

Cholesterol in Class C GPCRs: Role, Relevance, and Localization

Abstract
:1. Introduction
1.1. Cholesterol–Membrane Interactions
1.2. GPCR–Cholesterol Interactions
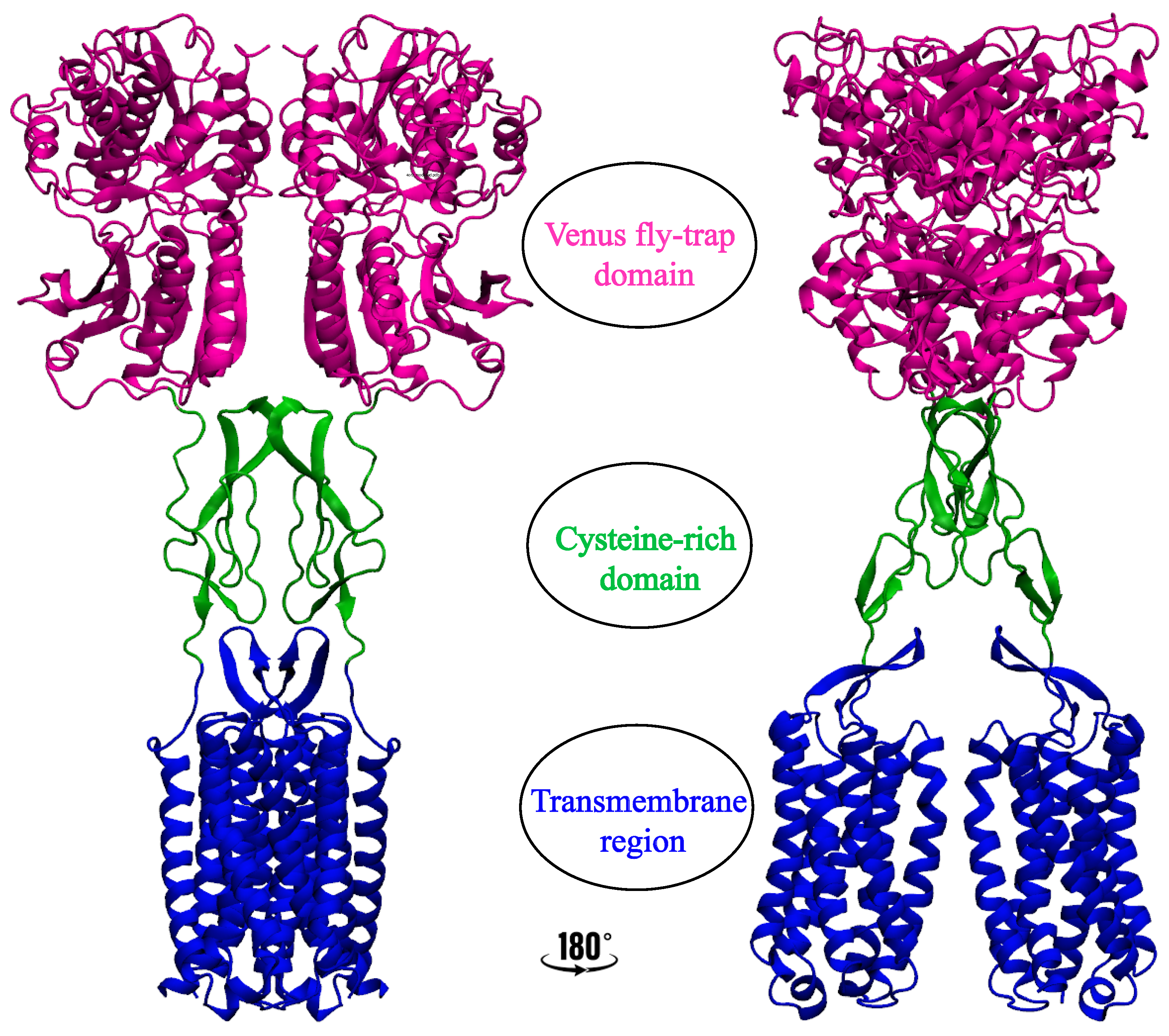
2. Significance and Interaction Sites of Cholesterol in Class C GPCRs
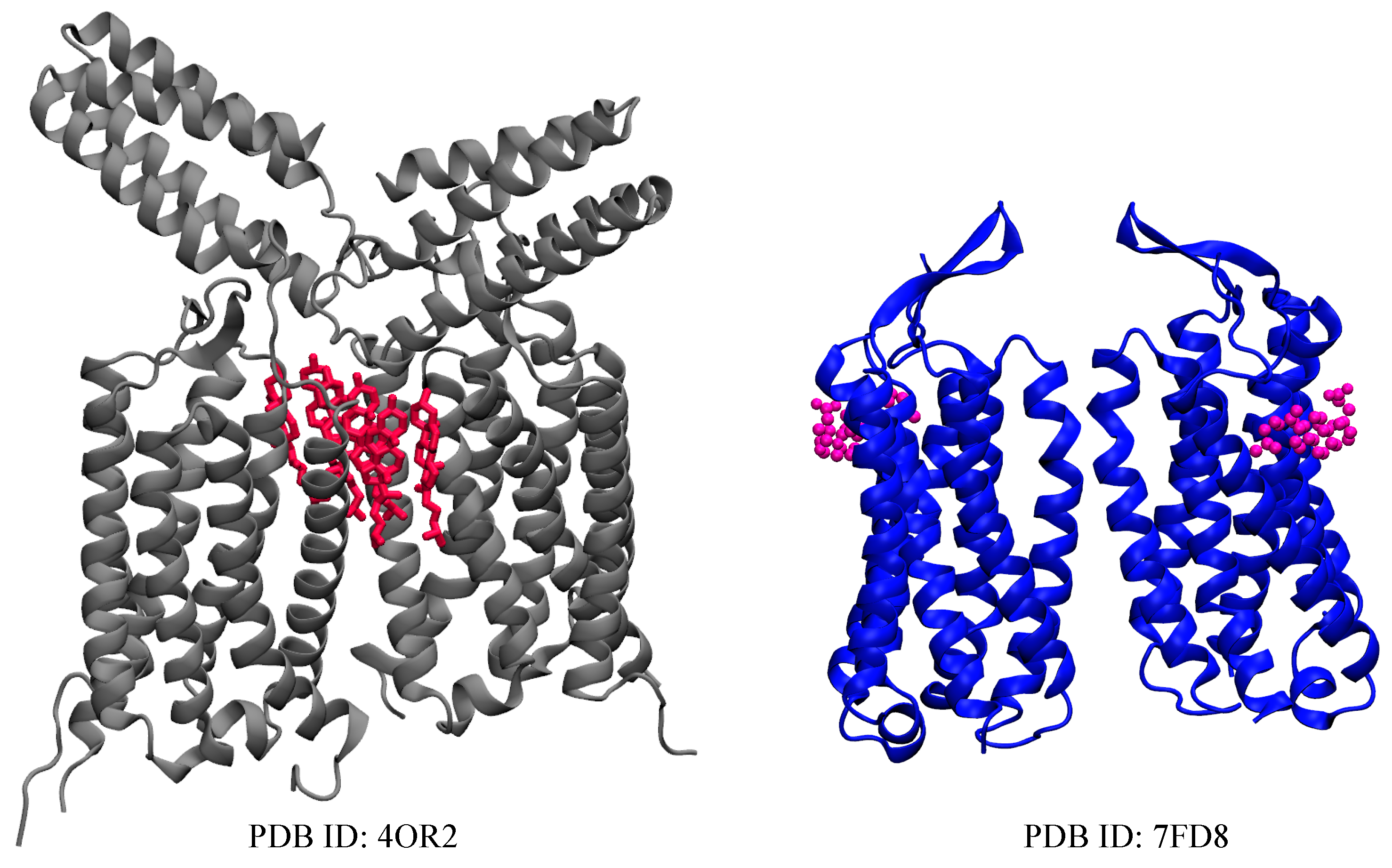
2.1. Metabotropic Glutamate Receptors (mGluRs)
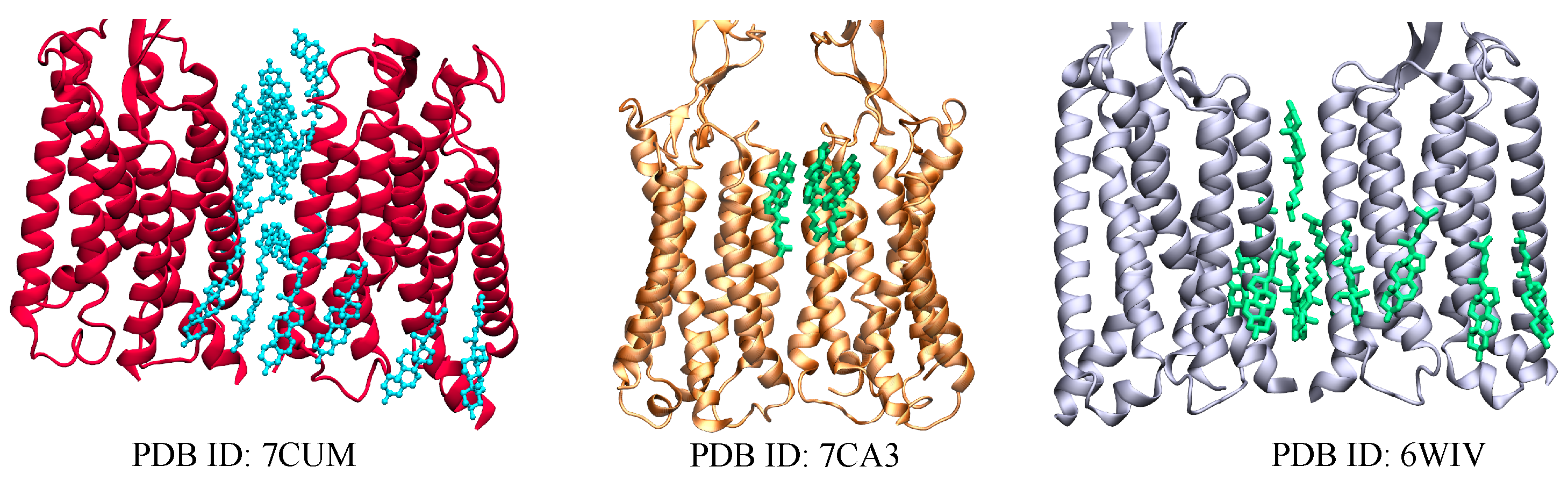
2.2. GABAB Receptors
2.3. Taste Receptor
2.4. Retinoic Acid-Inducible Orphan G Protein-Coupled Receptors (RAIGs)
2.5. Calcium-Sensing Receptor-Related Receptor
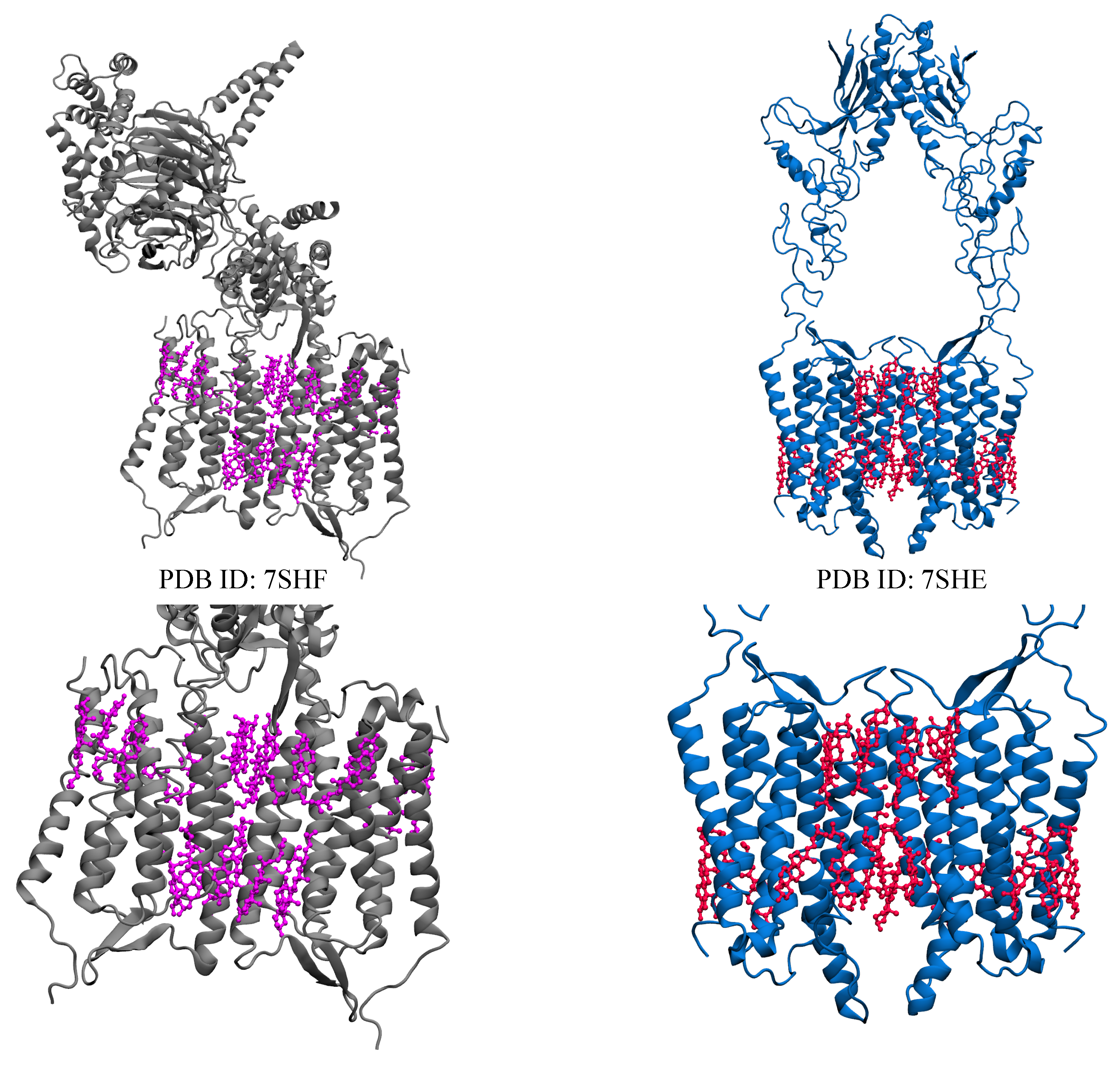
2.6. Orphan Receptor
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GPCR | G-protein-coupled receptors |
| CHS | Cholesteryl hemisuccinate |
| CLR | Cholesterol |
| mGluR | Metabotropic glutamate receptors |
| CRAC | Cholesterol Recognition/Interaction Amino Acid Consensus (CRAC) |
| MD(S) | Molecular dynamics (simulation) |
| GABA | Gamma-Aminobutyric acid |
| TAS1R1 | Taste 1 receptor member 1 |
| TAS1R2 | Taste 1 receptor member 2 |
| TAS1R3 | Taste 1 receptor member 3 |
| RORs | Retinoid-related orphan receptors |
| CASR | calcium-sensing receptor |
| TM(D) | Transmembrane (domain) |
| NTD | N-terminal domain |
| VSMCs | Vascular smooth muscle cells |
| PLC | phospholipase C |
| MMP-2 | Matrix metalloproteinase-2 |
| ERK | Extracellular signal-regulated kinase |
| MAPK | Mitogen-activated protein kinase |
| AMPK | AMP-activated protein kinase |
References
- Goddard, A.D.; Watts, A. Regulation of G protein-coupled receptors by palmitoylation and cholesterol. BMC Biol. 2012, 10, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Fan, Z.; Rovira, X.; Xue, L.; Roux, S.; Brabet, I.; Xin, M.; Pin, J.P.; Rondard, P.; Liu, J. Allosteric ligands control the activation of a class C GPCR heterodimer by acting at the transmembrane interface. eLife 2021, 10, e70188. [Google Scholar] [CrossRef] [PubMed]
- Chun, L.; Zhang, W.H.; Liu, J.F. Structure and ligand recognition of class C GPCRs. Acta Pharmacol. Sin. 2012, 33, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurice, P.; Kamal, M.; Jockers, R. Asymmetry of GPCR oligomers supports their functional relevance. Trends Pharmacol. Sci. 2011, 32, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Soffientini, U.; Graham, A. Intracellular cholesterol transport proteins: Roles in health and disease. Clin. Sci. 2016, 130, 1843–1859. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.A.; Chattopadhyay, A. Statin-Induced Chronic Cholesterol Depletion Switches GPCR Endocytosis and Trafficking: Insights from the Serotonin 1A Receptor. ACS Chem. Neurosci. 2020, 11, 453–465. [Google Scholar] [CrossRef]
- Lee, A.G. Interfacial Binding Sites for Cholesterol on G Protein-Coupled Receptors. Biophys. J. 2019, 116, 1586–1597. [Google Scholar] [CrossRef]
- Zakany, F.; Kovacs, T.; Panyi, G.; Varga, Z. Direct and indirect cholesterol effects on membrane proteins with special focus on potassium channels. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158706. [Google Scholar] [CrossRef]
- Zocher, M.; Zhang, C.; Rasmussen, S.G.; Kobilka, B.K.; Müller, D.J. Cholesterol increases kinetic, energetic, and mechanical stability of the human β2-adrenergic receptor. Proc. Natl. Acad. Sci. USA 2012, 109, E3463–E3472. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Chattopadhyay, A. Molecular dynamics simulations of GPCR-cholesterol interaction: An emerging paradigm. Biochim. Biophys. Acta-(BBA)-Biomembr. 2015, 1848, 1775–1782. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Prasanna, X.; Mohole, M.; Chattopadhyay, A. exploring GPCR-lipid interactions by Molecular dynamic simulation. J. Phys. Chem. B 2018, 122, 5727–5737. [Google Scholar] [CrossRef] [PubMed]
- Hedger, G.; Koldsø, H.; Chavent, M.; Siebold, C.; Rohatgi, R.; Sansom, M.S.P. Cholesterol Interaction Sites on the Transmembrane Domain of the Hedgehog Signal Transducer and Class F G Protein-Coupled Receptor Smoothened. Structure 2019, 27, 549–559.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polasa, A.; Moradi, M. Deciphering the Inter-domain Decoupling in the Gram-negative Bacterial Membrane Insertase. bioRxiv 2022. [Google Scholar] [CrossRef]
- Govind Kumar, V.; Polasa, A.; Agrawal, S.; Kumar, T.K.S.; Moradi, M. Binding affinity estimation from restrained umbrella sampling simulations. Nat. Comput. Sci. 2022, 3, 59–70. [Google Scholar] [CrossRef]
- Polasa, A.; Mosleh, I.; Losey, J.; Abbaspourrad, A.; Beitle, R.; Moradi, M. Developing a rational approach to designing recombinant proteins for peptide-directed nanoparticle synthesis. Nanoscale Adv. 2022, 4, 3161–3171. [Google Scholar] [CrossRef] [PubMed]
- Govind Kumar, V.; Ogden, D.S.; Isu, U.H.; Polasa, A.; Losey, J.; Moradi, M. Prefusion spike protein conformational changes are slower in SARS-CoV-2 than in SARS-CoV-1. J. Biol. Chem. 2022, 298, 101814. [Google Scholar] [CrossRef]
- Polasa, A.; Hettige, J.; Kalyan, I.; Moradi, M. An investigation of the YidC-mediated membrane insertion of Pf3 coat protein using molecular dynamics simulations. Front. Mol. Biosci. 2022, 9, 954262. [Google Scholar] [CrossRef]
- Immadisetty, K.; Moradi, M. Mechanistic Picture for Chemomechanical Coupling in a Bacterial Proton-Coupled Oligopeptide Transporter from Streptococcus Thermophilus. J. Phys. Chem. B 2021, 125, 9738–9750. [Google Scholar] [CrossRef]
- Gorvin, C.M. Calcium-sensing receptor signaling—How human disease informs biology. Curr. Opin. Endocr. Metab. Res. 2021, 16, 10–18. [Google Scholar] [CrossRef]
- Ling, S.; Shi, P.; Liu, S.; Meng, X.; Zhou, Y.; Sun, W.; Chang, S.; Zhang, X.; Zhang, L.; Shi, C.; et al. Structural mechanism of cooperative activation of the human calcium-sensing receptor by Ca2+ ions and L-tryptophan. Cell Res. 2021, 31, 383–394. [Google Scholar] [CrossRef]
- Ray, K. Chapter Six—Calcium-Sensing Receptor: Trafficking, Endocytosis, Recycling, and Importance of Interacting Proteins. In Trafficking of GPCRs; Wu, G., Ed.; Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2015; Volume 132, pp. 127–150. [Google Scholar] [CrossRef]
- Kniazeff, J.; Prézeau, L.; Rondard, P.; Pin, J.P.; Goudet, C. Dimers and beyond: The functional puzzles of class C GPCRs. Pharmacol. Ther. 2011, 130, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Mao, C.; Xu, C.; Jin, N.; Zhang, H.; Shen, D.D.; Shen, Q.; Wang, X.; Hou, T.; Chen, Z.; et al. Structural basis of GABAB receptor—Gi protein coupling. Nature 2021, 594, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Fu, Z.; Frangaj, A.; Liu, J.; Mosyak, L.; Shen, T.; Slavkovich, V.N.; Ray, K.M.; Taura, J.; Cao, B.; et al. Structure of human GABAB receptor in an inactive state. Nature 2020, 584, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Terunuma, M. Diversity of structure and function of GABAB receptors: A complexity of GABAB-mediated signaling. Proc. Jpn. Acad. Ser. B 2018, 94, 390–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaye, H.; Ishchenko, A.; Lam, J.H.; Han, G.W.; Xue, L.; Rondard, P.; Pin, J.P.; Katritch, V.; Gati, C.; Cherezov, V. Structural basis of the activation of a metabotropic GABA receptor. Nature 2020, 584, 298–303. [Google Scholar] [CrossRef]
- Vafabakhsh, R.; Levitz, J.; Isacoff, E.Y. Conformational dynamics of a class C G-protein-coupled receptor. Nature 2015, 524, 497–501. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.Y.; Kim, S.K.; Goddard, W.A. G protein coupling and activation of the metabotropic GABAB heterodimer. Nat. Commun. 2022, 13, 4612. [Google Scholar] [CrossRef]
- Shaye, H.; Stauch, B.; Gati, C.; Cherezov, V. Molecular mechanisms of metabotropic GABAB receptor function. Sci. Adv. 2021, 7, eabg3362. [Google Scholar] [CrossRef]
- Geng, Y.; Bush, M.; Mosyak, L.; Wang, F.; Fan, Q.R. Structural mechanism of ligand activation in human GABAB receptor. Nature 2013, 504, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, Y.; Gao, Y. Chapter Four—Asymmetric activation of class C GPCRs. In G Protein-Coupled Receptors—Part B; Shukla, A.K., Ed.; Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2023; Volume 195, pp. 77–87. [Google Scholar] [CrossRef]
- Ellaithy, A.; Gonzalez-Maeso, J.; Logothetis, D.A.; Levitz, J. Structural and Biophysical Mechanisms of Class C G Protein-Coupled Receptor Function. Trends Biochem. Sci. 2020, 45, 1049–1064. [Google Scholar] [CrossRef]
- McCullock, T.W.; Kammermeier, P.J. The evidence for and consequences of metabotropic glutamate receptor heterodimerization. Neuropharmacology 2021, 199, 108801. [Google Scholar] [CrossRef] [PubMed]
- Møller, T.C.; Moreno-Delgado, D.; Pin, J.P.; Kniazeff, J. Class CG protein-coupled receptors: Reviving old couples with new partners. Biophys. Rep. 2017, 3, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salon, J.A.; Lodowski, D.T.; Palczewski, K. The Significance of G Protein-Coupled Receptor Crystallography for Drug Discovery. Pharmacol. Rev. 2011, 63, 901–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Wang, C.; Gregory, K.J.; Han, G.W.; Cho, H.P.; Xia, Y.; Niswender, C.M.; Katritch, V.; Meiler, J.; Cherezov, V.; et al. Structure of a class C GPCR metabotropic glutamate receptor 1 bound to an allosteric modulator. Science 2014, 344, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Zhou, Q.; Labroska, V.; Qin, S.; Darbalaei, S.; Wu, Y.; Yuliantie, E.; Xie, L.; Tao, H.; Cheng, J.; et al. G protein-coupled receptors: Structure-and function-based drug discovery. Signal Transduct. Target. Ther. 2021, 6, 7. [Google Scholar] [CrossRef]
- Gómez-Santacana, X.; Panarello, S.; Rovira, X.; Llebaria, A. Photoswitchable allosteric modulators for metabotropic glutamate receptors. Curr. Opin. Pharmacol. 2022, 66, 102266. [Google Scholar] [CrossRef]
- Orgován, Z.; Ferenczy, G.G.; Keserű, G.M. Allosteric Molecular Switches in Metabotropic Glutamate Receptors. ChemMedChem 2021, 16, 81–93. [Google Scholar] [CrossRef]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295. [Google Scholar] [CrossRef] [Green Version]
- Doumazane, E.; Scholler, P.; Zwier, J.M.; Trinquet, E.; Rondard, P.; Pin, J.P. A new approach to analyze cell surface protein complexes reveals specific heterodimeric metabotropic glutamate receptors. FASEB J. 2011, 25, 66–77. [Google Scholar] [CrossRef]
- Seven, A.B.; Barros-Álvarez, X.; de Lapeyrière, M.; Papasergi-Scott, M.M.; Robertson, M.J.; Zhang, C.; Nwokonko, R.M.; Gao, Y.; Meyerowitz, J.G.; Rocher, J.P.; et al. G-protein activation by a metabotropic glutamate receptor. Nature 2021, 595, 450–454. [Google Scholar] [CrossRef]
- Fang, W.; Yang, F.; Xu, C.; Ling, S.; Lin, L.; Zhou, Y.; Sun, W.; Wang, X.; Liu, P.; Rondard, P.; et al. Structural basis of the activation of metabotropic glutamate receptor 3. Cell Res. 2022, 32, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Han, S.; Cai, X.; Tan, Q.; Zhou, K.; Wang, D.; Wang, X.; Du, J.; Yi, C.; Chu, X.; et al. Structures of Gi-bound metabotropic glutamate receptors mGlu2 and mGlu4. Nature 2021, 594, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, C.; Cannone, G.; Briot, J.; Rottier, K.; Berizzi, A.E.; Huang, C.Y.; Quast, R.B.; Hoh, F.; Banères, J.L.; Malhaire, F.; et al. Agonists and allosteric modulators promote signaling from different metabotropic glutamate receptor 5 conformations. Cell Rep. 2021, 36, 109648. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, D.; Fan, H.; Xu, C.; Tai, L.; Lin, S.; Han, S.; Tan, Q.; Wang, X.; Xu, T.; et al. Structures of human mGlu2 and mGlu7 homo- and heterodimers. Nature 2021, 594, 589–593. [Google Scholar] [CrossRef]
- Frangaj, A.; Fan, Q.R. Structural biology of GABAB receptor. Neuropharmacology 2018, 136, 68–79. [Google Scholar] [CrossRef]
- Schamber, M.R.; Vafabakhsh, R. Mechanism of sensitivity modulation in the calcium-sensing receptor via electrostatic tuning. Nat. Commun. 2022, 13, 2194. [Google Scholar] [CrossRef]
- Wen, T.; Wang, Z.; Chen, X.; Ren, Y.; Lu, X.; Xing, Y.; Lu, J.; Chang, S.; Zhang, X.; Shen, Y.; et al. Structural basis for activation and allosteric modulation of full-length calcium-sensing receptor. Sci. Adv. 2021, 7, eabg1483. [Google Scholar] [CrossRef]
- Robbins, M.J.; Michalovich, D.; Hill, J.; Calver, A.R.; Medhurst, A.D.; Gloger, I.; Sims, M.; Middlemiss, D.N.; Pangalos, M.N. Molecular Cloning and Characterization of Two Novel Retinoic Acid-Inducible Orphan G-Protein-Coupled Receptors (GPRC5B and GPRC5C). Genomics 2000, 67, 8–18. [Google Scholar] [CrossRef]
- Mafi, A.; Kim, S.K.; Chou, K.C.; Güthrie, B.; Goddard, W.A.I.I.I. Predicted Structure of Fully Activated Tas1R3/1R3 Homodimer Bound to G Protein and Natural Sugars: Structural Insights into G Protein Activation by a Class C Sweet Taste Homodimer with Natural Sugars. J. Am. Chem. Soc. 2021, 143, 16824–16838. [Google Scholar] [CrossRef]
- Belloir, C.; Brulé, M.; Tornier, L.; Neiers, F.; Briand, L. Biophysical and functional characterization of the human TAS1R2 sweet taste receptor overexpressed in a HEK293S inducible cell line. Sci. Rep. 2021, 11, 22238. [Google Scholar] [CrossRef]
- Ahmad, R.; Dalziel, J.E. G Protein-Coupled Receptors in Taste Physiology and Pharmacology. Front. Pharmacol. 2020, 11, 587664. [Google Scholar] [CrossRef] [PubMed]
- Nuemket, N.; Yasui, N.; Kusakabe, Y.; Nomura, Y.; Atsumi, N.; Akiyama, S.; Nango, E.; Kato, Y.; Kaneko, M.K.; Takagi, J.; et al. Structural basis for perception of diverse chemical substances by T1r taste receptors. Nat. Commun. 2017, 8, 15530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, D.N.; Singh, S.; Laboute, T.; Strutzenberg, T.S.; Qiu, X.; Wu, D.; Novick, S.J.; Robinson, C.V.; Griffin, P.R.; Hunt, J.F.; et al. Cryo-EM structure of human GPR158 receptor coupled to the RGS7-Gβ5 signaling complex. Science 2022, 375, 86–91. [Google Scholar] [CrossRef]
- Darira, S.V.; Sutton, L.P. Chapter Seven—The interaction, mechanism and function of GPR158-RGS7 cross-talk. In G Protein-Coupled Receptors—Part A; Shukla, A.K., Ed.; Progress in Molecular Biology and Translational Science; Academic Press: Cambridge, MA, USA, 2022; Volume 193, pp. 167–176. [Google Scholar] [CrossRef]
- Kindt, K.S.; Akturk, A.; Jarysta, A.; Day, M.; Beirl, A.; Flonard, M.; Tarchini, B. EMX2-GPR156-Gαi reverses hair cell orientation in mechanosensory epithelia. Nat. Commun. 2021, 12, 2861. [Google Scholar] [CrossRef]
- Jørgensen, C.V.; Bräuner-Osborne, H. Pharmacology and physiological function of the orphan GPRC6A receptor. Basic Clin. Pharmacol. Toxicol. 2020, 126, 77–87. [Google Scholar] [CrossRef]
- Orlandi, C.; Cao, Y.; Martemyanov, K.A. Orphan Receptor GPR179 Forms Macromolecular Complexes with Components of Metabotropic Signaling Cascade in Retina ON-Bipolar Neurons. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7153–7161. [Google Scholar] [CrossRef] [Green Version]
- Guixà-González, R.; Albasanz, J.L.; Rodriguez-Espigares, I.; Pastor, M.; Sanz, F.; Martí-Solano, M.; Manna, M.; Martinez-Seara, H.; Hildebrand, P.W.; Martín, M.; et al. Membrane cholesterol access into a G-protein-coupled receptor. Nat. Commun. 2017, 8, 14505. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, X.; Mohole, M.; Chattopadhyay, A.; Sengupta, D. Role of cholesterol-mediated effects in GPCR heterodimers. Chem. Phys. Lipids 2020, 227, 104852. [Google Scholar] [CrossRef]
- Jakubík, J.; El-Fakahany, E.E. Allosteric Modulation of GPCRs of Class A by Cholesterol. Int. J. Mol. Sci. 2021, 22, 1953. [Google Scholar] [CrossRef]
- Harkey, T.; Govind Kumar, V.; Hettige, J.; Tabari, S.H.; Immadisetty, K.; Moradi, M. The Role of a Crystallographically Unresolved Cytoplasmic Loop in Stabilizing the Bacterial Membrane Insertase YidC2. Sci. Rep. 2019, 9, 14451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Jeong, E.; Jeong, J.H.; Kim, Y.; Cho, Y. Structural Basis for Activation of the Heterodimeric GABAB Receptor. J. Mol. Biol. 2020, 432, 5966–5984. [Google Scholar] [CrossRef]
- Park, J.; Zuo, H.; Frangaj, A.; Fu, Z.; Yen, L.Y.; Zhang, Z.; Mosyak, L.; Slavkovich, V.N.; Liu, J.; Ray, K.M.; et al. Symmetric activation and modulation of the human calcium-sensing receptor. Proc. Natl. Acad. Sci. 2021, 118, e2115849118. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Barrantes, F. How cholesterol interacts with membrane proteins: An exploration of cholesterol-binding sites including CRAC, CARC, and tilted domains. Front. Physiol. 2013, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dingjan, T.; Futerman, A.H. The fine-tuning of cell membrane lipid bilayers accentuates their compositional complexity. BioEssays 2021, 43, 2100021. [Google Scholar] [CrossRef]
- Pinkwart, K.; Schneider, F.; Lukoseviciute, M.; Sauka-Spengler, T.; Lyman, E.; Eggeling, C.; Sezgin, E. Nanoscale dynamics of cholesterol in the cell membrane. J. Biol. Chem. 2019, 294, 12599–12609. [Google Scholar] [CrossRef] [Green Version]
- Pucadyil, T.J.; Chattopadhyay, A. Role of cholesterol in the function and organization of G-protein coupled receptors. Prog. Lipid Res. 2006, 45, 295–333. [Google Scholar] [CrossRef]
- Arora, A.; Raghuraman, H.; Chattopadhyay, A. Influence of cholesterol and ergosterol on membrane dynamics: A fluorescence approach. Biochem. Biophys. Res. Commun. 2004, 318, 920–926. [Google Scholar] [CrossRef]
- Chimento, A.; Casaburi, I.; Avena, P.; Trotta, F.; De Luca, A.; Rago, V.; Pezzi, V.; Sirianni, R. Cholesterol and Its Metabolites in Tumor Growth: Therapeutic Potential of Statins in Cancer Treatment. Front. Endocrinol. 2019, 9, 807. [Google Scholar] [CrossRef] [Green Version]
- Ohvo-Rekilä, H.; Ramstedt, B.; Leppimäki, P.; Peter Slotte, J. Cholesterol interactions with phospholipids in membranes. Prog. Lipid Res. 2002, 41, 66–97. [Google Scholar] [CrossRef]
- McMullen, T.P.W.; Lewis, R.N.A.H.; McElhaney, R.N. Cholesterol–phospholipid interactions, the liquid-ordered phase and lipid rafts in model and biological membranes. Curr. Opin. Colloid Interface Sci. 2004, 8, 459–468. [Google Scholar] [CrossRef]
- Gater, D.L.; Réat, V.; Czaplicki, G.; Saurel, O.; Milon, A.; Jolibois, F.; Cherezov, V. Hydrogen Bonding of Cholesterol in the Lipidic Cubic Phase. Langmuir 2013, 29, 8031–8038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowlands, L.J.; Marks, A.; Sanderson, J.M.; Law, R.V. 17O NMR spectroscopy as a tool to study hydrogen bonding of cholesterol in lipid bilayers. Chem. Commun. 2020, 56, 14499–14502. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.G.W.; Jacobson, K. A Role for Lipid Shells in Targeting Proteins to Caveolae, Rafts, and Other Lipid Domains. Science 2002, 296, 1821–1825. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Yahi, N. Molecular insights into amyloid regulation by membrane cholesterol and sphingolipids: Common mechanisms in neurodegenerative diseases. Expert Rev. Mol. Med. 2010, 12, e27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantini, J.; Garmy, N.; Mahfoud, R.; Yahi, N. Lipid rafts: Structure, function and role in HIV, Alzheimer’s and prion diseases. Expert Rev. Mol. Med. 2002, 4, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.T.; Kreutzberger, A.J.B.; Lee, J.; Kiessling, V.; Tamm, L.K. The role of cholesterol in membrane fusion. Chem. Phys. Lipids 2016, 199, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Kenworthy, A.K.; Sanders, C.R. Cholesterol as a co-solvent and a ligand for membrane proteins. Protein Sci. 2014, 23, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Najafinobar, N.; Mellander, L.J.; Kurczy, M.E.; Dunevall, J.; Angerer, T.B.; Fletcher, J.S.; Cans, A.S. Cholesterol Alters the Dynamics of Release in Protein Independent Cell Models for Exocytosis. Sci. Rep. 2016, 6, 33702. [Google Scholar] [CrossRef] [Green Version]
- Yeagle, P.L. Modulation of membrane function by cholesterol. Biochimie 1991, 73, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Peränen, J.; Schreiner, R.; Wieland, F.; Kurzchalia, T.V.; Simons, K. VIP21/caveolin is a cholesterol-binding protein. Proc. Natl. Acad. Sci. USA 1995, 92, 10339–10343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, J.A.; Young, K.E.; Beachy, P.A. Cholesterol Modification of Hedgehog Signaling Proteins in Animal Development. Science 1996, 274, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Sutter, M.L.; Console, L.; Fahner, A.F.; Samodelov, S.L.; Gai, Z.; Ciarimboli, G.; Indiveri, C.; Kullak-Ublick, G.A.; Visentin, M. The role of cholesterol recognition (CARC/CRAC) mirror codes in the allosterism of the human organic cation transporter 2 (OCT2, SLC22A2). Biochem. Pharmacol. 2021, 194, 114840. [Google Scholar] [CrossRef] [PubMed]
- Taghon, G.; Rowe, J.; Kapolka, N.; Isom, D. Predictable cholesterol binding sites in GPCRs lack consensus motifs. Structure 2021, 29, 499–506.e3. [Google Scholar] [CrossRef] [PubMed]
- de Vries, M.; Herrmann, A.; Veit, M. A cholesterol consensus motif is required for efficient intracellular transport and raft association of a group 2 HA of influenza virus. Biochem. J. 2014, 465, 305–314. [Google Scholar] [CrossRef]
- Kuwabara, P.E.; Labouesse, M. The sterol-sensing domain: Multiple families, a unique role? Trends Genet. 2002, 18, 193–201. [Google Scholar] [CrossRef]
- Martín, V.; Carrillo, G.; Torroja, C.; Guerrero, I. The sterol-sensing domain of Patched protein seems to control Smoothened activity through Patched vesicular trafficking. Curr. Biol. CB 2001, 11, 601–607. [Google Scholar] [CrossRef] [Green Version]
- Jafurulla, M.; Tiwari, S.; Chattopadhyay, A. Identification of cholesterol recognition amino acid consensus (CRAC) motif in G-protein coupled receptors. Biochem. Biophys. Res. Commun. 2011, 404, 569–573. [Google Scholar] [CrossRef]
- Sarkar, P.; Chattopadhyay, A. Cholesterol in GPCR Structures: Prevalence and Relevance. J. Membr. Biol. 2022, 255, 99–106. [Google Scholar] [CrossRef]
- Epand, R.M. Cholesterol and the interaction of proteins with membrane domains. Prog. Lipid Res. 2006, 45, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yao, Z.x.; Degenhardt, B.; Teper, G.; Papadopoulos, V. Cholesterol binding at the cholesterol recognition/ interaction amino acid consensus (CRAC) of the peripheral-type benzodiazepine receptor and inhibition of steroidogenesis by an HIV TAT-CRAC peptide. Proc. Natl. Acad. Sci. USA 2001, 98, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Mozumder, S.; Bej, A.; Mukherjee, S.; Sengupta, J.; Chattopadhyay, A. Structure, dynamics and lipid interactions of serotonin receptors: Excitements and challenges. Biophys. Rev. 2021, 13, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Paila, Y.D.; Jafurulla, M.; Chattopadhyay, A. Membrane cholesterol depletion from live cells enhances the function of human serotonin1A receptors. Biochem. Biophys. Res. Commun. 2009, 389, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Di Scala, C.; Evans, L.S.; Williamson, P.T.F.; Barrantes, F.J. A mirror code for protein–cholesterol interactions in the two leaflets of biological membranes. Sci. Rep. 2016, 6, 21907. [Google Scholar] [CrossRef] [Green Version]
- Di Scala, C.; Baier, C.; Evans, L.; Williamson, P.; Fantini, J.; Barrantes, F. Relevance of CARC and CRAC Cholesterol-Recognition Motifs in the Nicotinic Acetylcholine Receptor and Other Membrane-Bound Receptors. Curr. Top. Membr. 2017, 80, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Azzaz, F.; Chahinian, H.; Yahi, N.; Di Scala, C.; Baier, C.J.; Barrantes, F.J.; Fantini, J. Chapter 7—Cholesterol-recognizing amino acid consensus motifs in transmembrane proteins: Comparative analysis of in silico studies and structural data. In Cholesterol; Bukiya, A.N., Dopico, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 127–145. [Google Scholar] [CrossRef]
- Hanson, M.A.; Cherezov, V.; Griffith, M.T.; Roth, C.B.; Jaakola, V.P.; Chien, E.Y.; Velasquez, J.; Kuhn, P.; Stevens, R.C. A Specific Cholesterol Binding Site Is Established by the 2.8 Å Structure of the Human β2-Adrenergic Receptor. Structure 2008, 16, 897–905. [Google Scholar] [CrossRef] [Green Version]
- McGraw, C.; Koretz, K.S.; Oseid, D.; Lyman, E.; Robinson, A.S. Cholesterol Dependent Activity of the Adenosine A2A Receptor Is Modulated via the Cholesterol Consensus Motif. Molecules 2022, 27, 3529. [Google Scholar] [CrossRef]
- Desai, A.; Miller, L. Sensitivity of cholecystokinin receptors to membrane cholesterol content. Front. Endocrinol. 2012, 3, 123. [Google Scholar] [CrossRef] [Green Version]
- Adamian, L.; Naveed, H.; Liang, J. Lipid-binding surfaces of membrane proteins: Evidence from evolutionary and structural analysis. Biochim. Biophys. Acta (BBA) Biomembr. 2011, 1808, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Yabe, D.; Xia, Z.P.; Adams, C.M.; Rawson, R.B. Three mutations in sterol-sensing domain of SCAP block interaction with insig and render SREBP cleavage insensitive to sterols. Proc. Natl. Acad. Sci. USA 2002, 99, 16672–16677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Espenshade, P.J.; Wright, M.E.; Yabe, D.; Gong, Y.; Aebersold, R.; Goldstein, J.L.; Brown, M.S. Crucial Step in Cholesterol Homeostasis: Sterols Promote Binding of SCAP to INSIG-1, a Membrane Protein that Facilitates Retention of SREBPs in ER. Cell 2002, 110, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukiya, A.N.; Dopico, A.M. Common structural features of cholesterol binding sites in crystallized soluble proteins. J. Lipid Res. 2017, 58, 1044–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rone, M.B.; Fan, J.; Papadopoulos, V. Cholesterol transport in steroid biosynthesis: Role of protein–protein interactions and implications in disease states. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2009, 1791, 646–658. [Google Scholar] [CrossRef] [Green Version]
- Naito, T.; Ercan, B.; Krshnan, L.; Triebl, A.; Koh, D.H.Z.; Wei, F.Y.; Tomizawa, K.; Torta, F.T.; Wenk, M.R.; Saheki, Y. Movement of accessible plasma membrane cholesterol by the GRAMD1 lipid transfer protein complex. eLife 2019, 8, e51401. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Hua, T.; Liu, Z.J.; Liu, H.; Zhao, S. MD Simulations Revealing Special Activation Mechanism of Cannabinoid Receptor 1. Front. Mol. Biosci. 2022, 9, 860035. [Google Scholar] [CrossRef]
- Nieto Gutierrez, A.; McDonald, P.H. GPCRs: Emerging anti-cancer drug targets. Cell. Signal. 2018, 41, 65–74. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- Sriram, K.; Insel, P.A. GPCRs as targets for approved drugs: How many targets and how many drugs? Mol. Pharmacol. 2018, 93, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Niswender, C.M.; Jones, C.K.; Lin, X.; Bubser, M.; Thompson Gray, A.; Blobaum, A.L.; Engers, D.W.; Rodriguez, A.L.; Loch, M.T.; Daniels, J.S.; et al. Development and antiparkinsonian activity of VU0418506, a selective positive allosteric modulator of metabotropic glutamate receptor 4 homomers without activity at mGlu2/4 heteromers. ACS Chem. Neurosci. 2016, 7, 1201–1211. [Google Scholar] [CrossRef] [Green Version]
- Moreau, C.J.; Audic, G.; Lemel, L.; García-Fernández, M.D.; Nieścierowicz, K. Interactions of cholesterol molecules with GPCRs in different states: A comparative analysis of GPCRs’ structures. Biochim. Biophys. Acta (BBA) Biomembr. 2023, 1865, 184100. [Google Scholar] [CrossRef] [PubMed]
- Salas-Estrada, L.A.; Leioatts, N.; Romo, T.D.; Grossfield, A. Lipids Alter Rhodopsin Function via Ligand-like and Solvent-like Interactions. Biophys. J. 2018, 114, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimpl, G.; Fahrenholz, F. Cholesterol as stabilizer of the oxytocin receptor. Biochim. Biophys. Acta (BBA) Biomembr. 2002, 1564, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Marino, K.A.; Prada-Gracia, D.; Provasi, D.; Filizola, M. Impact of Lipid Composition and Receptor Conformation on the Spatio-temporal Organization of μ-Opioid Receptors in a Multi-component Plasma Membrane Model. PLoS Comput. Biol. 2016, 12, e1005240. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Chattopadhyay, A. Identification of Cholesterol Binding Sites in the Serotonin1A Receptor. J. Phys. Chem. B 2012, 116, 12991–12996. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, G.; Sircar, R.; Kong, J.H.; Nachtergaele, S.; Sagner, A.; Byrne, E.F.; Covey, D.F.; Siebold, C.; Rohatgi, R. Cholesterol activates the G-protein coupled receptor smoothened to promote hedgehog signaling. eLife 2016, 5, e20304. [Google Scholar] [CrossRef]
- Geiger, J.; Sexton, R.; Al-Sahouri, Z.; Lee, M.Y.; Chun, E.; Harikumar, K.G.; Miller, L.J.; Beckstein, O.; Liu, W. Evidence that specific interactions play a role in the cholesterol sensitivity of G protein-coupled receptors. Biochim. Biophys. Acta (BBA) Biomembr. 2021, 1863, 183557. [Google Scholar] [CrossRef] [PubMed]
- Paila, Y.D.; Chattopadhyay, A. The function of G-protein coupled receptors and membrane cholesterol: Specific or general interaction? Glycoconj. J. 2009, 26, 711–720. [Google Scholar] [CrossRef]
- Augustyn, B.; Stepien, P.; Poojari, C.; Mobarak, E.; Polit, A.; Wisniewska-Becker, A.; Róg, T. Cholesteryl Hemisuccinate Is Not a Good Replacement for Cholesterol in Lipid Nanodiscs. J. Phys. Chem. B 2019, 123, 9839–9845. [Google Scholar] [CrossRef]
- Kulig, W.; Tynkkynen, J.; Javanainen, M.; Manna, M.; Rog, T.; Vattulainen, I.; Jungwirth, P. How well does cholesteryl hemisuccinate mimic cholesterol in saturated phospholipid bilayers? J. Mol. Model. 2014, 20, 2121. [Google Scholar] [CrossRef]
- Kulig, W.; Jurkiewicz, P.; Olżyńska, A.; Tynkkynen, J.; Javanainen, M.; Manna, M.; Rog, T.; Hof, M.; Vattulainen, I.; Jungwirth, P. Experimental determination and computational interpretation of biophysical properties of lipid bilayers enriched by cholesteryl hemisuccinate. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 422–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, P.; Bhat, A.; Chattopadhyay, A. Lysine 101 in the CRAC Motif in Transmembrane Helix 2 Confers Cholesterol-Induced Thermal Stability to the Serotonin1A Receptor. J. Membr. Biol. 2022, 255, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, X.; Chattopadhyay, A.; Sengupta, D. Cholesterol modulates the dimer interface of the β2-adrenergic receptor via cholesterol occupancy sites. Biophys. J. 2014, 106, 1290–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oddi, S.; Dainese, E.; Fezza, F.; Lanuti, M.; Barcaroli, D.; De Laurenzi, V.; Centonze, D.; Maccarrone, M. Functional characterization of putative cholesterol binding sequence (CRAC) in human type-1 cannabinoid receptor. J. Neurochem. 2011, 116, 858–865. [Google Scholar] [CrossRef]
- Oddi, S.; Dainese, E.; Sandiford, S.; Fezza, F.; Lanuti, M.; Chiurchiù, V.; Totaro, A.; Catanzaro, G.; Barcaroli, D.; De Laurenzi, V.; et al. Effects of palmitoylation of Cys 415 in helix 8 of the CB 1 cannabinoid receptor on membrane localization and signaling. Br. J. Pharmacol. 2012, 165, 2635–2651. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, P.; Chattopadhyay, A. Cholesterol interaction motifs in G protein-coupled receptors: Slippery hot spots? Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1481. [Google Scholar] [CrossRef]
- Mazzitelli, M.; Presto, P.; Antenucci, N.; Meltan, S.; Neugebauer, V. Recent Advances in the Modulation of Pain by the Metabotropic Glutamate Receptors. Cells 2022, 11, 2608. [Google Scholar] [CrossRef]
- Kroon, T.; Dawitz, J.; Kramvis, I.; Anink, J.; Obermayer, J.; Verhoog, M.B.; Wilbers, R.; Goriounova, N.A.; Idema, S.; Baayen, J.C.; et al. Group I mGluR-mediated activation of martinotti cells inhibits local cortical circuitry in human cortex. Front. Cell. Neurosci. 2019, 13, 315. [Google Scholar] [CrossRef] [Green Version]
- Pin, J.P.; De Colle, C.; Bessis, A.S.; Acher, F. New perspectives for the development of selective metabotropic glutamate receptor ligands. Eur. J. Pharmacol. 1999, 375, 277–294. [Google Scholar] [CrossRef]
- Sejdiu, B.I.; Tieleman, D.P. Lipid-protein interactions are a unique property and defining feature of G protein-coupled receptors. Biophys. J. 2020, 118, 1887–1900. [Google Scholar] [CrossRef]
- Kurth, M.; Lolicato, F.; Sandoval, A.; Amaya-Espinosa, H.; Teslenko, A.; Sinning, I.; Beck, R.; Brügger, B.; Aponte-Santamaria, C. Cholesterol Localization Around the Metabotropic Glutamate Receptor 2. J. Phys. Chem. B 2020, 124, 9061–9078. [Google Scholar] [CrossRef] [PubMed]
- Liauw, B.W.H.; Foroutan, A.; Schamber, M.R.; Lu, W.; Samareh Afsari, H.; Vafabakhsh, R. Conformational fingerprinting of allosteric modulators in metabotropic glutamate receptor 2. eLife 2022, 11, e78982. [Google Scholar] [CrossRef] [PubMed]
- Bordi, F.; Ugolini, A. Group I metabotropic glutamate receptors: Implications for brain diseases. Prog. Neurobiol. 1999, 59, 55–79. [Google Scholar] [CrossRef] [PubMed]
- Liauw, B.W.H.; Afsari, H.S.; Vafabakhsh, R. Conformational rearrangement during activation of a metabotropic glutamate receptor. Nat. Chem. Biol. 2021, 17, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Yeagle, P. The biophysics and cell biology of cholesterol: An hypothesis for the essential role of cholesterol in mammalian cells. In Cholesterol in Membrane Models; CRC Press: Boca Raton, FL, USA, 1993; pp. 1–12. [Google Scholar]
- Sooksawate, T.; Simmonds, M. Influence of membrane cholesterol on modulation of the GABAA receptor by neuroactive steroids and other potentiators. Br. J. Pharmacol. 2001, 134, 1303–1311. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, F.; Jefferson, J.R.; Kier, A.B.; Knittel, J.; Scallen, T.J.; Wood, W.G.; Hapala, I. Membrane cholesterol dynamics: Cholesterol domains and kinetic pools. Proc. Soc. Exp. Biol. Med. 1991, 196, 235–252. [Google Scholar] [CrossRef]
- Marlow, B.; Kuenze, G.; Li, B.; Sanders, C.R.; Meiler, J. Structural determinants of cholesterol recognition in helical integral membrane proteins. Biophys. J. 2021, 120, 1592–1604. [Google Scholar] [CrossRef]
- Kumari, R.; Castillo, C.; Francesconi, A. Agonist-dependent signaling by group I metabotropic glutamate receptors is regulated by association with lipid domains. J. Biol. Chem. 2013, 288, 32004–32019. [Google Scholar] [CrossRef] [Green Version]
- Lei, T.; Hu, Z.; Ding, R.; Chen, J.; Li, S.; Zhang, F.; Pu, X.; Zhao, N. Exploring the Activation Mechanism of a Metabotropic Glutamate Receptor Homodimer via Molecular Dynamics Simulation. ACS Chem. Neurosci. 2020, 11, 133–145. [Google Scholar] [CrossRef]
- Thibado, J.K.; Tano, J.Y.; Lee, J.; Salas-Estrada, L.; Provasi, D.; Strauss, A.; Marcelo Lamim Ribeiro, J.; Xiang, G.; Broichhagen, J.; Filizola, M.; et al. Differences in interactions between transmembrane domains tune the activation of metabotropic glutamate receptors. eLife 2021, 10, e67027. [Google Scholar] [CrossRef]
- Francesconi, A.; Kumari, R.; Zukin, R.S. Regulation of group I metabotropic glutamate receptor trafficking and signaling by the caveolar/lipid raft pathway. J. Neurosci. 2009, 29, 3590–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caraci, F.; Nicoletti, F.; Copani, A. Metabotropic glutamate receptors: The potential for therapeutic applications in Alzheimer’s disease. Curr. Opin. Pharmacol. 2018, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, F.; Bruno, V.; Ngomba, R.T.; Gradini, R.; Battaglia, G. Metabotropic glutamate receptors as drug targets: What’s new? Curr. Opin. Pharmacol. 2015, 20, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, J.Y.; Wall, B.A.; Wangari-Talbot, J.; Chen, S. Metabotropic glutamate receptors in cancer. Neuropharmacology 2017, 115, 193–202. [Google Scholar]
- Immadisetty, K.; Hettige, J.; Moradi, M. What Can and Cannot Be Learned from Molecular Dynamics Simulations of Bacterial Proton-Coupled Oligopeptide Transporter GkPOT? J. Phys. Chem. B 2017, 121, 3644–3656. [Google Scholar] [CrossRef]
- Moradi, M.; Babin, V.; Roland, C.; Sagui, C. The Adaptively Biased Molecular Dynamics method revisited: New capabilities and an application. J. Phys. Conf. Ser. 2015, 640, 12020. [Google Scholar] [CrossRef]
- Govind Kumar, V.; Agrawal, S.; Kumar, T.K.S.; Moradi, M. Mechanistic Picture for Monomeric Human Fibroblast Growth Factor 1 Stabilization by Heparin Binding. J. Phys. Chem. B 2021, 125, 12690–12697. [Google Scholar] [CrossRef]
- Baier, C.J.; Fantini, J.; Barrantes, F.J. Disclosure of cholesterol recognition motifs in transmembrane domains of the human nicotinic acetylcholine receptor. Sci. Rep. 2011, 1, 69. [Google Scholar] [CrossRef] [Green Version]
- Bruno, A.; Costantino, G.; de Fabritiis, G.; Pastor, M.; Selent, J. Membrane-sensitive conformational states of helix 8 in the metabotropic Glu2 receptor, a class C GPCR. PLoS ONE 2012, 7, e42023. [Google Scholar] [CrossRef]
- Nutt, D.J.; Malizia, A.L. New insights into the role of the GABAA–benzodiazepine receptor in psychiatric disorder. Br. J. Psychiatry 2001, 179, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Burmakina, S.; Geng, Y.; Chen, Y.; Fan, Q.R. Heterodimeric coiled-coil interactions of human GABAB receptor. Proc. Natl. Acad. Sci. USA 2014, 111, 6958–6963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassetti, D. Keeping the Balance: GABAB Receptors in the Developing Brain and Beyond. Brain Sci. 2022, 12, 419. [Google Scholar] [CrossRef]
- Sooksawate, T.; Simmonds, M. Effects of membrane cholesterol on the sensitivity of the GABAA receptor to GABA in acutely dissociated rat hippocampal neurones. Neuropharmacology 2001, 40, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Hoon, M.A.; Adler, E.; Lindemeier, J.; Battey, J.F.; Ryba, N.J.; Zuker, C.S. Putative mammalian taste receptors: A class of taste-specific GPCRs with distinct topographic selectivity. Cell 1999, 96, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Vigues, S.; Dotson, C.D.; Munger, S.D. The receptor basis of sweet taste in mammals. Results Probl. Cell Differ. 2009, 47, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Prasad Pydi, S.; Singh, N.; Upadhyaya, J.; Pal Bhullar, R.; Chelikani, P. The third intracellular loop plays a critical role in bitter taste receptor activation. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Pydi, S.P.; Jafurulla, M.; Wai, L.; Bhullar, R.P.; Chelikani, P.; Chattopadhyay, A. Cholesterol modulates bitter taste receptor function. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 2081–2087. [Google Scholar] [CrossRef]
- Ilegems, E.; Iwatsuki, K.; Kokrashvili, Z.; Benard, O.; Ninomiya, Y.; Margolskee, R.F. REEP2 Enhances Sweet Receptor Function by Recruitment to Lipid Rafts. J. Neurosci. 2010, 30, 13774–13783. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekar, J.; Mueller, K.L.; Hoon, M.A.; Adler, E.; Feng, L.; Guo, W.; Zuker, C.S.; Ryba, N.J. T2Rs function as bitter taste receptors. Cell 2000, 100, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Shaik, F.A.; Medapati, M.R.; Chelikani, P. Cholesterol modulates the signaling of chemosensory bitter taste receptor T2R14 in human airway cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L45–L57. [Google Scholar] [CrossRef]
- Sarkanen, J.R.; Nykky, J.; Siikanen, J.; Selinummi, J.; Ylikomi, T.; Jalonen, T.O. Cholesterol supports the retinoic acid-induced synaptic vesicle formation in differentiating human SH-SY5Y neuroblastoma cells. J. Neurochem. 2007, 102, 1941–1952. [Google Scholar] [CrossRef]
- Harada, Y.; Yokota, C.; Habas, R.; Slusarski, D.C.; He, X. Retinoic acid-inducible G protein-coupled receptors bind to frizzled receptors and may activate non-canonical Wnt signaling. Biochem. Biophys. Res. Commun. 2007, 358, 968–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Lotan, R. Molecular Cloning and Characterization of a Novel Retinoic Acid-inducible Gene That Encodes a Putative G Protein-coupled Receptor*. J. Biol. Chem. 1998, 273, 35008–35015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bräuner-Osborne, H.; Jensen, A.A.; Sheppard, P.O.; Brodin, B.; Krogsgaard-Larsen, P.; O’Hara, P. Cloning and characterization of a human orphan family C G-protein coupled receptor GPRC5D1GenBank accession Nos. for GPRC5C: AF207989, for Gprc5d: AF218809 and for GPRC5D: AF209923.1. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2001, 1518, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.H.; Grossmann, M.; Ji, I. G Protein-coupled Receptors: I. Diversity of receptor-ligand interactions. J. Biol. Chem. 1998, 273, 17299–17302. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M.; MacLeod, R.J. Extracellular calcium sensing and extracellular calcium signaling. Physiol. Rev. 2001, 81, 239–297. [Google Scholar] [CrossRef]
- Hannan, F.M.; Kallay, E.; Chang, W.; Brandi, M.L.; Thakker, R.V. The calcium-sensing receptor in physiology and in calcitropic and noncalcitropic diseases. Nat. Rev. Endocrinol. 2019, 15, 33–51. [Google Scholar] [CrossRef]
- Kifor, O.; Diaz, R.; Butters, R.; Kifor, I.; Brown, E.M. The calcium-sensing receptor is localized in caveolin-rich plasma membrane domains of bovine parathyroid cells. J. Biol. Chem. 1998, 273, 21708–21713. [Google Scholar] [CrossRef] [Green Version]
- Augé, N.; Maupas-Schwalm, F.; Elbaz, M.; Thiers, J.C.; Waysbort, A.; Itohara, S.; Krell, H.W.; Salvayre, R.; Nègre-Salvayre, A. Role for matrix metalloproteinase-2 in oxidized low-density lipoprotein–induced activation of the sphingomyelin/ceramide pathway and smooth muscle cell proliferation. Circulation 2004, 110, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Aoyagi, M.; Yamamoto, M.; Azuma, H.; Nagashima, G.; Niimi, Y.; Tamaki, M.; Hirakawa, K.; Yamamoto, K. Immunolocalization of matrix metalloproteinases in rabbit carotid arteries after balloon denudation. Histochem. Cell Biol. 1998, 109, 97–102. [Google Scholar] [CrossRef]
- Sundararaman, S.S.; van der Vorst, E.P. Calcium-Sensing Receptor (CaSR), Its Impact on Inflammation and the Consequences on Cardiovascular Health. Int. J. Mol. Sci. 2021, 22, 2478. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, L.; Zielke, H.R.; Cheng, L.; Xiao, R.; Crow, M.T.; Stetler-Stevenson, W.G.; Froehlich, J.; Lakatta, E.G. Increased expression of 72-kd type IV collagenase (MMP-2) in human aortic atherosclerotic lesions. Am. J. Pathol. 1996, 148, 121. [Google Scholar] [PubMed]
- Molostvov, G.; Fletcher, S.; Bland, R.; Zehnder, D. Extracellular calcium-sensing receptor mediated signaling is involved in human vascular smooth muscle cell proliferation and apoptosis. Cell. Physiol. Biochem. 2008, 22, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Kong, F.J.; Bai, S.Z.; He, W.; Xing, W.J.; Xi, Y.H.; Li, G.W.; Guo, J.; Li, H.Z.; Wu, L.Y.; et al. Involvement of calcium-sensing receptor in oxLDL-induced MMP-2 production in vascular smooth muscle cells via PI3K/Akt pathway. Mol. Cell. Biochem. 2012, 362, 115–122. [Google Scholar] [CrossRef]
- Tang, X.l.; Wang, Y.; Li, D.l.; Luo, J.; Liu, M.y. Orphan G protein-coupled receptors (GPCRs): Biological functions and potential drug targets. Acta Pharmacol. Sin. 2012, 33, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Jetten, A.M.; Takeda, Y.; Slominski, A.; Kang, H.S. Retinoic acid-related orphan receptor γ (RORγ): Connecting sterol metabolism to regulation of the immune system and autoimmune disease. Curr. Opin. Toxicol. 2018, 8, 66–80. [Google Scholar] [CrossRef]
- Jeong, E.; Kim, Y.; Jeong, J.; Cho, Y. Structure of the class C orphan GPCR GPR158 in complex with RGS7-Gβ5. Nat. Commun. 2021, 12, 6805. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.P.; Orlandi, C.; Song, C.; Oh, W.C.; Muntean, B.S.; Xie, K.; Filippini, A.; Xie, X.; Satterfield, R.; Yaeger, J.D.W.; et al. Orphan receptor GPR158 controls stress-induced depression. eLife 2018, 7, e33273. [Google Scholar] [CrossRef]
- Khrimian, L.; Obri, A.; Ramos-Brossier, M.; Rousseaud, A.; Moriceau, S.; Nicot, A.S.; Mera, P.; Kosmidis, S.; Karnavas, T.; Saudou, F.; et al. Gpr158 mediates osteocalcin’s regulation of cognition. J. Exp. Med. 2017, 214, 2859–2873. [Google Scholar] [CrossRef] [Green Version]
- Insel, P.A.; Wilderman, A.; Zambon, A.C.; Snead, A.N.; Murray, F.; Aroonsakool, N.; McDonald, D.S.; Zhou, S.; McCann, T.; Zhang, L.; et al. G Protein–Coupled Receptor (GPCR) Expression in Native Cells: “Novel” endoGPCRs as Physiologic Regulators and Therapeutic Targets. Mol. Pharmacol. 2015, 88, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Civelli, O. Orphan GPCRs and Neuromodulation. Neuron 2012, 76, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlandi, C.; Posokhova, E.; Masuho, I.; Ray, T.A.; Hasan, N.; Gregg, R.G.; Martemyanov, K.A. GPR158/179 regulate G protein signaling by controlling localization and activity of the RGS7 complexes. J. Cell Biol. 2012, 197, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Hollinger, S.; Hepler, J.R. Cellular Regulation of RGS Proteins: Modulators and Integrators of G Protein Signaling. Pharmacol. Rev. 2002, 54, 527–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, E.M.; Wilkie, T.M. GTPase-Activating Proteins for Heterotrimeric G Proteins: Regulators of G Protein Signaling (RGS) and RGS-Like Proteins. Annu. Rev. Biochem. 2000, 69, 795–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | PDB ID * | Number of Sterols in TM |
|---|---|---|
| mGluR1 | 4OR2 [36] | 6 CLR |
| mGluR5 | 7FD8 [45] | 2 CHS |
| GABA | 6WIV [24] | 10 CLR |
| 7CUM [65] | 16 CLR | |
| 7CA3 [65] | 3 CLR | |
| CaSR | 7SIM [66] | 8 CLR |
| 7SIL [66] | 8 CLR | |
| Orphan receptor (GPR158) | 7SHF [56] | 22 CLR |
| 7SHE [56] | 22 CLR ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isu, U.H.; Badiee, S.A.; Khodadadi, E.; Moradi, M. Cholesterol in Class C GPCRs: Role, Relevance, and Localization. Membranes 2023, 13, 301. https://doi.org/10.3390/membranes13030301
Isu UH, Badiee SA, Khodadadi E, Moradi M. Cholesterol in Class C GPCRs: Role, Relevance, and Localization. Membranes. 2023; 13(3):301. https://doi.org/10.3390/membranes13030301
Chicago/Turabian StyleIsu, Ugochi H., Shadi A Badiee, Ehsaneh Khodadadi, and Mahmoud Moradi. 2023. "Cholesterol in Class C GPCRs: Role, Relevance, and Localization" Membranes 13, no. 3: 301. https://doi.org/10.3390/membranes13030301
APA StyleIsu, U. H., Badiee, S. A., Khodadadi, E., & Moradi, M. (2023). Cholesterol in Class C GPCRs: Role, Relevance, and Localization. Membranes, 13(3), 301. https://doi.org/10.3390/membranes13030301