. Vaccines. 2020; 8(4):588. https://doi.org/10.3390/vaccines8040588
Chicago/Turabian StyleModak, Tejashree H., and Marta Gomez-Chiarri. 2020. "Contrasting Immunomodulatory Effects of Probiotic and Pathogenic Bacteria on Eastern Oyster, Crassostrea Virginica, Larvae" Vaccines 8, no. 4: 588. https://doi.org/10.3390/vaccines8040588
APA StyleModak, T. H., & Gomez-Chiarri, M. (2020). Contrasting Immunomodulatory Effects of Probiotic and Pathogenic Bacteria on Eastern Oyster, Crassostrea Virginica, Larvae. Vaccines, 8(4), 588. https://doi.org/10.3390/vaccines8040588