
Predictive Markers of Immunogenicity and Efficacy for Human Vaccines


 , , , and
, , , and
Abstract
:1. Introduction
2. Identification of Biomarkers and Signatures of Vaccine Responses
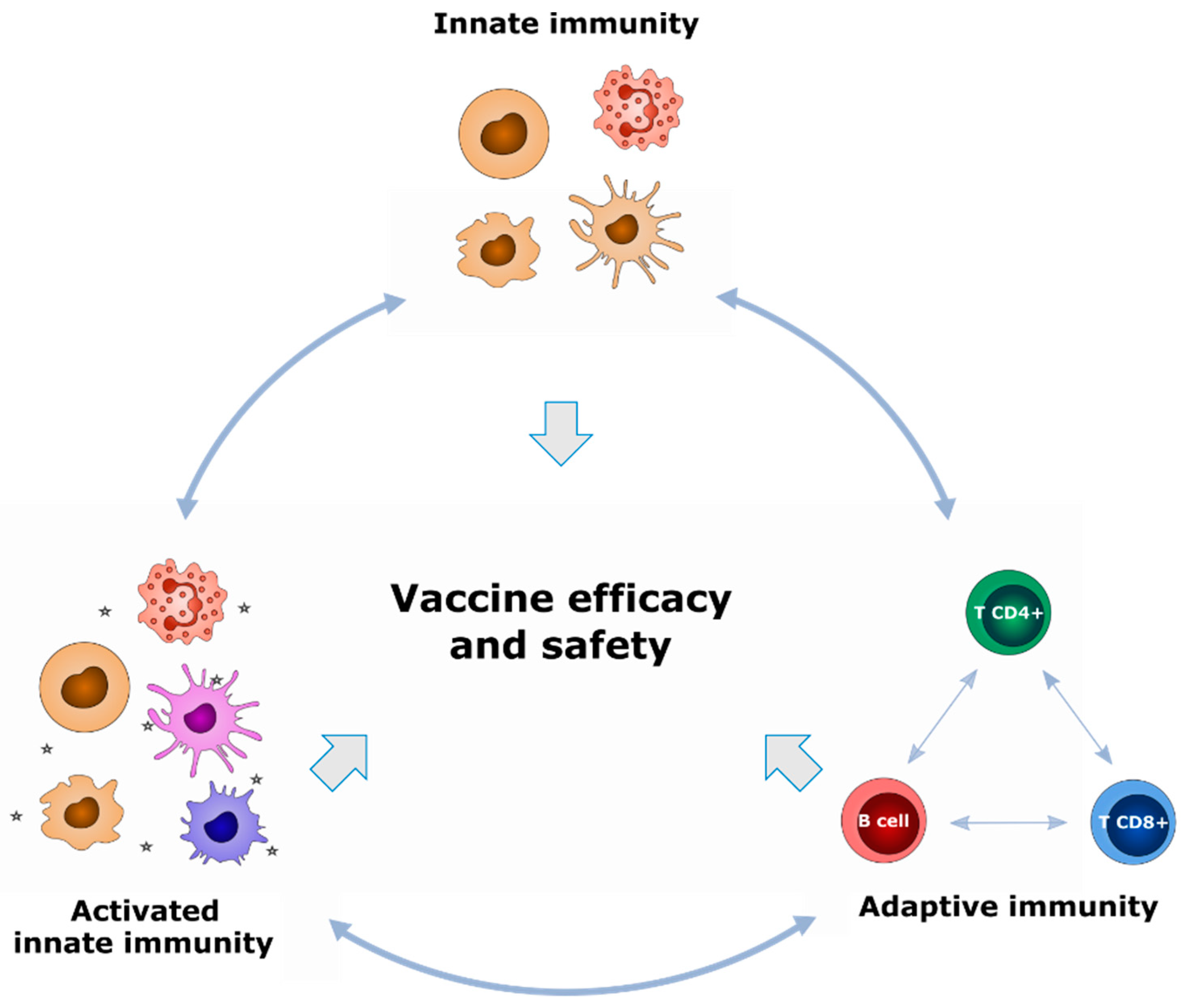
2.1. What Types of Biomarkers Can Be Used to Define Vaccine Signatures?
2.2. From What Samples Can We Identify Vaccine Response Biomarkers?
2.3. At What Time Should We Identify Vaccine Response Biomarkers?
3. Conventional High-Throughput Technologies to Assess Vaccine Responses
3.1. High-Dimensional Flow and Mass Cytometry
3.2. Cytokine Profiling
3.3. OMICS Technologies
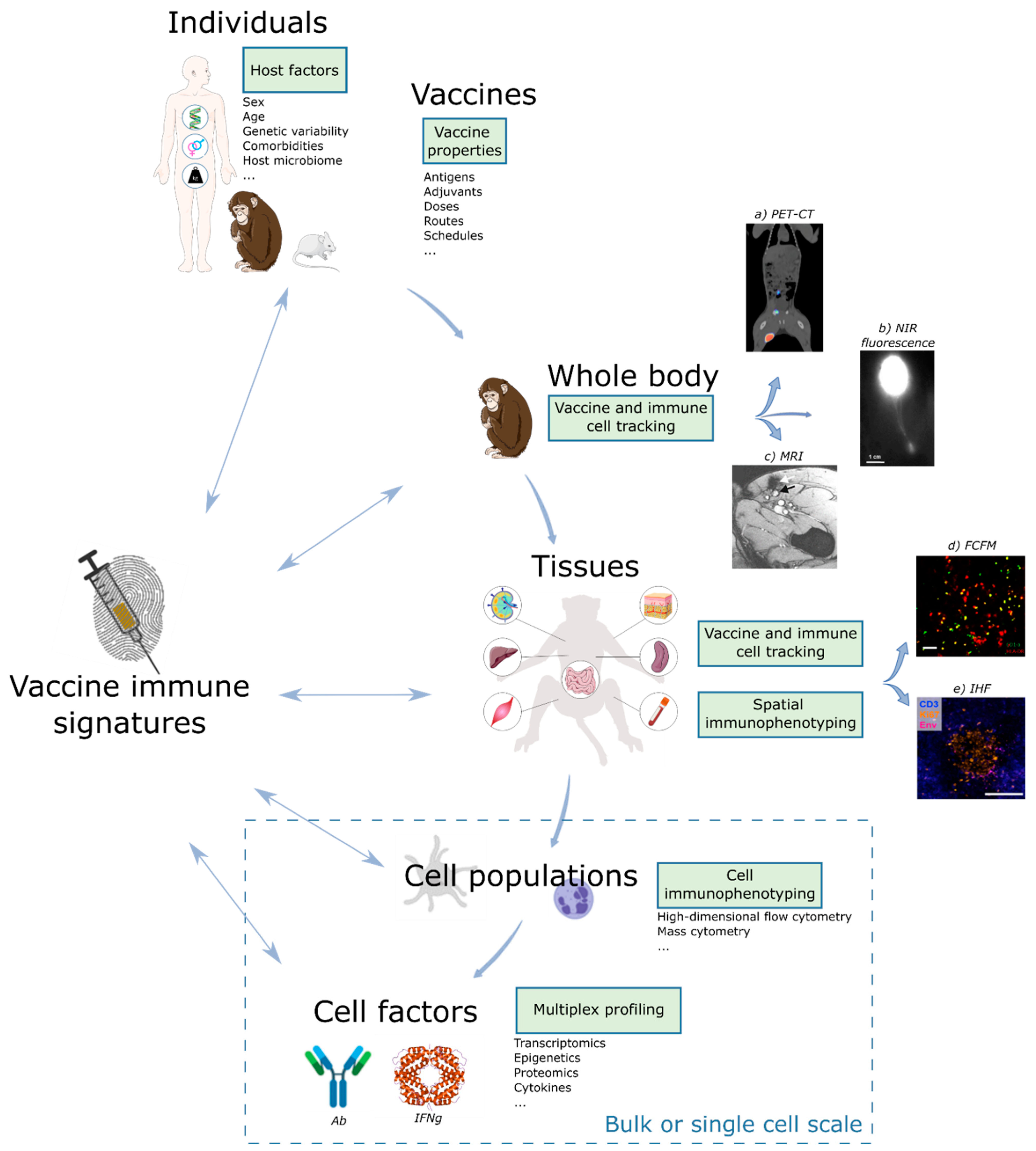
4. Imaging Technologies to Refine and Expand Vaccine Signatures
4.1. In Vivo Imaging of Vaccine Trafficking and the Immune Response
4.1.1. Whole Body Imaging of Vaccine Distribution and the Immune Response
4.1.2. In Vivo Microscopic Imaging of the Interactions between Vaccines and Immune Cells
4.2. Ex Vivo Multiparametric Analyses
5. Bioinformatics and Statistical Tools to Build Predictive Models of Vaccine Responses
5.1. Analysis of High-Dimensional Biological Data
5.2. Machine Learning and In Silico Models
6. Vaccine Signatures in Preclinical Models to Improve Human Vaccination Strategies
6.1. Defining New Correlates of Protection
6.2. Stepping up Personalized Vaccinology
6.3. Improving Vaccine Formulation and Administration
6.4. Deciphering Mechanisms That Underly Immune Protective Responses
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heaton, P.M. Challenges of Developing Novel Vaccines With Particular Global Health Importance. Front. Immunol. 2020, 11, 517290. [Google Scholar] [CrossRef]
- World Heath Organization (WHO). Global Vaccine Action Plan 2011–2020. Available online: https://www.who.int/publications/i/item/global-vaccine-action-plan-2011–2020 (accessed on 20 May 2021).
- Trovato, M.; Sartorius, R.; D’Apice, L.; Manco, R.; De Berardinis, P. Viral Emerging Diseases: Challenges in Developing Vaccination Strategies. Front. Immunol. 2020, 11, 2130. [Google Scholar] [CrossRef] [PubMed]
- Montalvo Zurbia-Flores, G.; Rollier, C.S.; Reyes-Sandoval, A. Re-thinking yellow fever vaccines: Fighting old foes with new generation vaccines. Hum. Vaccines Immunother. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Group, B.D.W. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharm. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Galassie, A.C.; Link, A.J. Proteomic contributions to our understanding of vaccine and immune responses. Proteom. Clin. Appl. 2015, 9, 972–989. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J.; Wang, S.; Jia, M.; Li, D.; Liu, Y.; Li, Z.; Zhu, H.; Xu, H.; Sun, M.; Lu, L.; et al. A Systems Vaccinology Approach Reveals Temporal Transcriptomic Changes of Immune Responses to the Yellow Fever 17D Vaccine. J. Immunol. 2017, 199, 1476–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakaya, H.I.; Clutterbuck, E.; Kazmin, D.; Wang, L.; Cortese, M.; Bosinger, S.E.; Patel, N.B.; Zak, D.E.; Aderem, A.; Dong, T.; et al. Systems biology of immunity to MF59-adjuvanted versus nonadjuvanted trivalent seasonal influenza vaccines in early childhood. Proc. Natl. Acad. Sci. USA 2016, 113, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, H.I.; Hagan, T.; Duraisingham, S.S.; Lee, E.K.; Kwissa, M.; Rouphael, N.; Frasca, D.; Gersten, M.; Mehta, A.K.; Gaujoux, R.; et al. Systems Analysis of Immunity to Influenza Vaccination across Multiple Years and in Diverse Populations Reveals Shared Molecular Signatures. Immunity 2015, 43, 1186–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogorelyy, M.V.; Minervina, A.A.; Touzel, M.P.; Sycheva, A.L.; Komech, E.A.; Kovalenko, E.I.; Karganova, G.G.; Egorov, E.S.; Komkov, A.Y.; Chudakov, D.M.; et al. Precise tracking of vaccine-responding T cell clones reveals convergent and personalized response in identical twins. Proc. Natl. Acad. Sci. USA 2018, 115, 12704–12709. [Google Scholar] [CrossRef] [Green Version]
- Rechtien, A.; Richert, L.; Lorenzo, H.; Martrus, G.; Hejblum, B.; Dahlke, C.; Kasonta, R.; Zinser, M.; Stubbe, H.; Matschl, U.; et al. Systems Vaccinology Identifies an Early Innate Immune Signature as a Correlate of Antibody Responses to the Ebola Vaccine rVSV-ZEBOV. Cell Rep. 2017, 20, 2251–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, C.E.; White, J.M.; Viola, N.T.; Gibson, H.M. In vivo Imaging Technologies to Monitor the Immune System. Front. Immunol. 2020, 11, 1067. [Google Scholar] [CrossRef] [PubMed]
- Rashidian, M.; Keliher, E.J.; Bilate, A.M.; Duarte, J.N.; Wojtkiewicz, G.R.; Jacobsen, J.T.; Cragnolini, J.; Swee, L.K.; Victora, G.D.; Weissleder, R.; et al. Noninvasive imaging of immune responses. Proc. Natl. Acad. Sci. USA 2015, 112, 6146–6151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.L.; Poland, G.A. Personalized vaccinology: One size and dose might not fit both sexes. Vaccine 2013, 31, 2599–2600. [Google Scholar] [CrossRef]
- Kennedy, R.B.; Ovsyannikova, I.G.; Pankratz, V.S.; Vierkant, R.A.; Jacobson, R.M.; Ryan, M.A.; Poland, G.A. Gender effects on humoral immune responses to smallpox vaccine. Vaccine 2009, 27, 3319–3323. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.L.; Hodgson, A.; Robinson, D.P. Mechanisms of sex disparities in influenza pathogenesis. J. Leukoc. Biol. 2012, 92, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyanja, E.; Ssemaganda, A.; Ngauv, P.; Cubas, R.; Perrin, H.; Srinivasan, D.; Canderan, G.; Lawson, B.; Kopycinski, J.; Graham, A.S.; et al. Immune activation alters cellular and humoral responses to yellow fever 17D vaccine. J. Clin. Investig. 2014, 124, 3147–3158. [Google Scholar] [CrossRef] [Green Version]
- Omersel, J.; Karas Kuželički, N. Vaccinomics and Adversomics in the Era of Precision Medicine: A Review Based on HBV, MMR, HPV, and COVID-19 Vaccines. J. Clin. Med. 2020, 9, 3561. [Google Scholar] [CrossRef]
- Poland, G.A.; Ovsyannikova, I.G.; Jacobson, R.M. Application of pharmacogenomics to vaccines. Pharmacogenomics 2009, 10, 837–852. [Google Scholar] [CrossRef] [Green Version]
- Davila, S.; Froeling, F.E.; Tan, A.; Bonnard, C.; Boland, G.J.; Snippe, H.; Hibberd, M.L.; Seielstad, M. New genetic associations detected in a host response study to hepatitis B vaccine. Genes Immun. 2010, 11, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Zhang, L.; Zhang, W.; Wu, X.; Li, Y.; Yan, B.; Zhu, X.; Liu, X.; Yang, C.; Xu, J.; et al. A genome-wide association study identifies polymorphisms in the HLA-DR region associated with non-response to hepatitis B vaccination in Chinese Han populations. Hum. Mol. Genet. 2014, 23, 2210–2219. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.W.; Chen, C.F.; Lai, S.K.; Lin, H.H.; Chu, C.C.; Wang, L.Y. SNP rs7770370 in HLA-DPB1 loci as a major genetic determinant of response to booster hepatitis B vaccination: Results of a genome-wide association study. J. Gastroenterol. Hepatol. 2015, 30, 891–899. [Google Scholar] [CrossRef]
- Nishida, N.; Sugiyama, M.; Sawai, H.; Nishina, S.; Sakai, A.; Ohashi, J.; Khor, S.S.; Kakisaka, K.; Tsuchiura, T.; Hino, K.; et al. Key HLA-DRB1-DQB1 haplotypes and role of the BTNL2 gene for response to a hepatitis B vaccine. Hepatology 2018, 68, 848–858. [Google Scholar] [CrossRef]
- Scepanovic, P.; Alanio, C.; Hammer, C.; Hodel, F.; Bergstedt, J.; Patin, E.; Thorball, C.W.; Chaturvedi, N.; Charbit, B.; Abel, L.; et al. Human genetic variants and age are the strongest predictors of humoral immune responses to common pathogens and vaccines. Genome Med. 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Mosca, F.; Tritto, E.; Muzzi, A.; Monaci, E.; Bagnoli, F.; Iavarone, C.; O’Hagan, D.; Rappuoli, R.; De Gregorio, E. Molecular and cellular signatures of human vaccine adjuvants. Proc. Natl. Acad. Sci. USA 2008, 105, 10501–10506. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Rouphael, N.; Duraisingham, S.; Romero-Steiner, S.; Presnell, S.; Davis, C.; Schmidt, D.S.; Johnson, S.E.; Milton, A.; Rajam, G.; et al. Molecular signatures of antibody responses derived from a systems biology study of five human vaccines. Nat. Immunol. 2014, 15, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, P.; Curtis, N. Factors That Influence the Immune Response to Vaccination. Clin. Microbiol. Rev. 2019, 32, e00084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerman, M.E.; Barouch, D.H.; Alter, G. Systems serology for evaluation of HIV vaccine trials. Immunol. Rev. 2017, 275, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Van Gils, M.J.; Sanders, R.W. Broadly neutralizing antibodies against HIV-1: Templates for a vaccine. Virology 2013, 435, 46–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.; Hua, Z.; Liu, C.; Lin, L.; Chen, R.; Hou, B. Characterization of T-Dependent and T-Independent B Cell Responses to a Virus-like Particle. J. Immunol. 2017, 198, 3846–3856. [Google Scholar] [CrossRef]
- Chaudhury, S.; Duncan, E.H.; Atre, T.; Storme, C.K.; Beck, K.; Kaba, S.A.; Lanar, D.E.; Bergmann-Leitner, E.S. Identification of Immune Signatures of Novel Adjuvant Formulations Using Machine Learning. Sci. Rep. 2018, 8, 17508. [Google Scholar] [CrossRef] [Green Version]
- Palgen, J.L.; Tchitchek, N.; Rodriguez-Pozo, A.; Jouhault, Q.; Abdelhouahab, H.; Dereuddre-Bosquet, N.; Contreras, V.; Martinon, F.; Cosma, A.; Lévy, Y.; et al. Innate and secondary humoral responses are improved by increasing the time between MVA vaccine immunizations. NPJ Vaccines 2020, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Palgen, J.L.; Tchitchek, N.; Huot, N.; Elhmouzi-Younes, J.; Lefebvre, C.; Rosenbaum, P.; Dereuddre-Bosquet, N.; Martinon, F.; Hocini, H.; Cosma, A.; et al. NK cell immune responses differ after prime and boost vaccination. J. Leukoc. Biol. 2019, 105, 1055–1073. [Google Scholar] [CrossRef]
- Palgen, J.L.; Tchitchek, N.; Elhmouzi-Younes, J.; Delandre, S.; Namet, I.; Rosenbaum, P.; Dereuddre-Bosquet, N.; Martinon, F.; Cosma, A.; Lévy, Y.; et al. Prime and Boost Vaccination Elicit a Distinct Innate Myeloid Cell Immune Response. Sci. Rep. 2018, 8, 3087. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, P.; Tchitchek, N.; Joly, C.; Stimmer, L.; Hocini, H.; Dereuddre-Bosquet, N.; Beignon, A.S.; Chapon, C.; Levy, Y.; Le Grand, R.; et al. Molecular and Cellular Dynamics in the Skin, the Lymph Nodes, and the Blood of the Immune Response to Intradermal Injection of Modified Vaccinia Ankara Vaccine. Front. Immunol. 2018, 9, 870. [Google Scholar] [CrossRef]
- Rosenbaum, P.; Tchitchek, N.; Joly, C.; Rodriguez Pozo, A.; Stimmer, L.; Langlois, S.; Hocini, H.; Gosse, L.; Pejoski, D.; Cosma, A.; et al. Vaccine Inoculation Route Modulates Early Immunity and Consequently Antigen-Specific Immune Response. Front. Immunol. 2021, 12, 1362. [Google Scholar] [CrossRef]
- Wimmers, F.; Pulendran, B. Emerging technologies for systems vaccinology—Multi-omics integration and single-cell (epi)genomic profiling. Curr. Opin. Immunol. 2020, 65, 57–64. [Google Scholar] [CrossRef]
- Natrajan, M.S.; Rouphael, N.; Lai, L.; Kazmin, D.; Jensen, T.L.; Weiss, D.S.; Ibegbu, C.; Sztein, M.B.; Hooper, W.F.; Hill, H.; et al. Systems Vaccinology for a Live Attenuated Tularemia Vaccine Reveals Unique Transcriptional Signatures That Predict Humoral and Cellular Immune Responses. Vaccines 2019, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagan, T.; Cortese, M.; Rouphael, N.; Boudreau, C.; Linde, C.; Maddur, M.S.; Das, J.; Wang, H.; Guthmiller, J.; Zheng, N.Y.; et al. Antibiotics-Driven Gut Microbiome Perturbation Alters Immunity to Vaccines in Humans. Cell 2019, 178, 1313–1328.e1313. [Google Scholar] [CrossRef]
- Lindsay, K.E.; Bhosle, S.M.; Zurla, C.; Beyersdorf, J.; Rogers, K.A.; Vanover, D.; Xiao, P.; Araínga, M.; Shirreff, L.M.; Pitard, B.; et al. Visualization of early events in mRNA vaccine delivery in non-human primates via PET-CT and near-infrared imaging. Nat. Biomed. Eng. 2019, 3, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Salabert, N.; Todorova, B.; Martinon, F.; Boisgard, R.; Zurawski, G.; Zurawski, S.; Dereuddre-Bosquet, N.; Cosma, A.; Kortulewski, T.; Banchereau, J.; et al. Intradermal injection of an anti-Langerin-HIVGag fusion vaccine targets epidermal Langerhans cells in nonhuman primates and can be tracked in vivo. Eur. J. Immunol. 2016, 46, 689–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, I.J.; Lesterhuis, W.J.; Barentsz, J.O.; Verdijk, P.; van Krieken, J.H.; Boerman, O.C.; Oyen, W.J.; Bonenkamp, J.J.; Boezeman, J.B.; Adema, G.J.; et al. Magnetic resonance tracking of dendritic cells in melanoma patients for monitoring of cellular therapy. Nat. Biotechnol. 2005, 23, 1407–1413. [Google Scholar] [CrossRef] [PubMed]
- Todorova, B.; Salabert, N.; Tricot, S.; Boisgard, R.; Rathaux, M.; Le Grand, R.; Chapon, C. Fibered Confocal Fluorescence Microscopy for the Noninvasive Imaging of Langerhans Cells in Macaques. Contrast Media Mol. Imaging 2017, 2017, 3127908. [Google Scholar] [CrossRef] [Green Version]
- Ols, S.; Yang, L.; Thompson, E.A.; Pushparaj, P.; Tran, K.; Liang, F.; Lin, A.; Eriksson, B.; Karlsson Hedestam, G.B.; Wyatt, R.T.; et al. Route of Vaccine Administration Alters Antigen Trafficking but Not Innate or Adaptive Immunity. Cell Rep. 2020, 30, 3964–3971.e3967. [Google Scholar] [CrossRef]
- Wong, M.T.; Ong, D.E.; Lim, F.S.; Teng, K.W.; McGovern, N.; Narayanan, S.; Ho, W.Q.; Cerny, D.; Tan, H.K.; Anicete, R.; et al. A High-Dimensional Atlas of Human T Cell Diversity Reveals Tissue-Specific Trafficking and Cytokine Signatures. Immunity 2016, 45, 442–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Ji, C.; Shen, Q.; Liu, W.; Qin, F.X.; Wu, A. Tissue-specific deconvolution of immune cell composition by integrating bulk and single-cell transcriptomes. Bioinformatics 2020, 36, 819–827. [Google Scholar] [CrossRef]
- DeGottardi, Q.; Gates, T.J.; Yang, J.; James, E.A.; Malhotra, U.; Chow, I.T.; Simoni, Y.; Fehlings, M.; Newell, E.W.; DeBerg, H.A.; et al. Ontogeny of different subsets of yellow fever virus-specific circulatory CXCR5+ CD4+ T cells after yellow fever vaccination. Sci. Rep. 2020, 10, 15686. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, A.; Zhang, J.H.; Oxman, M.N.; Johnson, G.R.; Hayward, A.R.; Caulfield, M.J.; Irwin, M.R.; Clair, J.; Smith, J.G.; Stanley, H.; et al. Varicella-zoster virus-specific immune responses to herpes zoster in elderly participants in a trial of a clinically effective zoster vaccine. J. Infect. Dis. 2009, 200, 1068–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchanathan, V.; Chaudhri, G.; Karupiah, G. Protective immunity against secondary poxvirus infection is dependent on antibody but not on CD4 or CD8 T-cell function. J. Virol. 2006, 80, 6333–6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, B. Smallpox vaccines: Targets of protective immunity. Immunol. Rev. 2011, 239, 8–26. [Google Scholar] [CrossRef]
- Querec, T.D.; Akondy, R.S.; Lee, E.K.; Cao, W.; Nakaya, H.I.; Teuwen, D.; Pirani, A.; Gernert, K.; Deng, J.; Marzolf, B.; et al. Systems biology approach predicts immunogenicity of the yellow fever vaccine in humans. Nat. Immunol. 2009, 10, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Gonnet, J.; Poncelet, L.; Meriaux, C.; Gonçalves, E.; Weiss, L.; Tchitchek, N.; Pedruzzi, E.; Soria, A.; Boccara, D.; Vogt, A.; et al. Mechanisms of innate events during skin reaction following intradermal injection of seasonal influenza vaccine. J. Proteom. 2020, 216, 103670. [Google Scholar] [CrossRef]
- Gourley, T.S.; Wherry, E.J.; Masopust, D.; Ahmed, R. Generation and maintenance of immunological memory. Semin Immunol. 2004, 16, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, K.; Tokoyoda, K.; Hanazawa, A.; Hayashizaki, K.; Zehentmeier, S.; Hosokawa, H.; Iwamura, C.; Koseki, H.; Tumes, D.J.; Radbruch, A.; et al. Type II membrane protein CD69 regulates the formation of resting T-helper memory. Proc. Natl. Acad. Sci. USA 2012, 109, 7409–7414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monath, T.P. Yellow fever: An update. Lancet Infect. Dis. 2001, 1, 11–20. [Google Scholar] [CrossRef]
- Darrah, P.A.; Zeppa, J.J.; Maiello, P.; Hackney, J.A.; Wadsworth, M.H., 2nd; Hughes, T.K.; Pokkali, S.; Swanson, P.A., 2nd; Grant, N.L.; Rodgers, M.A.; et al. Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature 2020, 577, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Pattyn, J.; Van Keer, S.; Tjalma, W.; Matheeussen, V.; Van Damme, P.; Vorsters, A. Infection and vaccine-induced HPV-specific antibodies in cervicovaginal secretions. A review of the literature. Papillomavirus Res. 2019, 8, 100185. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, S.K.; Kulkarni, R.R.; Weldon, W.C.; Silveira, E.L.V.; Ahmed, H.; Gunisetty, S.; Chandele, A.; Antia, R.; Verma, H.; Sutter, R.; et al. Immune Priming and Long-term Persistence of Memory B Cells After Inactivated Poliovirus Vaccine in Macaque Models: Support for at least 2 Doses. Clin. Infect. Dis. 2018, 67, S66–S77. [Google Scholar] [CrossRef]
- Tsang, J.S.; Schwartzberg, P.L.; Kotliarov, Y.; Biancotto, A.; Xie, Z.; Germain, R.N.; Wang, E.; Olnes, M.J.; Narayanan, M.; Golding, H.; et al. Global analyses of human immune variation reveal baseline predictors of postvaccination responses. Cell 2014, 157, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Human Immunology Project Consortium—Center for Human Immunology (HIPC-CHI) Signatures Project Team and HIPC-I Consortium. Multicohort analysis reveals baseline transcriptional predictors of influenza vaccination responses. Sci. Immunol. 2017, 2, eaal4656. [Google Scholar] [CrossRef] [Green Version]
- Kotliarov, Y.; Sparks, R.; Martins, A.J.; Mulè, M.P.; Lu, Y.; Goswami, M.; Kardava, L.; Banchereau, R.; Pascual, V.; Biancotto, A.; et al. Broad immune activation underlies shared set point signatures for vaccine responsiveness in healthy individuals and disease activity in patients with lupus. Nat. Med. 2020, 26, 618–629. [Google Scholar] [CrossRef]
- Fourati, S.; Cristescu, R.; Loboda, A.; Talla, A.; Filali, A.; Railkar, R.; Schaeffer, A.K.; Favre, D.; Gagnon, D.; Peretz, Y.; et al. Pre-vaccination inflammation and B-cell signalling predict age-related hyporesponse to hepatitis B vaccination. Nat. Commun. 2016, 7, 10369. [Google Scholar] [CrossRef]
- Pauthner, M.; Havenar-Daughton, C.; Sok, D.; Nkolola, J.P.; Bastidas, R.; Boopathy, A.V.; Carnathan, D.G.; Chandrashekar, A.; Cirelli, K.M.; Cottrell, C.A.; et al. Elicitation of Robust Tier 2 Neutralizing Antibody Responses in Nonhuman Primates by HIV Envelope Trimer Immunization Using Optimized Approaches. Immunity 2017, 46, 1073–1088.e1076. [Google Scholar] [CrossRef] [Green Version]
- Mahnke, Y.D.; Roederer, M. Optimizing a multicolor immunophenotyping assay. Clin. Lab. Med. 2007, 27, 469–485. [Google Scholar] [CrossRef] [Green Version]
- Reeves, P.M.; Sluder, A.E.; Paul, S.R.; Scholzen, A.; Kashiwagi, S.; Poznansky, M.C. Application and utility of mass cytometry in vaccine development. FASEB J. 2018, 32, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Futamura, K.; Sekino, M.; Hata, A.; Ikebuchi, R.; Nakanishi, Y.; Egawa, G.; Kabashima, K.; Watanabe, T.; Furuki, M.; Tomura, M. Novel full-spectral flow cytometry with multiple spectrally-adjacent fluorescent proteins and fluorochromes and visualization of in vivo cellular movement. Cytom. A 2015, 87, 830–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzer, M.H.; Nolan, G.P. Mass Cytometry: Single Cells, Many Features. Cell 2016, 165, 780–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, L.R.; Leipold, M.D.; Pedersen, C.B.; Maecker, H.T. The anatomy of single cell mass cytometry data. Cytom. A 2019, 95, 156–172. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, S.C.; Roederer, M. Eleven-color flow cytometry. A powerful tool for elucidation of the complex immune system. Clin. Lab. Med. 2001, 21, 697–712. [Google Scholar] [PubMed]
- Boesch, M.; Cosma, A.; Sopper, S. Flow Cytometry: To Dump or Not To Dump. J. Immunol. 2018, 201, 1813–1815. [Google Scholar] [CrossRef] [Green Version]
- Pejoski, D.; Tchitchek, N.; Rodriguez Pozo, A.; Elhmouzi-Younes, J.; Yousfi-Bogniaho, R.; Rogez-Kreuz, C.; Clayette, P.; Dereuddre-Bosquet, N.; Lévy, Y.; Cosma, A.; et al. Identification of Vaccine-Altered Circulating B Cell Phenotypes Using Mass Cytometry and a Two-Step Clustering Analysis. J. Immunol. 2016, 196, 4814–4831. [Google Scholar] [CrossRef] [Green Version]
- Subrahmanyam, P.B.; Holmes, T.H.; Lin, D.; Su, L.F.; Obermoser, G.; Banchereau, J.; Pascual, V.; García-Sastre, A.; Albrecht, R.A.; Palucka, K.; et al. Mass Cytometry Defines Virus-Specific CD4+ T Cells in Influenza Vaccination. Immunohorizons 2020, 4, 774–788. [Google Scholar] [CrossRef]
- Lingblom, C.M.D.; Kowli, S.; Swaminathan, N.; Maecker, H.T.; Lambert, S.L. Baseline immune profile by CyTOF can predict response to an investigational adjuvanted vaccine in elderly adults. J. Transl. Med. 2018, 16, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, D.; Davis, M.M. New approaches to understanding the immune response to vaccination and infection. Vaccine 2015, 33, 5271–5281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbarasu, D.; Raja, C.P.; Raja, A. Multiplex analysis of cytokines/chemokines as biomarkers that differentiate healthy contacts from tuberculosis patients in high endemic settings. Cytokine 2013, 61, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Ashton, N.J.; Dobson, R.J.B.; Andersson, L.M.; Yilmaz, A.; Blennow, K.; Gisslen, M.; Zetterberg, H. Proteomic blood profiling in mild, severe and critical COVID-19 patients. Sci. Rep. 2021, 11, 6357. [Google Scholar] [CrossRef] [PubMed]
- Chauvat, A.; Benhamouda, N.; Gey, A.; Lemoine, F.M.; Paulie, S.; Carrat, F.; Gougeon, M.L.; Rozenberg, F.; Krivine, A.; Cherai, M.; et al. Clinical validation of IFNγ/IL-10 and IFNγ/IL-2 FluoroSpot assays for the detection of Tr1 T cells and influenza vaccine monitoring in humans. Hum. Vaccines Immunother. 2014, 10, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttner, A.; Combescure, C.; Grillet, S.; Haks, M.C.; Quinten, E.; Modoux, C.; Agnandji, S.T.; Brosnahan, J.; Dayer, J.A.; Harandi, A.M.; et al. A dose-dependent plasma signature of the safety and immunogenicity of the rVSV-Ebola vaccine in Europe and Africa. Sci. Transl. Med. 2017, 9, eaaj1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galson, J.D.; Pollard, A.J.; Trück, J.; Kelly, D.F. Studying the antibody repertoire after vaccination: Practical applications. Trends Immunol. 2014, 35, 319–331. [Google Scholar] [CrossRef]
- Teraguchi, S.; Saputri, D.S.; Llamas-Covarrubias, M.A.; Davila, A.; Diez, D.; Nazlica, S.A.; Rozewicki, J.; Ismanto, H.S.; Wilamowski, J.; Xie, J.; et al. Methods for sequence and structural analysis of B and T cell receptor repertoires. Comput. Struct. Biotechnol. J. 2020, 18, 2000–2011. [Google Scholar] [CrossRef]
- Cotugno, N.; Ruggiero, A.; Santilli, V.; Manno, E.C.; Rocca, S.; Zicari, S.; Amodio, D.; Colucci, M.; Rossi, P.; Levy, O.; et al. OMIC Technologies and Vaccine Development: From the Identification of Vulnerable Individuals to the Formulation of Invulnerable Vaccines. J. Immunol. Res. 2019, 2019, 8732191. [Google Scholar] [CrossRef] [Green Version]
- Arunachalam, P.S.; Charles, T.P.; Joag, V.; Bollimpelli, V.S.; Scott, M.K.D.; Wimmers, F.; Burton, S.L.; Labranche, C.C.; Petitdemange, C.; Gangadhara, S.; et al. T cell-inducing vaccine durably prevents mucosal SHIV infection even with lower neutralizing antibody titers. Nat. Med. 2020, 26, 932–940. [Google Scholar] [CrossRef]
- Routhu, N.K.; Cheedarla, N.; Gangadhara, S.; Bollimpelli, V.S.; Boddapati, A.K.; Shiferaw, A.; Rahman, S.A.; Sahoo, A.; Edara, V.V.; Lai, L.; et al. A modified vaccinia Ankara vector-based vaccine protects macaques from SARS-CoV-2 infection, immune pathology, and dysfunction in the lungs. Immunity 2021, 54, 542–556.e549. [Google Scholar] [CrossRef]
- Afik, S.; Yates, K.B.; Bi, K.; Darko, S.; Godec, J.; Gerdemann, U.; Swadling, L.; Douek, D.C.; Klenerman, P.; Barnes, E.J.; et al. Targeted reconstruction of T cell receptor sequence from single cell RNA-seq links CDR3 length to T cell differentiation state. Nucleic Acids Res. 2017, 45, e148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.S.; Zhou, J.Q.; Han, J.; Schmitz, A.J.; Rizk, A.A.; Alsoussi, W.B.; Lei, T.; Amor, M.; McIntire, K.M.; Meade, P.; et al. Human germinal centres engage memory and naive B cells after influenza vaccination. Nature 2020, 586, 127–132. [Google Scholar] [CrossRef]
- Horns, F.; Dekker, C.L.; Quake, S.R. Memory B Cell Activation, Broad Anti-influenza Antibodies, and Bystander Activation Revealed by Single-Cell Transcriptomics. Cell Rep. 2020, 30, 905–913.e906. [Google Scholar] [CrossRef] [Green Version]
- Cirelli, K.M.; Carnathan, D.G.; Nogal, B.; Martin, J.T.; Rodriguez, O.L.; Upadhyay, A.A.; Enemuo, C.A.; Gebru, E.H.; Choe, Y.; Viviano, F.; et al. Slow Delivery Immunization Enhances HIV Neutralizing Antibody and Germinal Center Responses via Modulation of Immunodominance. Cell 2020, 180, 206. [Google Scholar] [CrossRef] [PubMed]
- Waickman, A.T.; Victor, K.; Li, T.; Hatch, K.; Rutvisuttinunt, W.; Medin, C.; Gabriel, B.; Jarman, R.G.; Friberg, H.; Currier, J.R. Dissecting the heterogeneity of DENV vaccine-elicited cellular immunity using single-cell RNA sequencing and metabolic profiling. Nat. Commun. 2019, 10, 3666. [Google Scholar] [CrossRef] [Green Version]
- James, M.L.; Gambhir, S.S. A molecular imaging primer: Modalities, imaging agents, and applications. Physiol. Rev. 2012, 92, 897–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrielson, K.; Maronpot, R.; Monette, S.; Mlynarczyk, C.; Ramot, Y.; Nyska, A.; Sysa-Shah, P. In Vivo Imaging With Confirmation by Histopathology for Increased Rigor and Reproducibility in Translational Research: A Review of Examples, Options, and Resources. Ilar J. 2018, 59, 80–98. [Google Scholar] [CrossRef]
- Ottobrini, L.; Martelli, C.; Trabattoni, D.L.; Clerici, M.; Lucignani, G. In vivo imaging of immune cell trafficking in cancer. Eur. J. Nucl. Med. Mol. Imaging 2011, 38, 949–968. [Google Scholar] [CrossRef]
- Romain, G.; van Gulck, E.; Epaulard, O.; Oh, S.; Li, D.; Zurawski, G.; Zurawski, S.; Cosma, A.; Adam, L.; Chapon, C.; et al. CD34-derived dendritic cells transfected ex vivo with HIV-Gag mRNA induce polyfunctional T-cell responses in nonhuman primates. Eur. J. Immunol. 2012, 42, 2019–2030. [Google Scholar] [CrossRef] [Green Version]
- Ahrens, E.T.; Bulte, J.W. Tracking immune cells in vivo using magnetic resonance imaging. Nat. Rev. Immunol. 2013, 13, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Brewer, K.D.; Lake, K.; Pelot, N.; Stanford, M.M.; DeBay, D.R.; Penwell, A.; Weir, G.M.; Karkada, M.; Mansour, M.; Bowen, C.V. Clearance of depot vaccine SPIO-labeled antigen and substrate visualized using MRI. Vaccine 2014, 32, 6956–6962. [Google Scholar] [CrossRef]
- Ferguson, P.M.; Slocombe, A.; Tilley, R.D.; Hermans, I.F. Using magnetic resonance imaging to evaluate dendritic cell-based vaccination. PLoS ONE 2013, 8, e65318. [Google Scholar] [CrossRef] [PubMed]
- Daldrup-Link, H.E.; Golovko, D.; Ruffell, B.; Denardo, D.G.; Castaneda, R.; Ansari, C.; Rao, J.; Tikhomirov, G.A.; Wendland, M.F.; Corot, C.; et al. MRI of tumor-associated macrophages with clinically applicable iron oxide nanoparticles. Clin. Cancer Res. 2011, 17, 5695–5704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makela, A.V.; Gaudet, J.M.; Foster, P.J. Quantifying tumor associated macrophages in breast cancer: A comparison of iron and fluorine-based MRI cell tracking. Sci. Rep. 2017, 7, 42109. [Google Scholar] [CrossRef] [PubMed]
- Floris, S.; Blezer, E.L.; Schreibelt, G.; Döpp, E.; van der Pol, S.M.; Schadee-Eestermans, I.L.; Nicolay, K.; Dijkstra, C.D.; de Vries, H.E. Blood-brain barrier permeability and monocyte infiltration in experimental allergic encephalomyelitis: A quantitative MRI study. Brain 2004, 127, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wells, J.; Buist, R.; Peeling, J.; Yong, V.W.; Mitchell, J.R. Active inflammation increases the heterogeneity of MRI texture in mice with relapsing experimental allergic encephalomyelitis. Magn. Reson. Imaging 2014, 32, 168–174. [Google Scholar] [CrossRef]
- Tremblay, M.L.; Davis, C.; Bowen, C.V.; Stanley, O.; Parsons, C.; Weir, G.; Karkada, M.; Stanford, M.M.; Brewer, K.D. Using MRI cell tracking to monitor immune cell recruitment in response to a peptide-based cancer vaccine. Magn. Reson. Med. 2018, 80, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Meller, B.; Frohn, C.; Brand, J.M.; Lauer, I.; Schelper, L.F.; von Hof, K.; Kirchner, H.; Richter, E.; Baehre, M. Monitoring of a new approach of immunotherapy with allogenic (111)In-labelled NK cells in patients with renal cell carcinoma. Eur. J. Nucl. Med. Mol. Imaging 2004, 31, 403–407. [Google Scholar] [CrossRef]
- Yuki, Y.; Nochi, T.; Harada, N.; Katakai, Y.; Shibata, H.; Mejima, M.; Kohda, T.; Tokuhara, D.; Kurokawa, S.; Takahashi, Y.; et al. In vivo molecular imaging analysis of a nasal vaccine that induces protective immunity against botulism in nonhuman primates. J Immunol. 2010, 185, 5436–5443. [Google Scholar] [CrossRef]
- Pektor, S.; Hilscher, L.; Walzer, K.C.; Miederer, I.; Bausbacher, N.; Loquai, C.; Schreckenberger, M.; Sahin, U.; Diken, M.; Miederer, M. In vivo imaging of the immune response upon systemic RNA cancer vaccination by FDG-PET. EJNMMI Res. 2018, 8, 80. [Google Scholar] [CrossRef]
- Coates, E.E.; Costner, P.J.; Nason, M.C.; Herrin, D.M.; Conant, S.; Herscovitch, P.; Sarwar, U.N.; Holman, L.; Mitchell, J.; Yamshchikov, G.; et al. Lymph Node Activation by PET/CT Following Vaccination With Licensed Vaccines for Human Papillomaviruses. Clin. Nucl. Med. 2017, 42, 329–334. [Google Scholar] [CrossRef]
- Vadalà, M.; Cistaro, A.; Quartuccio, N.; Calcagni, M.L.; Fania, P.; Margotti, S.; Schiera, I.G.; Laurino, C.; Palmieri, B. 18F-FDG-PET brain imaging may highlight brain metabolic alterations in dysautonomic syndrome after human papilloma virus vaccination. Nucl Med. Commun. 2020, 41, 1275–1282. [Google Scholar] [CrossRef]
- Mingos, M.; Howard, S.; Giacalone, N.; Kozono, D.; Jacene, H. Systemic Immune Response to Vaccination on FDG-PET/CT. Nucl. Med. Mol. Imaging 2016, 50, 358–361. [Google Scholar] [CrossRef] [Green Version]
- Shirone, N.; Shinkai, T.; Yamane, T.; Uto, F.; Yoshimura, H.; Tamai, H.; Imai, T.; Inoue, M.; Kitano, S.; Kichikawa, K.; et al. Axillary lymph node accumulation on FDG-PET/CT after influenza vaccination. Ann. Nucl. Med. 2012, 26, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Aarntzen, E.H.; Srinivas, M.; De Wilt, J.H.; Jacobs, J.F.; Lesterhuis, W.J.; Windhorst, A.D.; Troost, E.G.; Bonenkamp, J.J.; van Rossum, M.M.; Blokx, W.A.; et al. Early identification of antigen-specific immune responses in vivo by [18F]-labeled 3’-fluoro-3’-deoxy-thymidine ([18F]FLT) PET imaging. Proc. Natl. Acad. Sci. USA 2011, 108, 18396–18399. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Li, F.; Niu, G.; Chen, X. PET imaging of inflammation biomarkers. Theranostics 2013, 3, 448–466. [Google Scholar] [CrossRef] [Green Version]
- Rashidian, M.; LaFleur, M.W.; Verschoor, V.L.; Dongre, A.; Zhang, Y.; Nguyen, T.H.; Kolifrath, S.; Aref, A.R.; Lau, C.J.; Paweletz, C.P.; et al. Immuno-PET identifies the myeloid compartment as a key contributor to the outcome of the antitumor response under PD-1 blockade. Proc. Natl. Acad. Sci. USA 2019, 116, 16971–16980. [Google Scholar] [CrossRef] [Green Version]
- Freise, A.C.; Wu, A.M. In vivo imaging with antibodies and engineered fragments. Mol. Immunol. 2015, 67, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.M.; Saverymuttu, S.H.; Reavy, H.J.; Danpure, H.J.; Osman, S.; Lavender, J.P. Imaging of inflammation with indium-111 tropolonate labeled leukocytes. J. Nucl. Med. 1983, 24, 39–44. [Google Scholar]
- Watson, H.A.; Durairaj, R.R.P.; Ohme, J.; Alatsatianos, M.; Almutairi, H.; Mohammed, R.N.; Vigar, M.; Reed, S.G.; Paisey, S.J.; Marshall, C.; et al. L-Selectin Enhanced T Cells Improve the Efficacy of Cancer Immunotherapy. Front. Immunol. 2019, 10, 1321. [Google Scholar] [CrossRef] [PubMed]
- Heskamp, S.; Raavé, R.; Boerman, O.; Rijpkema, M.; Goncalves, V.; Denat, F. (89)Zr-Immuno-Positron Emission Tomography in Oncology: State-of-the-Art (89)Zr Radiochemistry. Bioconjug. Chem. 2017, 28, 2211–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.; Carroll, L.; Yahioglu, G.; Aboagye, E.O.; Miller, P.W. Antibody Fragment and Affibody ImmunoPET Imaging Agents: Radiolabelling Strategies and Applications. Chem. Med. Chem. 2018, 13, 2466–2478. [Google Scholar] [CrossRef]
- Cruz, L.J.; Tacken, P.J.; Bonetto, F.; Buschow, S.I.; Croes, H.J.; Wijers, M.; de Vries, I.J.; Figdor, C.G. Multimodal imaging of nanovaccine carriers targeted to human dendritic cells. Mol. Pharm. 2011, 8, 520–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorova, B.; Adam, L.; Culina, S.; Boisgard, R.; Martinon, F.; Cosma, A.; Ustav, M.; Kortulewski, T.; Le Grand, R.; Chapon, C. Electroporation as a vaccine delivery system and a natural adjuvant to intradermal administration of plasmid DNA in macaques. Sci. Rep. 2017, 7, 4122. [Google Scholar] [CrossRef]
- Cao, Q.; Huang, Q.; Mohan, C.; Li, C. Small-Animal PET/CT Imaging of Local and Systemic Immune Response Using (64)Cu-αCD11b. J. Nucl. Med. 2019, 60, 1317–1324. [Google Scholar] [CrossRef] [Green Version]
- Nigam, S.; McCarl, L.; Kumar, R.; Edinger, R.S.; Kurland, B.F.; Anderson, C.J.; Panigrahy, A.; Kohanbash, G.; Edwards, W.B. Preclinical ImmunoPET Imaging of Glioblastoma-Infiltrating Myeloid Cells Using Zirconium-89 Labeled Anti-CD11b Antibody. Mol. Imaging Biol. 2020, 22, 685–694. [Google Scholar] [CrossRef]
- Warram, J.M.; de Boer, E.; Sorace, A.G.; Chung, T.K.; Kim, H.; Pleijhuis, R.G.; van Dam, G.M.; Rosenthal, E.L. Antibody-based imaging strategies for cancer. Cancer Metastasis Rev. 2014, 33, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Chanier, T.; Chames, P. Nanobody Engineering: Toward Next Generation Immunotherapies and Immunoimaging of Cancer. Antibodies 2019, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blykers, A.; Schoonooghe, S.; Xavier, C.; D’Hoe, K.; Laoui, D.; D’Huyvetter, M.; Vaneycken, I.; Cleeren, F.; Bormans, G.; Heemskerk, J.; et al. PET Imaging of Macrophage Mannose Receptor-Expressing Macrophages in Tumor Stroma Using 18F-Radiolabeled Camelid Single-Domain Antibody Fragments. J. Nucl. Med. 2015, 56, 1265–1271. [Google Scholar] [CrossRef] [Green Version]
- Van Elssen, C.; Rashidian, M.; Vrbanac, V.; Wucherpfennig, K.W.; Habre, Z.E.; Sticht, J.; Freund, C.; Jacobsen, J.T.; Cragnolini, J.; Ingram, J.; et al. Noninvasive Imaging of Human Immune Responses in a Human Xenograft Model of Graft-Versus-Host Disease. J. Nucl. Med. 2017, 58, 1003–1008. [Google Scholar] [CrossRef] [Green Version]
- Chandrupatla, D.; Molthoff, C.F.M.; Lammertsma, A.A.; van der Laken, C.J.; Jansen, G. The folate receptor β as a macrophage-mediated imaging and therapeutic target in rheumatoid arthritis. Drug Deliv. Transl. Res. 2019, 9, 366–378. [Google Scholar] [CrossRef] [Green Version]
- Varasteh, Z.; Mohanta, S.; Li, Y.; López Armbruster, N.; Braeuer, M.; Nekolla, S.G.; Habenicht, A.; Sager, H.B.; Raes, G.; Weber, W.; et al. Targeting mannose receptor expression on macrophages in atherosclerotic plaques of apolipoprotein E-knockout mice using (68)Ga-NOTA-anti-MMR nanobody: Non-invasive imaging of atherosclerotic plaques. EJNMMI Res. 2019, 9, 5. [Google Scholar] [CrossRef]
- Movahedi, K.; Schoonooghe, S.; Laoui, D.; Houbracken, I.; Waelput, W.; Breckpot, K.; Bouwens, L.; Lahoutte, T.; De Baetselier, P.; Raes, G.; et al. Nanobody-based targeting of the macrophage mannose receptor for effective in vivo imaging of tumor-associated macrophages. Cancer Res. 2012, 72, 4165–4177. [Google Scholar] [CrossRef] [Green Version]
- Mall, S.; Yusufi, N.; Wagner, R.; Klar, R.; Bianchi, H.; Steiger, K.; Straub, M.; Audehm, S.; Laitinen, I.; Aichler, M.; et al. Immuno-PET Imaging of Engineered Human T Cells in Tumors. Cancer Res. 2016, 76, 4113–4123. [Google Scholar] [CrossRef] [Green Version]
- Tavaré, R.; McCracken, M.N.; Zettlitz, K.A.; Knowles, S.M.; Salazar, F.B.; Olafsen, T.; Witte, O.N.; Wu, A.M. Engineered antibody fragments for immuno-PET imaging of endogenous CD8+ T cells in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 1108–1113. [Google Scholar] [CrossRef] [Green Version]
- Tavaré, R.; Escuin-Ordinas, H.; Mok, S.; McCracken, M.N.; Zettlitz, K.A.; Salazar, F.B.; Witte, O.N.; Ribas, A.; Wu, A.M. An Effective Immuno-PET Imaging Method to Monitor CD8-Dependent Responses to Immunotherapy. Cancer Res. 2016, 76, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Rashidian, M.; Ingram, J.R.; Dougan, M.; Dongre, A.; Whang, K.A.; LeGall, C.; Cragnolini, J.J.; Bierie, B.; Gostissa, M.; Gorman, J.; et al. Predicting the response to CTLA-4 blockade by longitudinal noninvasive monitoring of CD8 T cells. J. Exp. Med. 2017, 214, 2243–2255. [Google Scholar] [CrossRef]
- Woodham, A.W.; Zeigler, S.H.; Zeyang, E.L.; Kolifrath, S.C.; Cheloha, R.W.; Rashidian, M.; Chaparro, R.J.; Seidel, R.D.; Garforth, S.J.; Dearling, J.L.; et al. In vivo detection of antigen-specific CD8+ T cells by immuno-positron emission tomography. Nat. Methods 2020, 17, 1025–1032. [Google Scholar] [CrossRef]
- Faye, N.; Fournier, L.; Balvay, D.; Wilhelm, C.; Broqueres-You, D.; Bruneval, P.; Clément, O. Antitumoral Effect of Mural Cells Assessed With High-Resolution MRI and Fluorescence Microscopy. Ajr. Am. J. Roentgenol. 2015, 205, W11–W18. [Google Scholar] [CrossRef]
- Al-Gubory, K.H. Shedding light on fibered confocal fluorescence microscopy: Applications in biomedical imaging and therapies. J. Biophotonics 2019, 12, e201900146. [Google Scholar] [CrossRef]
- Mahe, B.; Vogt, A.; Liard, C.; Duffy, D.; Abadie, V.; Bonduelle, O.; Boissonnas, A.; Sterry, W.; Verrier, B.; Blume-Peytavi, U.; et al. Nanoparticle-based targeting of vaccine compounds to skin antigen-presenting cells by hair follicles and their transport in mice. J. Investig. Derm. 2009, 129, 1156–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmchen, F.; Denk, W. Deep tissue two-photon microscopy. Nat. Methods 2005, 2, 932–940. [Google Scholar] [CrossRef]
- Rattanapak, T.; Birchall, J.C.; Young, K.; Kubo, A.; Fujimori, S.; Ishii, M.; Hook, S. Dynamic visualization of dendritic cell-antigen interactions in the skin following transcutaneous immunization. PLoS ONE 2014, 9, e89503. [Google Scholar] [CrossRef]
- Miller, M.J.; Wei, S.H.; Parker, I.; Cahalan, M.D. Two-photon imaging of lymphocyte motility and antigen response in intact lymph node. Science 2002, 296, 1869–1873. [Google Scholar] [CrossRef] [Green Version]
- Bousso, P.; Robey, E. Dynamics of CD8+ T cell priming by dendritic cells in intact lymph nodes. Nat. Immunol. 2003, 4, 579–585. [Google Scholar] [CrossRef]
- Deguine, J.; Bousso, P. Dynamics of NK cell interactions in vivo. Immunol. Rev. 2013, 251, 154–159. [Google Scholar] [CrossRef]
- Bousso, P.; Bhakta, N.R.; Lewis, R.S.; Robey, E. Dynamics of thymocyte-stromal cell interactions visualized by two-photon microscopy. Science 2002, 296, 1876–1880. [Google Scholar] [CrossRef]
- Tan, W.C.C.; Nerurkar, S.N.; Cai, H.Y.; Ng, H.H.M.; Wu, D.; Wee, Y.T.F.; Lim, J.C.T.; Yeong, J.; Lim, T.K.H. Overview of multiplex immunohistochemistry/immunofluorescence techniques in the era of cancer immunotherapy. Cancer Commun. 2020, 40, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Muntifering, M.; Castranova, D.; Gibson, G.A.; Meyer, E.; Kofron, M.; Watson, A.M. Clearing for Deep Tissue Imaging. Curr. Protoc. Cytom. 2018, 86, e38. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Alieva, M.; Wellens, L.M.; Ariese, H.C.R.; Jamieson, P.R.; Vonk, A.M.; Amatngalim, G.D.; Hu, H.; Oost, K.C.; Snippert, H.J.G.; et al. High-resolution 3D imaging of fixed and cleared organoids. Nat. Protoc. 2019, 14, 1756–1771. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Germain, R.N.; Gerner, M.Y. Multiplex, quantitative cellular analysis in large tissue volumes with clearing-enhanced 3D microscopy (Ce3D). Proc. Natl. Acad. Sci. USA 2017, 114, E7321–E7330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walch, A.; Rauser, S.; Deininger, S.O.; Höfler, H. MALDI imaging mass spectrometry for direct tissue analysis: A new frontier for molecular histology. Histochem. Cell Biol. 2008, 130, 421–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caprioli, R.M.; Farmer, T.B.; Gile, J. Molecular imaging of biological samples: Localization of peptides and proteins using MALDI-TOF MS. Anal. Chem. 1997, 69, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Ryan, N.M.; Hess, J.A.; de Villena, F.P.; Leiby, B.E.; Shimada, A.; Yu, L.; Yarmahmoodi, A.; Petrovsky, N.; Zhan, B.; Bottazzi, M.E.; et al. Onchocerca volvulus bivalent subunit vaccine induces protective immunity in genetically diverse collaborative cross recombinant inbred intercross mice. NPJ Vaccines 2021, 6, 17. [Google Scholar] [CrossRef]
- Yagnik, G.; Liu, Z.; Rothschild, K.J.; Lim, M.J. Highly Multiplexed Immunohistochemical MALDI-MS Imaging of Biomarkers in Tissues. J. Am. Soc. Mass Spectrom. 2021, 32, 977–988. [Google Scholar] [CrossRef]
- Oetjen, J.; Aichler, M.; Trede, D.; Strehlow, J.; Berger, J.; Heldmann, S.; Becker, M.; Gottschalk, M.; Kobarg, J.H.; Wirtz, S.; et al. MRI-compatible pipeline for three-dimensional MALDI imaging mass spectrometry using PAXgene fixation. J. Proteom. 2013, 90, 52–60. [Google Scholar] [CrossRef]
- Kriegsmann, K.; Longuespée, R.; Hundemer, M.; Zgorzelski, C.; Casadonte, R.; Schwamborn, K.; Weichert, W.; Schirmacher, P.; Harms, A.; Kazdal, D.; et al. Combined Immunohistochemistry after Mass Spectrometry Imaging for Superior Spatial Information. Proteom. Clin. Appl. 2019, 13, e1800035. [Google Scholar] [CrossRef] [Green Version]
- Porta Siegel, T.; Hamm, G.; Bunch, J.; Cappell, J.; Fletcher, J.S.; Schwamborn, K. Mass Spectrometry Imaging and Integration with Other Imaging Modalities for Greater Molecular Understanding of Biological Tissues. Mol. Imaging Biol. 2018, 20, 888–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giesen, C.; Wang, H.A.; Schapiro, D.; Zivanovic, N.; Jacobs, A.; Hattendorf, B.; Schüffler, P.J.; Grolimund, D.; Buhmann, J.M.; Brandt, S.; et al. Highly multiplexed imaging of tumor tissues with subcellular resolution by mass cytometry. Nat. Methods 2014, 11, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Bandura, D.R.; Baranov, V.I.; Ornatsky, O.I.; Antonov, A.; Kinach, R.; Lou, X.; Pavlov, S.; Vorobiev, S.; Dick, J.E.; Tanner, S.D. Mass cytometry: Technique for real time single cell multitarget immunoassay based on inductively coupled plasma time-of-flight mass spectrometry. Anal. Chem. 2009, 81, 6813–6822. [Google Scholar] [CrossRef] [PubMed]
- Baharlou, H.; Canete, N.P.; Cunningham, A.L.; Harman, A.N.; Patrick, E. Mass Cytometry Imaging for the Study of Human Diseases-Applications and Data Analysis Strategies. Front. Immunol. 2019, 10, 2657. [Google Scholar] [CrossRef]
- Zhao, Y.; Uduman, M.; Siu, J.H.Y.; Tull, T.J.; Sanderson, J.D.; Wu, Y.B.; Zhou, J.Q.; Petrov, N.; Ellis, R.; Todd, K.; et al. Spatiotemporal segregation of human marginal zone and memory B cell populations in lymphoid tissue. Nat. Commun. 2018, 9, 3857. [Google Scholar] [CrossRef]
- Damond, N.; Engler, S.; Zanotelli, V.R.T.; Schapiro, D.; Wasserfall, C.H.; Kusmartseva, I.; Nick, H.S.; Thorel, F.; Herrera, P.L.; Atkinson, M.A.; et al. A Map of Human Type 1 Diabetes Progression by Imaging Mass Cytometry. Cell Metab. 2019, 29, 755–768.e755. [Google Scholar] [CrossRef] [Green Version]
- Buchberger, A.R.; DeLaney, K.; Johnson, J.; Li, L. Mass Spectrometry Imaging: A Review of Emerging Advancements and Future Insights. Anal. Chem. 2018, 90, 240–265. [Google Scholar] [CrossRef]
- Durand, M.; Walter, T.; Pirnay, T.; Naessens, T.; Gueguen, P.; Goudot, C.; Lameiras, S.; Chang, Q.; Talaei, N.; Ornatsky, O.; et al. Human lymphoid organ cDC2 and macrophages play complementary roles in T follicular helper responses. J. Exp. Med. 2019, 216, 1561–1581. [Google Scholar] [CrossRef] [Green Version]
- Gerner, M.Y.; Kastenmuller, W.; Ifrim, I.; Kabat, J.; Germain, R.N. Histo-cytometry: A method for highly multiplex quantitative tissue imaging analysis applied to dendritic cell subset microanatomy in lymph nodes. Immunity 2012, 37, 364–376. [Google Scholar] [CrossRef] [Green Version]
- Gerner, M.Y.; Torabi-Parizi, P.; Germain, R.N. Strategically localized dendritic cells promote rapid T cell responses to lymph-borne particulate antigens. Immunity 2015, 42, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Petrovas, C.; Ferrando-Martinez, S.; Gerner, M.Y.; Casazza, J.P.; Pegu, A.; Deleage, C.; Cooper, A.; Hataye, J.; Andrews, S.; Ambrozak, D.; et al. Follicular CD8 T cells accumulate in HIV infection and can kill infected cells in vitro via bispecific antibodies. Sci. Transl. Med. 2017, 9, eaag2285. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Germain, R.N.; Gerner, M.Y. High-dimensional cell-level analysis of tissues with Ce3D multiplex volume imaging. Nat. Protoc. 2019, 14, 1708–1733. [Google Scholar] [CrossRef]
- Kotov, D.I.; Pengo, T.; Mitchell, J.S.; Gastinger, M.J.; Jenkins, M.K. Chrysalis: A New Method for High-Throughput Histo-Cytometry Analysis of Images and Movies. J. Immunol. 2019, 202, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.W.; da Silva Santos, L.B.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Silva, J.; Domingues, D.; Lopes, F.M. RNA-Seq differential expression analysis: An extended review and a software tool. PLoS ONE 2017, 12, e0190152. [Google Scholar] [CrossRef] [Green Version]
- Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [CrossRef] [PubMed]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Qiu, P.; Simonds, E.F.; Bendall, S.C.; Gibbs, K.D., Jr.; Bruggner, R.V.; Linderman, M.D.; Sachs, K.; Nolan, G.P.; Plevritis, S.K. Extracting a cellular hierarchy from high-dimensional cytometry data with SPADE. Nat. Biotechnol. 2011, 29, 886–891. [Google Scholar] [CrossRef] [Green Version]
- Van Gassen, S.; Callebaut, B.; Van Helden, M.J.; Lambrecht, B.N.; Demeester, P.; Dhaene, T.; Saeys, Y. FlowSOM: Using self-organizing maps for visualization and interpretation of cytometry data. Cytom. A 2015, 87, 636–645. [Google Scholar] [CrossRef]
- Amirel, A.D.; Davis, K.L.; Tadmor, M.D.; Simonds, E.F.; Levine, J.H.; Bendall, S.C.; Shenfeld, D.K.; Krishnaswamy, S.; Nolan, G.P.; Pe’er, D. viSNE enables visualization of high dimensional single-cell data and reveals phenotypic heterogeneity of leukemia. Nat. Biotechnol. 2013, 31, 545–552. [Google Scholar] [CrossRef] [Green Version]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2018, arXiv:1802.03426. [Google Scholar]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M.; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zagar, M.; et al. Integrated analysis of multimodal single-cell data. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mitchell, T.M. Machine Learning; McGraw-Hill, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Gonzalez-Dias, P.; Lee, E.K.; Sorgi, S.; de Lima, D.S.; Urbanski, A.H.; Silveira, E.L.; Nakaya, H.I. Methods for predicting vaccine immunogenicity and reactogenicity. Hum. Vaccine Immunother. 2020, 16, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Jojic, V.; Kidd, B.; Shen-Orr, S.; Price, J.; Jarrell, J.; Tse, T.; Huang, H.; Lund, P.; Maecker, H.T.; et al. Apoptosis and other immune biomarkers predict influenza vaccine responsiveness. Mol. Syst. Biol. 2013, 9, 659. [Google Scholar] [CrossRef] [PubMed]
- Tomic, A.; Tomic, I.; Rosenberg-Hasson, Y.; Dekker, C.L.; Maecker, H.T.; Davis, M.M. SIMON, an Automated Machine Learning System, Reveals Immune Signatures of Influenza Vaccine Responses. J. Immunol. 2019, 203, 749–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Sullivan, N.L.; Rouphael, N.; Yu, T.; Banton, S.; Maddur, M.S.; McCausland, M.; Chiu, C.; Canniff, J.; Dubey, S.; et al. Metabolic Phenotypes of Response to Vaccination in Humans. Cell 2017, 169, 862–877.e817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazmin, D.; Nakaya, H.I.; Lee, E.K.; Johnson, M.J.; van der Most, R.; van den Berg, R.A.; Ballou, W.R.; Jongert, E.; Wille-Reece, U.; Ockenhouse, C.; et al. Systems analysis of protective immune responses to RTS,S malaria vaccination in humans. Proc. Natl. Acad. Sci. USA 2017, 114, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, D.J.; Lythgoe, M.P. Application of “Systems Vaccinology” to Evaluate Inflammation and Reactogenicity of Adjuvanted Preventative Vaccines. J. Immunol. Res. 2015, 2015, 909406. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Tofte, J.; Vrahatis, A.; Josefsen, K.; Mehlsen, J.; Winther, K. Investigating the aetiology of adverse events following HPV vaccination with systems vaccinology. Cell Mol. Life Sci. 2019, 76, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Trtica-Majnaric, L.; Zekic-Susac, M.; Sarlija, N.; Vitale, B. Prediction of influenza vaccination outcome by neural networks and logistic regression. J. Biomed. Inf. 2010, 43, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, H.I.; Wrammert, J.; Lee, E.K.; Racioppi, L.; Marie-Kunze, S.; Haining, W.N.; Means, A.R.; Kasturi, S.P.; Khan, N.; Li, G.M.; et al. Systems biology of vaccination for seasonal influenza in humans. Nat. Immunol. 2011, 12, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.T.; Kennedy, R.B.; Grill, D.E.; Oberg, A.L.; Goergen, K.M.; Ovsyannikova, I.G.; Haralambieva, I.H.; Poland, G.A. Integration of Immune Cell Populations, mRNA-Seq, and CpG Methylation to Better Predict Humoral Immunity to Influenza Vaccination: Dependence of mRNA-Seq/CpG Methylation on Immune Cell Populations. Front. Immunol. 2017, 8, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvandeh, S.; Poland, G.A.; Kennedy, R.B.; McKinney, B.A. Multi-Level Model to Predict Antibody Response to Influenza Vaccine Using Gene Expression Interaction Network Feature Selection. Microorganisms 2019, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avey, S.; Mohanty, S.; Wilson, J.; Zapata, H.; Joshi, S.R.; Siconolfi, B.; Tsang, S.; Shaw, A.C.; Kleinstein, S.H. Multiple network-constrained regressions expand insights into influenza vaccination responses. Bioinformatics 2017, 33, i208–i216. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, E.; Bonduelle, O.; Soria, A.; Loulergue, P.; Rousseau, A.; Cachanado, M.; Bonnabau, H.; Thiebaut, R.; Tchitchek, N.; Behillil, S.; et al. Innate gene signature distinguishes humoral versus cytotoxic responses to influenza vaccination. J. Clin. Investig. 2019, 129, 1960–1971. [Google Scholar] [CrossRef]
- Wall, N.; Godlee, A.; Geh, D.; Jones, C.; Faustini, S.; Harvey, R.; Penn, R.; Chanouzas, D.; Nightingale, P.; O’Shea, M.; et al. Latent cytomegalovirus infection and previous capsular polysaccharide vaccination predict poor vaccine responses in older adults, independent of chronic kidney disease. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Babji, S.; Manickavasagam, P.; Chen, Y.H.; Jeyavelu, N.; Jose, N.V.; Praharaj, I.; Syed, C.; Kaliappan, S.P.; John, J.; Giri, S.; et al. Immune predictors of oral poliovirus vaccine immunogenicity among infants in South India. NPJ Vaccines 2020, 5, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janes, H.E.; Cohen, K.W.; Frahm, N.; De Rosa, S.C.; Sanchez, B.; Hural, J.; Magaret, C.A.; Karuna, S.; Bentley, C.; Gottardo, R.; et al. Higher T-Cell Responses Induced by DNA/rAd5 HIV-1 Preventive Vaccine Are Associated With Lower HIV-1 Infection Risk in an Efficacy Trial. J. Infect. Dis. 2017, 215, 1376–1385. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, L.; Janes, H.; Frahm, N.; Isaacs, A.; Kim, J.H.; Montefiori, D.; McElrath, M.J.; Tomaras, G.D.; Gilbert, P.B. Predictors of durable immune responses six months after the last vaccination in preventive HIV vaccine trials. Vaccine 2017, 35, 1184–1193. [Google Scholar] [CrossRef] [Green Version]
- Dérian, N.; Bellier, B.; Pham, H.P.; Tsitoura, E.; Kazazi, D.; Huret, C.; Mavromara, P.; Klatzmann, D.; Six, A. Early Transcriptome Signatures from Immunized Mouse Dendritic Cells Predict Late Vaccine-Induced T-Cell Responses. PLoS Comput. Biol. 2016, 12, e1004801. [Google Scholar] [CrossRef] [Green Version]
- Germain, R.N. Will Systems Biology Deliver Its Promise and Contribute to the Development of New or Improved Vaccines? What Really Constitutes the Study of “Systems Biology“ and How Might Such an Approach Facilitate Vaccine Design. Cold Spring Harb Perspect. Biol. 2018, 10, a033308. [Google Scholar] [CrossRef]
- Ravindran, R.; Loebbermann, J.; Nakaya, H.I.; Khan, N.; Ma, H.; Gama, L.; Machiah, D.K.; Lawson, B.; Hakimpour, P.; Wang, Y.C.; et al. The amino acid sensor GCN2 controls gut inflammation by inhibiting inflammasome activation. Nature 2016, 531, 523–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antia, R.; Ganusov, V.V.; Ahmed, R. The role of models in understanding CD8+ T-cell memory. Nat. Rev. Immunol. 2005, 5, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Le, D.; Miller, J.D.; Ganusov, V.V. Mathematical modeling provides kinetic details of the human immune response to vaccination. Front. Cell Infect. Microbiol. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.E.; Peña, J.M.; Jayamohan, J.; Jérusalem, A. Mechanistic models versus machine learning, a fight worth fighting for the biological community? Biol. Lett. 2018, 14, 20170660. [Google Scholar] [CrossRef]
- Barré-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Future Sci. Oa 2015, 1, Fso63. [Google Scholar] [CrossRef] [Green Version]
- Kiros, T.G.; Levast, B.; Auray, G.; Strom, S.; van Kessel, J.; Gerdts, V. The Importance of Animal Models in the Development of Vaccines. In Innovation in Vaccinology: From Design, through to Delivery and Testing; Baschieri, S., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 251–264. [Google Scholar]
- Klasse, P.J.; Nixon, D.F.; Moore, J.P. Immunogenicity of clinically relevant SARS-CoV-2 vaccines in nonhuman primates and humans. Sci. Adv. 2021, 7, eabe8065. [Google Scholar] [CrossRef]
- Rappuoli, R.; Black, S.; Bloom, D.E. Vaccines and global health: In search of a sustainable model for vaccine development and delivery. Sci. Transl. Med. 2019, 11, eaaw2888. [Google Scholar] [CrossRef]
- Plotkin, S.A.; Gilbert, P.B. Nomenclature for immune correlates of protection after vaccination. Clin. Infect. Dis 2012, 54, 1615–1617. [Google Scholar] [CrossRef]
- Van Damme, P.; Van Herck, K. A review of the long-term protection after hepatitis A and B vaccination. Travel Med. Infect. Dis. 2007, 5, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.; Cunningham, A.F.; Fooks, A.R. The immune response to rabies virus infection and vaccination. Vaccine 2010, 28, 3896–3901. [Google Scholar] [CrossRef]
- Wieten, R.W.; Jonker, E.F.; van Leeuwen, E.M.; Remmerswaal, E.B.; Ten Berge, I.J.; de Visser, A.W.; van Genderen, P.J.; Goorhuis, A.; Visser, L.G.; Grobusch, M.P.; et al. A Single 17D Yellow Fever Vaccination Provides Lifelong Immunity; Characterization of Yellow-Fever-Specific Neutralizing Antibody and T-Cell Responses after Vaccination. PLoS ONE 2016, 11, e0149871. [Google Scholar] [CrossRef] [Green Version]
- Center for Viral Systems Biology CViSB. SARS-CoV-2 (hCoV-19) Mutation Situation Reports. Available online: https://outbreak.info/situation-reports (accessed on 28 April 2021).
- Thakur, A.; Pedersen, L.E.; Jungersen, G. Immune markers and correlates of protection for vaccine induced immune responses. Vaccine 2012, 30, 4907–4920. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B. Systems vaccinology: Probing humanity‘s diverse immune systems with vaccines. Proc. Natl. Acad. Sci. USA 2014, 111, 12300–12306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epaulard, O.; Adam, L.; Poux, C.; Zurawski, G.; Salabert, N.; Rosenbaum, P.; Dereuddre-Bosquet, N.; Zurawski, S.; Flamar, A.L.; Oh, S.; et al. Macrophage-and neutrophil-derived TNF-α instructs skin langerhans cells to prime antiviral immune responses. J. Immunol. 2014, 193, 2416–2426. [Google Scholar] [CrossRef] [Green Version]
- Adam, L.; Tchitchek, N.; Todorova, B.; Rosenbaum, P.; Joly, C.; Poux, C.; Chapon, C.; Spetz, A.L.; Ustav, M.; Le Grand, R.; et al. Innate Molecular and Cellular Signature in the Skin Preceding Long-Lasting T Cell Responses after Electroporated DNA Vaccination. J. Immunol. 2020, 204, 3375–3388. [Google Scholar] [CrossRef]
- Liard, C.; Munier, S.; Arias, M.; Joulin-Giet, A.; Bonduelle, O.; Duffy, D.; Shattock, R.J.; Verrier, B.; Combadière, B. Targeting of HIV-p24 particle-based vaccine into differential skin layers induces distinct arms of the immune responses. Vaccine 2011, 29, 6379–6391. [Google Scholar] [CrossRef]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and T(H)1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- McMahan, K.; Yu, J.; Mercado, N.B.; Loos, C.; Tostanoski, L.H.; Chandrashekar, A.; Liu, J.; Peter, L.; Atyeo, C.; Zhu, A.; et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021, 590, 630–634. [Google Scholar] [CrossRef]
- Arrode-Brusés, G.; Moussa, M.; Baccard-Longere, M.; Villinger, F.; Chebloune, Y. Long-term central and effector SHIV-specific memory T cell responses elicited after a single immunization with a novel lentivector DNA vaccine. PLoS ONE 2014, 9, e110883. [Google Scholar] [CrossRef]
- Hansen, S.G.; Zak, D.E.; Xu, G.; Ford, J.C.; Marshall, E.E.; Malouli, D.; Gilbride, R.M.; Hughes, C.M.; Ventura, A.B.; Ainslie, E.; et al. Prevention of tuberculosis in rhesus macaques by a cytomegalovirus-based vaccine. Nat. Med. 2018, 24, 130–143. [Google Scholar] [CrossRef]
- Elhmouzi-Younes, J.; Palgen, J.L.; Tchitchek, N.; Delandre, S.; Namet, I.; Bodinham, C.L.; Pizzoferro, K.; Lewis, D.J.M.; Le Grand, R.; Cosma, A.; et al. In depth comparative phenotyping of blood innate myeloid leukocytes from healthy humans and macaques using mass cytometry. Cytom. A 2017, 91, 969–982. [Google Scholar] [CrossRef]
- Mentzer, A.J.; O’Connor, D.; Pollard, A.J.; Hill, A.V. Searching for the human genetic factors standing in the way of universally effective vaccines. Philos. Trans. R Soc. Lond B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef]
- Weinberg, A.; Pang, L.; Johnson, M.J.; Caldas, Y.; Cho, A.; Tovar-Salazar, A.; Canniff, J.; Schmader, K.E.; Popmihajlov, Z.; Levin, M.J. The Effect of Age on the Immunogenicity of the Live Attenuated Zoster Vaccine Is Predicted by Baseline Regulatory T Cells and Varicella-Zoster Virus-Specific T Cell Immunity. J. Virol 2019, 93, e00305-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heiden, M.; Berbers, G.A.M.; Fuentes, S.; van Zelm, M.C.; Boots, A.M.H.; Buisman, A.M. An Explorative Biomarker Study for Vaccine Responsiveness after a Primary Meningococcal Vaccination in Middle-Aged Adults. Front. Immunol. 2017, 8, 1962. [Google Scholar] [CrossRef]
- Alcorn, J.F.; Avula, R.; Chakka, A.B.; Schwarzmann, W.E.; Nowalk, M.P.; Lin, C.J.; Ortiz, M.A.; Horne, W.T.; Chandran, U.R.; Nagg, J.P.; et al. Differential gene expression in peripheral blood mononuclear cells from children immunized with inactivated influenza vaccine. Hum. Vaccine Immunother. 2020, 16, 1782–1790. [Google Scholar] [CrossRef]
- Idoko, O.T.; Smolen, K.K.; Wariri, O.; Imam, A.; Shannon, C.P.; Dibassey, T.; Diray-Arce, J.; Darboe, A.; Strandmark, J.; Ben-Othman, R.; et al. Clinical Protocol for a Longitudinal Cohort Study Employing Systems Biology to Identify Markers of Vaccine Immunogenicity in Newborn Infants in The Gambia and Papua New Guinea. Front. Pediatr. 2020, 8, 197. [Google Scholar] [CrossRef]
- Merino, K.M.; Slisarenko, N.; Taylor, J.M.; Falkenstein, K.P.; Gilbert, M.H.; Bohm, R.P.; Blanchard, J.L.; Ardeshir, A.; Didier, E.S.; Kim, W.K.; et al. Clinical and Immunological Metrics During Pediatric Rhesus Macaque Development. Front. Pediatr. 2020, 8, 388. [Google Scholar] [CrossRef]
- Terao, K. Essentials for starting a pediatric clinical study (3): Dynamic changes in early development of immune system in macaque monkeys—the significance from standpoint of preclinical toxicity test using nonhuman primates. J. Toxicol. Sci. 2009, 34 (Suppl. 2), SP321–SP325. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Xu, H.; Liu, D.; Veazey, R.S.; Wang, X. Development of serum antibodies during early infancy in rhesus macaques: Implications for humoral immune responses to vaccination at birth. Vaccine 2014, 32, 5337–5342. [Google Scholar] [CrossRef] [Green Version]
- Levast, B.; Schulz, S.; Hurk, S.; Gerdts, V. Animal models for neonatal diseases in humans. Vaccine 2013, 31, 2489–2499. [Google Scholar] [CrossRef] [PubMed]
- Ramos, L.; Lunney, J.K.; Gonzalez-Juarrero, M. Neonatal and infant immunity for tuberculosis vaccine development: Importance of age-matched animal models. Dis. Model. Mech. 2020, 13, dmm045740. [Google Scholar] [CrossRef]
- Rosario, M.; Fulkerson, J.; Soneji, S.; Parker, J.; Im, E.J.; Borthwick, N.; Bridgeman, A.; Bourne, C.; Joseph, J.; Sadoff, J.C.; et al. Safety and immunogenicity of novel recombinant BCG and modified vaccinia virus Ankara vaccines in neonate rhesus macaques. J. Virol. 2010, 84, 7815–7821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesarwani, A.; Sahu, P.; Jain, K.; Sinha, P.; Mohan, K.V.; Nagpal, P.S.; Singh, S.; Zaidi, R.; Nagarajan, P.; Upadhyay, P. The safety and efficacy of BCG encapsulated alginate particle (BEAP) against M.tb H37Rv infection in Macaca mulatta: A pilot study. Sci. Rep. 2021, 11, 3049. [Google Scholar] [CrossRef] [PubMed]
- Abel, K. The rhesus macaque pediatric SIV infection model—A valuable tool in understanding infant HIV-1 pathogenesis and for designing pediatric HIV-1 prevention strategies. Curr. Hiv Res. 2009, 7, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, A.D., 2nd; Dennis, M.; Eudailey, J.; Walter, K.L.; Cronin, K.; Alam, S.M.; Choudhary, N.; Tuck, R.H.; Hudgens, M.; Kozlowski, P.A.; et al. HIV Env-Specific IgG Antibodies Induced by Vaccination of Neonatal Rhesus Macaques Persist and Can Be Augmented by a Late Booster Immunization in Infancy. mSphere 2020, 5, e00162-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bricker, K.M.; Obregon-Perko, V.; Uddin, F.; Williams, B.; Uffman, E.A.; Garrido, C.; Fouda, G.G.; Geleziunas, R.; Robb, M.; Michael, N.; et al. Therapeutic vaccination of SIV-infected, ART-treated infant rhesus macaques using Ad48/MVA in combination with TLR-7 stimulation. PLoS Pathog. 2020, 16, e1008954. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Bradley, T.; Williams, W.B.; Cain, D.W.; Montefiori, D.C.; Saunders, K.O.; Parks, R.J.; Edwards, R.W.; Ferrari, G.; Mueller, O.; et al. Neonatal Rhesus Macaques Have Distinct Immune Cell Transcriptional Profiles following HIV Envelope Immunization. Cell Rep. 2020, 30, 1553–1569.e1556. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.J.; Hill, A.V. Human genetic susceptibility to infectious disease. Nat. Rev. Genet. 2012, 13, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Noll, K.E.; Ferris, M.T.; Heise, M.T. The Collaborative Cross: A Systems Genetics Resource for Studying Host-Pathogen Interactions. Cell Host Microbe 2019, 25, 484–498. [Google Scholar] [CrossRef] [Green Version]
- Olafsdottir, T.A.; Lindqvist, M.; Nookaew, I.; Andersen, P.; Maertzdorf, J.; Persson, J.; Christensen, D.; Zhang, Y.; Anderson, J.; Khoomrung, S.; et al. Comparative Systems Analyses Reveal Molecular Signatures of Clinically tested Vaccine Adjuvants. Sci. Rep. 2016, 6, 39097. [Google Scholar] [CrossRef] [Green Version]
- Kasturi, S.P.; Kozlowski, P.A.; Nakaya, H.I.; Burger, M.C.; Russo, P.; Pham, M.; Kovalenkov, Y.; Silveira, E.L.V.; Havenar-Daughton, C.; Burton, S.L.; et al. Adjuvanting a Simian Immunodeficiency Virus Vaccine with Toll-Like Receptor Ligands Encapsulated in Nanoparticles Induces Persistent Antibody Responses and Enhanced Protection in TRIM5α Restrictive Macaques. J. Virol. 2017, 91, e01844-16. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E.A.; Ols, S.; Miura, K.; Rausch, K.; Narum, D.L.; Spångberg, M.; Juraska, M.; Wille-Reece, U.; Weiner, A.; Howard, R.F.; et al. TLR-adjuvanted nanoparticle vaccines differentially influence the quality and longevity of responses to malaria antigen Pfs25. JCI Insight 2018, 3, e120692. [Google Scholar] [CrossRef]
- Ciabattini, A.; Pettini, E.; Fiorino, F.; Lucchesi, S.; Pastore, G.; Brunetti, J.; Santoro, F.; Andersen, P.; Bracci, L.; Pozzi, G.; et al. Heterologous Prime-Boost Combinations Highlight the Crucial Role of Adjuvant in Priming the Immune System. Front. Immunol. 2018, 9, 380. [Google Scholar] [CrossRef]
- Elsner, R.A.; Shlomchik, M.J. Germinal Center and Extrafollicular B Cell Responses in Vaccination, Immunity, and Autoimmunity. Immunity 2020, 53, 1136–1150. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Yin, Q. B Cell Response to Vaccination. Immunol. Investig. 2021, 1–22. [Google Scholar] [CrossRef]
- Ben-Othman, R.; Cai, B.; Liu, A.C.; Varankovich, N.; He, D.; Blimkie, T.M.; Lee, A.H.; Gill, E.E.; Novotny, M.; Aevermann, B.; et al. Systems Biology Methods Applied to Blood and Tissue for a Comprehensive Analysis of Immune Response to Hepatitis B Vaccine in Adults. Front. Immunol. 2020, 11, 580373. [Google Scholar] [CrossRef]
- Eslamizar, L.; Petrovas, C.; Leggat, D.J.; Furr, K.; Lifton, M.L.; Levine, G.; Ma, S.; Fletez-Brant, C.; Hoyland, W.; Prabhakaran, M.; et al. Recombinant MVA-prime elicits neutralizing antibody responses by inducing antigen-specific B cells in the germinal center. NPJ Vaccines 2021, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- de Bree, L.C.J.; Koeken, V.; Joosten, L.A.B.; Aaby, P.; Benn, C.S.; van Crevel, R.; Netea, M.G. Non-specific effects of vaccines: Current evidence and potential implications. Semin. Immunol. 2018, 39, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Joosten, L.A.; Latz, E.; Mills, K.H.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, aaf1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covián, C.; Fernández-Fierro, A.; Retamal-Díaz, A.; Díaz, F.E.; Vasquez, A.E.; Lay, M.K.; Riedel, C.A.; González, P.A.; Bueno, S.M.; Kalergis, A.M. BCG-Induced Cross-Protection and Development of Trained Immunity: Implication for Vaccine Design. Front. Immunol. 2019, 10, 2806. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: Can it offer protection against COVID-19? Nat. Rev. Immunol. 2020, 20, 335–337. [Google Scholar] [CrossRef]
- Palgen, J.L.; Feraoun, Y.; Dzangué-Tchoupou, G.; Joly, C.; Martinon, F.; Le Grand, R.; Beignon, A.S. Optimize Prime/Boost Vaccine Strategies: Trained Immunity as a New Player in the Game. Front. Immunol. 2021, 12, 612747. [Google Scholar] [CrossRef]
- Sarkar, S.; Heise, M.T. Mouse Models as Resources for Studying Infectious Diseases. Clinthera 2019, 41, 1912–1922. [Google Scholar] [CrossRef] [Green Version]
- Ermann, J.; Glimcher, L.H. After GWAS: Mice to the rescue? Curr. Opin Immunol. 2012, 24, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, R.; Khan, N.; Nakaya, H.I.; Li, S.; Loebbermann, J.; Maddur, M.S.; Park, Y.; Jones, D.P.; Chappert, P.; Davoust, J.; et al. Vaccine activation of the nutrient sensor GCN2 in dendritic cells enhances antigen presentation. Science 2014, 343, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Ng’uni, T.; Chasara, C.; Ndhlovu, Z.M. Major Scientific Hurdles in HIV Vaccine Development: Historical Perspective and Future Directions. Front. Immunol. 2020, 11, 590780. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Eraslan, B.; Wieland, T.; Hallström, B.; Hopf, T.; Zolg, D.P.; Zecha, J.; Asplund, A.; Li, L.H.; Meng, C.; et al. A deep proteome and transcriptome abundance atlas of 29 healthy human tissues. Mol. Syst. Biol. 2019, 15, e8503. [Google Scholar] [CrossRef]
- Maecker, H.T.; McCoy, J.P.; Nussenblatt, R. Standardizing immunophenotyping for the Human Immunology Project. Nat. Rev. Immunol. 2012, 12, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeys, Y.; Van Gassen, S.; Lambrecht, B.N. Computational flow cytometry: Helping to make sense of high-dimensional immunology data. Nat. Rev. Immunol. 2016, 16, 449–462. [Google Scholar] [CrossRef]
- Fruhwirth, G.O.; Kneilling, M.; de Vries, I.J.M.; Weigelin, B.; Srinivas, M.; Aarntzen, E. The Potential of In Vivo Imaging for Optimization of Molecular and Cellular Anti-cancer Immunotherapies. Mol. Imaging Biol. 2018, 20, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torcellan, T.; Stolp, J.; Chtanova, T. In Vivo Imaging Sheds Light on Immune Cell Migration and Function in Cancer. Front. Immunol. 2017, 8, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Machine Learning Algorithm | Principle | Advantages | Drawbacks |
|---|---|---|---|
| Linear regression | It assumes a linear relationship between input variables and output and thus, attempts to model this relationship by fitting a linear equation to the observed data There are several implementations of this model, of which the most commonly used is ordinary least squares, which tends to minimize the residual sum of the squares between the observed and predicted targets. |
|
|
| Linear discriminant analysis (LDA) | It is used to identify to which class samples belong to, certain statistical properties of the data are first calculated and then substituted into the LDA equation. The statistical properties consist of the mean and variance for the case of a single input and the means and covariance matrix for multiple inputs. |
|
|
| Random Forest | It builds a number of decision trees on bootstrapped training sets and considers a random sample of m predictors to be split candidates from the full set of p predictors to overcome the problem of high variance. Therefore, on average, the strong predictor is not considered and other predictors have a better chance. This process can be thought of as decorrelating of the trees, thereby making the average of the resulting trees less variable and hence more accurate and reliable. |
|
|
| Support vector machine | It converts a non-linear separable problem by transforming it onto another higher dimensional space and thus, the problem becomes linearly separable. This is accomplished using various types of so-called kernel functions. Then, classification is performed by finding the hyperplane that well separates the classes of samples. |
|
|
| Discriminant analysis via mixed integer programming (DAMIP) | It is a classification model based on a very powerful supervised-learning approach used primarily in the biomedical field. It is a discrete support vector machine coupled with a powerful embedded feature-selection module [176]. |
|
|
| Vaccine | Vaccinees | Predicted Responses | Predictors | Machine Learning Method | Performance * | Reference |
|---|---|---|---|---|---|---|
| Yellow fever vaccine (YF-17D) | Healthy adults | The magnitude of the activated CD8+ T cell and neutralizing Ab responses | Early blood transcriptional signatures | ClaNC and DAMIP | Up to 90% and 100% respectively | [52] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) | Patients 50–89 years old suffering from multiple chronic medical conditions | The magnitude of plasma HAI Ab response | Baseline signatures among 26 input continuous or categorical variables inc. previous vaccination, low grade chronic inflammation, chronic infections, blood cell counts | Neural network (multilayer perceptron (MLP), radial-basis function network (RBFN) and probabilistic network (PNN)) and Logistic regression | 72.5% of average hit rate across 10 samples | [184] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) | Healthy adults | The magnitude of plasma HAI Ab response | Early blood transcriptional signatures | DAMIP | Up to 90% | [185] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) | Healthy adults, inc. young (20–30 years) and older subjects (60 to 89 years) | The magnitude of plasma HAI Ab response | Baseline blood transcriptional, cytokines and cell populations signatures | Logistic regression | 84% | [178] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) and pandemic H1N1 (pH1N1) vaccine | Healthy adults | The magnitude of the Ab response | Baseline HAI titer, blood cell populations, transcripts and pathways signatures | Diagonal linear discriminant analysis (for cell frequency data and when cell frequency and pathway status were combined); or partial least square (for data dimension reduction due to the large number of genes) followed by linear discriminant analysis (PLS-LDA) for transcript data alone | 0.86 of AUROC | [60] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) over 5 seasons | Human adults, inc. elderlies (>65 years) | The magnitude of plasma HAI Ab response | Early blood transcriptional signatures | DAMIP and artificial neural network classifier | >80% | [10] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) | Healthy adults (50 to 74 years) | The magnitude of the B-cell ELISPOT and plasma HAI Ab responses | Early blood cell composition, mRNA-Seq, and DNA methylation signatures | The ensemble learner (inc. Generalized linear models, Recursive Partitioning, and Regression Trees), and random forest models | 0.64–0.79 of AUROC | [186] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) | Healthy adults | The magnitude of plasma HAI Ab response | Baseline HAI titer and blood transcriptional signatures | Gaussian Mixture Model (GMM) | R2 = 0.64 for the correlation between observed and predicted data | [187] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) | Healthy adults | The magnitude of the Ab response | Early blood transcriptional signatures | Logistic Multiple Network-constrained Regression | 69% | [188] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) over 8 seasons | Healthy adults | The magnitude of the specific Ab response | Baseline blood cell populations signatures | 128 machine learning algorithms suitable for classification using Sequential Iterative Modeling “OverNight” (SIMON), inc. Diagonal Discriminant Analysis, Partial Least Squares, Linear Discriminant Analysis, Logic Regression, Neural Network, Random Forest | Up to 0.92 of AUROC | [179] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) given transcutaneously, intradermally or intramuscularly | Healthy adults | The magnitude of the specific T CD8+ and Ab responses | Early blood transcriptional and serum cytokines signatures | Logistic regression | 0.93 to 0.96 of AUROC | [189] |
| Seasonal Trivalent Inactivated influenza Vaccine (TIV) and 23-valent pneumococcal polysaccharide vaccine | Old patients (>65 years) with chronic kidney disease with or without non-dialysis | The magnitude of the HAI Ab and anti-PnPS IgG responses | Baseline signatures among 30 input continuous or categorical variables inc. previous vaccinations, low grade chronic inflammation, chronic infections, blood cell counts | Multivariable linear regression model | p < 0.05 | [190] |
| RTS,S malaria vaccine | Healthy adults | The protection against CHMI | Early blood transcriptional signatures | DAMIP | >80% | [181] |
| Candidate malaria vaccine composed of a Self-Assembling Protein Nanoparticles presenting the malarial circumsporozoite protein (CSP) adjuvanted with three different liposomal formulations: liposome plus Alum, liposome plus QS21, or both | Rhesus macaques | Adjuvant condition | Vaccine-induced immune response signatures among many variables inc. serology, fluorospot, ICS from blood, liver, LN and spleen | Random forest followed by Linear regression analysis | 92% | [32] |
| Live-attenuated varicella zoster virus (VZV) vaccine | Healthy adults, inc. younger (25–40 years) and older (60–79 years) | The magnitude of the specific T and IgG responses | Early blood transcriptional, metabolite clusters, cytokines, and cell populations signatures | Multivariate regression model (Partial least square) | p < 0.05 | [180] |
| Monovalent oral polio vaccine type 3 (mOPV3) | Infants aged 6–11 months | Seroconversion or shedding of vaccine virus as a marker of vaccine “take” | Baseline enteric pathogens blood cell populations, and plasma cytokines signatures | Random forest | 58% | [191] |
| Two distinct live attenuated Tularemia vaccine administered by scarification | Healthy humans | The magnitude of the specific Ab and activated CD4 and CD8 T cell responses | Early blood transcriptional signatures | Logistic regression | 26% of mean misclassification error | [39] |
| rVSV-ZEBOV | Healthy adults | The magnitude of the Ab response | Early blood transcriptional, plasma cytokine and cell populations signatures | Sparse partial least-squares followed by multivariable linear regression | 0.77 of root square residuals leave-one-out explaining 55% of the variability | [12] |
| DNA/rAd5 HIV-1 preventive candidate vaccine | Healthy adults | HIV infection | Magnitude and quality of CD4 and CD8 T cells | PCA followed by Cox proportional hazards regression model, and Logistic regression with lasso | Up to 0.75 of AUROC | [192] |
| Seven preventive HIV-1 vaccine regimens (inc. DNA, NYVAC, ALVAC, MVA, AIDSVAX) | Healthy adults | The magnitude of long-term immune responses | Baseline demographic variables and peak immune responses | Regularized random forest and linear regression models | R = 0.91 for the correlation between observed andpredicted data | [193] |
| 41 different vaccine vectors all expressing the same antigen | Mice | The quality of late T-cell responses | Early transcriptome of dendritic cells | Random forest | Up to 98% | [194] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Tilbeurgh, M.; Lemdani, K.; Beignon, A.-S.; Chapon, C.; Tchitchek, N.; Cheraitia, L.; Marcos Lopez, E.; Pascal, Q.; Le Grand, R.; Maisonnasse, P.; et al. Predictive Markers of Immunogenicity and Efficacy for Human Vaccines. Vaccines 2021, 9, 579. https://doi.org/10.3390/vaccines9060579
Van Tilbeurgh M, Lemdani K, Beignon A-S, Chapon C, Tchitchek N, Cheraitia L, Marcos Lopez E, Pascal Q, Le Grand R, Maisonnasse P, et al. Predictive Markers of Immunogenicity and Efficacy for Human Vaccines. Vaccines. 2021; 9(6):579. https://doi.org/10.3390/vaccines9060579
Chicago/Turabian StyleVan Tilbeurgh, Matthieu, Katia Lemdani, Anne-Sophie Beignon, Catherine Chapon, Nicolas Tchitchek, Lina Cheraitia, Ernesto Marcos Lopez, Quentin Pascal, Roger Le Grand, Pauline Maisonnasse, and et al. 2021. "Predictive Markers of Immunogenicity and Efficacy for Human Vaccines" Vaccines 9, no. 6: 579. https://doi.org/10.3390/vaccines9060579
APA StyleVan Tilbeurgh, M., Lemdani, K., Beignon, A. -S., Chapon, C., Tchitchek, N., Cheraitia, L., Marcos Lopez, E., Pascal, Q., Le Grand, R., Maisonnasse, P., & Manet, C. (2021). Predictive Markers of Immunogenicity and Efficacy for Human Vaccines. Vaccines, 9(6), 579. https://doi.org/10.3390/vaccines9060579