Efficacy of a Turkey Herpesvirus Vectored Newcastle Disease Vaccine against Genotype VII.1.1 Virus: Challenge Route Affects Shedding Pattern

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccine and Vaccination
2.3. Serology
2.4. Challenge Virus
2.5. Challenge Infections and Post-Challenge Samplings
2.6. ND Challenge Virus Shedding Measurement
2.7. Statistical Analysis
3. Results
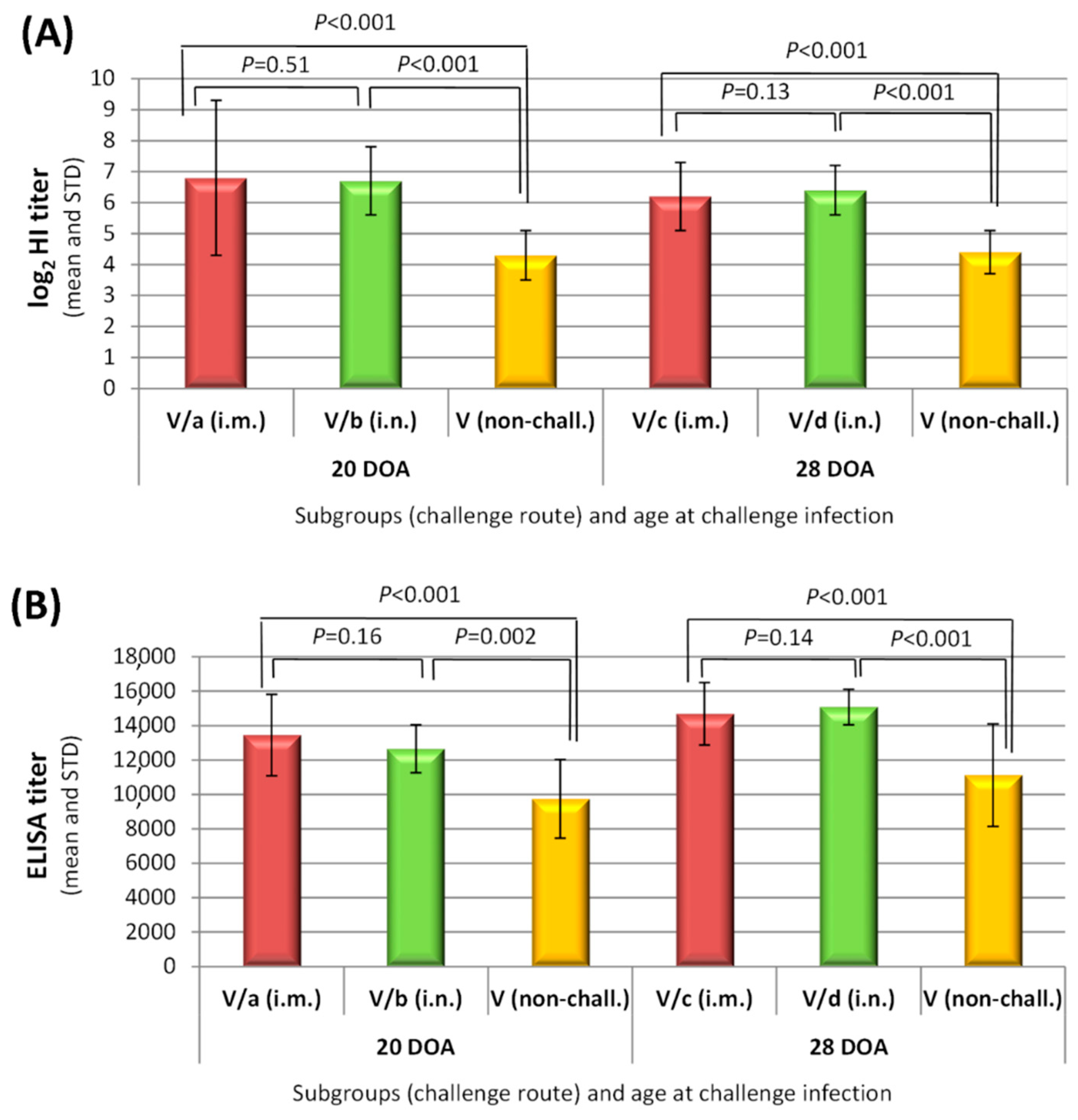
3.1. Vaccine-Take Detection and Humoral Immune Response to Vaccination
3.2. Efficacy against NDV Challenge
3.2.1. Prevention of Clinical Signs and Mortality
3.2.2. Effect of the Challenge Route on Challenge Virus Shedding by Vaccinated Chickens
3.2.3. Humoral Immune Response to Challenge
3.3. Comparison of the Challenge Virus F Gene Sequence with F Gene Insert of rHVT-ND
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hines, N.L.; Miller, C.L. Avian paramyxovirus serotype-1: A review of disease distribution, clinical symptoms, and laboratory diagnostics. Vet. Med. Int. 2012, 2012, 708216. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Haddas, R.; Simanov, L.; Lublin, A.; Rehmani, S.F.; Wajid, A.; Bibi, T.; Khan, T.A.; Yaqub, T.; Setiyaningsih, S.; et al. Identification of new sub-genotypes of virulent Newcastle disease virus with potential panzootic features. Infect. Genet. Evol. 2015, 29, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Rui, Z.; Juan, P.; Jingliang, S.; Jixun, Z.; Xiaoting, W.; Shouping, Z.; Xiaojiao, L.; Guozhong, Z. Phylogenetic characterization of Newcastle disease virus isolated in the mainland of China during 2001–2009. Vet. Microbiol. 2010, 141, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Hu, J.; Hu, S.; Liu, X.; Wang, X.; Zhu, J.; Liu, X. Strong innate immune response and cell death in chicken splenocytes infected with genotype VIId Newcastle disease virus. Virol. J. 2012, 9, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenglei, H.; Liu, X. Immune response-induced NDV pathology. Br. J. Virol. 2015, 2, 25–27. [Google Scholar]
- Czegledi, A.; Ujvari, D.; Somogyi, E.; Wehmann, E.; Werner, O.; Lomniczi, B. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications. Virus Res. 2006, 120, 36–48. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Ramey, A.M.; Qiu, X.; Bahl, J.; Afonso, C.L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infect. Genet. Evol. 2016, 39, 22–34. [Google Scholar] [CrossRef] [Green Version]
- Awad, A.M.; Sedeik, M.E.; Abdelkariem, A.A. Isolation, molecular characterization and pathotyping of Newcastle disease viruses from field outbreaks among broiler flocks in Egypt from 2014–2015. Int. J. Curr. Res. 2015, 7, 12925–12934. [Google Scholar]
- Mohamed, M.H.A.; Kumar, S.; Paldurai, A.; Samal, S.K. Sequence analysis of fusion protein gene of Newcastle disease virus isolated from outbreaks in Egypt during 2006. Virol. J. 2011, 8, 237. [Google Scholar] [CrossRef] [Green Version]
- Orabi, A.; Hussein, A.; Saleh, A.A.; El-Magd, M.A.; Munir, M. Evolutionary insights into the fusion protein of Newcastle disease virus isolated from vaccinated chickens in 2016 in Egypt. Arch. Virol. 2017, 162, 3069–3079. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.M.; Samy, A.; Soliman, M.A.; Arafa, A.; Zanaty, A.; Hassan, M.K.; Sultan, A.H.; Bazid, A.I.; Hussein, A.H. Genotypic and pathogenic characterization of genotype VII Newcastle disease viruses isolated from commercial farms in Egypt and evaluation of heterologous antibody responses. Arch. Virol. 2017, 162, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines-A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Gallili, G.E.; Ben-Nathan, D. Newcastle disease vaccines. Biotechnol. Adv. 1998, 16, 343–366. [Google Scholar] [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef]
- Van Boven, M.; Bouma, A.; Fabri, T.H.; Katsma, E.; Hartog, L.; Koch, G. Herd immunity to Newcastle disease virus in poultry by vaccination. Avian Pathol. 2008, 37, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Meulemans, G. Control by vaccination. In Newcastle Disease; Alexander, D.J., Ed.; Kluwer Academic Publishers (Springer US): Boston, MA, USA, 1988; pp. 318–332. [Google Scholar] [CrossRef]
- Dortmans, J.C.; Peeters, B.P.; Koch, G. Newcastle disease virus outbreaks: Vaccine mismatch or inadequate application? Vet. Microbiol. 2012, 160, 17–22. [Google Scholar] [CrossRef]
- Dortmans, J.C.; Venema-Kemper, S.; Peeters, B.P.; Koch, G. Field vaccinated chickens with low antibody titres show equally insufficient protection against matching and non-matching genotypes of virulent Newcastle disease virus. Vet. Microbiol. 2014, 172, 100–107. [Google Scholar] [CrossRef]
- Bennejean, G.; Guittet, M.; Picault, J.P.; Bouquet, J.F.; Devaux, B.; Gaudry, D.; Moreau, Y. Vaccination of one-day-old chicks against newcastle disease using inactivated oil adjuvant vaccine and/or live vaccine. Avian Pathol. 1978, 7, 15–27. [Google Scholar] [CrossRef]
- Current and Future Applications of Viral-Vectored Recombinant Vaccines in Poultry. Available online: https://en.engormix.com/poultry-industry/articles/current-future-applications-viral-t39410.htm (accessed on 4 September 2020).
- Nagai, Y.; Hamaguchi, M.; Toyoda, T. Molecular biology of Newcastle disease virus. Prog. Vet. Microbiol. Immunol. 1989, 5, 16–64. [Google Scholar]
- Morgan, R.W.; Gelb, J., Jr.; Schreurs, C.S.; Lutticken, D.; Rosenberger, J.K.; Sondermeijer, P.J. Protection of chickens from Newcastle and Marek’s diseases with a recombinant herpesvirus of turkeys vaccine expressing the Newcastle disease virus fusion protein. Avian Dis. 1992, 36, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector Newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Palya, V.; Kiss, I.; Tatár-Kis, T.; Mató, T.; Felföldi, B.; Gardin, Y. Advancement in vaccination against Newcastle disease: Recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions. Avian Dis. 2012, 56, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Palya, V.; Tatar-Kis, T.; Mato, T.; Felfoldi, B.; Kovacs, E.; Gardin, Y. Onset and long-term duration of immunity provided by a single vaccination with a turkey herpesvirus vector ND vaccine in commercial layers. Vet. Immunol. Immunopathol. 2014, 158, 105–115. [Google Scholar] [CrossRef]
- Rauw, F.; Gardin, Y.; Palya, V.; Anbari, S.; Lemaire, S.; Boschmans, M.; van den Berg, T.; Lambrecht, B. Improved vaccination against Newcastle disease by an in ovo recombinant HVT-ND combined with an adjuvanted live vaccine at day-old. Vaccine 2010, 28, 823–833. [Google Scholar] [CrossRef]
- Sonoda, K.; Sakaguchi, M.; Okamura, H.; Yokogawa, K.; Tokunaga, E.; Tokiyoshi, S.; Kawaguchi, Y.; Hirai, K. Development of an effective polyvalent vaccine against both Marek’s and Newcastle diseases based on recombinant Marek’s disease virus type 1 in commercial chickens with maternal antibodies. J. Virol. 2000, 74, 3217–3226. [Google Scholar] [CrossRef] [Green Version]
- Afonso, C.L.; Miller, P.J.; Grund, C.; Koch, G.; Peeters, B.; Selleck, P.W.; Srinivas, G.B. Newcastle Disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2018; pp. 964–983. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a Real-Time Reverse-Transcription PCR for Detection of Newcastle Disease Virus RNA in Clinical Samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.H.; Kwon, H.J.; Kim, T.E.; Kim, J.H.; Yoo, H.S.; Kim, S.J. Variation of a newcastle disease virus hemagglutinin-neuraminidase linear epitope. J. Clin. Microbiol. 2008, 46, 1541–1544. [Google Scholar] [CrossRef] [Green Version]
- Kapczynski, D.R.; King, D.J. Protection of chickens against overt clinical disease and determination of viral shedding following vaccination with commercially available Newcastle disease virus vaccines upon challenge with highly virulent virus from the California 2002 exotic Newcastle disease outbreak. Vaccine 2005, 23, 3424–3433. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; de Almeida, R.S.; Gil, P.; Majó, N.; Nofrarías, M.; Briand, F.-X.; Jestin, V.; Albina, E. Can genotype mismatch really affect the level of protection conferred by Newcastle disease vaccines against heterologous virulent strains? Vaccine 2018, 36, 3917–3925. [Google Scholar] [CrossRef] [PubMed]
- Tatar-Kis, T.; Fischer, E.A.J.; Cazaban, C.; Walko-Kovacs, E.; Homonnay, Z.G.; Velkers, F.C.; Palya, V.; Stegeman, J.A. A Herpesvirus of Turkey-Based Vector Vaccine Reduces Transmission of Newcastle Disease Virus in Commercial Broiler Chickens with Maternally Derived Antibodies. Vaccines 2020, 8, 614. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Groups | Subgroups | Age at Challenge | Route of Challenge b | Number of Birds |
|---|---|---|---|---|
| Vaccinated a | V/a | 20 DOA c | Intra-muscular | 20 |
| V/b | 20 DOA | Intra-nasal | 20 | |
| V/c | 28 DOA | Intra-muscular | 20 | |
| V/d | 28 DOA | Intra-nasal | 20 | |
| V/Nch d | - | - | 12 | |
| Control | C/a | 20 DOA | Intra-muscular | 10 |
| C/b | 20 DOA | Intra-nasal | 10 | |
| C/c | 28 DOA | Intra-muscular | 10 | |
| C/d | 28 DOA | Intra-nasal | 10 |
| Age of Chickens (Day) | 7 | 15 | 20 | 36 | 43 | ||
|---|---|---|---|---|---|---|---|
| Sample type | Feather pulp | Feather pulp | spleen | Feather pulp | spleen | spleen | Feather pulp |
| Positivity a | 90% | 100% | 100% | 100% | 100% | 100% | 58% |
| Positivity (positives/tested) | 18/20 | 20/20 | 10/10 | 20/20 | 20/20 | 12/12 | 7/12 |
| Ct value mean ± STD (rHVT-ND) | 31.0 ± 3.8 | 24.8 ± 3.8 | 29.0 ± 0.9 | 29.7 ± 5.6 | 30.6 ± 1.9 | 31.6 ± 3.3 | 36.8 ± 3.6 |
| Ct value mean ± STD (ovotransferrin) | 23.1 ± 0.7 | 23.2 ± 0.9 | 18.9 ± 0.5 | 23.5 ± 0.9 | 18.9 ± 0.9 | 18.8 ± 0.6 | 24.0 ± 1.5 |
| ΔCt b | 7.9 ± 3.6 | 1.6 ± 3.8 | 10.1 ± 1.2 | 6.2 ± 5.8 | 11.7 ± 1.9 | 12.9 ± 3.2 | 12.8 ± 3.7 |
| Method | Age at Sampling (Day) | 7 | 15 | 20 | 28 | 35 | 43 | |
|---|---|---|---|---|---|---|---|---|
| Parameter | Group | |||||||
| IDScreen® ND Indirect ELISA | ELISA titer mean ± STD | Vaccinated | 31 ± 39 | 2516 ± 1936 | 3964 ± 1697 | 8608 ± 2215 | 9743 ± 2294 | 11,119 ± 2976 |
| Control | NS | NS | 137 ± 287 | 55 ± 105 | 86 ± 184 | 1 ± 0 | ||
| Positivity | Vaccinated | 0% | 80% | 100% | 100% | 100% | 100% | |
| Control | NS | NS | 0% | 0% | 0% | 0% | ||
| HI test | Log2 HI titer mean ± STD | Vaccinated | 0.1 ± 0.2 | 0.9 ± 0.5 | 1.6 ± 0.9 | 2.5 ± 0.7 | 4.3 ± 0.8 | 4.4 ± 0.7 |
| Control | NS | NS | 0.0 ± 0.0 | 0.2 ± 0.4 | 1.9 ± 0.2 | 1.7 ± 0.4 | ||
| Positivity | Vaccinated | 0% | 0% | 55% | 90% | 100% | 100% | |
| Control | NS | NS | 0% | 0% | 0% | 0% | ||
| Age at Challenge | Route of Challenge | Sub-Group | Oro-Nasal Swabs | Cloacal Swabs | ||||
|---|---|---|---|---|---|---|---|---|
| Ct (Mean ± STD and Range) | p-Value § | Positivity # | Ct (Mean ± STD and Range) | p-Value § | Positivity # | |||
| 20 DOA * | i.m. | V/a (vaccinated) | 35.0 ± 3.2 (25.8–36.0) | p < 0.001 | 1/10 | 34.4 ± 3.4 (26.7–36.0) | p < 0.001 | 2/10 |
| C/a (control) | 16.2 ± 0.9 (14.4–17.5) | 10/10 | 19.4 ± 1.6 (16.6–22.5) | 10/10 | ||||
| i.n. | V/b (vaccinated) | 29.8 ± 3.2 (26.2–34.9) | p < 0.001 | 10/10 | 35.4 ± 1.3 (32.4–36.0) | p < 0.001 | 2/10 | |
| C/b (control) | 19.7 ± 1.4 (18.4–22.6) | 10/10 | 22.3 ± 2.0 (19.3–26.2) | 10/10 | ||||
| 28 DOA | i.m. | V/c (vaccinated) | 35.8 ± 0.7 (33.8–36.0) | p < 0.001 | 1/10 | 36.0 ± 0.0 (36.0–36.0) | p < 0.001 | 0/10 |
| C/c (control) | 16.7 ± 1.6 (14.2–19.0) | 10/10 | 21.0 ± 2.6 (16.6–23.9) | 10/10 | ||||
| i.n. | V/d (vaccinated) | 30.1 ± 4.1 (25.2–36.0) | p < 0.001 | 9/10 | 36.0 ± 0.0 (36.0–36.0) | p < 0.001 | 0/10 | |
| C/d (control) | 18.8 ± 1.0 (16.8–20.3) | 10/10 | 23.9 ± 2.2 (19.4–27.0) | 10/10 | ||||
| Subgroup | Non-Challenged | Intra-Muscularly Challenged | Intra-Nasally Challenged |
|---|---|---|---|
| ELISA titre (mean ± STD) | 119 ± 92 | 621 ± 872 | 590 ± 389 |
| Positivity | 0% | 10% | 15% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palya, V.; Tatár-Kis, T.; Arafa, A.S.A.; Felföldi, B.; Mató, T.; Setta, A. Efficacy of a Turkey Herpesvirus Vectored Newcastle Disease Vaccine against Genotype VII.1.1 Virus: Challenge Route Affects Shedding Pattern. Vaccines 2021, 9, 37. https://doi.org/10.3390/vaccines9010037
Palya V, Tatár-Kis T, Arafa ASA, Felföldi B, Mató T, Setta A. Efficacy of a Turkey Herpesvirus Vectored Newcastle Disease Vaccine against Genotype VII.1.1 Virus: Challenge Route Affects Shedding Pattern. Vaccines. 2021; 9(1):37. https://doi.org/10.3390/vaccines9010037
Chicago/Turabian StylePalya, Vilmos, Tímea Tatár-Kis, Abdel Satar A. Arafa, Balázs Felföldi, Tamás Mató, and Ahmed Setta. 2021. "Efficacy of a Turkey Herpesvirus Vectored Newcastle Disease Vaccine against Genotype VII.1.1 Virus: Challenge Route Affects Shedding Pattern" Vaccines 9, no. 1: 37. https://doi.org/10.3390/vaccines9010037
APA StylePalya, V., Tatár-Kis, T., Arafa, A. S. A., Felföldi, B., Mató, T., & Setta, A. (2021). Efficacy of a Turkey Herpesvirus Vectored Newcastle Disease Vaccine against Genotype VII.1.1 Virus: Challenge Route Affects Shedding Pattern. Vaccines, 9(1), 37. https://doi.org/10.3390/vaccines9010037