


Effect of Cytochrome C on the Conductance of Asolectin Membranes and the Occurrence of Through Pores at Different pHs
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lipid, Cytochrome C, and Electrolytes
2.2. Planar Lipid Bilayer Membranes
2.3. Electrical Measurements
2.4. Measurement of Conductance and Estimation of the Radius of Pores in the Membrane
2.5. Asymmetric Addition of Cyt C and Measurement of the Difference in Surface Potential
3. Results
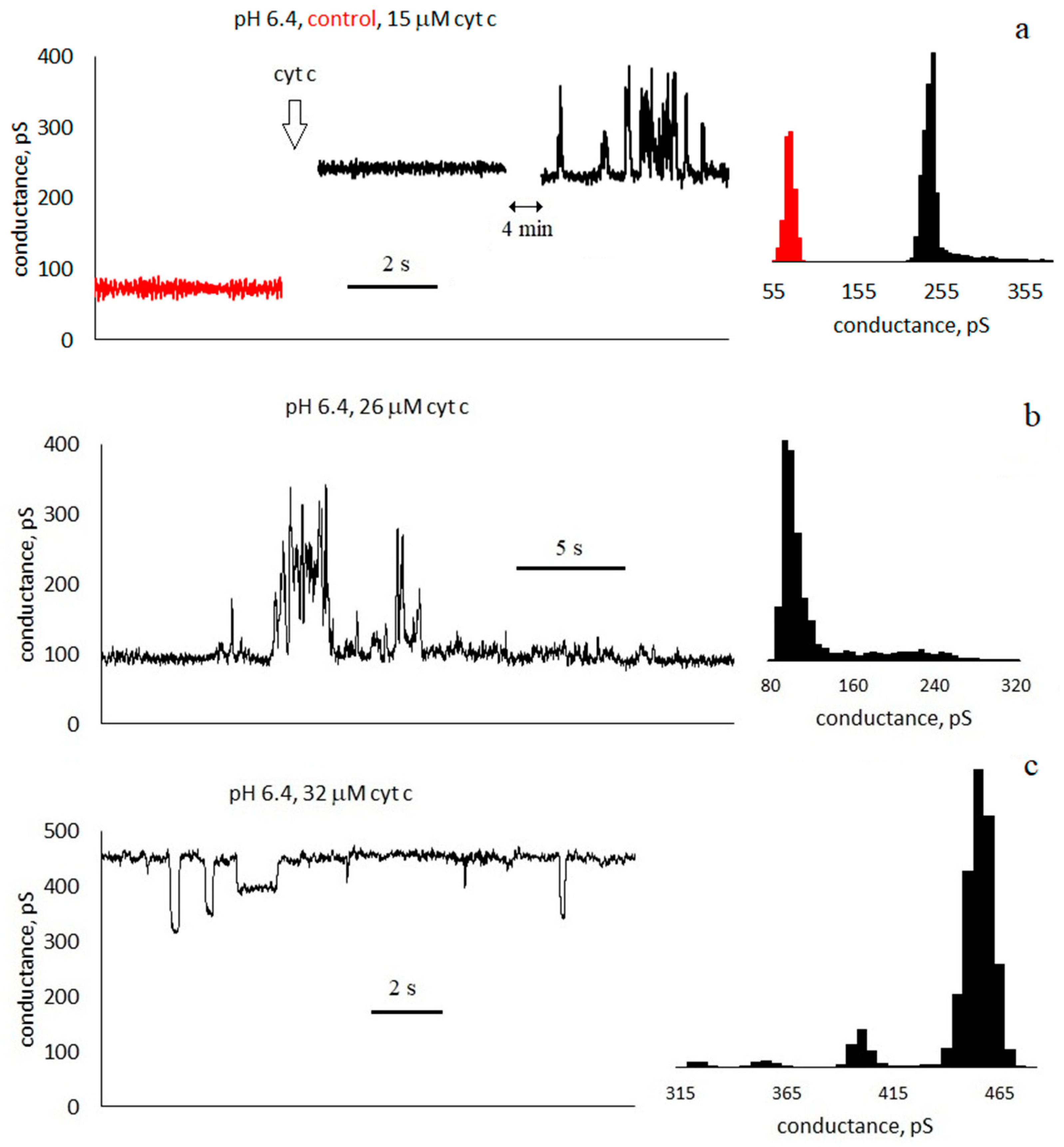
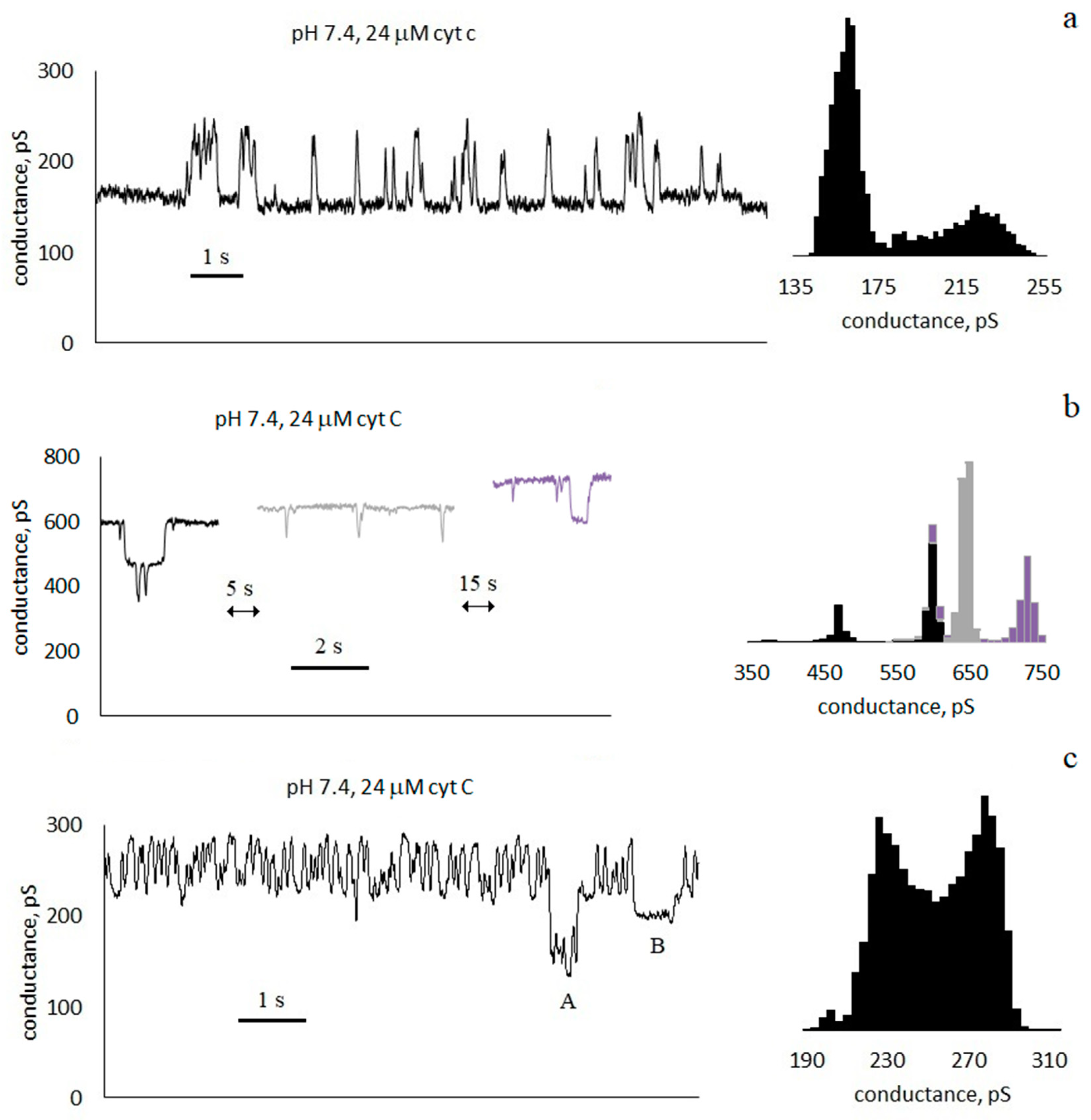
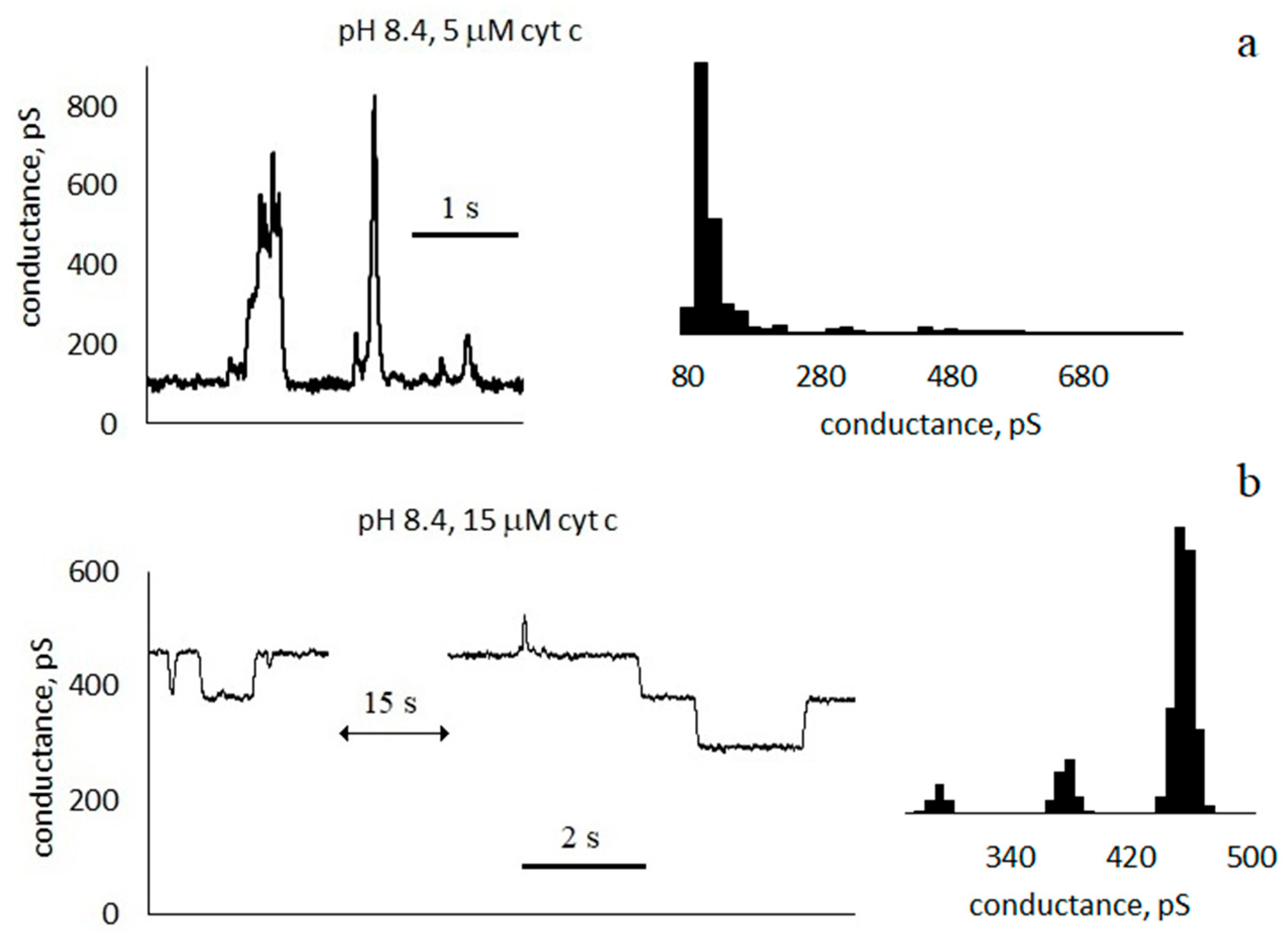
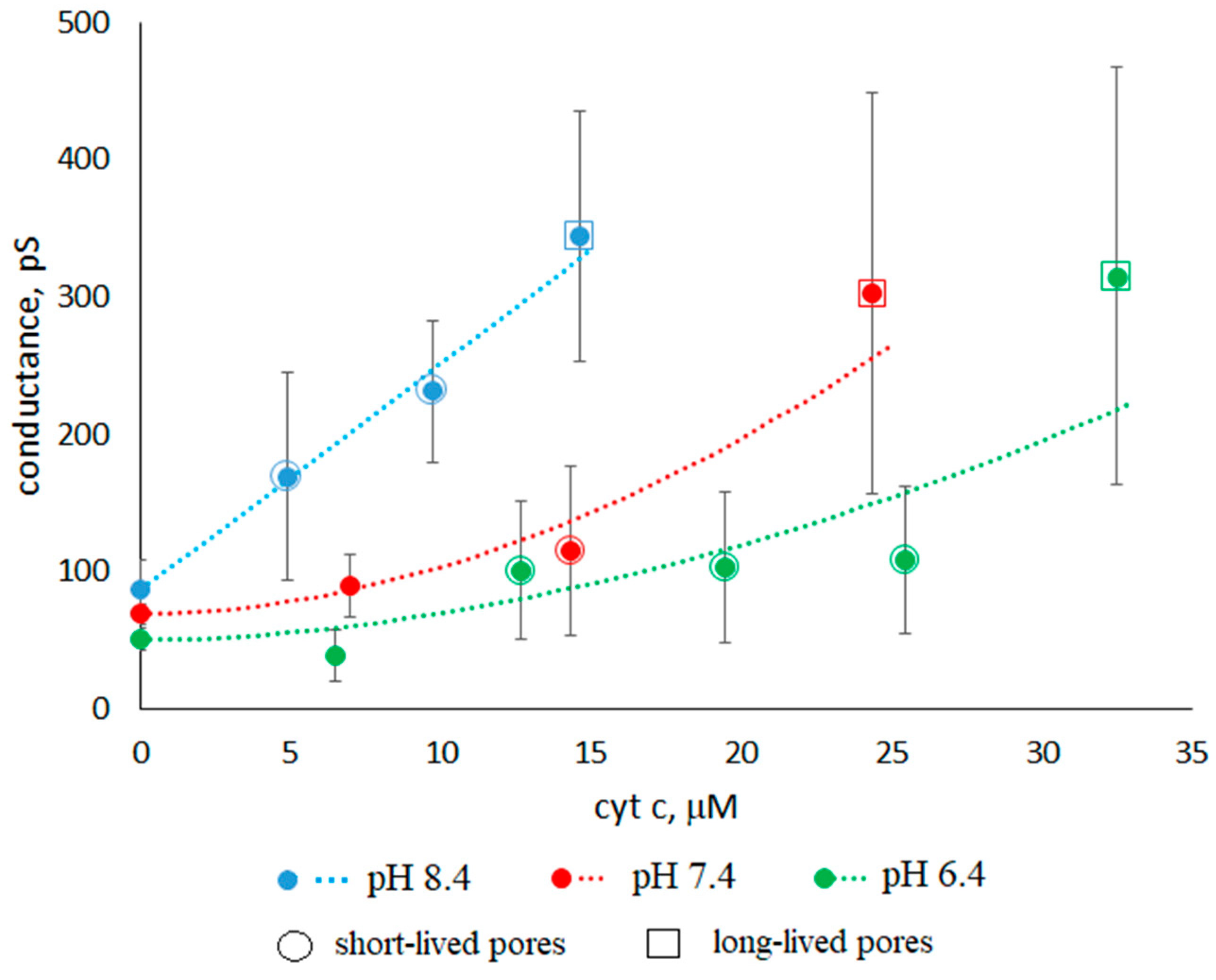
3.1. Conductance of Asolectin Membranes
3.2. Characteristics of Through Pores
- Pulses with a duration much less than the interpulse interval . For the trace in Figure 3a, the average pulse duration is τ = 100 ± 90 ms, and the frequency is f ≈ 2 Hz).
- Pulses with a duration approximately equal to half of the interpulse interval. For the trace in Figure 3c, the average pulse duration is τ = 60 ± 40 ms, and the frequency is f ≈ 8 Hz. In this case, in the conductance histogram there are two peaks of approximately the same size. The distributions of the durations of the opened and closed states are presented in the Supplementary Materials (Figure S2).
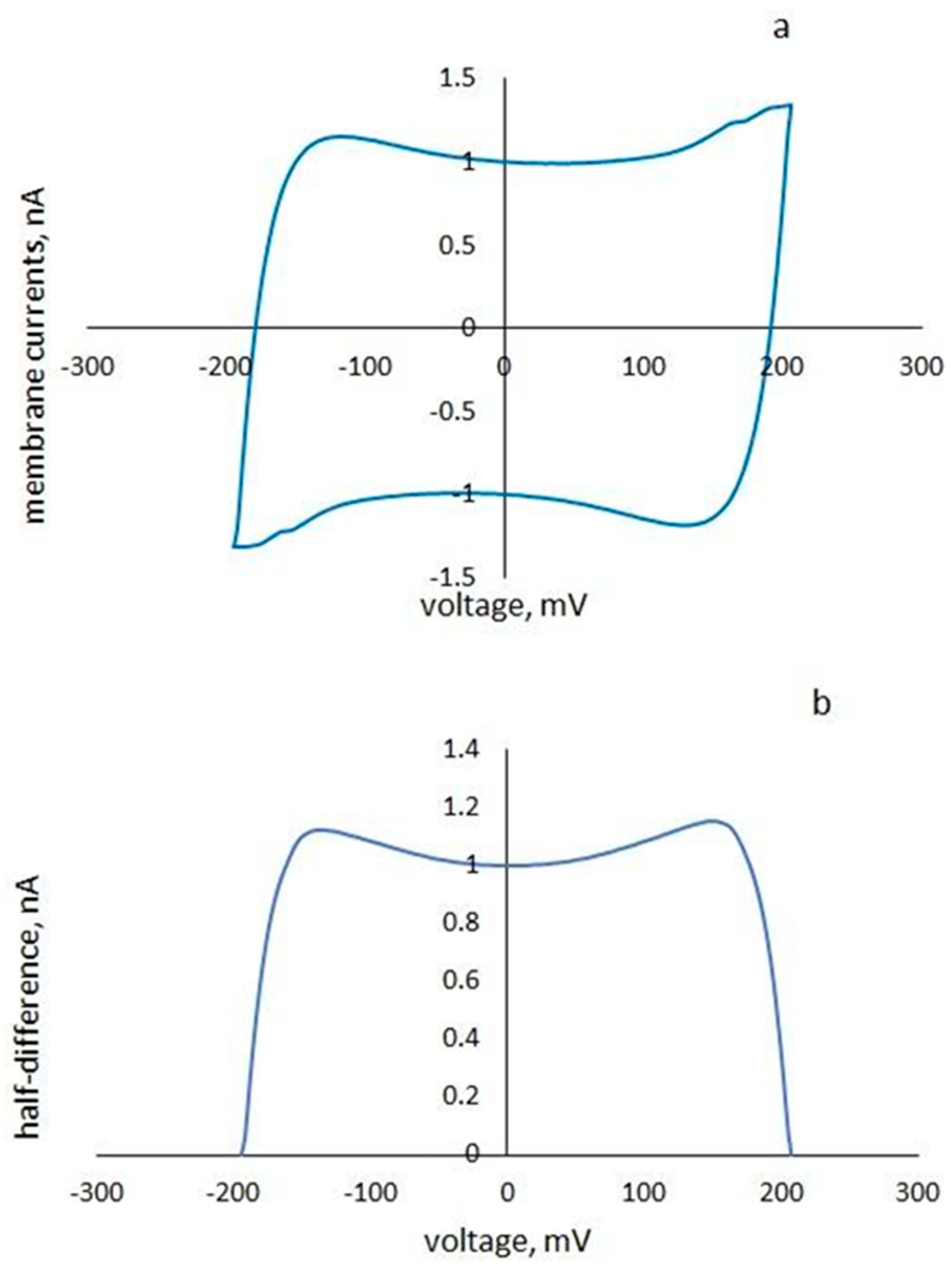
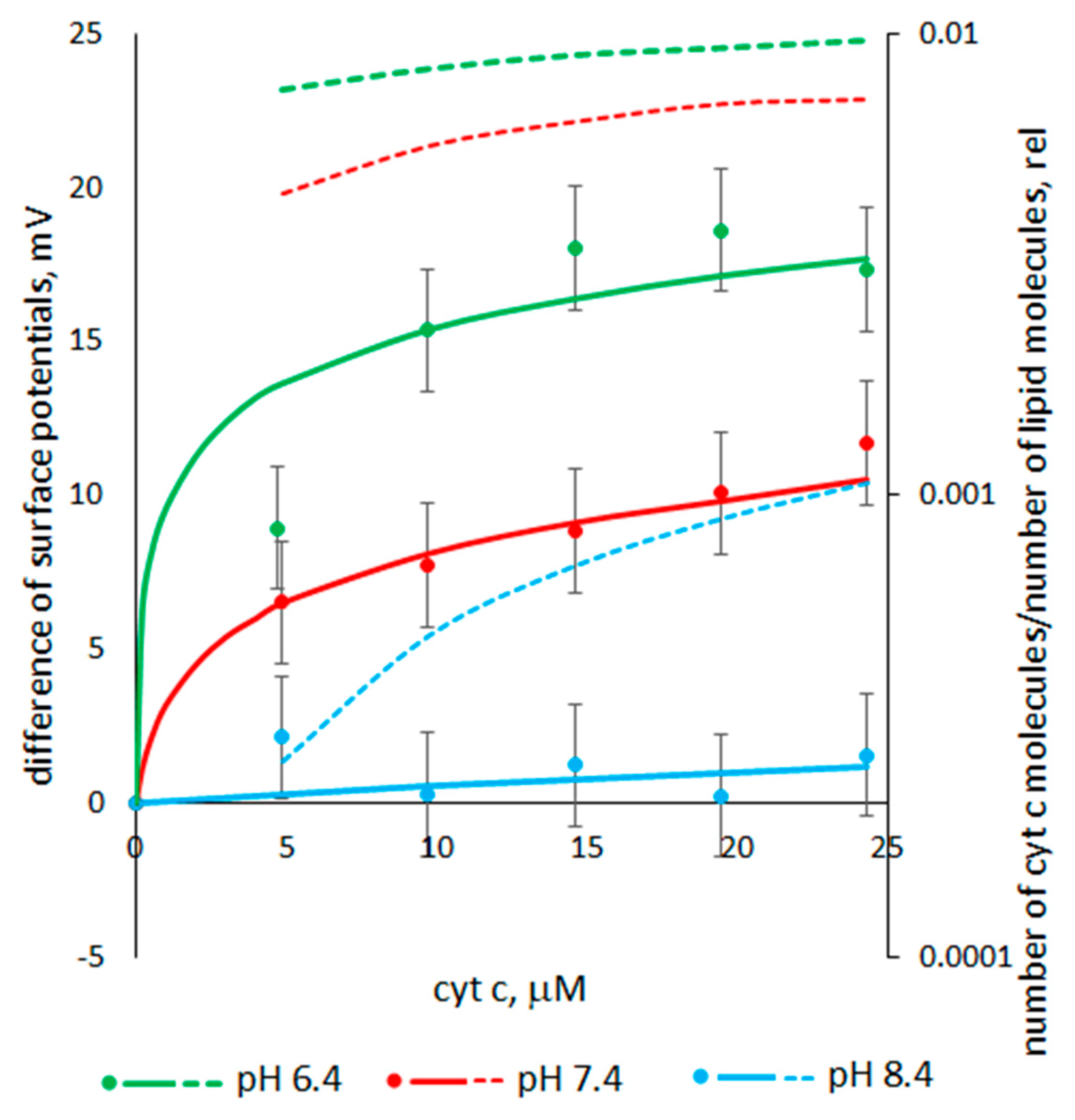
3.3. The Dependence of the Cyt C-Induced Difference in the Surface Potentials on pH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gorbenko, G.P.; Molotkovsky, J.G.; Kinnunen, P.K.J. Cytochrome c interaction with cardiolipin/phosphatidylcholine model membranes: Effect of cardiolipin protonation. Biophys. J. 2006, 90, 4093–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinibaldi, F.; Howes, B.D.; Piro, M.C.; Polticelli, F.; Bombelli, C.; Ferri, T.; Coletta, M.; Smulevich, G.; Santucci, R. Extended cardiolipin anchorage to cytochrome c: A model for protein–mitochondrial membrane binding. JBIC J. Biol. Inorg. Chem. 2010, 15, 689–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rytömaa, M.; Kinnunen, P.K. Reversibility of the Binding of Cytochrome c to Liposomes: Implications for Lipid-Protein Interactions (∗). J. Biol. Chem. 1995, 270, 3197–3202. [Google Scholar] [CrossRef] [Green Version]
- Milorey, B.; Schweitzer-Stenner, R.; Kurbaj, R.; Malyshka, D. pH-induced switch between different modes of cytochrome c binding to cardiolipin-containing liposomes. ACS Omega 2019, 4, 1386–1400. [Google Scholar] [CrossRef] [Green Version]
- Zuckermann, M.J.; Heimburg, T. Insertion and pore formation driven by adsorption of proteins onto lipid bilayer membrane–water interfaces. Biophys. J. 2001, 81, 2458–2472. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Dimitriadis, E.K. Cytochrome c adsorption to supported, anionic lipid bilayers studied via atomic force microscopy. Biophys. J. 2004, 87, 3234–3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweitzer-Stenner, R. Relating the multi-functionality of cytochrome c to membrane binding and structural conversion. Biophys. Rev. 2018, 10, 1151–1185. [Google Scholar] [CrossRef]
- Tuominen, E.K.J.; Wallace, C.J.A.; Kinnunen, P.K.J. Phospholipid-cytochrome c interaction: Evidence for the extended lipid anchorage. J. Biol. Chem. 2002, 277, 8822–8826. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, A.; Contreras, L.M.; Villalaín, J. Two-dimensional infrared correlation spectroscopy study of the interaction of oxidized and reduced cytochrome c with phospholipid model membranes. Biochim. Biophys. Acta (BBA) Biomembr. 2007, 1768, 2409–2420. [Google Scholar] [CrossRef] [Green Version]
- Bergstrom, C.L.; Beales, P.A.; Lv, Y.; Vanderlick, T.K.; Groves, J.T. Cytochrome c causes pore formation in cardiolipin-containing membranes. Proc. Natl. Acad. Sci. 2013, 110, 6269–6274. [Google Scholar] [CrossRef] [Green Version]
- Kitt, J.P.; Bryce, D.A.; Minteer, S.D.; Harris, J.M. Raman spectroscopy reveals selective interactions of cytochrome c with cardiolipin that correlate with membrane permeability. J. Am. Chem. Soc. 2017, 139, 3851–3860. [Google Scholar] [CrossRef] [PubMed]
- Belikova, N.A.; Vladimirov, Y.A.; Osipov, A.N.; Kapralov, A.A.; Tyurin, V.A.; Potapovich, M.V.; Basova, L.V.; Peterson, J.; Kurnikov, I.V.; Kagan, V.E. Peroxidase activity and structural transitions of cytochrome c bound to cardiolipin-containing membranes. Biochemistry 2006, 45, 4998–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, V.E.; Tyurin, V.A.; Jiang, J.; Tyurina, Y.Y.; Ritov, V.B.; Amoscato, A.A.; Osipov, A.N.; Belikova, N.A.; Kapralov, A.A.; Kini, V.; et al. Cytochrome c acts as a cardiolipin oxygenase required for release of proapoptotic factors. Nat. Chem. Biol. 2005, 1, 223–232. [Google Scholar] [CrossRef]
- Firsov, A.M.; Kotova, E.A.; Korepanova, E.A.; Osipov, A.N.; Antonenko, Y.N. Peroxidative permeabilization of liposomes induced by cytochrome c/cardiolipin complex. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 767–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchkov, M.N.; Vassarais, R.A.; Korepanova, E.A.; Osipov, A.N. Cytochrome c produces pores in cardiolipin-containing planar bilayer lipid membranes in the presence of hydrogen peroxide. Biochim. Biophys. Acta (BBA) Biomembr. 2013, 1828, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonov, V.F.; Puchkov, M.N.; Korepanova, E.A.; Nemchenko, O.Y.; Borodulin, V. Soft perforation of cardiolipin-containing planar lipid bilayer membrane by cytochrome c and H2O2. Eur. Biophys. J. 2014, 43, 469–476. [Google Scholar] [CrossRef]
- Kagan, V.E.; Borisenko, G.G.; Tyurina, Y.Y.; Tyurin, V.A.; Jiang, J.; Potapovich, A.I.; Fujii, Y. Oxidative lipidomics of apoptosis: Redox catalytic interactions of cytochrome c with cardiolipin and phosphatidylserine. Free. Radic. Biol. Med. 2004, 37, 1963–1985. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Wang, Z.; Huang, X.; Ge, Z.; Hu, B. Influence of structured water layers on protein adsorption process: A case study of cytochrome c and carbon nanotube interactions and its implications. J. Phys. Chem. B 2019, 124, 684–694. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, C.; Li, Z.; Lu, Z.; Li, Y.; Yin, J.J.; Zhou, Y.T.; Gao, X.; Fang, Y.; Nie, G.; et al. Unraveling stress-induced toxicity properties of graphene oxide and the underlying mechanism. Adv. Mater. 2012, 24, 5391–5397. [Google Scholar] [CrossRef]
- Xu, J.; Vanderlick, T.K.; Beales, P.A. Lytic and non-lytic permeabilization of cardiolipin-containing lipid bilayers induced by cytochrome c. PLoS ONE 2013, 8, e69492. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S.E.; Daum, G. Lipids of mitochondria. Prog. Lipid Res. 2013, 52, 590–614. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Llopis, J.; Deveraux, Q.L.; Tsien, R.Y.; Reed, J.C. Changes in intramitochondrial and cytosolic pH: Early events that modulate caspase activation during apoptosis. Nat. Cell Biol. 2000, 2, 318–325. [Google Scholar] [CrossRef]
- Mueller, P.; Rudin, D.O.; Ti Tien, H.; Wescott, W.C. Reconstitution of cell membrane structure in vitro and its transformation into an excitable system. Nature 1962, 194, 979–980. [Google Scholar] [CrossRef] [PubMed]
- DeBlois, R.W.; Bean, C.P.; Wesley, R.K. Electrokinetic measurements with submicron particles and pores by the resistive pulse technique. J. Colloid Interface Sci. 1977, 61, 323–335. [Google Scholar] [CrossRef]
- Glaser, R.W.; Leikin, S.L.; Chernomordik, L.V.; Pastushenko, V.F.; Sokirko, A.I. Reversible electrical breakdown of lipid bilayers: Formation and evolution of pores. Biochim. Biophys. Acta (BBA) Biomembr. 1988, 940, 275–287. [Google Scholar] [CrossRef]
- McLaughlin, S. The electrostatic properties of membranes. Annu. Rev. Biophys. Biophys. Chem. 1989, 18, 113–136. [Google Scholar] [CrossRef]
- Konstantinova, A.N.; Sokolov, V.S.; Jimenez-Munguia, I.; Finogenova, O.A.; Ermakov, Y.A.; Gorbunova, Y.G. Adsorption and photodynamic efficiency of meso-tetrakis (p-sulfonatophenyl) porphyrin on the surface of bilayer lipid membranes. J. Photochem. Photobiol. B Biol. 2018, 189, 74–80. [Google Scholar] [CrossRef]
- Alvarez, O.; Latorre, R. Voltage-dependent capacitance in lipid bilayers made from monolayers. Biophys. J. 1978, 21, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, E.Y.; Anosov, A.A. Bilayer Lipid Membrane as Memcapacitance: Capacitance–Voltage Pinched Hysteresis and Negative Insertion Conductance. Membranes 2023, 13, 97. [Google Scholar] [CrossRef]
- Cherny, V.V.; Sokolov, V.S.; Abidor, I.G. 330-Determination of surface charge of bilayer lipid membranes. Bioelectrochem. Bioenerg. 1980, 7, 413–420. [Google Scholar] [CrossRef]
- Anosov, A.A.; Smirnova, E.Y.; Korepanova, E.A.; Shogenov, I.M. The effects of SDS at subsolubilizing concentrations on the planar lipid bilayer permeability: Two kinds of current fluctuations. Chem. Phys. Lipids 2019, 218, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.; Papahadjopoulos, D. Phase transitions and phase separations in phospholipid membranes induced by changes in temperature, pH, and concentration of bivalent cations. Biochemistry 1975, 14, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Nagle, J.F.; Tristram-Nagle, S. Structure of lipid bilayers. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 2000, 1469, 159–195. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.C.; Myers, J.B.; Folta, T.; Shoja, V.; Heath, L.S.; Onufriev, A. H++: A server for estimating p K as and adding missing hydrogens to macromolecules. Nucleic Acids Res. 2005, 33 (Suppl. S2), W368–W371. [Google Scholar] [CrossRef] [PubMed]
- Anandakrishnan, R.; Aguilar, B.; Onufriev, A.V. H++ 3.0: Automating p K prediction and the preparation of biomolecular structures for atomistic molecular modeling and simulations. Nucleic Acids Res. 2012, 40, W537–W541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, E.Y.; Kozhomkulov, E.T.; Vasserman, A.N.; Vosnesensky, S.A.; Shevchenko, E.V.; Morozov, Y.V.; Antonov, V.F. Permeability of bilayer lipid membranes in phase transition. The significance of intermolecular phosphate-phosphate hydrogen bonding. Chem. Phys. Lipids 1986, 41, 173–180. [Google Scholar] [CrossRef]
- Verkleij, A.J.; De Kruyff, B.; Ververgaert, P.T.; Tocanne, J.F.; Van Deenen, L.L.M. The influence of pH, Ca2+ and protein on the thermotropic behaviour of the negatively charged phospholipid, phosphatidylglycerol. Biochim. Biophys. Acta (BBA) Biomembr. 1974, 339, 432–437. [Google Scholar] [CrossRef]
- Papahadjopoulos, D. Surface properties of acidic phospholipids: Interaction of monolayers and hydrated liquid crystals with uni- and bi-valent metal ions. Biochim. Biophys. Acta (BBA) Biomembr. 1968, 163, 240–254. [Google Scholar] [CrossRef]
- Teissie, J. Interaction of cytochrome c with phospholipid monolayers. Orientation and penetration of protein as functions of the packing density of film, nature of the phospholipids, and ionic content of the aqueous phase. Biochemistry 1981, 20, 1554–1560. [Google Scholar] [CrossRef]
- Peschke, J.; Möhwald, H. Cytochrome c interaction with phospholipid monolayers and vesicles. Colloids Surf. 1987, 27, 305–323. [Google Scholar] [CrossRef]
- Li, J.; Sun, R.; Hao, C.; He, G.; Zhang, L.; Wang, J. The behavior of the adsorption of cytochrome c on lipid monolayers: A study by the Langmuir–Blodgett technique and theoretical analysis. Biophys. Chem. 2015, 205, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Antonov, V.F.; Anosov, A.A.; Norik, V.P.; Smirnova, E.Y. Soft perforation of planar bilayer lipid membranes of dipalmitoylphosphatidylcholine at the temperature of the phase transition from the liquid crystalline to the gel state. Eur. Biophys. J. 2005, 34, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Anosov, A.; Astanina, P.; Proskuryakov, I.; Koplak, O.; Morgunov, R. Surface and Structure of Phosphatidylcholine Membranes Reconstructed with CoFe2O4 Nanoparticles. Langmuir 2022, 38, 14517–14526. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.A.; Wang, M.A.; Weaver, J.C. Theory of electroporation of planar bilayer membranes: Predictions of the aqueous area, change in capacitance, and pore-pore separation. Biophys. J. 1994, 67, 42–56. [Google Scholar] [CrossRef] [Green Version]
- Anosov, A.; Koplak, O.; Smirnova, E.; Borisova, E.; Korepanova, E.; Derunets, A. Effect of Cobalt Ferrite Nanoparticles in a Hydrophilic Shell on the Conductance of Bilayer Lipid Membrane. Membranes 2022, 12, 1106. [Google Scholar] [CrossRef]
- Powell, K.T.; Derrick, E.G.; Weaver, J.C. A quantitative theory of reversible electrical breakdown in bilayer membranes. Bioelectrochem. Bioenerg. 1986, 15, 243–255. [Google Scholar] [CrossRef]
- Anosov, A.A.; Smirnova, E.Y.; Ryleeva, E.D.; Gligonov, I.A.; Korepanova, E.A.; Sharakshane, A.A. Estimation of the parameters of the Smoluchowski equation describing the occurrence of pores in a bilayer lipid membrane under soft poration. Eur. Phys. J. E 2020, 43, 1–9. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anosov, A.; Borisova, E.; Smirnova, E.; Korepanova, E.; Osipov, A. Effect of Cytochrome C on the Conductance of Asolectin Membranes and the Occurrence of Through Pores at Different pHs. Membranes 2023, 13, 268. https://doi.org/10.3390/membranes13030268
Anosov A, Borisova E, Smirnova E, Korepanova E, Osipov A. Effect of Cytochrome C on the Conductance of Asolectin Membranes and the Occurrence of Through Pores at Different pHs. Membranes. 2023; 13(3):268. https://doi.org/10.3390/membranes13030268
Chicago/Turabian StyleAnosov, Andrey, Elizaveta Borisova, Elena Smirnova, Eugenia Korepanova, and Anatoly Osipov. 2023. "Effect of Cytochrome C on the Conductance of Asolectin Membranes and the Occurrence of Through Pores at Different pHs" Membranes 13, no. 3: 268. https://doi.org/10.3390/membranes13030268
APA StyleAnosov, A., Borisova, E., Smirnova, E., Korepanova, E., & Osipov, A. (2023). Effect of Cytochrome C on the Conductance of Asolectin Membranes and the Occurrence of Through Pores at Different pHs. Membranes, 13(3), 268. https://doi.org/10.3390/membranes13030268