
Field Evaluation of Commercial Vaccines against Infectious Bovine Rhinotracheitis (Ibr) Virus Using Different Immunization Protocols

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccination Protocols
2.3. Vaccines
- Vaccine A: inactivated suspension of BoHV-1, bovine viral diarrhea virus (BVDV)-1, Campylobacter fetus, Histophilus somni and Leptospira spp in aluminum hydroxide adjuvant.
- Vaccine B: inactivated suspension of BoHV-1, BoHV-5; BVDV-1, BVDV-2; Leptospira spp and Campylobacter fetus in aluminum hydroxide adjuvant.
- Vaccine C: inactivated BoHV-1, BVDV-1, Campylobacter fetus, Leptospira spp and Histophilus somni in aluminum hydroxide adjuvant.
- Vaccine D: inactivated suspension of BoHV-1, BoHV-5, BVDV-1, BVDV-2, Leptospira spp and Campylobacter fetus in oil—adjuvant.
- Clostridial vaccine: Bacterin-toxoid combined with a water-soluble adjuvant containing Clostridium chauvoei, Clostridium septicum, Clostridium haemolyticum, Clostridium novyi, Clostridium sordellii, Clostridium perfringens types B and C and Clostridium perfringens type D.
- FMD vaccine: Oil emulsion vaccine containing inactivated Foot and Mouth Disease Virus types O1 Campos and A24 Cruzeiro.
2.4. Serum Neutralization (SN) Test
2.5. Indirect ELISA
2.6. Statistical Analysis
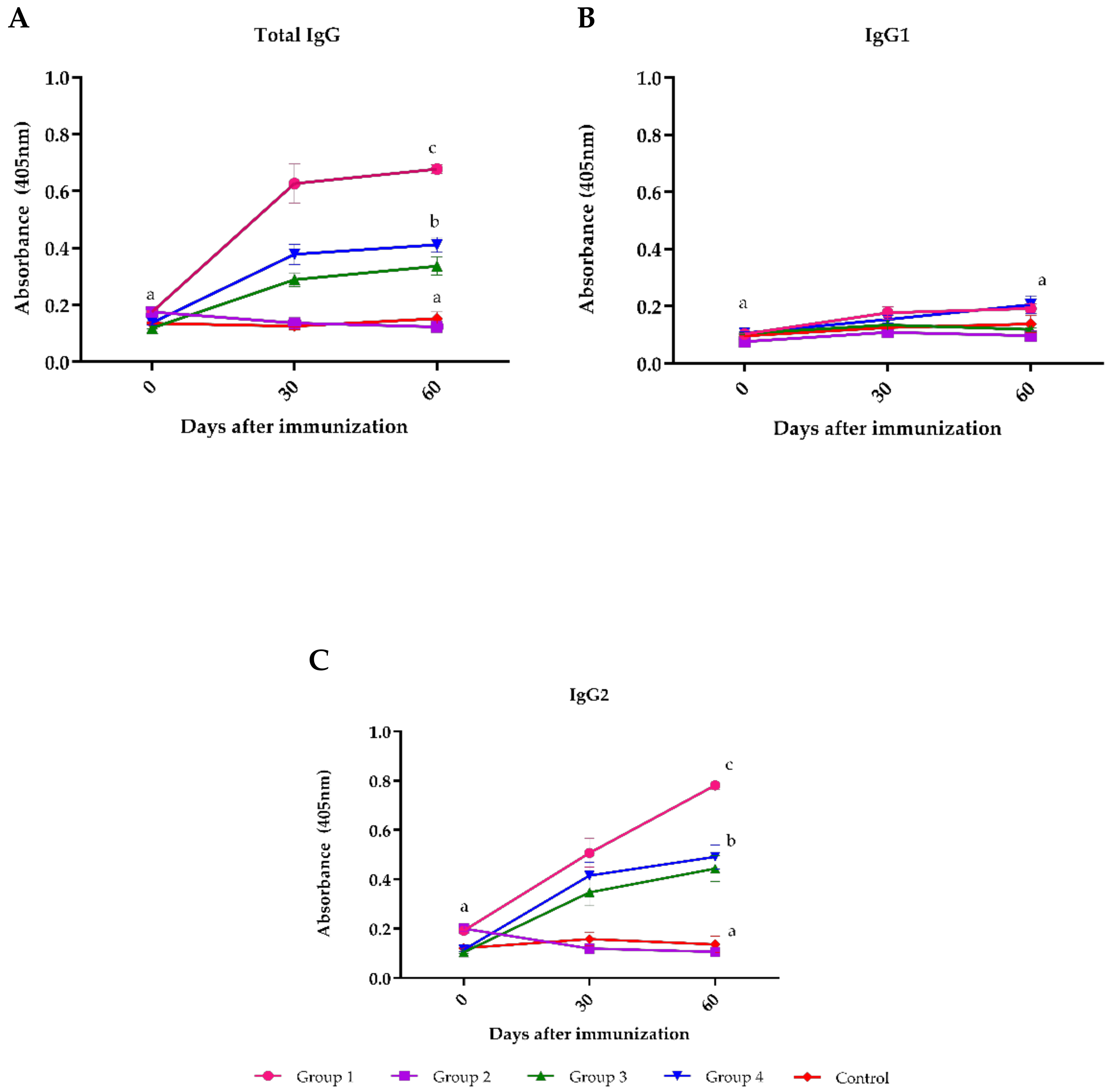
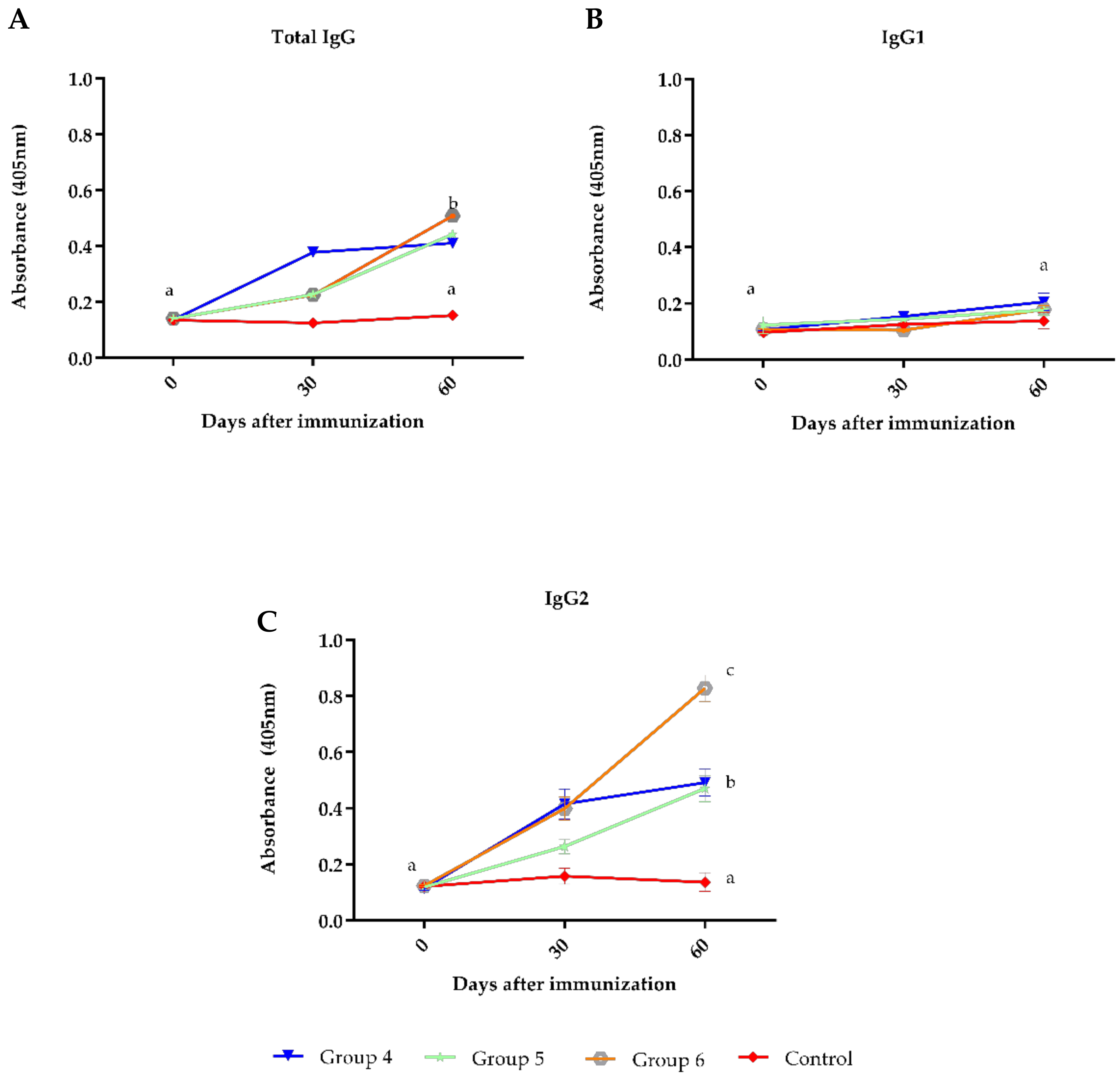
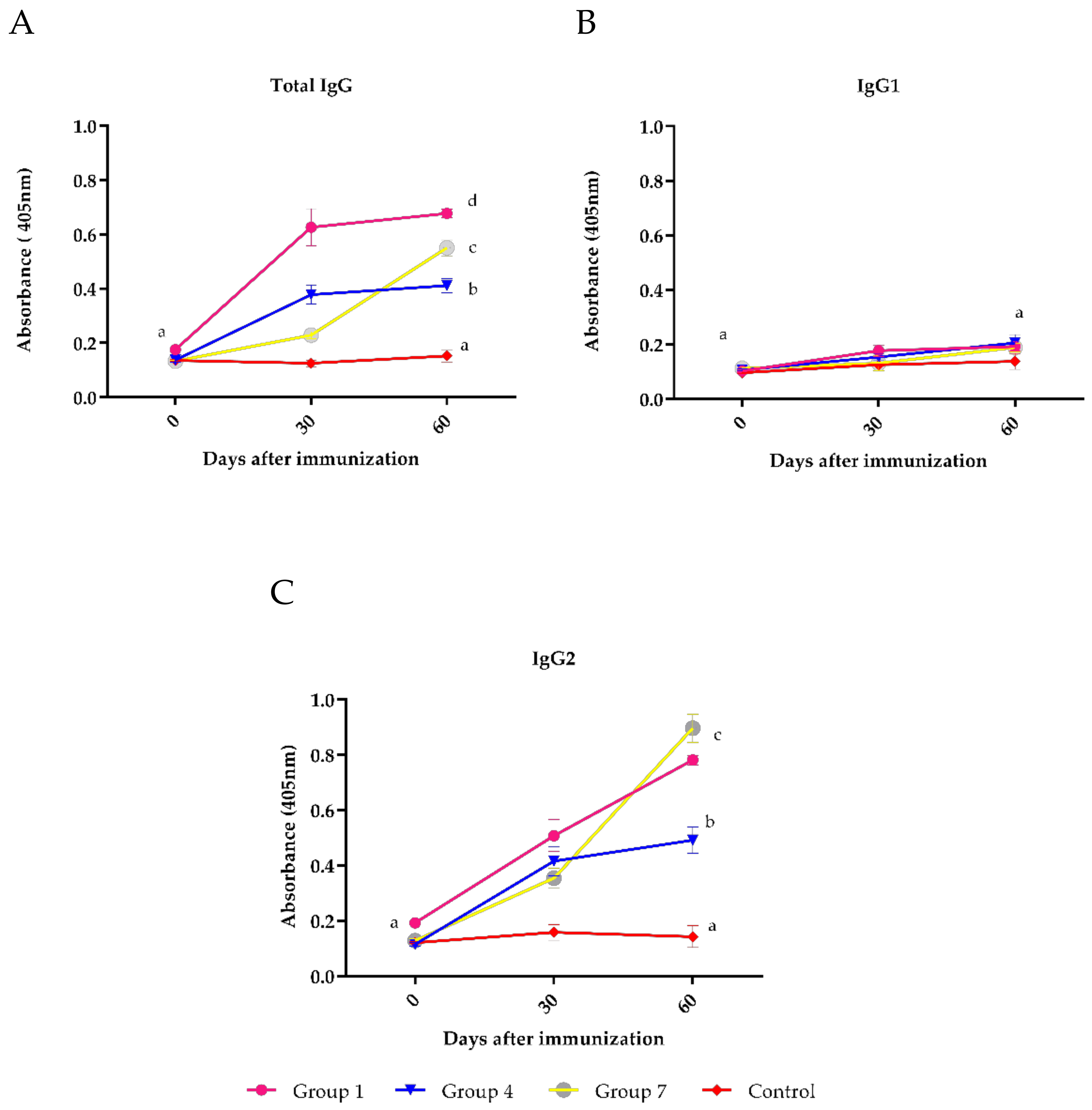
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engels, M.; Achermann, M. Pathogenesis of ruminant Herpesvirus infections. Vet. Microbiol. 1996, 54, 3–15. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses Universal Virus Database [ICTVdB]. Management Virus Taxonomy: 2015 Release EC 47, London, UK, July 2015. Email Ratification 2016 (MSL #30). Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy_releases (accessed on 10 August 2020).
- Ackermann, M.; Peterhans, E.; Wyler, R. DNA of bovine herpesvirus 1 in trigeminal ganglia of latently infected calves. Am. J. Vet. Res. 1982, 4, 36–40. [Google Scholar]
- Muylkens, B.; Thiry, J.; Kirten, P.; Schynts, F.; Thiry, E. Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet. Res. 2007, 38, 181–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rós, C.; Belák, S. Characterization of glycoprotein B gene from ruminant alphaherpesviruses. Virus Genes 2002, 24, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, M.; Weynants, V.; Godfroid, J.; Schynts, F.; Meyer, G.; Letesson, J.J.; Thiry, E. Effects of bovine herpesvirus type 1 infection in calves with maternal antibodies on immune response and virus latency. J. Clin. Microbiol. 2000, 38, 1885–1894. [Google Scholar] [CrossRef] [Green Version]
- Guarino, H.; Nuñez, A.; Repiso, M.; Gil, A.; Dargatz, D. Prevalence of serum antibodies to bovine herpesvirus-1 and bovine viral diarrhea virus in beef cattle in Uruguay. Prev. Vet. Med. 2008, 85, 34–40. [Google Scholar] [CrossRef]
- Alonzo, P.; Puentes, R.; Benavides, U.; Isnardi, F.; García, R.; Piaggio, J.; Cavestany, D.; Roses, G.; Maisonnave, J. Effect of Bovine Herpesvirus 1 infection on pregnancy rate of beef cows in Uruguay. Veterinaria 2012, 48, 5–12. [Google Scholar]
- Newcomer, B.W.; Cofield, L.G.; Walz, P.H.; Givens, M.D. Prevention of abortion in cattle following vaccination against bovine herpesvirus 1: A meta-analysis. Prev. Vet. Med. 2017, 138, 1–8. [Google Scholar] [CrossRef]
- Manoj, S.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Immunization with a dicistronic plasmid expressing a truncated form of bovine herpesvirus-1 glycoprotein D and the amino terminal subunit of glycoprotein B results in reduced gB-specific immune responses. Virology 2003, 313, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Spilki, F.R.; Franco, A.C.; Roehe, P.M. Analysis of isotype-specific antibody responses to bovine herpes viruses 1.1 and 1.2a allows to estimate the stage of infection. Braz. J. Microbiol. 2012, 43, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Petzhold, S.; Reckziegel, P.; Prado, J.; Teixeira, J.; Wald, V.; Esteves, P.; Spilki, F.; Roehe, P. Neutralizing antibodies to bovine herpesviruses types 1 (BHV-1) and 5 (BHV-5) induced by an experimental, oil-adjuvanted, BHV-1 vaccine. Braz. J. Vet. Res. Anim. Sci. 2001, 38, 184–187. [Google Scholar] [CrossRef]
- Silva, A.D.; Esteves, P.A.; Dezen, D.; Oliveira, A.P.; Spilki, F.R.; Campos, F.S.; Franco, A.C.; Roehe, P.M. Efficacy of a gE-deleted, bovine herpesvirus 1 (BoHV-1) inactivated vaccine. Pesq. Vet. Bras. 2009, 29, 545–551. [Google Scholar] [CrossRef]
- Campos, F.S.; Dezen, D.; Antunes, D.A.; Santos, H.F.; Arantes, T.S.; Cenci, A.; Gomes, F.; Lima, F.E.; Brito, W.M.; Filho, H.C.; et al. Efficacy of an inactivated, recombinant bovine herpesvirus type 5 (BoHV-5) vaccine. Vet. Microbiol. 2011, 148, 18–26. [Google Scholar] [CrossRef]
- Varela, A.P.; Holz, C.L.; Cibulski, S.P.; Teixeira, T.F.; Antunes, D.A.; Franco, A.C.; Roehe, L.R.; Oliveira, M.T.; Campos, F.S.; Dezen, D.; et al. Neutralizing antibodies to bovine herpesvirus types 1 (BoHV-1) and 5 (BoHV-5) and its subtypes. Vet. Microbiol. 2010, 142, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie-Dyck, S.; Kovacs-Nolan, J.; Snider, M.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Inclusion of the bovine neutrophil betadefensin 3 with glycoprotein D of bovine herpesvirus-1 in a DNA vaccine modulates immune responses of mice and cattle. Clin. Vaccine Immunol. 2014, 21, 463–477. [Google Scholar] [CrossRef] [Green Version]
- Estes, D.M.; Brown, W.C. Type 1 and type 2 responses in regulation of Ig isotype expression in cattle. Vet. Immunol. Immunopathol. 2002, 90, 1–10. [Google Scholar] [CrossRef]
- Puentes, R.; De Brun, L.; Algorta, A.; Da Silva, V.; Mansilla, F.; Sacco, G.; Llambí, S.; Capozzo, A.V. Evaluation of serological response to foot-and-mouth disease vaccination in BLV infected cows. BMC Vet. Res. 2016, 12, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE, 2020. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Available online: http://www.oie.int/manual-of-diagnostic-tests-and-vaccines-for-terrestrial-animals (accessed on 10 November 2020).
- Anziliero, D.; Martins, M.; Weiss, M.; Monteiro, F.L.; Ataide, C.F.; Weiblen, R.; Flores, E.F. Serological response to bovine herpesvírus 1 and 5 and bovine viral diarrhea virus induced by commercial vaccines. Ciênc. Rural 2015, 45, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.F.; da Silva, L.F.; Weiblen, R.; Flores, E.F. Immunogenicity of commercial inactivated bovine herpesvirus type 1 vaccines. Cienc. Rural 2007, 37, 1471–1474. [Google Scholar] [CrossRef] [Green Version]
- Vogel, F.; Flores, E.; Weiblen, R.; Kunrath, C. Neutralizing activity to bovine herpesvirus types 1 (BHV-1) and 5 (BHV-5) in sera of cattle immunized with vaccines against BHV-1. Ciênc. Rural 2002, 32, 881–883. [Google Scholar] [CrossRef] [Green Version]
- Walz, P.H.; Givens, M.D.; Rodning, S.P.; Riddell, K.P.; Brodersen, B.W.; Scruggs, D.; Grotelueschen, D. Evaluation of reproductive protection against bovine viral diarrhea virus and bovine herpesvirus-1 afforded by annual revaccination with modified-live viral or combination modified-live/killed viral vaccines after primary vaccination with modified-live viral vaccine. Vaccine 2017, 35, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Baccili, C.; Martin, C.; Silva, K.N.; Nichi, M.; Flores, E.; Vercesi Filho, A.; Pituco, E.; Gomes, V. Serological response against bovine herpesvirus and bovine viral diarrhea virus induced by commercial vaccines in Holstein heifers. Pesq. Vet. Bras. 2019, 39, 870–878. [Google Scholar] [CrossRef]
- Todryk, S.M. T Cell Memory to Vaccination. Vaccines 2018, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firacative, C.; Gressler, A.E.; Schubert, K.; Schulze, B.; Müller, U.; Brombacher, F.; von Bergen, M.; Alber, G. Identification of T helper (Th)1- and Th2-associated antigens of Cryptococcus neoformans in a murine model of pulmonary infection. Sci. Rep. 2018, 8, 2681. [Google Scholar] [CrossRef]
- Leites, M.; Puentes, R. Quantification of immunoglobulin isotypes against Bovine herpesvirus type 1 (BoHV-1) in vaccinated and naturally infected animals. In Proceedings of the XII Congreso Argentino de Virologia, Buenos Aires, Argentina, 26–28 September 2017. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Groups | Vaccines |
|---|---|
| 1 | A |
| 2 | B |
| 3 | C |
| 4 | D |
| 5 | D + Clostridial vaccine |
| 6 | D + FMD vaccine |
| 7 | A (1st dose) + D (2nd dose) |
| Control | Non-vaccinated |
| Groups | Vaccines | Number of Animals | Reagents | |
|---|---|---|---|---|
| 30 dpv | 60 dpv | |||
| 1 | A | 15 | 4 * #4(8) | 13 *,a #2(8) #2(12) #2(16) #1(24) #5(32) #1(64) |
| 2 | B | 15 | 0 * | 0 b |
| 3 | C | 15 | 1 * #1(8) | 1 b #1(8) |
| 4 | D | 15 | 2 * #2(8) | 3 b #2(8) #1(16) |
| 5 | D + Clostridial | 15 | 3 * #2(8) 1(12) | 3 b,c #2(16) #1(24) |
| 6 | D + FMD | 15 | 1 * #1(8) | 1 *,b #1(8) |
| 7 | A (1a dose) + D (2a dose) | 15 | 2 * #2(8) | 5 *,c #1(8) #1(12) 3(16) |
| Control | Non-vaccinated | 15 | 0 * | 0 *,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Brun, L.; Leites, M.; Furtado, A.; Campos, F.; Roehe, P.; Puentes, R. Field Evaluation of Commercial Vaccines against Infectious Bovine Rhinotracheitis (Ibr) Virus Using Different Immunization Protocols. Vaccines 2021, 9, 408. https://doi.org/10.3390/vaccines9040408
De Brun L, Leites M, Furtado A, Campos F, Roehe P, Puentes R. Field Evaluation of Commercial Vaccines against Infectious Bovine Rhinotracheitis (Ibr) Virus Using Different Immunization Protocols. Vaccines. 2021; 9(4):408. https://doi.org/10.3390/vaccines9040408
Chicago/Turabian StyleDe Brun, Laureana, Mauro Leites, Agustín Furtado, Fabricio Campos, Paulo Roehe, and Rodrigo Puentes. 2021. "Field Evaluation of Commercial Vaccines against Infectious Bovine Rhinotracheitis (Ibr) Virus Using Different Immunization Protocols" Vaccines 9, no. 4: 408. https://doi.org/10.3390/vaccines9040408
APA StyleDe Brun, L., Leites, M., Furtado, A., Campos, F., Roehe, P., & Puentes, R. (2021). Field Evaluation of Commercial Vaccines against Infectious Bovine Rhinotracheitis (Ibr) Virus Using Different Immunization Protocols. Vaccines, 9(4), 408. https://doi.org/10.3390/vaccines9040408