
The Adjuvants Polyphosphazene (PCEP) and a Combination of Curdlan Plus Leptin Promote a Th17-Type Immune Response to an Intramuscular Vaccine in Mice

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
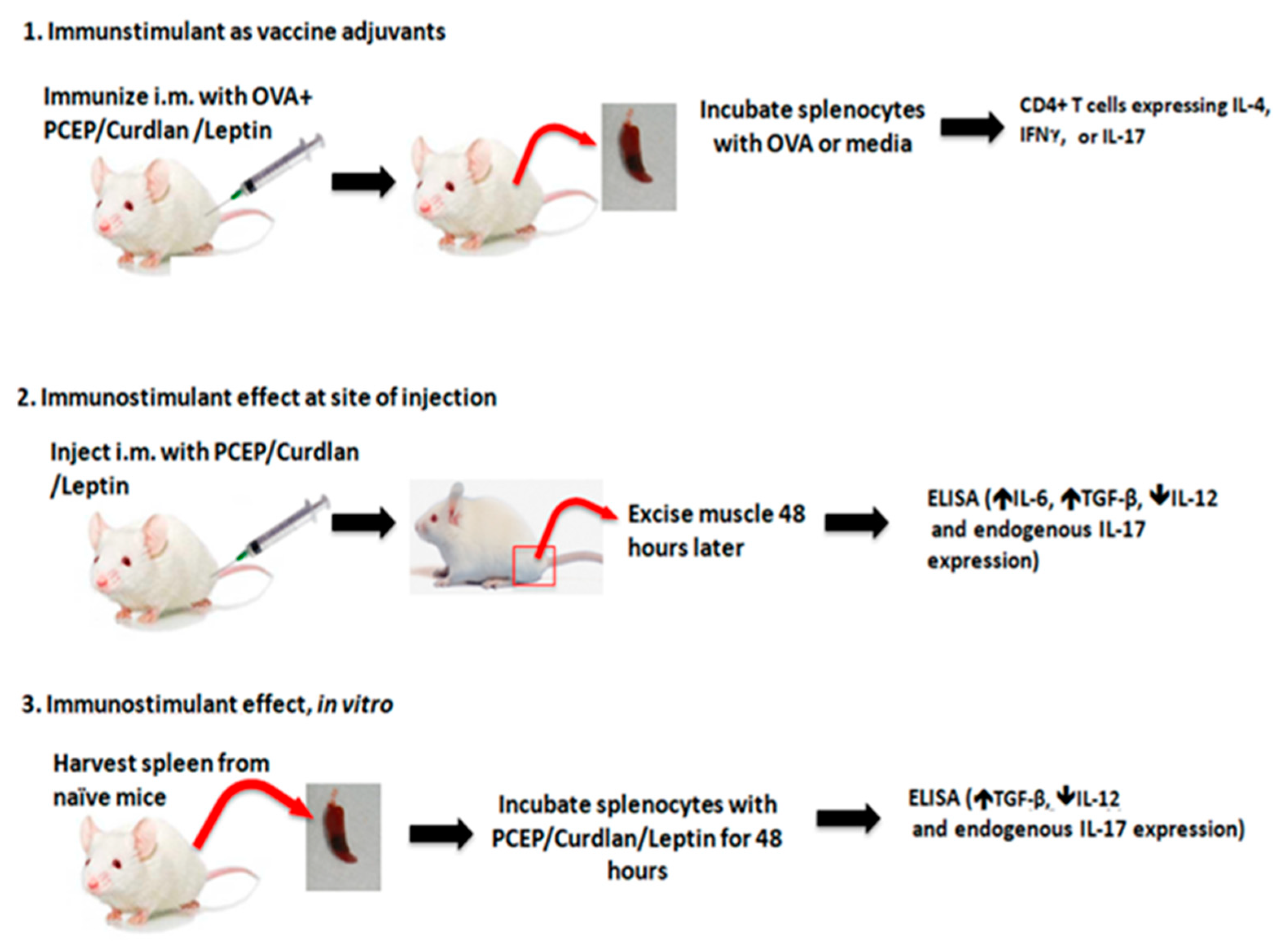
2.1. Animal Experiments
Mouse Immunization, Intramuscular Injections, and ex vivo Splenocytes Stimulations
2.2. Vaccine Trial
2.3. Ex Vivo Response to Immunostimulants
Isolation of Muscle Tissue
2.4. Isolation of Splenocytes
2.5. Cytokine Detection by ELISA in Mice
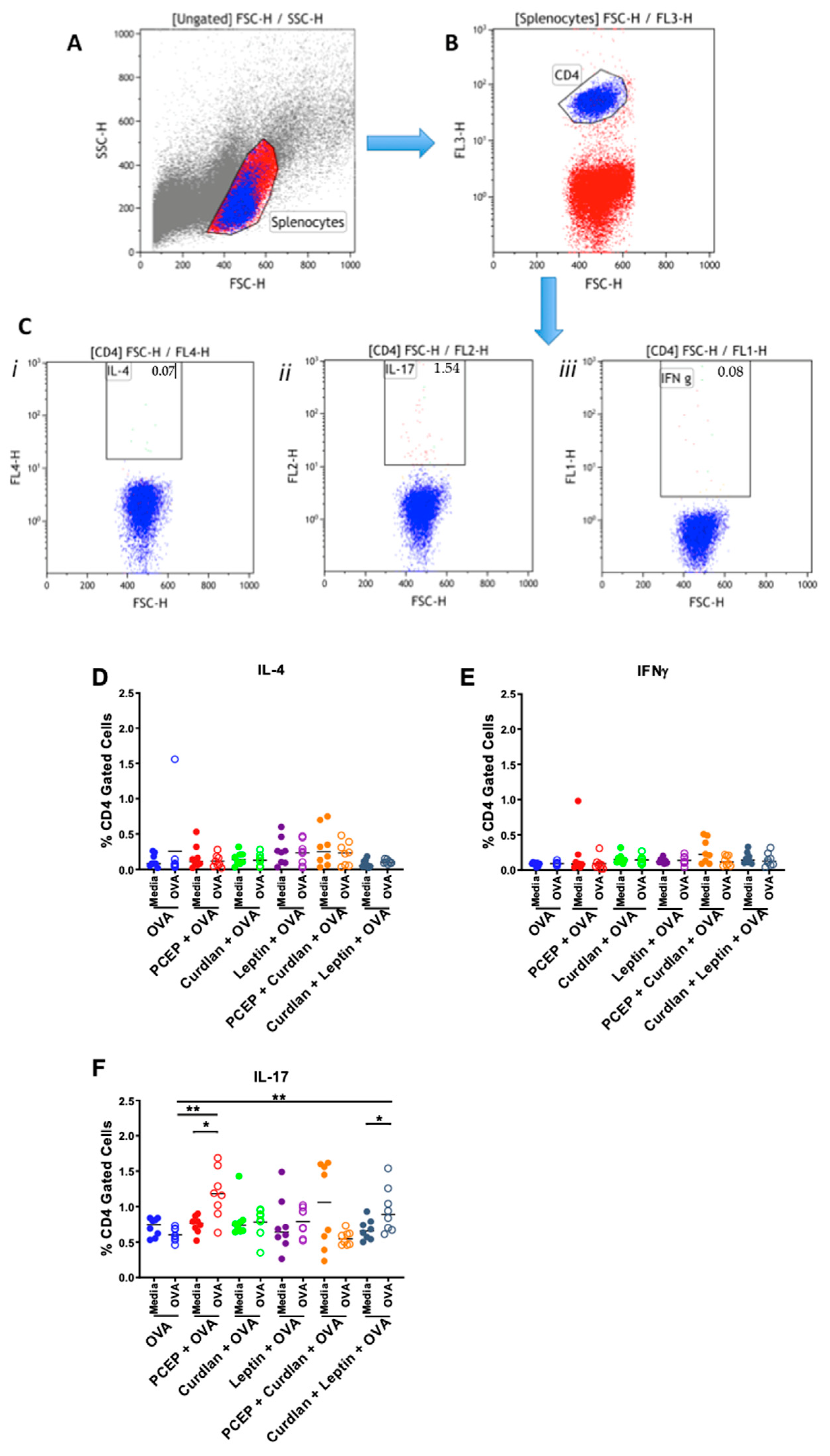
2.6. Splenocyte T cells Immunotyping Using Intracellular Flow Cytometry
2.7. Statistical Analyses
3. Results
3.1. CMI Response to Intramuscular OVA Vaccine Formulated with PCEP, Curdlan, and Leptin Adjuvants
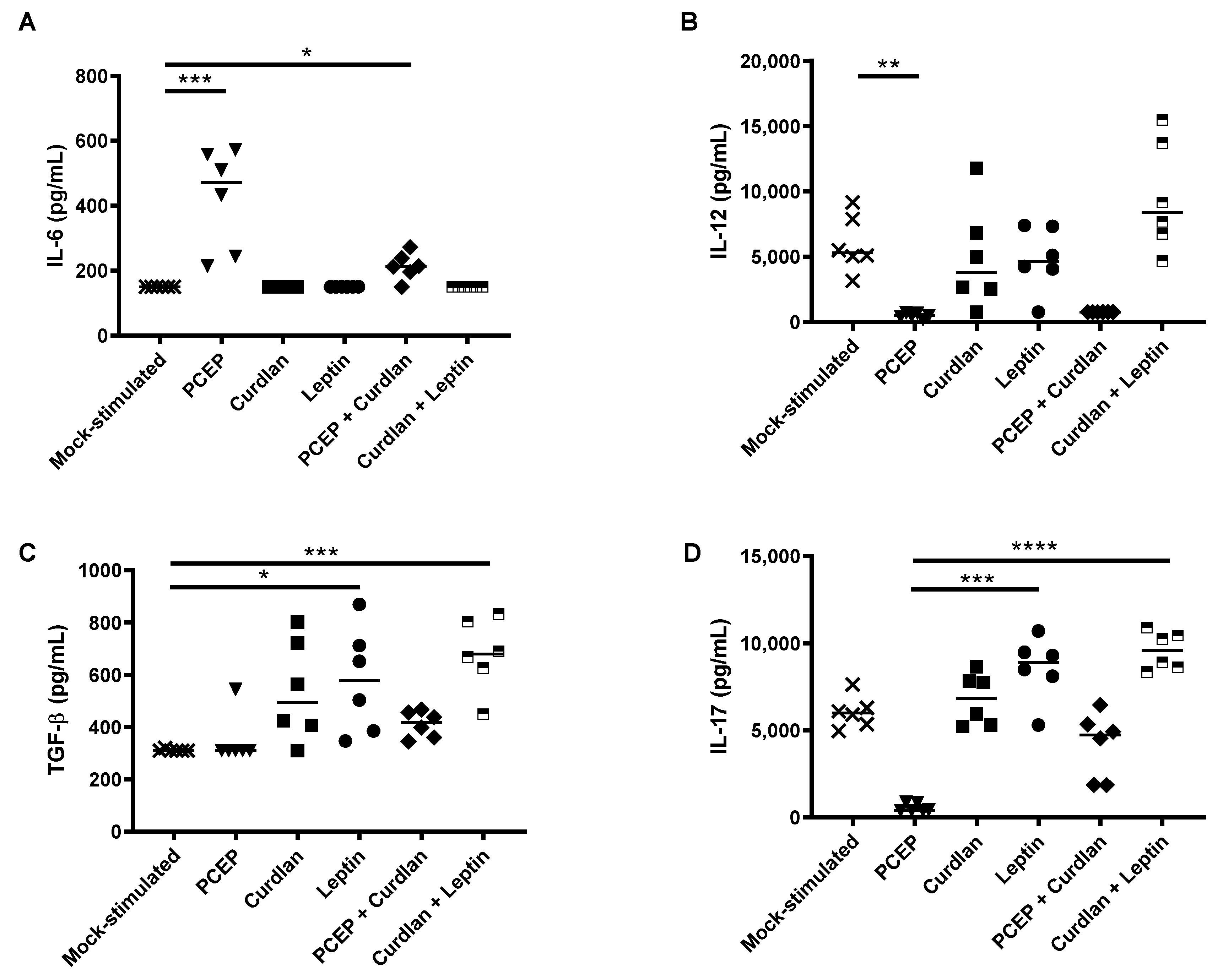
3.2. Local Cytokine Production in Response to PCEP, Curdlan, and/or Leptin Injected into Mouse Muscle
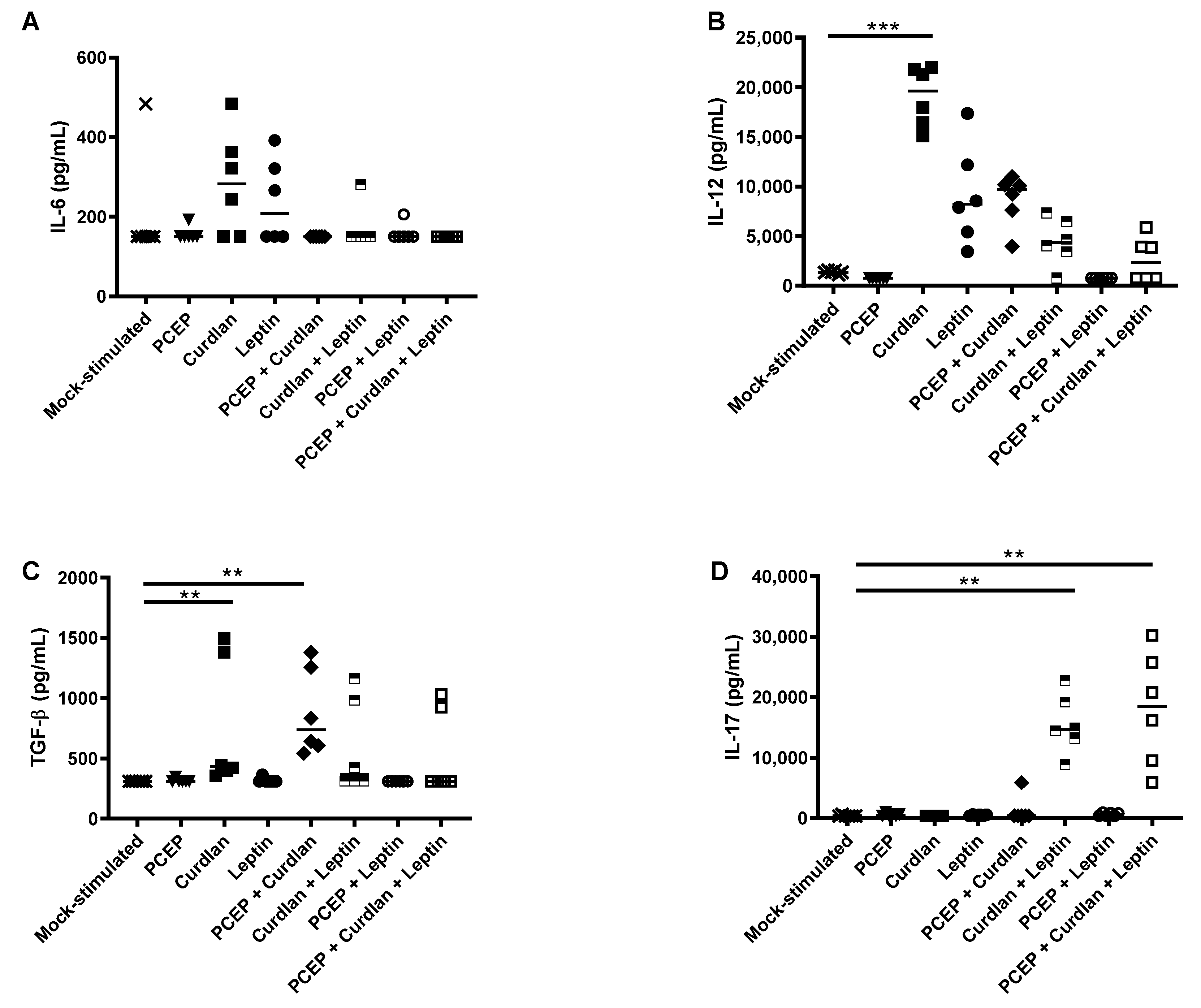
3.3. Primary Splenocyte Cytokine Response to Acute Immunostimulants Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romagnani, S. T-cell subsets (Th1 versus Th2). Ann. Allergy Asthma Immunol. 2020, 85, 9–18, quiz 18, 21. [Google Scholar] [CrossRef]
- Bettelli, E.; Oukka, M.; Kuchroo, V.K. T(H)-17 cells in the circle of immunity and autoimmunity. Nat. Immunol. 2007, 8, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Gurney, A.L. IL-17: Prototype member of an emerging cytokine family. J. Leukoc. Biol. 2002, 71, 1–8. [Google Scholar]
- Capone, A.; Volpe, E. Transcriptional Regulators of T Helper 17 Cell Differentiation in Health and Autoimmune Diseases. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yamasaki, R.; Yamaguchi, H.; Nagata, S.; Une, H.; Cui, Y.; Masaki, K.; Nakamuta, Y.; Iinuma, K.; Watanabe, M.; et al. Oligodendroglial connexin 47 regulates neuroinflammation upon autoimmune demyelination in a novel mouse model of multiple sclerosis. Proc. Natl. Acad. Sci. USA 2020, 117, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Littman, D.R. New traffic light on Th17 Avenue. Cell Res. 2017, 28, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Zeng, J.; Yuan, J.; Deng, X.; Huang, Y.; Chen, L.; Zhang, P.; Feng, H.; Liu, Z.; Wang, Z.; et al. MicroRNA-210 overexpression promotes psoriasis-like inflammation by inducing Th1 and Th17 cell differentiation. J. Clin. Investig. 2018, 128, 2551–2568. [Google Scholar] [CrossRef]
- Xie, A.; Robles, R.J.; Mukherjee, S.; Zhang, H.; Feldbrügge, L.; Csizmadia, E.; Wu, Y.; Enjyoji, K.; Moss, A.C.; Otterbein, L.E.; et al. HIF-1α-induced xenobiotic transporters promote Th17 responses in Crohn’s disease. J. Autoimmun. 2018, 94, 122–133. [Google Scholar] [CrossRef]
- Happel, K.I.; Zheng, M.; Young, E.; Quinton, L.J.; Lockhart, E.; Ramsay, A.J.; Shellito, J.E.; Schurr, J.R.; Bagby, G.J.; Nelson, S.; et al. Cutting edge: Roles of Toll-like receptor 4 and IL-23 in IL-17 expression in response to Klebsiella pneumoniae infection. J. Immunol. 2003, 170, 4432–4436. [Google Scholar] [CrossRef] [Green Version]
- Warfel, J.M.; Merkel, T.J. Bordetella pertussis infection induces a mucosal IL-17 response and long-lived Th17 and Th1 immune memory cells in nonhuman primates. Mucosal Immunol. 2013, 6, 787–796. [Google Scholar] [CrossRef]
- Ritter, K.; Sodenkamp, J.C.; Hölscher, A.; Behrends, J.; Hölscher, C. IL-6 Is Not Absolutely Essential for the Development of a TH17 Immune Response after an Aerosol Infection with Mycobacterium tuberculosis H37rv. Cells 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Yang, F.; Wang, Y.; Liao, Y.; Zhang, J.; Zeng, H.; Zou, Q.; Gu, J. Vaccination with a recombinant OprL fragment induces a Th17 response and confers serotype-independent protection against Pseudomonas aeruginosa infection in mice. Clin. Immunol. 2017, 183, 354–363. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Yang, Y.; Yin, Q.; Wang, Y.; Li, Y.; Wang, C.; Wong, S.M.; Wang, Y.; Goldfine, H.; et al. Recognition of conserved antigens by Th17 cells provides broad protection against pulmonaryHaemophilus influenzaeinfection. Proc. Natl. Acad. Sci. USA 2018, 115, E7149–E7157. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.R.; Tsoni, S.V.; Willment, J.A.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 is required for beta-glucan recognition and control of fungal infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Kang, H.S.; Kim, B.S. Th17 cells enhance viral persistence and inhibit T cell cytotoxicity in a model of chronic virus infection. J. Exp. Med. 2009, 206, 313–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Weaver, C.T.; Hatton, R.D.; Mangan, P.R.; Harrington, L.E. IL-17 Family Cytokines and the Expanding Diversity of Effector T Cell Lineages. Annu. Rev. Immunol. 2007, 25, 821–852. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.K.; Sabet, M.; Nguyen, K.P.L.; Valdez, P.A.; Gonzalez-Navajas, J.M.; Islam, S.; Mihajlov, I.; Fierer, J.; Insel, P.A.; Webster, N.J.; et al. Mucosal adjuvant activity of cholera toxin requires Th17 cells and protects against inhalation anthrax. Proc. Natl. Acad. Sci. USA 2010, 107, 10638–10643. [Google Scholar] [CrossRef] [Green Version]
- Mutwiri, G.; Gerdts, V.; Lopez, M.; Babiuk, L.A. Innate immunity and new adjuvants. Rev. Sci. Tech. IOE 2007, 26, 147–156. [Google Scholar] [CrossRef]
- Mutwiri, G.; Gerdts, V.; van Drunen Littel-van den Hurk, S.; Auray, G.; Eng, N.; Garlapati, S.; Babiuk, L.A.; Potter, A. Combination adjuvants: The next generation of adjuvants? Expert Rev. Vaccines 2011, 10, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Mutwiri, G.; Benjamin, P.; Soita, H.; Townsend, H.; Yost, R.; Roberts, B.; Andrianov, A.K.; Babiuk, L.A. Poly [di(sodium carboxylatoethylphenoxy)phosphazene] (PCEP) is a potent enhancer of mixed Th1/Th2 immune responses in mice immunized with influenza virus antigens. Vaccine 2007, 25, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Awate, S.; Wilson, H.L.; Lai, K.; Babiuk, L.A.; Mutwiri, G. Activation of adjuvant core response genes by the novel adjuvant PCEP. Mol. Immunol. 2012, 51, 292–303. [Google Scholar] [CrossRef]
- Mutwiri, G.; Benjamin, P.; Soita, H.; Babiuk, L.A. Co-administration of polyphosphazenes with CpG oligodeoxynucleotides strongly enhances immune responses in mice immunized with Hepatitis B virus surface antigen. Vaccine 2008, 26, 2680–2688. [Google Scholar] [CrossRef]
- Awate, S.; Wilson, H.L.; Singh, B.; Babiuk, L.A.; Mutwiri, G. The adjuvant PCEP induces recruitment of myeloid and lymphoid cells at the injection site and draining lymph node. Vaccine 2014, 32, 2420–2427. [Google Scholar] [CrossRef]
- Awate, S.; Eng, N.F.; Gerdts, V.; Babiuk, L.A.; Mutwiri, G. Caspase-1 Dependent IL-1beta Secretion and Antigen-Specific T-Cell Activation by the Novel Adjuvant, PCEP. Vaccines 2014, 2, 500–514. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Wentrup, F.; Cambi, A.; Adema, G.J.; Figdor, C.G. “Sweet talk”: Closing in on C type lectin signaling. Immunity 2005, 22, 399–400. [Google Scholar] [CrossRef] [Green Version]
- Goyal, S.; Castrillón-Betancur, J.C.; Klaile, E.; Slevogt, H. The Interaction of Human Pathogenic Fungi With C-Type Lectin Receptors. Front. Immunol. 2018, 9, 1261. [Google Scholar] [CrossRef] [Green Version]
- Osorio, F.; LeibundGut-Landmann, S.; Lochner, M.; Lahl, K.; Sparwasser, T.; Eberl, G.; Sousa, C.R.E. DC activated via dectin-1 convert Treg into IL-17 producers. Eur. J. Immunol. 2008, 38, 3274–3281. [Google Scholar] [CrossRef]
- Martínez, A.; Bono, C.; Gozalbo, D.; Goodridge, H.S.; Gil, M.L.; Yáñez, A. TLR2 and Dectin-1 Signaling in Mouse Hematopoietic Stem and Progenitor Cells Impacts the Ability of the Antigen Presenting Cells They Produce to Activate CD4 T Cells. Cells 2020, 9, 1317. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, X.; Cheng, Y.; Li, J.; Xiao, Y.; Zhang, Q.; Zong, A.; Zhong, C.; Wang, F. Preparation and in vitro immunomodulatory effect of curdlan sulfate. Carbohydr. Polym. 2014, 102, 852–861. [Google Scholar] [CrossRef]
- LeibundGut-Landmann, S.; Gross, O.; Robinson, M.J.; Osorio, F.; Slack, E.C.; Tsoni, S.V.; Schweighoffer, E.; Tybulewicz, V.L.J.; Brown, G.D.; Ruland, J.; et al. Syk- and CARD9-dependent coupling of innate immunity to the induction of T helper cells that produce interleukin 17. Nat. Immunol. 2007, 8, 630–638. [Google Scholar] [CrossRef]
- Agrawal, S.; Gupta, S.; Agrawal, A. Human Dendritic Cells Activated via Dectin-1 Are Efficient at Priming Th17, Cytotoxic CD8 T and B Cell Responses. PLoS ONE 2010, 5, e13418. [Google Scholar] [CrossRef]
- Schaefers, M.M.; Duan, B.; Mizrahi, B.; Lu, R.; Reznor, G.; Kohane, D.S.; Priebe, G.P. PLGA-encapsulation of the Pseudomonas aeruginosa PopB vaccine antigen improves Th17 responses and confers protection against experimental acute pneumonia. Vaccine 2018, 36, 6926–6932. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nat. Cell Biol. 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gómez-Reino, J.J.; Mera, J.J.G.-R.A.; Lago, F.; Gómez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef]
- La Cava, A. Leptin in inflammation and autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Procaccini, C.; Jirillo, E.; Matarese, G. Leptin as an immunomodulator. Mol. Asp. Med. 2012, 33, 35–45. [Google Scholar] [CrossRef]
- Sánchez-Margalet, V.; Martín-Romero, C.; Santos-Alvarez, J.; Goberna, R.; Najib, S.; Gonzalez-Yanes, C. Role of leptin as an immunomodulator of blood mononuclear cells: Mechanisms of action. Clin. Exp. Immunol. 2003, 133, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Dohn, G.L.; Fiedorek, F.T.; Tapscott, E.B.; Coleman, R.A. Leptin Directly Alters Lipid Partitioning in Skeletal Muscle. Diabetes 1997, 46, 1360–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margetic, S.; Gazzola, C.; Pegg, G.; Hill, R. Leptin: A review of its peripheral actions and interactions. Int. J. Obes. 2002, 26, 1407–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Liu, Y.; Shi, F.D.; Zou, H.; Matarese, G.; La Cava, A. Cutting edge: Leptin-induced RORgammat expression in CD4+ T cells promotes Th17 responses in systemic lupus erythematosus. J. Immunol. 2013, 190, 3054–3058. [Google Scholar] [CrossRef] [Green Version]
- La Cava, A.; Matarese, G. The weight of leptin in immunity. Nat. Rev. Immunol. 2004, 4, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Reis, B.S.; Lee, K.; Fanok, M.H.; Mascaraque, C.; Amoury, M.; Cohn, L.B.; Rogoz, A.; Dallner, O.S.; Moraes-Vieira, P.M.; Domingos, A.I.; et al. Leptin Receptor Signaling in T Cells Is Required for Th17 Differentiation. J. Immunol. 2015, 194, 5253–5260. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Chen, H.; Li, X.; Wei, J. Leptin: An unappreciated key player in SLE. Clin. Rheumatol. 2020, 39, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Chaffey, A.M. Identification of Immunostimulatory Adjuvants(s) that Will Promote a Th17-Type of Response. [Master’s Thesis, University of Saskatchewan], Saskatoon, SK, Canada. HARVEST. December 2017. Available online: http://hdl.handle.net/10388/7959 (accessed on 2 February 2021).
- Baca-Estrada, M.E.; Snider, M.; Tikoo, S.K.; Harland, R.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Immunogenicity of bovine herpesvirus 1 glycoprotein D in mice: Effect of antigen form on the induction of cellular and humoral immune responses. Viral. Immunol. 1996, 9, 11–22. [Google Scholar] [CrossRef]
- Magiri, R.; Lai, K.; Chaffey, A.; Wilson, H.; Berry, W.; Szafron, M.; Mutwiri, G. Response of immune response genes to adjuvants poly [di(sodium carboxylatoethylphenoxy)phosphazene] (PCEP), CpG oligodeoxynucleotide and emulsigen at intradermal injection site in pigs. Vet. Immunol. Immunopathol. 2016, 175, 57–63. [Google Scholar] [CrossRef]
- Acosta-Rodriguez, E.V.; Rivino, L.; Geginat, J.; Jarrossay, D.; Gattorno, M.; Lanzavecchia, A.; Sallusto, F.; Napolitani, G. Surface phenotype and antigenic specificity of human interleukin 17–producing T helper memory cells. Nat. Immunol. 2007, 8, 639–646. [Google Scholar] [CrossRef]
- Werner, J.L.; Gessner, M.A.; Lilly, L.M.; Nelson, M.P.; Metz, A.E.; Horn, D.; Dunaway, C.W.; Deshane, J.; Chaplin, D.D.; Weaver, C.T.; et al. Neutrophils Produce Interleukin 17A (IL-17A) in a Dectin-1- and IL-23-Dependent Manner during Invasive Fungal Infection. Infect. Immun. 2011, 79, 3966–3977. [Google Scholar] [CrossRef] [Green Version]
- Duluc, D.; Joo, H.; Ni, L.; Yin, W.; Upchurch, K.; Li, D.; Xue, Y.; Klucar, P.; Zurawski, S.; Zurawski, G.; et al. Induction and Activation of Human Th17 by Targeting Antigens to Dendritic Cells via Dectin-1. J. Immunol. 2014, 192, 5776–5788. [Google Scholar] [CrossRef] [Green Version]
- Rogers, N.C.; Slack, E.C.; Edwards, A.D.; Nolte, M.A.; Schulz, O.; Schweighoffer, E.; Williams, D.L.; Gordon, S.; Tybulewicz, V.L.; Brown, G.D.; et al. Syk-dependent cytokine induction by Dectin-1 reveals a novel pattern recognition pathway for C type lectins. Immunity 2005, 22, 507–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattioli, B.; Giordani, L.; Quaranta, M.G.; Viora, M. Leptin exerts an anti-apoptotic effect on human dendritic cells via the PI3K-Akt signaling pathway. FEBS Lett. 2009, 583, 1102–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Romero, C.; Santos-Alvarez, J.; Goberna, R.; Sánchez-Margalet, V. Human Leptin Enhances Activation and Proliferation of Human Circulating T Lymphocytes. Cell. Immunol. 2000, 199, 15–24. [Google Scholar] [CrossRef]
- Matarese, G.; De Rosa, V.; La Cava, A. Regulatory CD4 T cells: Sensing the environment. Trends Immunol. 2008, 29, 12–17. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, V.; Procaccini, C.; Calì, G.; Pirozzi, G.; Fontana, S.; Zappacosta, S.; La Cava, A.; Matarese, G. A Key Role of Leptin in the Control of Regulatory T Cell Proliferation. Immunity 2007, 26, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Takaku, M.; Zou, L.; Gu, A.-D.; Chou, W.-C.; Zhang, G.; Wu, B.; Kong, Q.; Thomas, S.Y.; Serody, J.S.; et al. Reversing SKI–SMAD4-mediated suppression is essential for TH17 cell differentiation. Nat. Cell Biol. 2017, 551, 105–109. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, G.; Wan, Y.Y. SKI and SMAD4 are essential for IL-21-induced Th17 differentiation. Mol. Immunol. 2019, 114, 260–268. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Tejedor, G.; Ipseiz, N.; Pène, J.; Morand, E.; Beaulieu, E.; Jorgensen, C.; Noël, D.; Djouad, F. A8.11 gilz-dependent activin a production by MSC inhibits TH17 differentiation. Ann. Rheum. Dis. 2014, 73, A80. [Google Scholar] [CrossRef]
- Ghoreschi, K.; Laurence, A.; Yang, X.-P.; Tato, C.M.; McGeachy, M.J.; Konkel, J.E.; Ramos, H.L.; Wei, L.; Davidson, T.S.; Bouladoux, N.; et al. Generation of pathogenic TH17 cells in the absence of TGF-β signalling. Nat. Cell Biol. 2010, 467, 967–971. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S. The role of transforming growth factorβ in T helper 17 differentiation. Immunology 2018, 155, 24–35. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Name of Antibody | Isotype | Concentration |
|---|---|---|---|
| CD4 | PerCP-Cy™5.5 labeled Rat Anti-Mouse CD4 | Rat (DA) IgG2a, κ | 0.0625 μg/test |
| CD8 | PE labeled Rat Anti-Mouse CD8b.2 | Rat (LOU) IgG1, κ | 0.25 μg/test |
| IL-4 | APC labeled Rat Anti-Mouse IL-4 | Rat IgG1 | 0.25 μg/test |
| IL-17 | PE labeled Rat Anti-Mouse IL-17A | Rat IgG1, κ | 0.25 μg/test |
| IFNγ | FITC labeled Rat Anti-Mouse IFNγ | Rat IgG1, κ | 0.25 μg/test |
| Cocktail | Antibodies and Fluorochrome |
|---|---|
| Cocktail 1 (C1) | PerCP-Cy™5.5 labeled Rat Anti-Mouse CD4 antibody |
| APC labeled Rat Anti-Mouse IL-4 antibody | |
| PE labeled Rat Anti-Mouse IL-17A antibody FITC labeled Rat Anti-Mouse IFNγ antibody | |
| Cocktail 2 (C2) | PerCP-Cy™5.5 labeled Rat Anti-Mouse CD4 antibody |
| PE labeled Rat Anti-Mouse CD8b.2 antibody | |
| FITC labeled Rat Anti-Mouse IFNγ antibody |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaffey, A.; Hamonic, G.; Chand, D.; Mutwiri, G.K.; Wilson, H.L. The Adjuvants Polyphosphazene (PCEP) and a Combination of Curdlan Plus Leptin Promote a Th17-Type Immune Response to an Intramuscular Vaccine in Mice. Vaccines 2021, 9, 507. https://doi.org/10.3390/vaccines9050507
Chaffey A, Hamonic G, Chand D, Mutwiri GK, Wilson HL. The Adjuvants Polyphosphazene (PCEP) and a Combination of Curdlan Plus Leptin Promote a Th17-Type Immune Response to an Intramuscular Vaccine in Mice. Vaccines. 2021; 9(5):507. https://doi.org/10.3390/vaccines9050507
Chicago/Turabian StyleChaffey, Alyssa, Glenn Hamonic, Dylan Chand, George K. Mutwiri, and Heather L. Wilson. 2021. "The Adjuvants Polyphosphazene (PCEP) and a Combination of Curdlan Plus Leptin Promote a Th17-Type Immune Response to an Intramuscular Vaccine in Mice" Vaccines 9, no. 5: 507. https://doi.org/10.3390/vaccines9050507
APA StyleChaffey, A., Hamonic, G., Chand, D., Mutwiri, G. K., & Wilson, H. L. (2021). The Adjuvants Polyphosphazene (PCEP) and a Combination of Curdlan Plus Leptin Promote a Th17-Type Immune Response to an Intramuscular Vaccine in Mice. Vaccines, 9(5), 507. https://doi.org/10.3390/vaccines9050507