
Purification of Adeno-Associated Virus (AAV) Serotype 2 from Spodoptera frugiperda (Sf9) Lysate by Chromatographic Nonwoven Membranes

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Experimental Procedure
2.1. Materials
2.2. Preparation of Ion-Exchange Nonwovens
2.3. Dynamic AAV2 Binding Capacity of AEX-TEA Membranes in Non-Competitive Conditions (Pure AAV2)
2.4. AAV2 Purification from Sf9 Cell Lysate with AEX-TEA Membranes
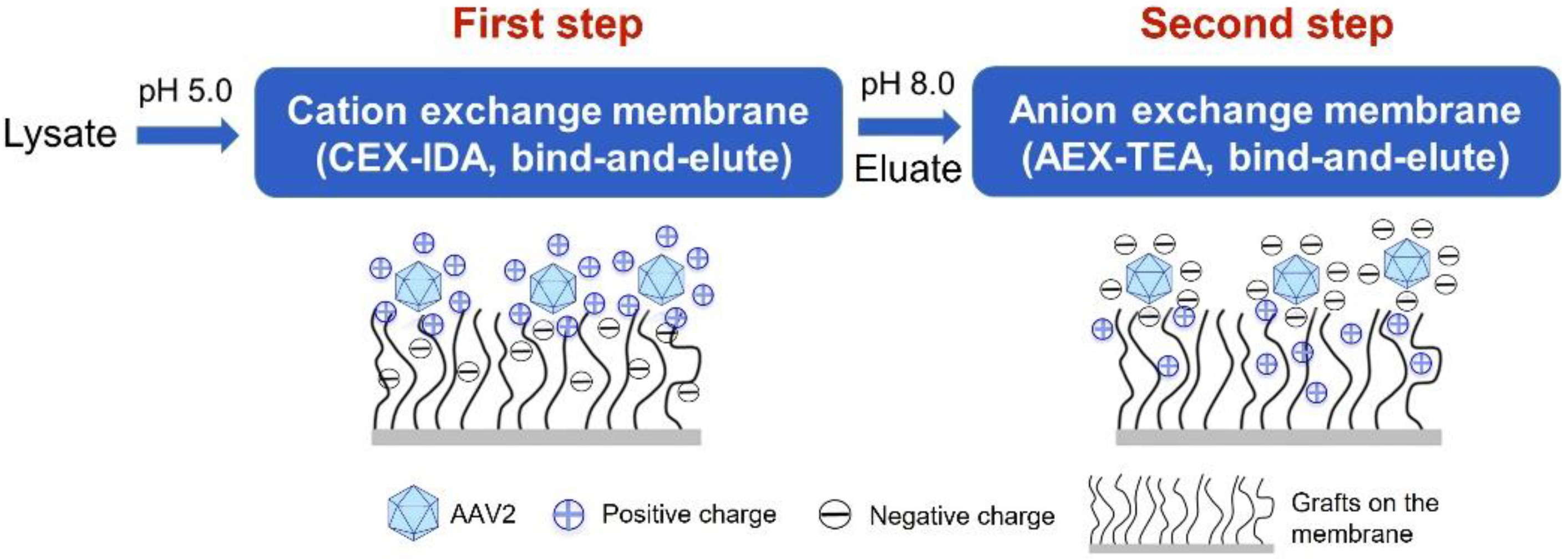
2.5. Purification of AAV2 from Cell Lysate by a Two-Step Membrane Chromatography Process
2.6. Sample Analysis
2.6.1. Total Protein Concentration, AAV2 Capsid (Full and Empty) Concentration, and dsDNA Concentration Measurement
2.6.2. Viral Genome (VG) Quantification by qPCR
2.6.3. Transduction Activity Assay
2.6.4. Capsid Imaging via Transmission Electron Microscopy (TEM)
3. Results and Discussion
3.1. Preparation of AEX-TEA Membrane
3.2. Pure AAV2 Bind-and-Elute Experiment with AEX-TEA Membrane
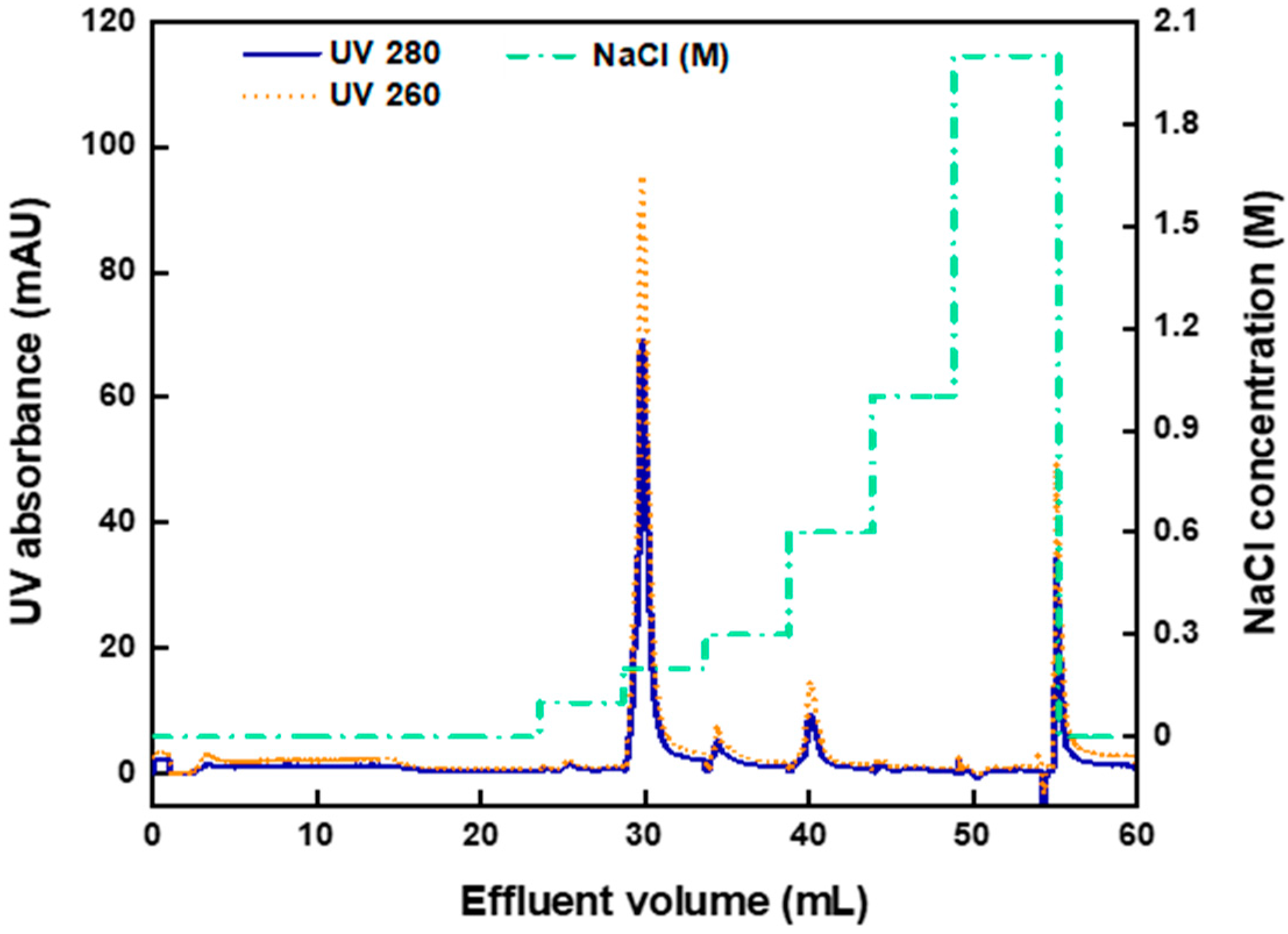
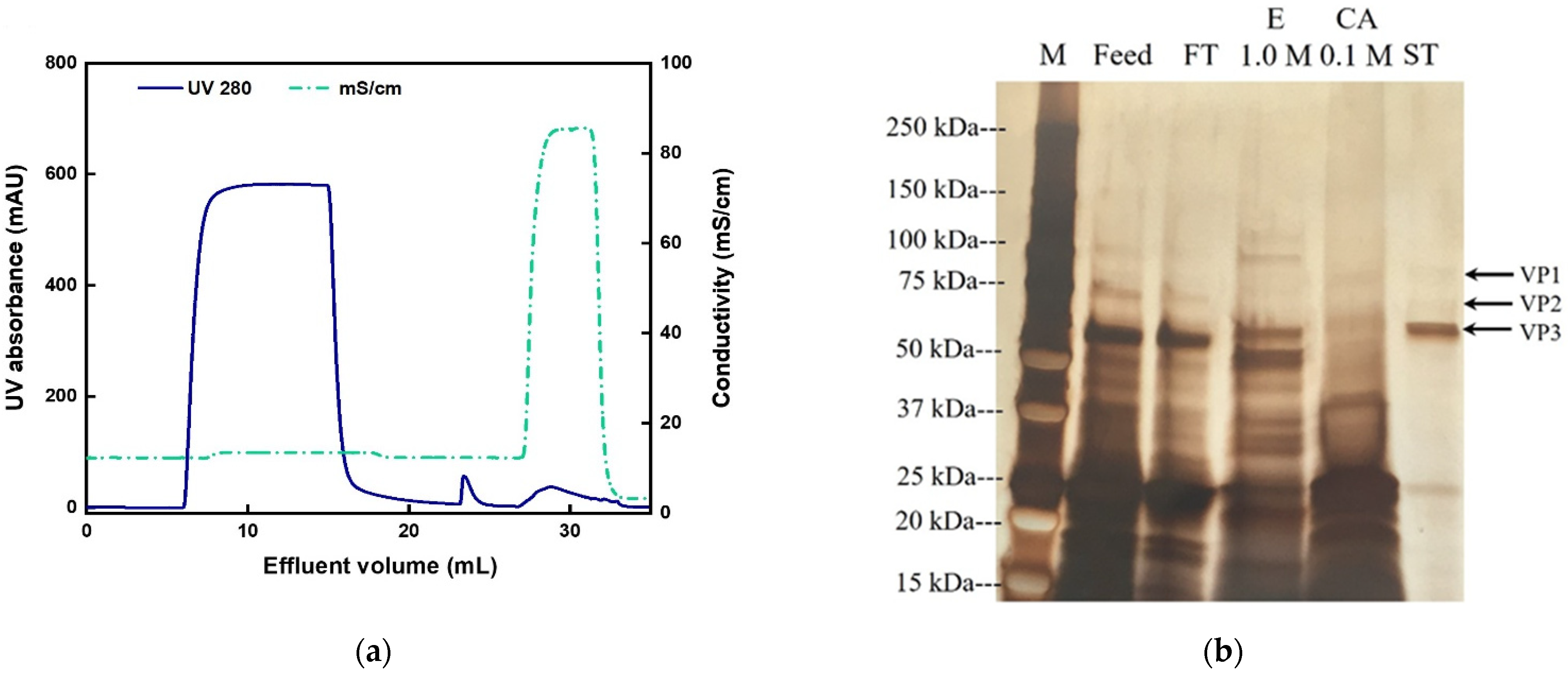
3.3. AAV2 Separation from Cell Lysate with Anion-Exchange Membranes
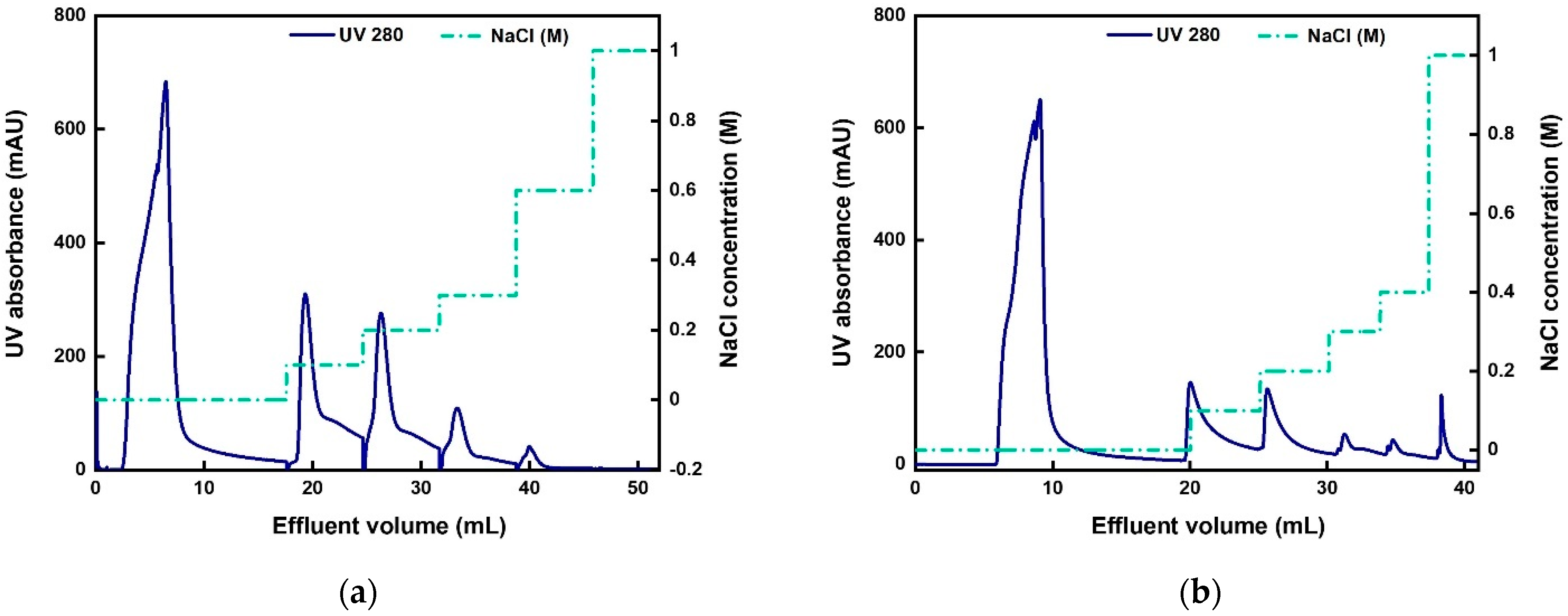
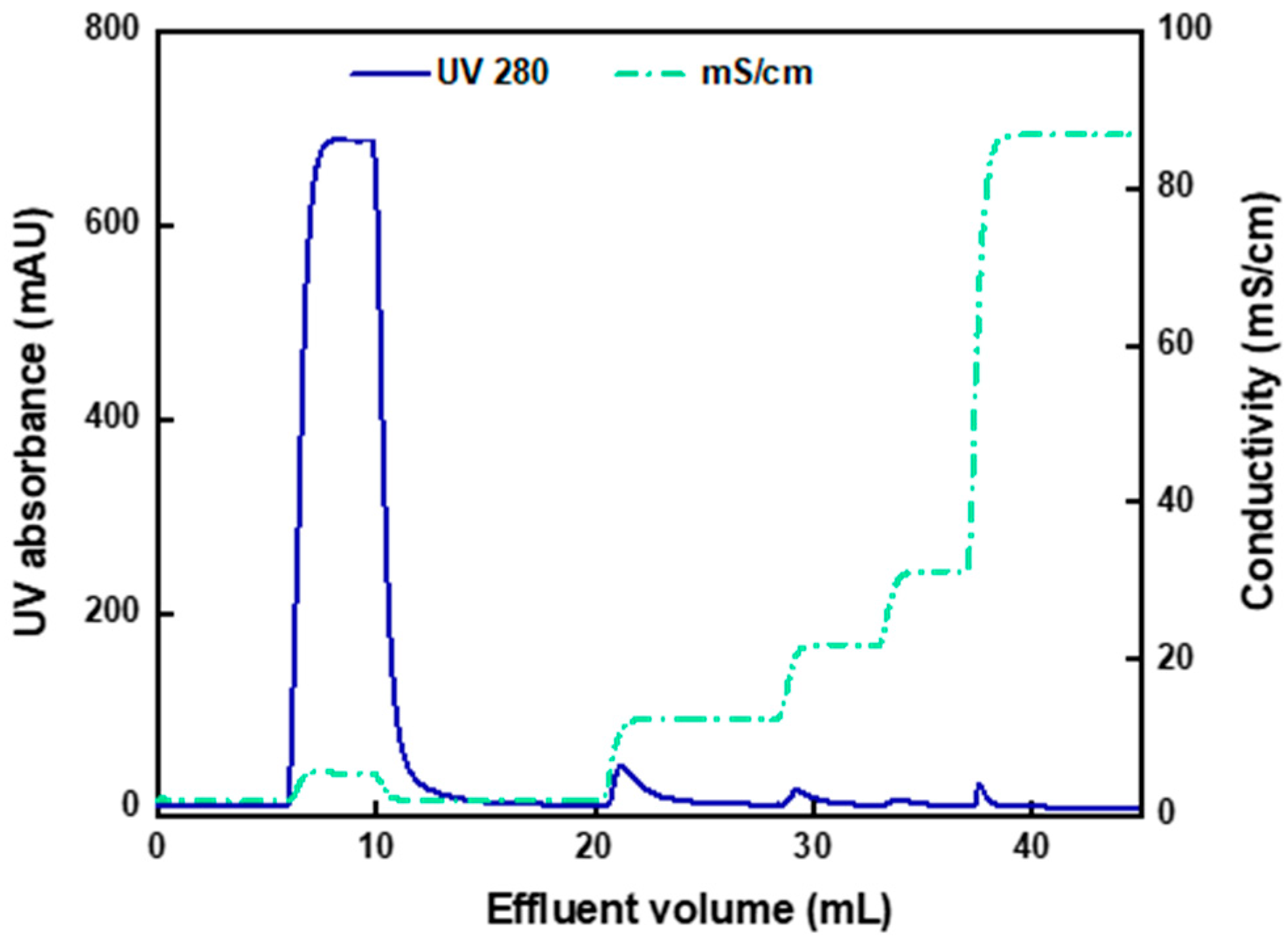
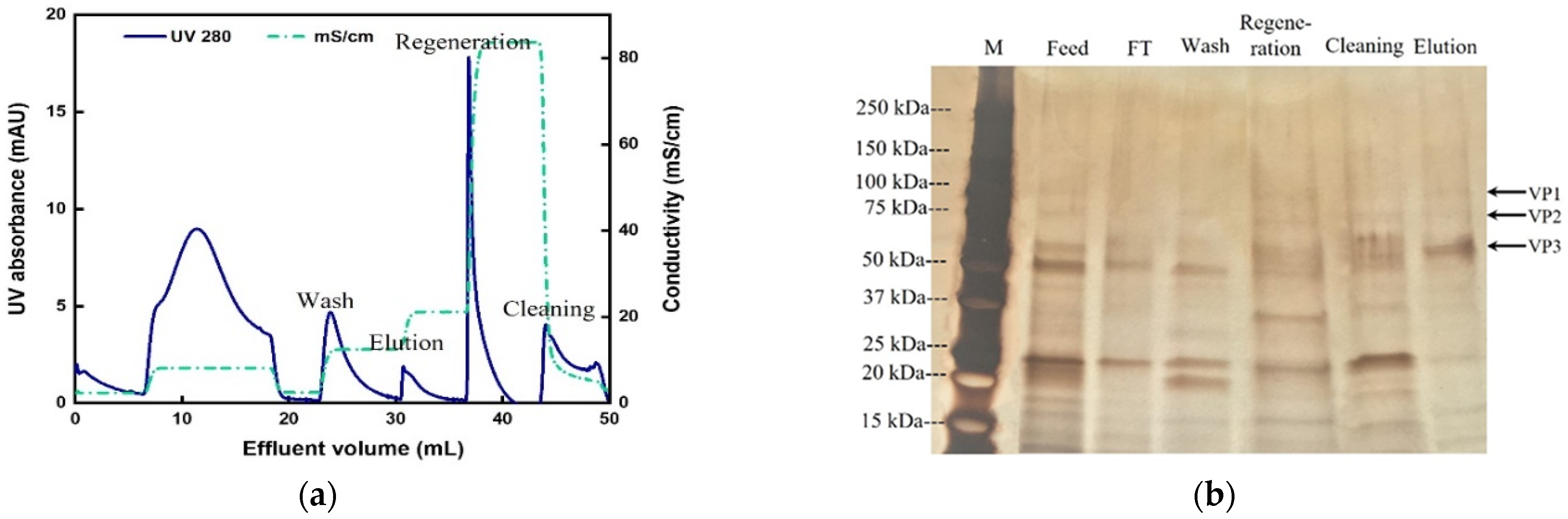
3.4. Two-Step Purification of AAV2 by Membrane Chromatography
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perrin, G.Q.; Herzog, R.W.; Markusic, D.M. Update on clinical gene therapy for hemophilia. Blood 2019, 133, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Nagasato, M.; Rin, Y.; Henmi, M.; Ino, Y.; Yachida, S.; Ohki, R.; Hiraoka, N.; Tagawa, M.; Aoki, K. Strong antitumor efficacy of a pancreatic tumor-targeting oncolytic adenovirus for neuroendocrine tumors. Cancer Med. 2017, 6, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.B.; Tan, X.Y.; Tampe, B.; Wilhelmi, T.; Hulshoff, M.S.; Saito, S.; Moser, T.; Kalluri, R.; Hasenfuss, G.; Zeisberg, E.M.; et al. High-fidelity CRISPR/Cas9-based gene-specific hydroxymethylation rescues gene expression and attenuates renal fibrosis. Nat. Commun. 2018, 9, 3509. [Google Scholar] [CrossRef]
- Kanao-Kanda, M.; Kanda, H.; Liu, S.; Roy, S.; Toborek, M.; Hao, S. Viral vector–mediated gene transfer of glutamic acid decarboxylase for chronic pain treatment: A literature review. Hum. Gene Ther. 2020, 31, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.C.; Wang, Z.L.; Xu, T.; He, Z.Y.; Wei, Y.Q. The approved gene therapy drugs worldwide: From 1998 to 2019. Biotechnol. Adv. 2020, 40, 107502. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.; Bak, H.; Tustian, A.D. Moving from the bench towards a large scale, industrial platform process for adeno-associated viral vector purification. Biotechnol. Bioeng. 2020, 117, 3199–3211. [Google Scholar] [CrossRef] [PubMed]
- Bulcha, J.T.; Wang, Y.; Ma, H.; Tai, P.W.L.; Gao, G.P. Viral vector platforms within the gene therapy landscape. Signal Transduct. Target. Ther. 2021, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- El Andari, J.; Grimm, D. Production, Processing, and Characterization of synthetic AAV Gene Therapy Vectors. Biotechnol. J. 2021, 16, 2000025. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Mallela, K.M.G.; Deorkar, N.; Brophy, G. Manufacturing challenges and rational formulation development for AAV viral vectors. J. Pharm. Sci. 2021, 110, 2609–2624. [Google Scholar] [CrossRef]
- Clement, N.; Grieger, J.C. Manufacturing of recombinant adeno-associated viral vectors for clinical trials. Mol. Ther.-Methods Clin. Dev. 2016, 3, 16002. [Google Scholar] [CrossRef] [Green Version]
- Nass, S.A.; Mattingly, M.A.; Woodcock, D.A.; Burnham, B.L.; Ardinger, J.A.; Osmond, S.E.; Frederick, A.M.; Scaria, A.; Cheng, S.H.; O’Riordan, C.R. Universal method for the purification of recombinant AAV vectors of differing serotypes. Mol. Ther.-Methods Clin. Dev. 2018, 9, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.H.; Su, C.; Wang, X.L.; Shu, Y.H.; Hu, S.C.; Zhao, C.Z.; Kuang, Y.T.; Chen, Y.W.; Li, Y.H.; Wei, Y.Q.; et al. A novel method to purify adenovirus based on increasing salt concentrations in buffer. Eur. J. Pharm. Sci. 2020, 141, 105090. [Google Scholar] [CrossRef] [PubMed]
- McNally, D.J.; Piras, B.A.; Willis, C.M.; Lockey, T.D.; Meagher, M.M. Development and optimization of a hydrophobic interaction chromatography-based method of AAV harvest, capture, and recovery. Mol. Ther.-Methods Clin. Dev. 2020, 19, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Nestola, P.; Peixoto, C.; Silva, R.; Alves, P.M.; Mota, J.P.B.; Carrondo, M.J.T. Improved virus purification processes for vaccines and gene therapy. Biotechnol. Bioeng. 2015, 112, 843–857. [Google Scholar] [CrossRef]
- Nadar, S.; Shooter, G.; Somasundaram, B.; Shave, E.; Baker, K.; Lua, L.H.L. Intensified downstream processing of monoclonal antibodies using membrane technology. Biotechnol. J. 2021, 16, 2000309. [Google Scholar] [CrossRef]
- Potter, M.; Lins, B.; Mietzsch, M.; Heilbronn, R.; van Vliet, K.; Chipman, P.; Agbandje-McKenna, M.; Cleaver, B.D.; Clement, N.; Byrne, B.J.; et al. A simplified purification protocol for recombinant adeno-associated virus vectors. Mol. Ther.-Methods Clin. Dev. 2014, 1, 14034. [Google Scholar] [CrossRef]
- Rieser, R.; Koch, J.; Faccioli, G.; Richter, K.; Menzen, T.; Biel, M.; Winter, G.; Michalakis, S. Comparison of different liquid chromatography-based purification strategies for adeno-associated virus vectors. Pharmaceutics 2021, 13, 748. [Google Scholar] [CrossRef]
- Dickerson, R.; Argento, C.; Pieracci, J.; Bakhshayeshi, M. Separating empty and full recombinant adeno-associated virus particles using isocratic anion exchange chromatography. Biotechnol. J. 2021, 16, 2000015. [Google Scholar] [CrossRef]
- Zydney, A.L. New developments in membranes for bioprocessing—A review. J. Membr. Sci. 2021, 620, 118804. [Google Scholar] [CrossRef]
- Kawka, K.; Madadkar, P.; Umatheva, U.; Shoaebargh, S.; Medina, M.F.C.; Lichty, B.D.; Ghosh, R.; Latulippe, D.R. Purification of therapeutic adenoviruses using laterally-fed membrane chromatography. J. Membr. Sci. 2019, 579, 351–358. [Google Scholar] [CrossRef]
- Vicente, T.; Fabar, R.; Alves, P.M.; Carrondo, M.J.T.; Mota, J.P.B. Impact of ligand density on the optimization of ion-exchange membrane chromatography for viral vector purification. Biotechnol. Bioeng. 2011, 108, 1347–1359. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, U. Bioseparation in antibody manufacturing: The good, the bad and the ugly. Biotechnol. Progr. 2008, 24, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, Q.; Shi, C.; Gou, J.; Gao, D.; Wang, H.; Yao, S.; Lin, D. Antibody capture with twin-column continuous chromatography: Effects of residence time, protein concentration and resin. Sep. Purif. Technol. 2020, 253, 117554. [Google Scholar] [CrossRef]
- Nestola, P.; Villain, L.; Peixoto, C.; Martins, D.L.; Alves, P.M.; Carrondo, M.J.T.; Mota, J.P.B. Impact of grafting on the design of new membrane adsorbents for adenovirus purification. J. Biotechnol. 2014, 181, 1–11. [Google Scholar] [CrossRef]
- Tran, R.; Joseph, J.R.; Sinclair, A.; Bracewell, D.; Zhou, Y.; Titchener-Hooker, N.J. A framework for the prediction of scale-up when using compressible chromatographic packings. Biotechnol. Progr. 2007, 23, 413–422. [Google Scholar]
- Vincent, D.; Kramberger, P.; Hudej, R.; Strancar, A.; Wang, Y.H.; Zhou, Y.H.; Velayudhan, A. The development of a monolith-based purification process for orthopoxvirus vaccinia virus lister strain. J. Chromatogr. A 2017, 1524, 87–100. [Google Scholar] [CrossRef]
- Grein, T.A.; Michalsky, R.; Lopez, M.V.; Czermak, P. Purification of a recombinant baculovirus of Autographa californica M nucleopolyhedrovirus by ion exchange membrane chromatography. J. Virol. Methods 2012, 183, 117–124. [Google Scholar] [CrossRef]
- Turnbull, J.; Wright, B.; Green, N.K.; Tarrant, R.; Roberts, I.; Hardick, O.; Bracewell, D.G. Adenovirus 5 recovery using nanofiber ion-exchange adsorbents. Biotechnol. Bioeng. 2019, 116, 1698–1709. [Google Scholar] [CrossRef]
- Junter, G.A.; Lebrun, L. Polysaccharide-based chromatographic adsorbents for virus purification and viral clearance. J. Pharm. Anal. 2020, 10, 291–312. [Google Scholar] [CrossRef]
- Boi, C.; Malavasi, A.; Carbonell, R.G.; Gilleskie, G. A direct comparison between membrane adsorber and packed column chromatography performance. J. Chromatogr. A 2020, 1612, 460629. [Google Scholar] [CrossRef]
- McNally, D.J.; Darling, D.; Farzaneh, F.; Levison, P.R.; Slater, N.K.H. Optimised concentration and purification of retroviruses using membrane chromatography. J. Chromatogr. A 2014, 1340, 24–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winderl, J.; Neumann, E.; Hubbuch, J. Exploration of fiber-based cation exchange adsorbents for the removal of monoclonal antibody aggregates. J. Chromatogr. A 2021, 1654, 462451. [Google Scholar] [CrossRef] [PubMed]
- Bhut, B.V.; Wickramasinghe, S.R.; Husson, S.M. Preparation of high-capacity, weak anion-exchange membranes for protein separations using surface-initiated atom transfer radical polymerization. J. Membr. Sci. 2008, 325, 176–183. [Google Scholar] [CrossRef]
- Ruscic, J.; Perry, C.; Mukhopadhyay, T.; Takeuchi, Y.; Bracewell, D.G. Lentiviral vector purification using nanofiber ion-exchange chromatography. Mol. Ther. Methods Clin. Dev. 2019, 15, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, S.; Crandall, C.; Schneiderman, S.; Menkhaus, T.J. Cellulose-graft-polyethyleneamidoamine Anion-Exchange Nanofiber Membranes for Simultaneous Protein Adsorption and Virus Filtration. ACS Appl. Nano. Mater. 2018, 1, 3321–3330. [Google Scholar] [CrossRef]
- Heller, M.; Li, Q.; Esinhart, K.; Pourdeyhimi, B.; Boi, C.; Carbonell, R.G. Heat induced grafting of poly(glycidyl methacrylate) on polybutylene terephthalate nonwovens for bioseparations. Ind. Eng. Chem. Res. 2020, 59, 5371–5380. [Google Scholar] [CrossRef]
- Lemma, S.M.; Boi, C.; Carbonell, R.G. Nonwoven ion-exchange membranes with high protein binding capacity for bioseparations. Membranes 2021, 11, 181. [Google Scholar] [CrossRef]
- Fan, J.X.; Boi, C.; Lemma, S.M.; Lavoie, J.; Carbonell, R.G. Iminodiacetic acid (IDA) cation-exchange nonwoven membranes for efficient capture of antibodies and antibody fragments. Membranes 2021, 11, 530. [Google Scholar] [CrossRef]
- Venkatakrishnan, B.; Yarbrough, J.; Domsic, J.; Bennett, A.; Bothner, B.; Kozyreva, O.G.; Samulski, R.J.; Muzyczka, N.; McKenna, R.; Agbandje-McKenna, M. Structure and dynamics of adeno-associated virus serotype 1 VP1-unique N-terminal domain and its role in capsid trafficking. J. Virol. 2013, 87, 4974–4984. [Google Scholar] [CrossRef]
- Santos, N.V.d.; Martins, M.; Santos-Ebinuma, V.C.; Ventura, S.P.M.; Coutinho, J.A.P.; Valentini, S.R.; Pereira, J.F.B. Aqueous biphasic systems composed of cholinium chloride and polymers as effective platforms for the purification of recombinant green fluorescent protein. ACS Sustain. Chem. Eng. 2018, 6, 9383–9393. [Google Scholar] [CrossRef]
- Tomono, T.; Hirai, Y.; Okada, H.; Miyagawa, Y.; Adachi, K.; Sakamoto, S.; Kawano, Y.; Chono, H.; Mineno, J.; Ishii, A.; et al. Highly efficient ultracentrifugation-free chromatographic purification of recombinant AAV serotype 9. Mol. Ther.-Methods Clin. Dev. 2018, 11, 180–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marichal-Gallardo, P.; Borner, K.; Pieler, M.M.; Sonntag-Buck, V.; Obr, M.; Bejarano, D.; Wolff, M.W.; Krausslich, H.G.; Reichl, U.; Grimm, D. Single-use capture purification of adeno-associated viral gene transfer vectors by membrane-based steric exclusion chromatography. Hum. Gene Ther. 2019, 32, 959–974. [Google Scholar] [CrossRef] [PubMed]
- Rakotondravao, H.M.; Ishizuka, N.; Sakakibara, K.; Wada, R.; Ichihashi, E.; Takahashi, R.; Takai, T.; Horiuchi, J.I.; Kumada, Y. Characterization of a macroporous epoxy-polymer based resin for the ion-exchange chromatography of therapeutic proteins. J. Chromatogr. A 2021, 1656, 462503. [Google Scholar] [CrossRef] [PubMed]
- Brument, N.; Morenweiser, R.; Blouin, V.; Toublanc, E.; Raimbaud, I.; Cherel, Y.; Folliot, S.; Gaden, F.; Boulanger, P.; Kroner-Lux, G.; et al. A versatile and scalable two-step ion-exchange chromatography process for the purification of recombinant adeno-associated virus serotypes-2 and-5. Mol. Ther. 2002, 6, 678–686. [Google Scholar] [CrossRef]
- Qu, G.; Bahr-Davidson, J.; Prado, J.; Tai, A.; Cataniag, F.; McDonnell, J.; Zhou, J.M.; Hauck, B.; Luna, J.; Sommer, J.M.; et al. Separation of adeno-associated virus type 2 empty particles from genome containing vectors by anion-exchange column chromatography. J. Virol. Methods 2007, 140, 183–192. [Google Scholar] [CrossRef]
- Lock, M.; Alvira, M.R.; Wilson, J.M. Analysis of particle content of recombinant adeno-associated virus serotype 8 vectors by ion-exchange chromatography. Hum. Gene Ther. Methods 2012, 23, 56–64. [Google Scholar] [CrossRef]
- Davidoff, A.M.; Ng, C.Y.C.; Sleep, S.; Gray, J.; Azam, S.; Zhao, Y.; McIntosh, J.H.; Karimipoor, M.; Nathwani, A.C. Purification of recombinant adeno-associated virus type 8 vectors by ion exchange chromatography generates clinical grade vector stock. J. Virol. Methods 2004, 121, 209–215. [Google Scholar] [CrossRef]
- Trilisky, E.I.; Lenhoff, A.M. Flow-dependent entrapment of large bioparticles in porous process media. Biotechnol. Bioeng. 2009, 104, 127–133. [Google Scholar] [CrossRef]
- Hardick, O.; Dods, S.; Stevens, B.; Bracewell, D.G. Nanofiber adsorbents for high productivity downstream processing. Biotechnol. Bioeng. 2013, 110, 1119–1128. [Google Scholar] [CrossRef]
- Heller, M.; Wimbish, R.; Gurgel, P.V.; Pourdeyhimi, B.; Carbonell, R.G. Reducing diffusion limitations in Ion exchange grafted membranes using high surface area nonwovens. J. Membr. Sci. 2016, 514, 53–64. [Google Scholar] [CrossRef]
- Tatarova, I.; Faber, R.; Denoyel, R.; Polakovic, M. Characterization of pore structure of a strong anion-exchange membrane adsorbent under different buffer and salt concentration conditions. J. Chromatogr. A 2009, 1216, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Mourão, C.A.; Marcuz, C.; Haupt, K.; Bueno, S.M.A. Polyacrylamide-alginate (PAAm-Alg) and phospho-L-tyrosine-linked PAAm-Alg monolithic cryogels: Purification of IgG from human serum. J. Chromatogr. B 2019, 1129, 121783. [Google Scholar] [CrossRef] [PubMed]
- Gardea-Torresdey, J.L.; Tiemann, K.J.; Armendariz, V.; Bess-Oberto, L.; Chianelli, R.R.; Rios, J.; Parsons, J.G.; Gamez, G. Characterization of Cr (VI) binding and reduction to Cr (III) by the agricultural byproducts of Avena monida (Oat) biomass. J. Hazard. Mater. A 2000, 80, 175–188. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AAV2 Sources | AAV2 from Clarified Sf9 Cell Lysate | Pure AAV2 with Full Capsids (Standard) |
|---|---|---|
| Capsids concentration (capsids/mL) 1 | 1.1 × 1013 | 1 × 1012 |
| VG/Capsids 2 | 0.19 | >0.9 |
| TU/mL 3 | 1.7 × 108 | 2.2 × 107 |
| Protein concentration 4 | ~10 mg/mL | 2.2 μg/mL |
| Membrane | AEX-TEA | Sartobind Q |
|---|---|---|
| Membrane type | Nonwoven | Microporous cast |
| Membrane volume (mL) | 0.1 | 0.08 |
| Specific surface area a (m2/g) | 0.86 b [50] | 0.90 [51] |
| Ligand density c (μmol/g) | 600.0 | 553.6 |
| Sample loading (capsids/mL of membrane volume) | 2.5 × 1013 | 2.5 × 1013 |
| % Capsids in FT d | 1.3% | 25.8% |
| % Capsids in the 0.1 and 0.2 M NaCl elution fraction | 87.9% | 59.0% |
| Number of bound capsids/mL of membrane e | 2.4 × 1013 | 1.9 × 1013 |
| Eluted capsid concentration (capsids/mL) f | 3.3 × 1011 | 1.9 × 1011 |
| VG/Capsids g | 0.2 | 0.4 |
| Non-capsid protein concentration (ng/mL) h | 4.4 × 103 (1.8 LRV) | 5.0 × 103 (1.5 LRV) |
| CEX-IDA Membrane (First Step) | AEX-TEA Membrane (Second Step) | |
|---|---|---|
| % Recovery | 1st Step capsid recovery: 87.3% | 2nd Step capsid recovery: 74.8% |
| 1st Step infective recovery: 88.1% | 2nd Step infective recovery: 86.3% | |
| 2-Step Combined Process | capsid recovery: 65.3%, infective recovery:76.0% | |
| Capsids concentration (capsids/mL) | 1.1 × 1011 | 2.4 × 1011 |
| VG/Capsids | 0.26 | 0.42 |
| Non-capsid protein concentration (ng/mL) | 1.4 × 104 (1.1 LRV) | 78.1 (LRV: 2.2) |
| Contaminant dsDNA (ng/mL) | 188.6 (1.5 LRV) | 0.4 (LRV: 2.7) |
| CEX-IDA Membrane (First Step) | AEX-TEA Membrane (Second Step) | |
|---|---|---|
| % Recovery | 1st Step capsid recovery: 90.6% | 2nd Step capsid recovery: 78.6% |
| 2-Step Combined Process | capsid recovery: 71.2% | |
| Capsids concentration (capsids/mL) | 1.2 × 1011 | 2.5 × 1011 |
| VG/Capsids | N/A | N/A |
| Non-capsid protein concentration (ng/mL) | 2.5 × 104 (0.8 LRV) | 636.4 (LRV: 1.6) |
| Contaminant dsDNA (ng/mL) | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Barbieri, E.; Shastry, S.; Menegatti, S.; Boi, C.; Carbonell, R.G. Purification of Adeno-Associated Virus (AAV) Serotype 2 from Spodoptera frugiperda (Sf9) Lysate by Chromatographic Nonwoven Membranes. Membranes 2022, 12, 944. https://doi.org/10.3390/membranes12100944
Fan J, Barbieri E, Shastry S, Menegatti S, Boi C, Carbonell RG. Purification of Adeno-Associated Virus (AAV) Serotype 2 from Spodoptera frugiperda (Sf9) Lysate by Chromatographic Nonwoven Membranes. Membranes. 2022; 12(10):944. https://doi.org/10.3390/membranes12100944
Chicago/Turabian StyleFan, Jinxin, Eduardo Barbieri, Shriarjun Shastry, Stefano Menegatti, Cristiana Boi, and Ruben G. Carbonell. 2022. "Purification of Adeno-Associated Virus (AAV) Serotype 2 from Spodoptera frugiperda (Sf9) Lysate by Chromatographic Nonwoven Membranes" Membranes 12, no. 10: 944. https://doi.org/10.3390/membranes12100944
APA StyleFan, J., Barbieri, E., Shastry, S., Menegatti, S., Boi, C., & Carbonell, R. G. (2022). Purification of Adeno-Associated Virus (AAV) Serotype 2 from Spodoptera frugiperda (Sf9) Lysate by Chromatographic Nonwoven Membranes. Membranes, 12(10), 944. https://doi.org/10.3390/membranes12100944