
CpG-ODN Signaling via Dendritic Cells-Expressing MyD88, but Not IL-10, Inhibits Allergic Sensitization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Allergen
2.3. Alum Gel Preparation
2.4. Experimental Protocol
2.5. Generation of Bone Marrow-Derived Dendritic Cells
2.6. Bronchoalveolar Lavage (BAL) Fluid Collection
2.7. ELISA for Cytokines
2.8. Lung Histopathology Analysis
2.9. Statistical Analyses
3. Results
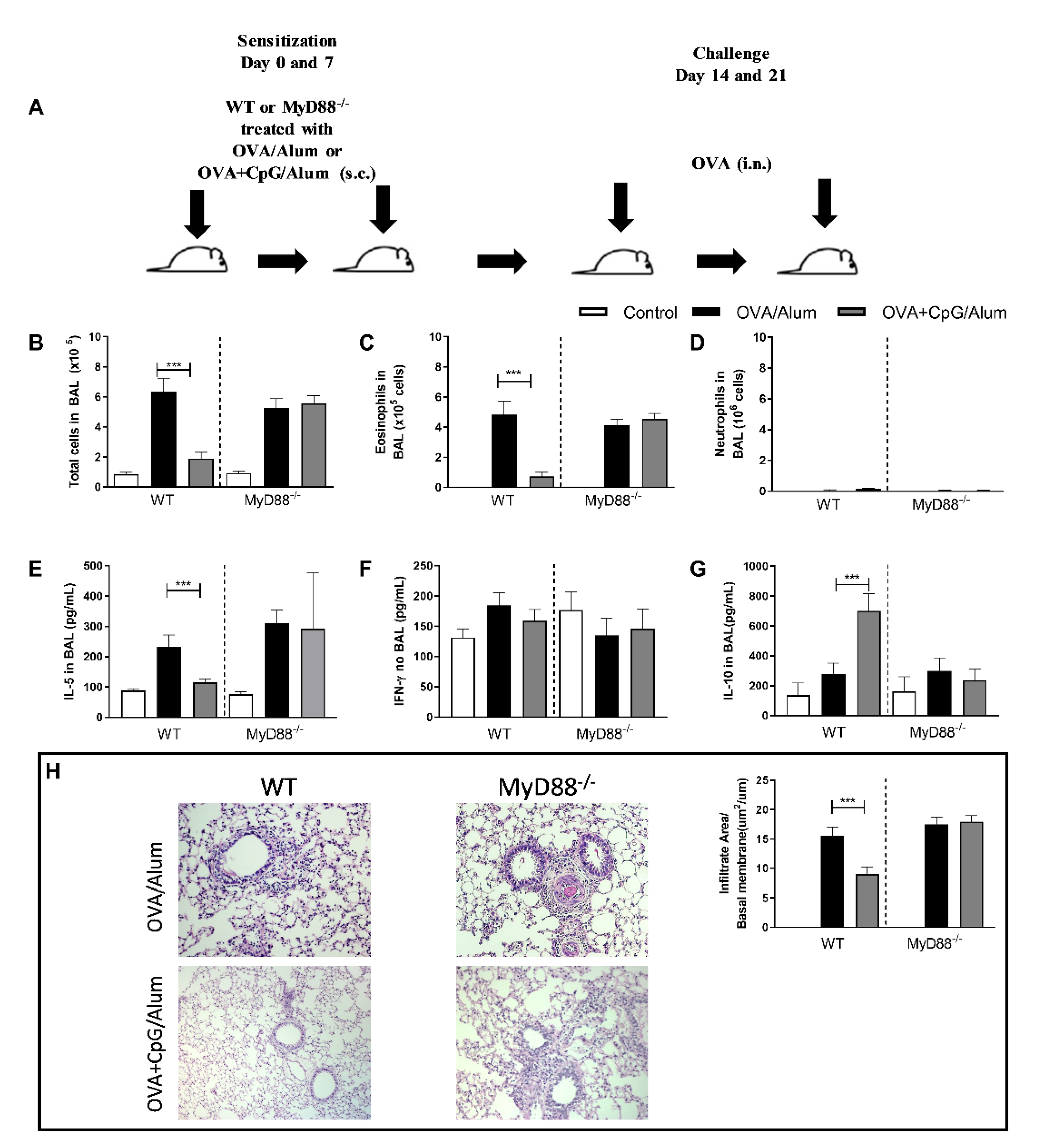
3.1. Inhibition of Th2 Allergic Immunity by CpG-ODN Is MyD88-Dependent and Associated with Increased Production of IL-10 in Lung
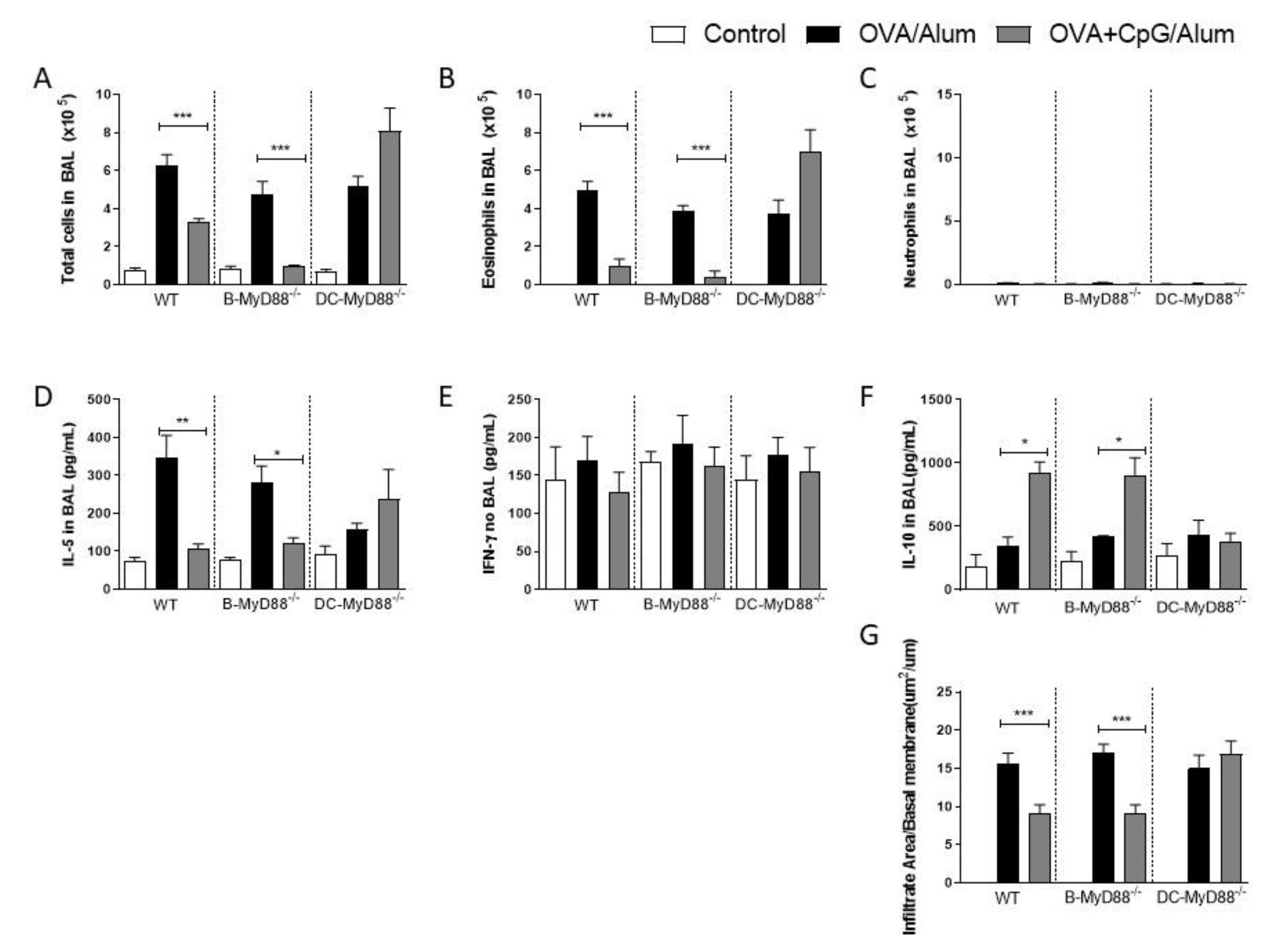
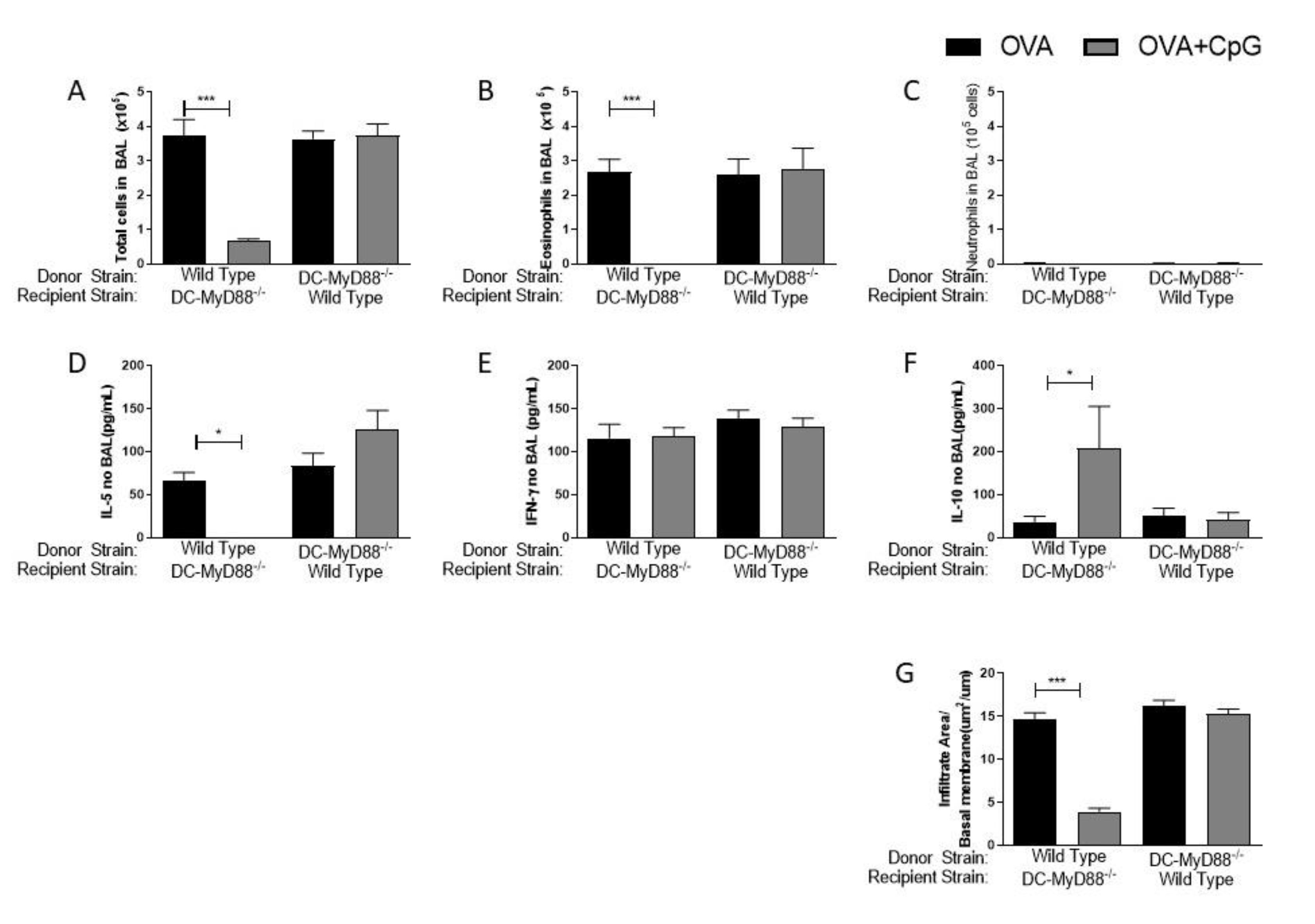
3.2. MyD88-Expressing Dendritic Cells, but Not B Cells, Are Necessary for the Inhibition of Allergic Sensitization by CpG-ODN Signaling
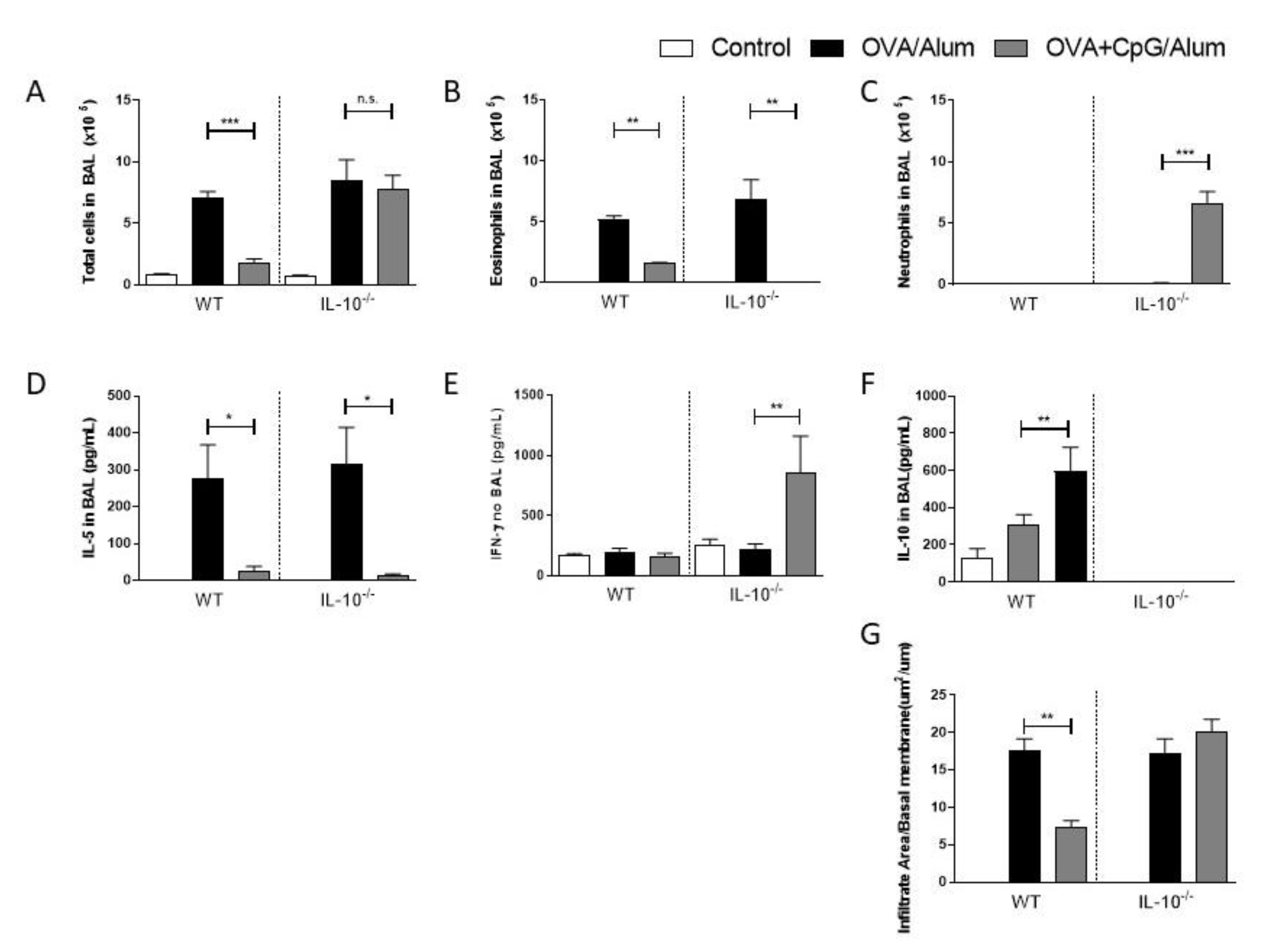
3.3. CpG-ODN in IL-10-Deficient Mice Induces Immune-Deviation towards a Th1-Dominated Airway Inflammation
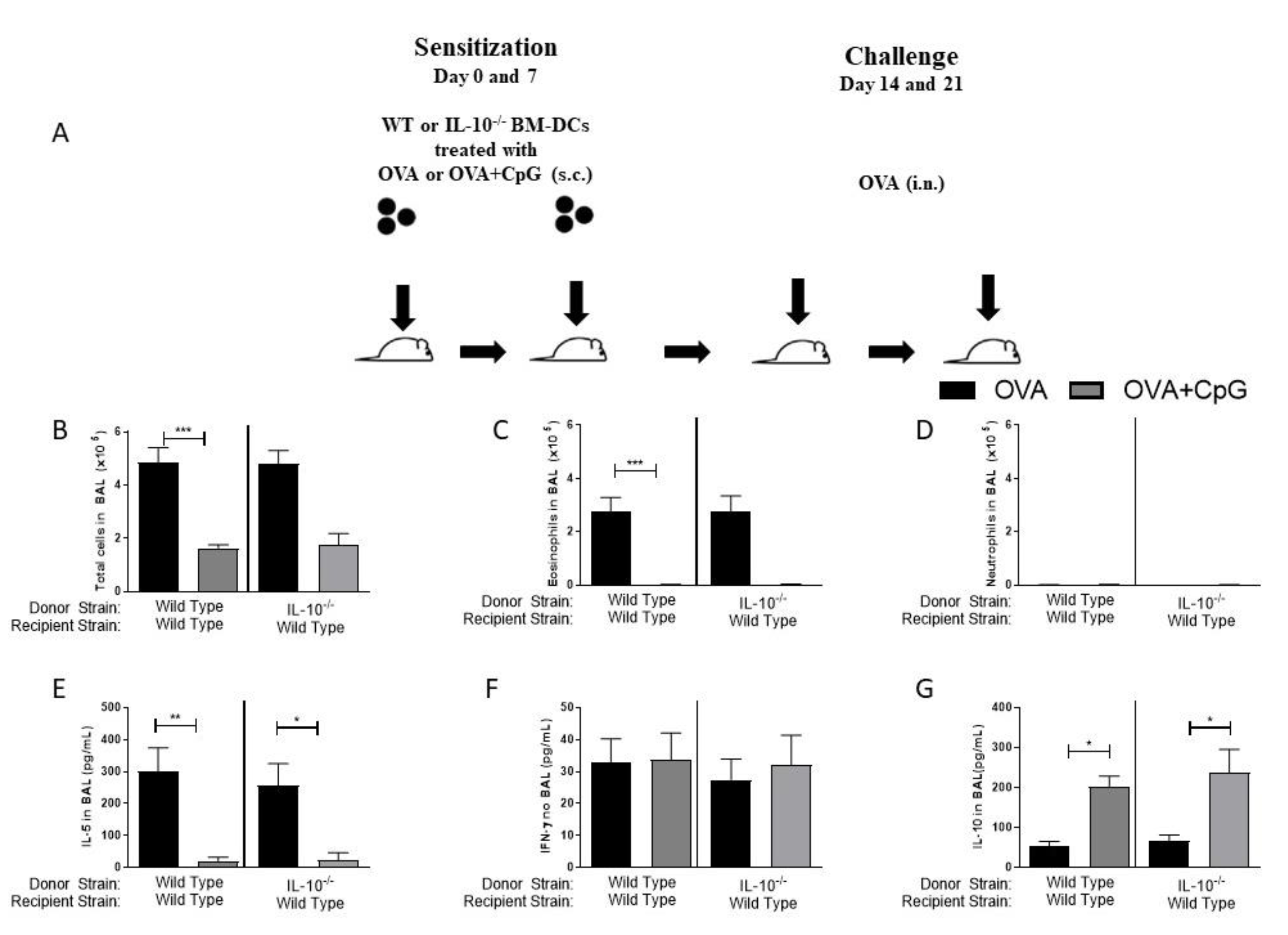
3.4. Sensitization with Bone Marrow-Derived Dendritic Cells Lacking IL-10 and Primed with OVA and CpG-ODN Prevent Allergic Lung Inflammation but Does Not Induce Th1 Immunity
3.5. MyD88 Expression on BM-DCs Mediates the CpG-ODN Anti-Allergic Effect
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zakeri, A.; Russo, M. Dual role of toll-like receptors in human and experimental asthma models. Front. Immunol. 2018, 9, 1027. [Google Scholar] [CrossRef] [Green Version]
- Karacs, J.; Reithofer, M.; Kitzmüller, C.; Kraller, M.; Schmalz, S.; Bleichert, S.; Huppa, J.B.; Stockinger, H.; Bohle, B.; Jahn-Schmid, B. Adjuvants and Vaccines Used in Allergen-Specific Immunotherapy Induce Neutrophil Extracellular Traps. Vaccines 2021, 9, 321. [Google Scholar] [CrossRef] [PubMed]
- Shirota, H.; Klinman, D.M. CpG Oligodeoxynucleotides as Adjuvants for Clinical Use. In Immunopotentiators in Modern Vaccines, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 163–198. ISBN 9780128040195. [Google Scholar]
- Casella, C.R.; Mitchell, T.C. Putting endotoxin to work for us: Monophosphoryl lipid a as a safe and effective vaccine adjuvant. Cell. Mol. Life Sci. 2008, 65, 3231–3240. [Google Scholar] [CrossRef] [Green Version]
- Steinhagen, F.; Kinjo, T.; Bode, C.; Klinman, D.M. TLR-based immune adjuvants. Vaccine 2011, 29, 3341–3355. [Google Scholar] [CrossRef] [Green Version]
- Hyer, R.; McGuire, D.K.; Xing, B.; Jackson, S.; Janssen, R. Safety of a two-dose investigational hepatitis B vaccine, HBsAg-1018, using a toll-like receptor 9 agonist adjuvant in adults. Vaccine 2018, 36, 2604–2611. [Google Scholar] [CrossRef]
- Moser, M.; Murphy, K.M. Dendritic cell regulation of TH1-TH2 development. Nat. Immunol. 2000, 1, 199–205. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. Lung dendritic cells in respiratory viral infection and asthma: From protection to immunopathology. Annu. Rev. Immunol. 2012, 30, 243–270. [Google Scholar] [CrossRef]
- Esterházy, D.; Loschko, J.; London, M.; Jove, V.; Oliveira, T.Y.; Mucida, D. Classical dendritic cells are required for dietary antigen-mediated induction of peripheral T reg cells and tolerance. Nat. Immunol. 2016, 17, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Kowalewicz-Kulbat, M.; Szpakowski, P.; Krawczyk, K.T.; Kowalski, M.L.; Kosinski, S.; Biet, F.; Rudnicka, W.; Locht, C. Decrease of il-5 production by naive t cells cocultured with il-18-producing bcg-pulsed dendritic cells from patients allergic to house dust mite. Vaccines 2021, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Jay, D.C.; Nadeau, K.C. Immune Mechanisms of Sublingual Immunotherapy. Curr. Allergy Asthma Rep. 2014, 14, 473. [Google Scholar] [CrossRef] [PubMed]
- Samarasinghe, R.; Tailor, P.; Tamura, T.; Kaisho, T.; Akira, S.; Ozato, K. Induction of an anti-inflammatory cytokine, IL-10, in dendritic cells after toll-like receptor signaling. J. Interf. Cytokine Res. 2006. [Google Scholar] [CrossRef] [PubMed]
- Gavett, S.H.; O’Hearn, D.J.; Li, X.; Huang, S.K.; Finkelman, F.D.; Wills-Karp, M. Interleukin 12 inhibits antigen-induced airway hyperresponsiveness, inflammation, and Th2 cytokine expression in mice. J. Exp. Med. 1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicari, A.P.; Chiodoni, C.; Vaure, C.; Aït-Yahia, S.; Dercamp, C.; Matsos, F.; Reynard, O.; Taverne, C.; Merle, P.; Colombo, M.P.; et al. Reversal of Tumor-induced Dendritic Cell Paralysis by CpG Immunostimulatory Oligonucleotide and Anti–Interleukin 10 Receptor Antibody. J. Exp. Med. 2002. [Google Scholar] [CrossRef] [PubMed]
- Mirotti, L.; Custódio, R.W.A.; Gomes, E.; Rammauro, F.; de Araujo, E.F.; Calich, V.L.G.; Russo, M. CpG-ODN shapes alum adjuvant activity signaling Via MyD88 and IL-10. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Nunes, F.P.B.; Alberca-Custódio, R.W.; Gomes, E.; Fonseca, D.M.; Yokoyama, N.H.; Labrada, A.; Russo, M. TLR9 agonist adsorbed to alum adjuvant prevents asthma-like responses induced by Blomia tropicalis mite extract. J. Leukoc. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Custodio, R.W.A.; Mirotti, L.; Gomes, E.; Nunes, F.P.B.; Vieira, R.S.; Graça, L.; Almeida, R.R.; Câmara, N.O.S.; Russo, M. Dendritic Cells Expressing MyD88 Molecule Are Necessary and Sufficient for CpG-Mediated Inhibition of IgE Production In Vivo. Cells 2019, 8, 1165. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Li, J.; Wu, K.; Azhati, B.; Rexiati, M. Culture and Identification of Mouse Bone Marrow-Derived Dendritic Cells and Their Capability to Induce T Lymphocyte Proliferation. Med. Sci. Monit. 2016. [Google Scholar] [CrossRef]
- Besnard, A.G.; Guillou, N.; Tschopp, J.; Erard, F.; Couillin, I.; Iwakura, Y.; Quesniaux, V.; Ryffel, B.; Togbe, D. NLRP3 inflammasome is required in murine asthma in the absence of aluminum adjuvant. Allergy Eur. J. Allergy Clin. Immunol. 2011, 66, 1047–1057. [Google Scholar] [CrossRef]
- Thomas, S.Y.; Whitehead, G.S.; Takaku, M.; Ward, J.M.; Xu, X.; Nakano, K.; Lyons-Cohen, M.R.; Nakano, H.; Gowdy, K.M.; Wade, P.A.; et al. MyD88-dependent dendritic and epithelial cell crosstalk orchestrates immune responses to allergens. Mucosal Immunol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Eisenbarth, S.C.; Piggott, D.A.; Huleatt, J.W.; Visintin, I.; Herrick, C.A.; Bottomly, K. Lipopolysaccharide-enhanced, Toll-like Receptor 4–dependent T Helper Cell Type 2 Responses to Inhaled Antigen. J. Exp. Med. 2002. [Google Scholar] [CrossRef] [Green Version]
- Eisenbarth, S.C. Use and limitations of alum-based models of allergy. Clin. Exp. Allergy 2008, 38, 1572–1575. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, D.M.; Wowk, P.F.; Paula, M.O.; Gembre, A.F.; Baruffi, M.D.; Fermino, M.L.; Turato, W.M.; Campos, L.W.; Silva, C.L.; Ramos, S.G.; et al. Requirement of MyD88 and Fas pathways for the efficacy of allergen-free immunotherapy. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 275–284. [Google Scholar] [CrossRef]
- Fiorentino, D.F.; Bond, M.W.; Mosmann, T.R. Two types of mouse t helper cell: IV. Th2 clones secrete a factor that inhibits cytokine production by Thl clones. J. Exp. Med. 1989. [Google Scholar] [CrossRef] [PubMed]
- Zuany-Amorim, C.; Hailé, S.; Leduc, D.; Dumarey, C.; Huerre, M.; Vargaftig, B.B.; Pretolani, M. Interleukin-10 inhibits antigen-induced cellular recruitment into the airways of sensitized mice. J. Clin. Investig. 1995. [Google Scholar] [CrossRef] [Green Version]
- Waibler, Z.; Anzaghe, M.; Konur, A.; Akira, S.; Müller, W.; Kalinke, U. Excessive CpG 1668 stimulation triggers IL-10 production by cDC that inhibits IFN-α responses by pDC. Eur. J. Immunol. 2008. [Google Scholar] [CrossRef]
- Schmitt, H.; Ulmschneider, J.; Billmeier, U.; Vieth, M.; Scarozza, P.; Sonnewald, S.; Reid, S.; Atreya, I.; Rath, T.; Zundler, S.; et al. The TLR9 Agonist Cobitolimod Induces IL10-Producing Wound Healing Macrophages and Regulatory T Cells in Ulcerative Colitis. J. Crohns Colitis 2020, 14, 508–524. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, J.D.; Calbo, S.; Maho-Vaillant, M.; Saussine, A.; Bagot, M.; Bensussan, A.; Musette, P. IL-10 produced by activated human B cells regulates CD4+ T-cell activation in vitro. Eur. J. Immunol. 2010, 40, 2686–2691. [Google Scholar] [CrossRef]
- Satitsuksanoa, P.; Daanje, M.; Akdis, M.; Boyd, S.D.; van de Veen, W. Biology and dynamics of B cells in the context of IgE-mediated food allergy. Allergy Eur. J. Allergy Clin. Immunol. 2020, 76, 1707–1717. [Google Scholar] [CrossRef]
- Humeniuk, P.; Dubiela, P.; Hoffmann-Sommergruber, K. Dendritic cells and their role in allergy: Uptake, proteolytic processing and presentation of allergens. Int. J. Mol. Sci. 2017, 18, 1491. [Google Scholar] [CrossRef] [Green Version]
- van de Veen, W.; Stanic, B.; Wirz, O.F.; Jansen, K.; Globinska, A.; Akdis, M. Role of regulatory B cells in immune tolerance to allergens and beyond. J. Allergy Clin. Immunol. 2016, 138, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Kushwah, R.; Hu, J. Role of dendritic cells in the induction of regulatory T cells. Cell Biosci. 2011, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Leigh, R.; Matsumoto, K.; Wattie, J.; Ellis, R.; O’Byrne, P.M.; Inman, M.D. Effect of interferon-γ on allergic airway responses in interferon-γ-deficient mice. Am. J. Respir. Crit. Care Med. 2002, 166, 451–456. [Google Scholar] [CrossRef]
- Nakajima, H.; Iwamoto, I.; Yoshida, S. Aerosolized recombinant interferon-gamma prevents antigen-induced eosinophil recruitment in mouse trachea. Am. Rev. Respir. Dis. 1993, 148, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Lack, G.; Bradley, K.L.; Hamelmann, E.; Renz, H.; Loader, J.; Leung, D.Y.; Larsen, G.; Gelfand, E.W. Nebulized IFN-gamma inhibits the development of secondary allergic responses in mice. J. Immunol. 1996, 157, 1432–1439. [Google Scholar]
- Iwamoto, I.; Nakajima, H.; Endo, H.; Yoshida, S. Interferon γ regulates antigen-induced eosinophil recruitment into the mouse airways by inhibiting the infiltration of CD4+ T cells. J. Exp. Med. 1993, 177, 573–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koya, T.; Matsuda, H.; Matsubara, S.; Miyahara, N.; Dakhama, A.; Takeda, K.; Gelfand, E.W. Differential effects of dendritic cell transfer on airway hyperresponsiveness and inflammation. Am. J. Respir. Cell Mol. Biol. 2009, 41, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Vargas, C.M.; Ramirez-Pineda, J.R.; Palmetshofer, A.; Grunewald, S.; Moll, H.; Berberich, C.; Erb, K.J. Mice vaccinated with allergen-pulsed myeloid dendritic cells are not protected from developing allergen-induced Th2 responses. Int. Arch. Allergy Immunol. 2005, 137, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wu, N.; Min, Z.; Li, Z.; Zhu, T.; Liu, C.; Zeng, Y.; Song, J.; Mao, R.; Ji, H.; et al. Adoptive transfer of bone marrow-derived dendritic cells (BMDCs) alleviates OVA-induced allergic airway inflammation in asthmatic mice. Sci. Rep. 2020, 10, 13915. [Google Scholar] [CrossRef]
- Min, Z.; Zeng, Y.; Zhu, T.; Cui, B.; Mao, R.; Jin, M.; Chen, Z. Lipopolysaccharide-Activated Bone Marrow-Derived Dendritic Cells Suppress Allergic Airway Inflammation by Ameliorating the Immune Microenvironment. Front. Immunol. 2021, 12, 1. [Google Scholar] [CrossRef]
- Bagaev, A.; Pichugin, A.; Nelson, E.L.; Agadjanyan, M.G.; Ghochikyan, A.; Ataullakhanov, R.I. Anticancer Mechanisms in Two Murine Bone Marrow–Derived Dendritic Cell Subsets Activated with TLR4 Agonists. J. Immunol. 2018, 200, 2656–2669. [Google Scholar] [CrossRef]
- Ren, S.; Wang, Q.; Zhang, Y.; Song, Y.; Dong, X.; Zhang, W.; Qin, X.; Liu, M.; Yu, T. Imiquimod enhances the potency of an exogenous BM-DC based vaccine against mouse melanoma. Int. Immunopharmacol. 2018, 64, 69–77. [Google Scholar] [CrossRef]
- Matisz, C.E.; Faz-López, B.; Thomson, E.; Al Rajabi, A.; Lopes, F.; Terrazas, L.I.; Wang, A.; Sharkey, K.A.; McKay, D.M. Suppression of colitis by adoptive transfer of helminth antigentreated dendritic cells requires interleukin-4 receptor-α signaling. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaide, E.G.; Krishnarajah, S.; Junker, F. Dendritic Cell Tumor Vaccination via Fc Gamma Receptor Targeting: Lessons Learned from Pre-Clinical and Translational Studies. Vaccines 2021, 9, 409. [Google Scholar] [CrossRef]
- Rehli, M. Of mice and men: Species variations of Toll-like receptor expression. Trends Immunol. 2002, 23, 375–378. [Google Scholar] [CrossRef]
- Hornung, V.; Rothenfusser, S.; Britsch, S.; Krug, A.; Jahrsdörfer, B.; Giese, T.; Endres, S.; Hartmann, G. Quantitative Expression of Toll-Like Receptor 1–10 mRNA in Cellular Subsets of Human Peripheral Blood Mononuclear Cells and Sensitivity to CpG Oligodeoxynucleotides. J. Immunol. 2002, 168, 4531–4537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoene, V.; Peiser, M.; Wanner, R. Human monocyte-derived dendritic cells express TLR9 and react directly to the CpG-A oligonucleotide D19. J. Leukoc. Biol. 2006, 80, 1328–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.; Peng, P.; Loschko, J.; Feng, L.; Pham, P.; Cui, W.; Lee, K.P.; Krug, A.B.; Jiang, A. Plasmacytoid dendritic cells cross-prime naive CD8 T cells by transferring antigen to conventional dendritic cells through exosomes. Proc. Natl. Acad. Sci. USA 2020, 117, 23730–23741. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberca, R.W.; Gomes, E.; Russo, M. CpG-ODN Signaling via Dendritic Cells-Expressing MyD88, but Not IL-10, Inhibits Allergic Sensitization. Vaccines 2021, 9, 743. https://doi.org/10.3390/vaccines9070743
Alberca RW, Gomes E, Russo M. CpG-ODN Signaling via Dendritic Cells-Expressing MyD88, but Not IL-10, Inhibits Allergic Sensitization. Vaccines. 2021; 9(7):743. https://doi.org/10.3390/vaccines9070743
Chicago/Turabian StyleAlberca, Ricardo Wesley, Eliane Gomes, and Momtchilo Russo. 2021. "CpG-ODN Signaling via Dendritic Cells-Expressing MyD88, but Not IL-10, Inhibits Allergic Sensitization" Vaccines 9, no. 7: 743. https://doi.org/10.3390/vaccines9070743
APA StyleAlberca, R. W., Gomes, E., & Russo, M. (2021). CpG-ODN Signaling via Dendritic Cells-Expressing MyD88, but Not IL-10, Inhibits Allergic Sensitization. Vaccines, 9(7), 743. https://doi.org/10.3390/vaccines9070743