


The Effect of Cholesterol in SOPC Lipid Bilayers at Low Temperatures


Abstract
:1. Introduction
2. Systems and Methods
3. Results and Discussion
3.1. Bilayer Characteristics
3.1.1. Mass Density Distribution
3.1.2. Electrostatic Potential
3.1.3. Isobaric Molar Heat Capacity
3.2. Structural Characteristics of the Lipids
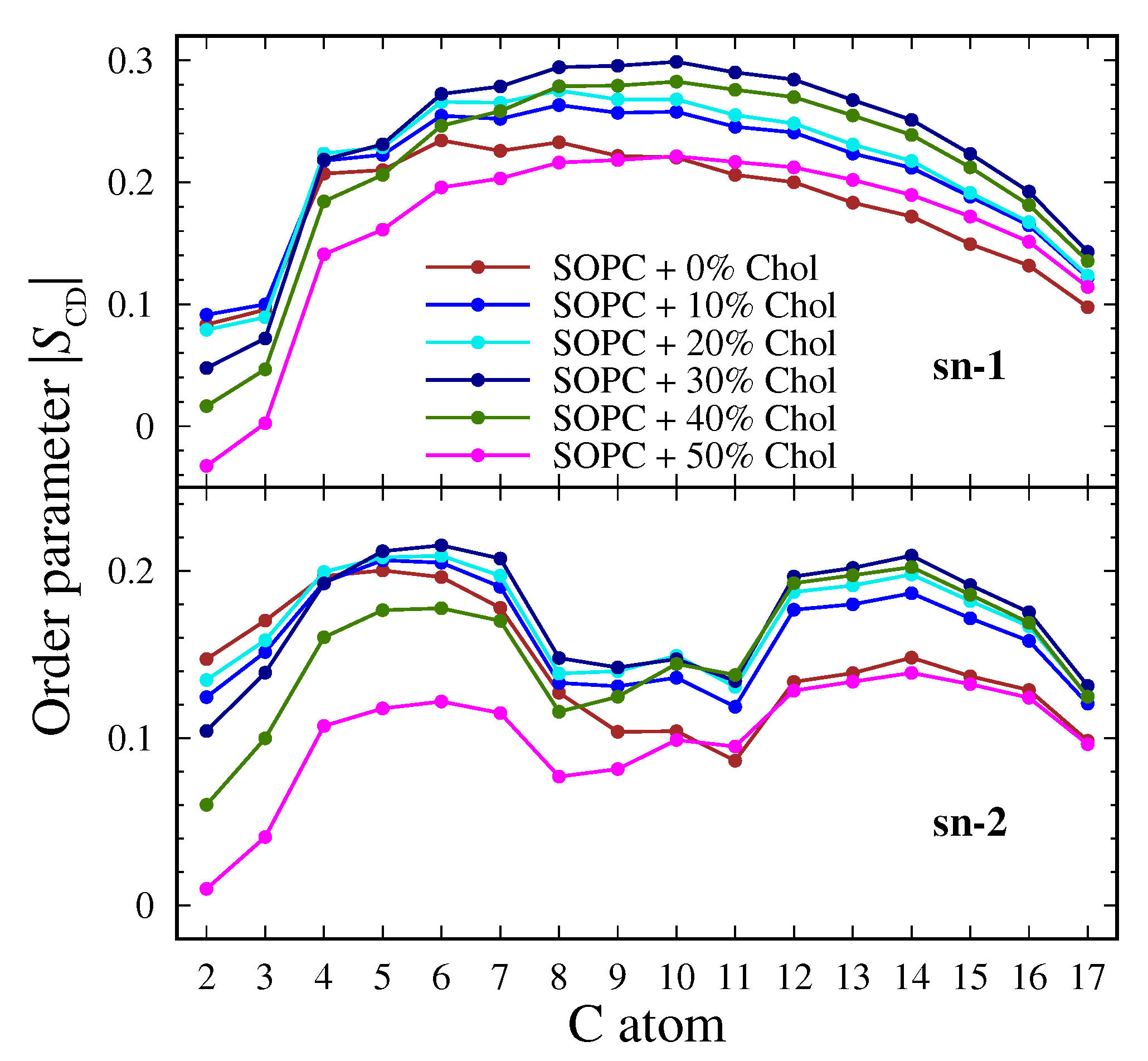
3.2.1. Order Parameter of the Lipid Tails
3.2.2. Tilt Angles of the Lipid Tails
3.3. Area per Lipid and Lateral Diffusion Coefficient
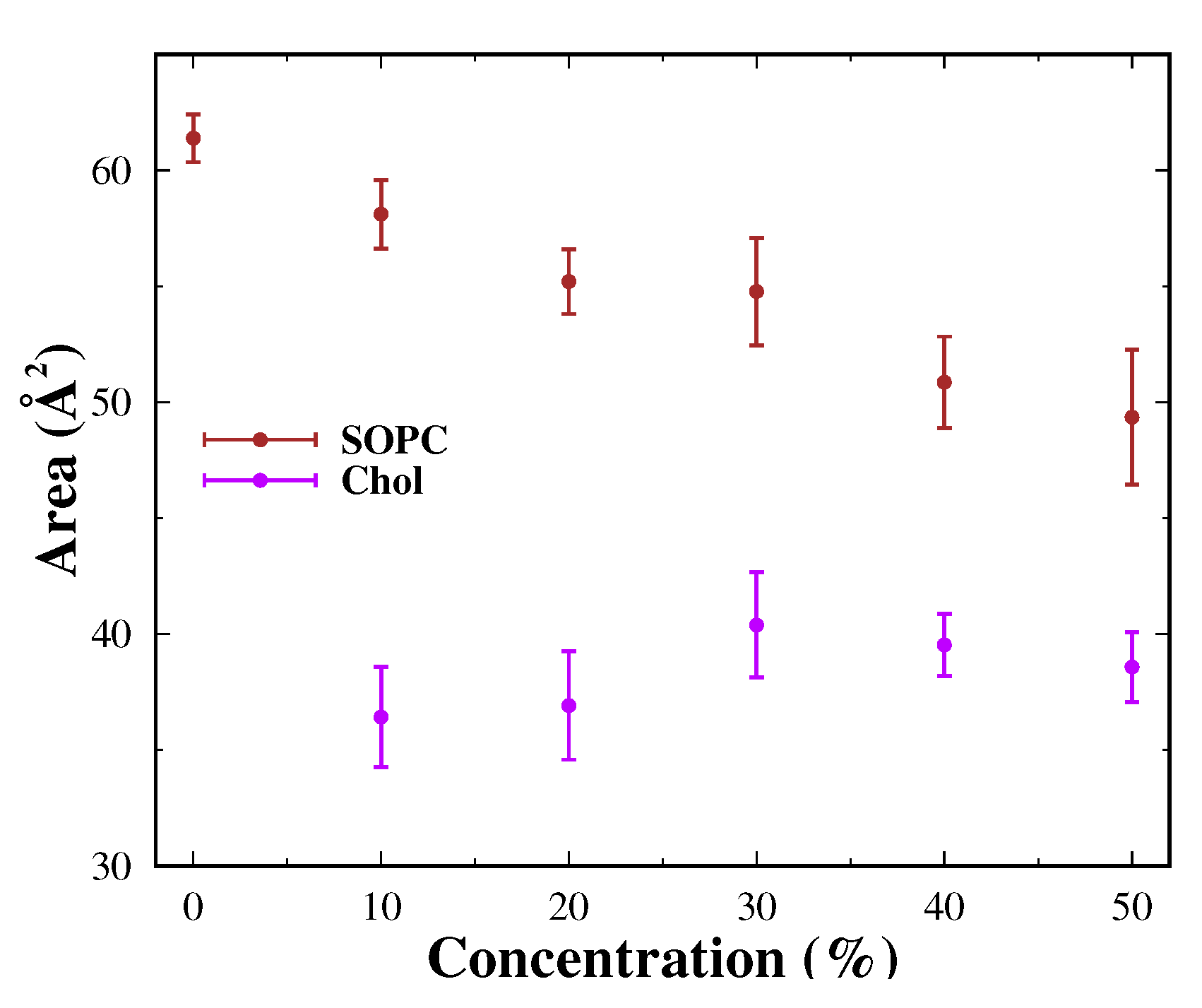
3.3.1. Area per Lipid
3.3.2. Lateral Diffusion Coefficient
3.4. Structural Behavior of the Membranes
3.4.1. Radial Distribution Function
3.4.2. Hydrogen Bonds
4. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stillwell, W. An Introduction to Biological Membranes: Composition, Structure and Function, 2nd ed.; Academic Press: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 7th ed.; W. H. Freeman: New York, NY, USA; Basingstoke, UK, 2017. [Google Scholar]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell. Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Wirtz, K.W.A. Phospholipid Transfer Proteins. Annu. Rev. Biochem. 1991, 60, 73–99. [Google Scholar] [CrossRef]
- Chamati, H. Theory of Phase Transitions: From Magnets to Biomembranes. Adv. Biomembr. Lipid Self-Assem. 2013, 17, 237. [Google Scholar] [CrossRef]
- Goldstein, R.E.; Leibler, S. Model for Lamellar Phases of Interacting Lipid Membranes. Phys. Rev. Lett. 1988, 61, 2213–2216. [Google Scholar] [CrossRef]
- Heimburg, T. Thermal Biophysics of Membranes; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Janiak, M.J.; Small, D.M.; Shipley, G.G. Temperature and compositional dependence of the structure of hydrated dimyristoyl lecithin. J. Biol. Chem. 1979, 254, 6068–6078. [Google Scholar] [CrossRef]
- Ipsen, J.H.; Karlström, G.; Mourtisen, O.G.; Wennerström, H.; Zuckermann, M.J. Phase equilibria in the phosphatidylcholine-cholesterol system. Biochim. Biophys. Acta Biomembr. 1987, 905, 162–172. [Google Scholar] [CrossRef]
- Gally, H.U.; Seelig, A.; Seelig, J. Cholesterol-induced rod-like motion of fatty acyl chains in lipid bilayers a deuterium magnetic resonance study. Hoppe-Seyler Z. Fur Physiol. Chem. 1976, 357, 1447–1450. [Google Scholar]
- Polozov, I.V.; Gawrisch, K. Characterization of the Liquid-Ordered State by Proton MAS NMR. Biophys. J. 2006, 90, 2051. [Google Scholar] [CrossRef] [Green Version]
- Genova, J.; Chamati, H.; Petrov, M. Physico-chemical characterizations of lipid membranes in presence of cholesterol. Adv. Biomembr. Lipid Self-Assem. 2020, 31, 1. [Google Scholar] [CrossRef]
- Róg, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. What Happens if Cholesterol Is Made Smoother. Biophys. J. 2007, 92, 3346–3357. [Google Scholar] [CrossRef] [Green Version]
- MacDermaid, C.M.; Kashyap, H.K.; DeVane, R.H.; Shinoda, W.; Klauda, J.B.; Klein, M.L.; Fiorin, G. Molecular dynamics simulations of cholesterol-rich membranes using a coarse-grained force field for cyclic alkanes. J. Chem. Phys. 2015, 143, 243144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monje-Galvan, V.; Klauda, J.B. Two sterols, two bilayers: Insights on membrane structure from molecular dynamics. Mol. Simul. 2017, 43, 1179–1188. [Google Scholar] [CrossRef]
- Phillips, R.; Kondev, J.; Theriot, J.; Garcia, H. Physical Biology of the Cell, 2nd ed.; Garland Science: London, UK, 2012. [Google Scholar]
- Karp, G. Cell and Molecular Biology: Concepts and Experiments, 7th ed.; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar]
- Sankaram, M.B.; Thompson, T.E. Modulation of phospholipid acyl chain order by cholesterol. A solid-state 2H nuclear magnetic resonance study. Biochemistry 1990, 29, 10676–10684. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, N.; Genova, J.; Chamati, H. Physical properties of SOPC lipid membranes containing cholesterol by molecular dynamics simulation. Adv. Biomembr. Lipid Self-Assem. 2021, 34, 1–30. [Google Scholar] [CrossRef]
- van der Ploeg, P.; Berendsen, H.J.C. Molecular dynamics simulation of a bilayer membrane. J. Chem. Phys. 1982, 76, 3271–3276. [Google Scholar] [CrossRef] [Green Version]
- Venable, R.M.; Zhang, Y.; Hardy, B.J.; Pastor, R.W. Molecular Dynamics Simulations of a Lipid Bilayer and of Hexadecane: An Investigation of Membrane Fluidity. Science 1993, 262, 223–226. [Google Scholar] [CrossRef]
- Feller, S.E.; Venable, R.M.; Pastor, R.W. Computer Simulation of a DPPC Phospholipid Bilayer: Structural Changes as a Function of Molecular Surface Area. Langmuir 1997, 13, 6555–6561. [Google Scholar] [CrossRef]
- Pastor, R.W.; Venable, R.M.; Feller, S.E. Lipid Bilayers, NMR Relaxation, and Computer Simulations. Acc. Chem. Res. 2002, 35, 438–446. [Google Scholar] [CrossRef]
- Brooks, B.R.; Bruccoleri, R.E.; Olafson, B.D.; States, D.J.; Swaminathan, S.; Karplus, M. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 1983, 4, 187–217. [Google Scholar] [CrossRef]
- Klauda, J.B.; Venable, R.M.; Freites, J.A.; O’Connor, J.W.; Tobias, D.J.; Mondragon-Ramirez, C.; Vorobyov, I.; MacKerell, A.D.; Pastor, R.W. Update of the CHARMM All-Atom Additive Force Field for Lipids: Validation on Six Lipid Types. J. Phys. Chem. B 2010, 114, 7830–7843. [Google Scholar] [CrossRef] [Green Version]
- Dickson, C.J.; Madej, B.D.; Skjevik, o.A.; Betz, R.M.; Teigen, K.; Gould, I.R.; Walker, R.C. Lipid14: The Amber Lipid Force Field. J. Chem. Theory Comput. 2014, 10, 865–879. [Google Scholar] [CrossRef]
- Leonard, A.N.; Wang, E.; Monje-Galvan, V.; Klauda, J.B. Developing and Testing of Lipid Force Fields with Applications to Modeling Cellular Membranes. Chem. Rev. 2019, 119, 6227–6269. [Google Scholar] [CrossRef]
- Sonne, J.; Jensen, M.Ø.; Hansen, F.Y.; Hemmingsen, L.; Peters, G.H. Reparameterization of All-Atom Dipalmitoylphosphatidylcholine Lipid Parameters Enables Simulation of Fluid Bilayers at Zero Tension. Biophys. J. 2007, 92, 4157–4167. [Google Scholar] [CrossRef] [Green Version]
- Högberg, C.J.; Nikitin, A.M.; Lyubartsev, A.P. Modification of the CHARMM force field for DMPC lipid bilayer. J. Comput. Chem. 2008, 29, 2359–2369. [Google Scholar] [CrossRef]
- Jämbeck, J.P.M.; Lyubartsev, A.P. Derivation and Systematic Validation of a Refined All-Atom Force Field for Phosphatidylcholine Lipids. J. Phys. Chem. B 2012, 116, 3164. [Google Scholar] [CrossRef]
- Grote, F.; Lyubartsev, A.P. Optimization of Slipids Force Field Parameters Describing Headgroups of Phospholipids. J. Phys. Chem. B 2020, 124, 8784–8793. [Google Scholar] [CrossRef]
- Antila, H.S.; Ferreira, T.M.; Ollila, O.H.S.; Miettinen, M.S. Using Open Data to Rapidly Benchmark Biomolecular Simulations: Phospholipid Conformational Dynamics. J. Chem. Inf. Model. 2021, 61, 938–949. [Google Scholar] [CrossRef]
- Antila, H.; Buslaev, P.; Favela-Rosales, F.; Ferreira, T.M.; Gushchin, I.; Javanainen, M.; Kav, B.; Madsen, J.J.; Melcr, J.; Miettinen, M.S.; et al. Headgroup Structure and Cation Binding in Phosphatidylserine Lipid Bilayers. J. Phys. Chem. B 2019, 123, 9066–9079. [Google Scholar] [CrossRef]
- Favela-Rosales, F.; Galván-Hernández, A.; Hernández-Cobos, J.; Kobayashi, N.; Carbajal-Tinoco, M.D.; Nakabayashi, S.; Ortega-Blake, I. A molecular dynamics study proposing the existence of statistical structural heterogeneity due to chain orientation in the POPC-cholesterol bilayer. Biophys. Chem. 2020, 257, 106275. [Google Scholar] [CrossRef]
- Ivanova, N.; Chamati, H. Physical properties of phospholipids at low temperatures through Slipid force field. J. Phys. Conf. Ser. 2023, 2436, 012025. [Google Scholar] [CrossRef]
- Genova, J.; Chamati, H.; Slavkova, Z.; Petrov, M. Differential Scanning Calorimetric Study of the Effect of Cholesterol on the Thermotropic Phase Behavior of the Phospholipid 1-Stearoyl-2-Oleoyl-sn-Glycero-3-Phosphocholine. J. Surfactants Deterg. 2019, 22, 1229. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L., III; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [Green Version]
- Jo, S.; Lim, J.B.; Klauda, J.B.; Im, W. CHARMM-GUI membrane builder for mixed bilayers and its application to yeast membranes. Biophys. J. 2009, 97, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theor. Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef]
- Hung, W.C.; Lee, M.T.; Chen, F.Y.; Huang, H.W. The Condensing Effect of Cholesterol in Lipid Bilayers. Biophys. J. 2007, 92, 3960–3967. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; McIntosh, T. Probing the ethanol-induced chain interdigitations in gel-state bilayers of mixed-chain phosphatidylcholines. Biophys. J. 1997, 72, 2702–2709. [Google Scholar] [CrossRef] [Green Version]
- Maulik, P.R.; Shipley, G.G. N-Parmitoyl Sphingomyelin Bilayers: Structure and Interactions with Cholesterol and Dipalmitoylphosphatidylcholine. Biochemistry 1996, 35, 8025–8034. [Google Scholar] [CrossRef]
- Tristram-Nagle, S.; Zhang, R.; Suter, R.M.; Worthington, C.R.; Sun, W.J.; Nagle, J.F. Measurement of chain tilt angle in fully hydrated bilayers of gel phase lecithins. Biophys. J. 1993, 64, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Tristram-Nagle, S.; Nagle, J.F. Effect of cholesterol on structural and mechanical properties of membranes depends on lipid chain saturation. Phys. Rev. E 2009, 80, 021931. [Google Scholar] [CrossRef] [Green Version]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; van der Spoel, D.; et al. GROMACS 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845. [Google Scholar] [CrossRef] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindah, E. Gromacs: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Lindahl; Abraham; Hess; Spoel, V.D. GROMACS 2021 Manual. In Zenodo; 2021. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Allen, M.P.; Tildesley, D.J.; Banavar, J.R. Computer Simulation of Liquids. Phys. Today 1989, 42, 105–106. [Google Scholar] [CrossRef]
- Ewald, P.P. Die Berechnung optischer und elektrostatischer Gitterpotentiale. Ann. Phys. 1921, 369, 253–287. [Google Scholar] [CrossRef] [Green Version]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.; Postma, J.P.; Van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Nosé, S. A molecular dynamics method for simulations in the canonical ensemble. Mol. Phys. 1984, 52, 255–268. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695–1697. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Nosé, S.; Klein, M. Constant pressure molecular dynamics for molecular systems. Mol. Phys. 1983, 50, 1055–1076. [Google Scholar] [CrossRef]
- Kandt, C.; Ash, W.L.; Peter Tieleman, D. Setting up and running molecular dynamics simulations of membrane proteins. Methods 2007, 41, 475–488. [Google Scholar] [CrossRef]
- Ivanova, N.; Ivanova, A. Testing the limits of model membrane simulations—bilayer composition and pressure scaling. J. Comput. Chem. 2018, 39, 387–396. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33. [Google Scholar] [CrossRef]
- Guixà-González, R.; Rodriguez-Espigares, I.; Ramírez-Anguita, J.M.; Carrió-Gaspar, P.; Martinez-Seara, H.; Giorgino, T.; Selent, J. MEMBPLUGIN: Studying membrane complexity in VMD. Bioinformatics 2014, 30, 1478–1480. [Google Scholar] [CrossRef] [Green Version]
- Barber, C.B.; Dobkin, D.P.; Huhdanpaa, H. The quickhull algorithm for convex hulls. ACM Trans. Math. Softw. 1996, 22, 469–483. [Google Scholar] [CrossRef] [Green Version]
- Ono, S.; Kondo, S. Molecular Theory of Surface Tension in Liquids. In Structure of Liquids / Struktur der Flüssigkeiten; Springer: Berlin, Germany, 1960; pp. 134–280. [Google Scholar] [CrossRef]
- Ivanova, N.; Ivanova, A. Influence of the dimensionality of the periodic boundary conditions on the transport of a drug–peptide complex across model cell membranes. J. Biomol. Struct. Dyn. 2022, 40, 5345–5356. [Google Scholar] [CrossRef]
- Doktorova, M.; LeVine, M.V.; Khelashvili, G.; Weinstein, H. A New Computational Method for Membrane Compressibility: Bilayer Mechanical Thickness Revisited. Biophys. J. 2019, 116, 487–502. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, X.; Makover, J.R.; Im, W.; Klauda, J.B. A systematic molecular dynamics simulation study of temperature dependent bilayer structural properties. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2520–2529. [Google Scholar] [CrossRef] [Green Version]
- Kučerka, N.; Nieh, M.P.; Katsaras, J. Fluid phase lipid areas and bilayer thicknesses of commonly used phosphatidylcholines as a function of temperature. Biochim. Biophys. Acta Biomembr. 2011, 1808, 2761–2771. [Google Scholar] [CrossRef] [PubMed]
- Levine, Y.K.; Wilkins, M.H.F. Structure of Oriented Lipid Bilayers. Nat. New Biol. 1971, 230, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Pencer, J.; Nieh, M.P.; Harroun, T.A.; Krueger, S.; Adams, C.; Katsaras, J. Bilayer thickness and thermal response of dimyristoylphosphatidylcholine unilamellar vesicles containing cholesterol, ergosterol and lanosterol: A small-angle neutron scattering study. Biochim. Biophys. Acta 2005, 1720, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Mills, T.T.; Tristram-Nagle, S.; Nagle, J.F. Cholesterol Perturbs Lipid Bilayers Nonuniversally. Phys. Rev. Lett. 2008, 100, 198103. [Google Scholar] [CrossRef]
- Gurtovenko, A.A.; Vattulainen, I. Calculation of the electrostatic potential of lipid bilayers from molecular dynamics simulations: Methodological issues. J. Chem. Phys. 2009, 130, 215107. [Google Scholar] [CrossRef]
- Tieleman, D.P.; Berendsen, H.J. Molecular dynamics simulations of a fully hydrated dipalmitoylphosphatidylcholine bilayer with different macroscopic boundary conditions and parameters. J. Chem. Phys. 1996, 105, 4871–4880. [Google Scholar] [CrossRef] [Green Version]
- Clarke, R.J. The dipole potential of phospholipid membranes and methods for its detection. Adv. Colloid Interface Sci. 2001, 89–90, 263–281. [Google Scholar] [CrossRef]
- Blume, A. Apparent molar heat capacities of phospholipids in aqueous dispersion. Effects of chain length and head group structure. Biochemistry 1983, 22, 5436–5442. [Google Scholar] [CrossRef]
- Ivanova, V.P.; Makarov, I.M.; Schäffer, T.E.; Heimburg, T. Analyzing Heat Capacity Profiles of Peptide-Containing Membranes: Cluster Formation of Gramicidin A. Biophys. J. 2003, 84, 2427–2439. [Google Scholar] [CrossRef] [Green Version]
- Blicher, A.; Wodzinska, K.; Fidorra, M.; Winterhalter, M.; Heimburg, T. The Temperature Dependence of Lipid Membrane Permeability, its Quantized Nature, and the Influence of Anesthetics. Biophys. J. 2009, 96, 4581–4591. [Google Scholar] [CrossRef] [Green Version]
- Vermeer, L.S.; de Groot, B.L.; Réat, V.; Milon, A.; Czaplicki, J. Acyl chain order parameter profiles in phospholipid bilayers: Computation from molecular dynamics simulations and comparison with 2H NMR experiments. Eur. Biophys. J. 2007, 36, 919–931. [Google Scholar] [CrossRef] [Green Version]
- Seelig, A.; Seelig, J. Dynamic structure of fatty acyl chains in a phospholipid bilayer measured by deuterium magnetic resonance. Biochemistry 1974, 13, 4839–4845. [Google Scholar] [CrossRef]
- Hofsäss, C.; Lindahl, E.; Edholm, O. Molecular dynamics simulations of phospholipid bilayers with cholesterol. Biophys. J. 2003, 84, 2192–2206. [Google Scholar] [CrossRef] [Green Version]
- Capponi, S.; Freites, J.A.; Tobias, D.J.; White, S.H. Interleaflet mixing and coupling in liquid-disordered phospholipid bilayers. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 354–362. [Google Scholar] [CrossRef]
- Wisniewska, A.; Wolnicka-Glubisz, A. ESR studies on the effect of cholesterol on chlorpromazine interaction with saturated and unsaturated liposome membranes. Biophys. Chem. 2004, 111, 43–52. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, J.; Heberle, F.A.; Mills, T.T.; Klawitter, P.; Huang, G.; Costanza, G.; Feigenson, G.W. Phase studies of model biomembranes: Complex behavior of DSPC/DOPC/Cholesterol. Biochim. Biophys. Acta Biomembr. 2007, 1768, 2764–2776. [Google Scholar] [CrossRef] [Green Version]
- Nagle, J.F. Experimentally determined tilt and bending moduli of single-component lipid bilayers. Chem. Phys. Lipids 2017, 205, 18–24. [Google Scholar] [CrossRef]
- Watts, A.; Harlos, K.; Marsh, D. Charge-induced tilt in ordered-phase phosphatidylglycerol bilayers Evidence from X-ray diffraction. Biochim. Biophys. Acta (BBA)-Biomembr. 1981, 645, 91–96. [Google Scholar] [CrossRef]
- Kučerka, N.; Gallová, J.; Uhríková, D.; Balgavý, P.; Bulacu, M.; Marrink, S.J.; Katsaras, J. Areas of Monounsaturated Diacylphosphatidylcholines. Biophys. J. 2009, 97, 1926–1932. [Google Scholar] [CrossRef] [Green Version]
- Kučerka, N.; Tristram-Nagle, S.; Nagle, J.F. Structure of fully hydrated fluid phase lipid bilayers with monounsaturated chains. J. Membr. Biol. 2005, 208, 193–202. [Google Scholar] [CrossRef]
- Koenig, B.W.; Strey, H.H.; Gawrisch, K. Membrane lateral compressibility determined by NMR and x-ray diffraction: Effect of acyl chain polyunsaturation. Biophys. J. 1997, 73, 1954–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molugu, T.R.; Brown, M.F. Cholesterol Effects on the Physical Properties of Lipid Membranes Viewed by Solid-state NMR Spectroscopy. In Cholesterol Modulation of Protein Function; Rosenhouse-Dantsker, A., Bukiya, A.N., Eds.; Series Title: Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2019; Volume 1115, pp. 99–133. [Google Scholar] [CrossRef]
- Smondyrev, A.M.; Berkowitz, M.L. Structure of Dipalmitoylphosphatidylcholine/Cholesterol Bilayer at Low and High Cholesterol Concentrations: Molecular Dynamics Simulation. Biophys. J. 1999, 77, 2075–2089. [Google Scholar] [CrossRef] [Green Version]
- Einstein, A. Zur Theorie der Brownschen Bewegung. Ann. Phys. 1906, 324, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Lindblom, G.; Orädd, G. Lipid lateral diffusion and membrane heterogeneity. Biochim. Biophys. Acta 2009, 1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrink, S.J.; Risselada, J.; Mark, A.E. Simulation of gel phase formation and melting in lipid bilayers using a coarse grained model. Chem. Phys. Lipids 2005, 135, 223–244. [Google Scholar] [CrossRef] [Green Version]
- Filippov, A.; Orädd, G.; Lindblom, G. Sphingomyelin structure influences the lateral diffusion and raft formation in lipid bilayers. Biophys. J. 2006, 90, 2086–2092. [Google Scholar] [CrossRef] [Green Version]
- Filippov, A.; Orädd, G.; Lindblom, G. Domain Formation in Model Membranes Studied by Pulsed-Field Gradient-NMR: The Role of Lipid Polyunsaturation. Biophys. J. 2007, 93, 3182–3190. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | N – SOPC | A – SOPC | N – Chol | A – Chol | N – Water | z |
|---|---|---|---|---|---|---|
| SOPC + 0% Chol | 256 | 66 | 0 | 0 | 13496 | 9.49 |
| SOPC + 10% Chol | 232 | 62 | 24 | 30 | 12,192 | 9.50 |
| SOPC + 20% Chol | 204 | 60 | 52 | 29 | 11,428 | 9.51 |
| SOPC + 30% Chol | 180 | 60 | 76 | 27 | 11,009 | 9.52 |
| SOPC + 40% Chol | 154 | 59 | 102 | 28 | 10,314 | 9.53 |
| SOPC + 50% Chol | 128 | 58 | 128 | 29 | 10,056 | 9.54 |
| System | Bilayer Thickness | Submergence Depth | Hydrophobic Layer |
|---|---|---|---|
| SOPC + 0% Chol | 4.11 | 1.19 | 0.59 |
| SOPC + 10% Chol | 4.31 | 1.25 | 0.78 |
| SOPC + 20% Chol | 4.35 | 1.41 | 0.98 |
| SOPC + 30% Chol | 4.19 | 1.32 | 1.13 |
| SOPC + 40% Chol | 4.21 | 1.37 | 1.56 |
| SOPC + 50% Chol | 4.10 | 1.43 | 1.81 |
| System | Standard Error | |
|---|---|---|
| SOPC + 0% Chol | 134 | 25 |
| SOPC + 10% Chol | 180 | 64 |
| SOPC + 20% Chol | 278 | 53 |
| SOPC + 30% Chol | 504 | 38 |
| SOPC + 40% Chol | 1484 | 43 |
| SOPC + 50% Chol | 3492 | 20 |
| System | Lateral Diffusion Coefficient | Standard Error |
|---|---|---|
| SOPC + 0% Chol | 1.2 | 0.2 |
| SOPC + 10% Chol | 1.0 | 0.5 |
| SOPC + 20% Chol | 1.5 | 0.7 |
| SOPC + 30% Chol | 0.8 | 0.8 |
| SOPC + 40% Chol | 1.4 | 1.1 |
| SOPC + 50% Chol | 1.9 | 1.8 |
| System | SOPC–Water | Chol–Water |
|---|---|---|
| SOPC + 0% Chol | 6.008 ± 2.448 | – |
| SOPC + 10% Chol | 5.612 ± 2.380 | 1.311 ± 1.187 |
| SOPC + 20% Chol | 5.346 ± 2.311 | 3.962 ± 2.014 |
| SOPC + 30% Chol | 4.834 ± 2.221 | 7.141 ± 2.621 |
| SOPC + 40% Chol | 4.534 ± 2.130 | 7.203 ± 2.719 |
| SOPC + 50% Chol | 3.991 ± 2.000 | 6.312 ± 2.526 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, N.; Chamati, H. The Effect of Cholesterol in SOPC Lipid Bilayers at Low Temperatures. Membranes 2023, 13, 275. https://doi.org/10.3390/membranes13030275
Ivanova N, Chamati H. The Effect of Cholesterol in SOPC Lipid Bilayers at Low Temperatures. Membranes. 2023; 13(3):275. https://doi.org/10.3390/membranes13030275
Chicago/Turabian StyleIvanova, Nikoleta, and Hassan Chamati. 2023. "The Effect of Cholesterol in SOPC Lipid Bilayers at Low Temperatures" Membranes 13, no. 3: 275. https://doi.org/10.3390/membranes13030275
APA StyleIvanova, N., & Chamati, H. (2023). The Effect of Cholesterol in SOPC Lipid Bilayers at Low Temperatures. Membranes, 13(3), 275. https://doi.org/10.3390/membranes13030275