
αvβ3 Integrin Expression and Mitogenic Effects by Thyroid Hormones in Chronic Lymphocytic Leukemia

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
3. Results
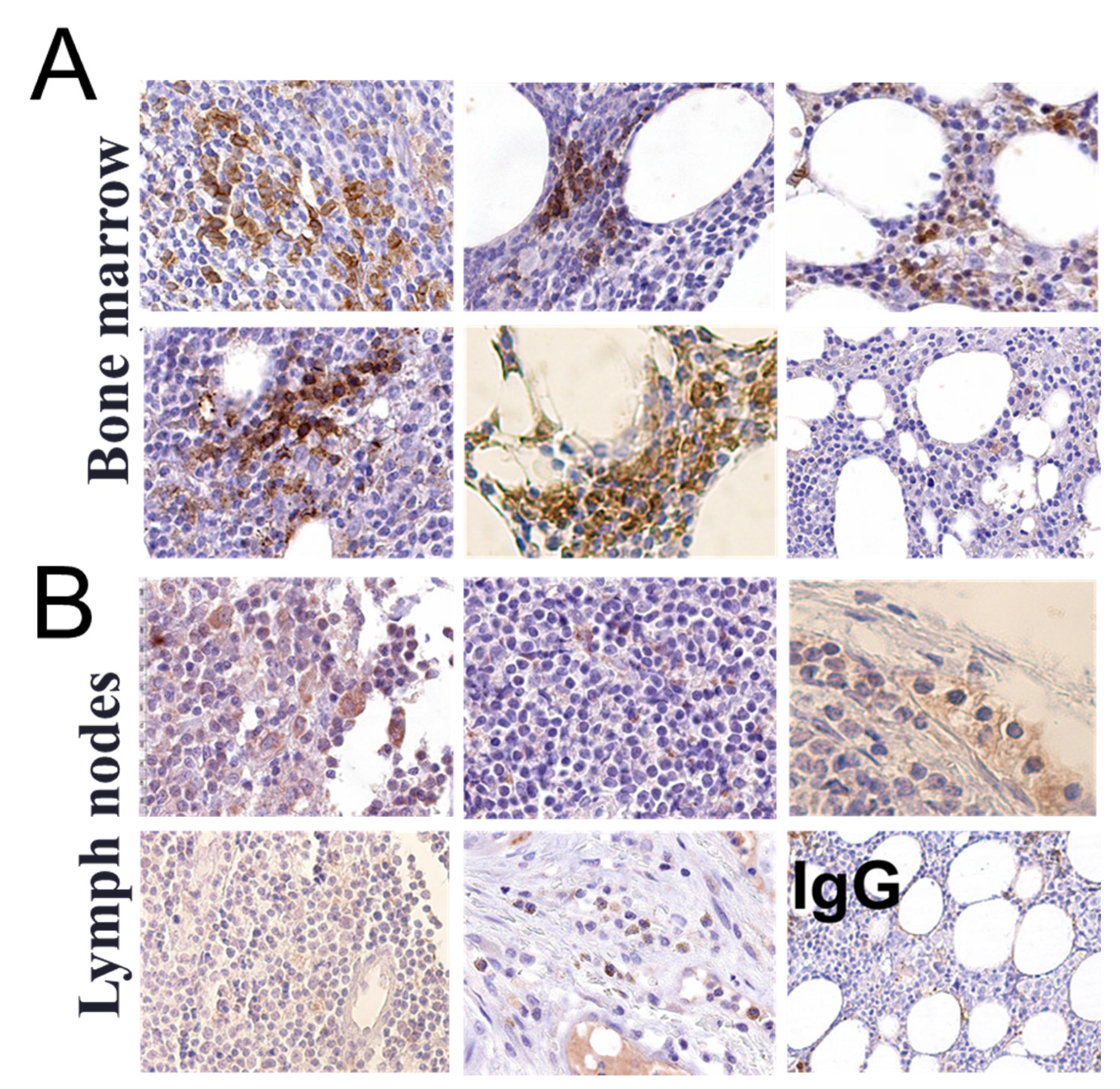
3.1. αvβ3 Integrin Is Expressed in Circulating CLL B Lymphocytes and in the Bone Marrow but Not in the Lymph Node of CLL Patients
3.2. Thyroid Hormones Induce αvβ3 Integrin Expression in Primary CLL Cells
3.3. Mitogenic Signals Are Induced by Thyroid Hormones in Primary CLL Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hus, I.; Roliński, J. Current concepts in diagnosis and treatment of chronic lymphocytic leukemia. Contemp. Oncol. 2015, 19, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaidano, G.; Foà, R.; Dalla-Favera, R. Molecular pathogenesis of chronic lymphocytic leukemia. J. Clin. Investig. 2012, 122, 3432–3438. [Google Scholar] [CrossRef] [Green Version]
- Messmer, B.T.; Messmer, D.; Allen, S.L.; Kolitz, J.E.; Kudalkar, P.; Cesar, D.; Murphy, E.J.; Koduru, P.; Ferrarini, M.; Zupo, S. In vivo measurements document the dynamic cellular kinetics of chronic lymphocytic leukemia B cells. J. Clin. Investig. 2005, 115, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Cacciatore, M.; Guarnotta, C.; Calvaruso, M.; Sangaletti, S.; Florena, A.M.; Franco, V.; Colombo, M.P.; Tripodo, C. Microenvironment-centred dynamics in aggressive B-cell lymphomas. Adv. Hematol. 2012, 2012, 138079. [Google Scholar] [CrossRef] [Green Version]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Giancotti, F. Integrin signalling during tumour progression. Nat. Rev. Mol. Cell Biol. 2004, 5, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Moeller, L.C.; Fuhrer, D. Thyroid hormone, thyroid hormone receptors, and cancer: A clinical perspective. Endocr. Relat. Cancer 2013, 20, R19–R29. [Google Scholar] [CrossRef] [Green Version]
- Davis, P.J.; Goglia, F.; Leonard, J.L. Nongenomic actions of thyroid hormone. Nat. Rev. Endocrinol. 2016, 12, 111–121. [Google Scholar] [CrossRef]
- Cody, V.; Davis, P.J.; Davis, F.B. Molecular modeling of the thyroid hormone interactions with alpha v beta 3 integrin. Steroids 2007, 72, 165–170. [Google Scholar] [CrossRef]
- Lin, H.Y.; Landersdorfer, C.B.; London, D.; Meng, R.; Lim, C.U.; Lin, C.; Lin, S.; Tang, H.Y.; Brown, D.; Van Scoy, B.; et al. Pharmacodynamic modeling of anti-cancer activity of tetraiodothyroacetic acid in a perfused cell culture system. PLoS Comput. Biol. 2011, 7, e1001073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Stupack, D.; Mathias, P.; Wang, Y.; Nemerow, G. Growth arrest of Epstein–Barr virus immortalized B lymphocytes by adenovirus-delivered ribozymes. Proc. Natl. Acad. Sci. USA 1997, 94, 8156–8161. [Google Scholar] [CrossRef] [Green Version]
- Wilder, R. Integrin alpha V beta 3 as a target for treatment of rheumatoid arthritis and related rheumatic diseases. Ann. Rheum. Dis. 2002, 61, ii96–ii99. [Google Scholar] [CrossRef]
- Till, K.J.; Spiller, D.G.; Harris, R.J.; Chen, H.; Zuzel, M.; Cawley, J.C. CLL, but not normal, B cells are dependent on autocrine VEGF and α4β1 integrin for chemokine-induced motility on and through endothelium. Blood 2005, 105, 4813–4819. [Google Scholar] [CrossRef]
- Plate, J.; Long, B.; Kelkar, S. Role of β2 integrins in the prevention of apoptosis induction in chronic lymphocytic leukemia B cells. Leukemia 2000, 14, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldini, L.G.; Cro, L.M. Structure and function of VLA integrins: Differential expression in B-cell leukemia/lymphoma. Leuk. Lymphoma 1994, 12, 197–203. [Google Scholar] [CrossRef]
- Takeuchi, H.; Katayama, I. Surface phenotype and adhesion activity of B-cell chronic lymphoid leukemias. Leuk. Lymphoma 1993, 10, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Cawley, J.; Burthem, J. Integrin function in chronic lymphocytic leukemia. Blood 1996, 87, 4780–4788. [Google Scholar] [CrossRef] [Green Version]
- Mateo, V.; Lagneaux, L.; Bron, D.; Biron, G.; Armant, M.; Delespesse, G.; Sarfati, M. CD47 ligation induces caspase-independent cell death in chronic lymphocytic leukemia. Nat. Med. 1999, 5, 1277. [Google Scholar] [CrossRef]
- Bairey, O.; Zimra, Y.; Rabizadeh, E.; Shaklai, M. Expression of adhesion molecules on leukemic B cells from chronic lymphocytic leukemia patients with predominantly splenic manifestations. IMAJ-RAMAT GAN 2004, 6, 147–151. [Google Scholar]
- De Rossi, G.; Zarcone, D.; Mauro, F.; Cerruti, G.; Tenca, C.; Puccetti, A.; Mandelli, F.; Grossi, C.E. Adhesion molecule expression on B-cell chronic lymphocytic leukemia cells: Malignant cell phenotypes define distinct disease subsets. Blood 1993, 81, 2679–2687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurisic, V.; Colovic, N.; Kraguljac, N.; Atkinson, H.D.; Colovic, M. Analysis of CD23 antigen expression in B-chronic lymphocytic leukaemia and its correlation with clinical parameters. Med. Oncol. 2008, 25, 315–322. [Google Scholar] [CrossRef]
- Fournier, S.; Tran, D.; Suter, U.; Biron, G.; Delespesse, G.; Sarfati, M. The in vivo expression of type B CD23 mRNA in B-chronic lymphocytic leukemic cells is associated with an abnormally low CD23 upregulation by IL-4: Comparison with their normal cellular counterparts. Leuk. Res. 1991, 15, 609–618. [Google Scholar] [CrossRef]
- Sarfati, M.; Chevret, S.; Chastang, C.; Biron, G.; Stryckmans, P.; Delespesse, G.; Binet, J.-L.; Merle-Beral, H.; Bron, D. Prognostic importance of serum soluble CD23 level in chronic lymphocytic leukemia. Blood 1996, 88, 4259–4264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ria, R.; Vacca, A.; Ribatti, D.; Di Raimondo, F.; Merchionne, F.; Dammacco, F. Alpha(v)beta(3) integrin engagement enhances cell invasiveness in human multiple MM. Haematologica 2002, 87, 836–845. [Google Scholar] [PubMed]
- Vacca, A.; Ria, R.; Presta, M.; Ribatti, D.; Iurlaro, M.; Merchionne, F.; Tanghetti, E.; Dammacco, F. Avb3 integrin engagement modulates cell adhesion, proliferation, and protease secretion in human lymphoid tumor cells. Exp. Hematol. 2001, 29, 993–1003. [Google Scholar] [CrossRef]
- Cohen, K.; Abadi, U.; Hercbergs, A.; Davis, P.J.; Ellis, M.; Ashur-Fabian, O. The induction of myeloma cell death and DNA damage by tetrac, a thyroid hormone derivative. Endocr. Relat. Cancer 2018, 25, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.; Ellis, M.; Khoury, S.; Davis, P.J.; Hercbergs, A.; Ashur-Fabian, O. Thyroid hormone is a MAPK-dependent growth factor for human myeloma cells acting via alphavbeta3 integrin. Mol. Cancer Res. 2011, 9, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinderman-Maman, E.; Cohen, K.; Weingarten, C.; Nabriski, D.; Twito, O.; Baraf, L.; Hercbergs, A.; Davis, P.J.; Werner, H.; Ellis, M.; et al. The thyroid hormone-alphavbeta3 integrin axis in ovarian cancer: Regulation of gene transcription and MAPK-dependent proliferation. Oncogene 2016, 35, 1977–1987. [Google Scholar] [CrossRef]
- Morris, D.; Wolff, F.; Upton, A.C. The influence of the thyroid gland on the survival of rats and mice bearing transplanted lymphoid leukemia. Cancer Res. 1957, 17, 325–328. [Google Scholar]
- Morris, D.M. The effect of thyroidectomy and thyroid-stimulating hormone on a transplanted acute leukemia in the Fischer rat. Cancer Res. 1960, 20, 373–376. [Google Scholar]
- Krashin, E.; Piekiełko-Witkowska, A.; Ellis, M.; Ashur-Fabian, O. Thyroid hormones and cancer: A comprehensive review of preclinical and clinical studies. Front. Endocrinol. 2019, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cayrol, F.; Sterle, H.A.; Díaz Flaqué, M.C.; Barreiro Arcos, M.L.; Cremaschi, G.A. Non-genomic actions of thyroid hormones regulate the growth and angiogenesis of T cell lymphomas. Front. Endocrinol. 2019, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abadi, U.; Weisz, A.; Kidron, D.; Katzav, A.; Hercbergs, A.; Davis, P.J.; Ellis, M.H.; Ashur-Fabian, O. αvβ3 Integrin Expression and Mitogenic Effects by Thyroid Hormones in Chronic Lymphocytic Leukemia. J. Clin. Med. 2021, 10, 1766. https://doi.org/10.3390/jcm10081766
Abadi U, Weisz A, Kidron D, Katzav A, Hercbergs A, Davis PJ, Ellis MH, Ashur-Fabian O. αvβ3 Integrin Expression and Mitogenic Effects by Thyroid Hormones in Chronic Lymphocytic Leukemia. Journal of Clinical Medicine. 2021; 10(8):1766. https://doi.org/10.3390/jcm10081766
Chicago/Turabian StyleAbadi, Uri, Avivit Weisz, Dvora Kidron, Aviva Katzav, Aleck Hercbergs, Paul J. Davis, Martin H. Ellis, and Osnat Ashur-Fabian. 2021. "αvβ3 Integrin Expression and Mitogenic Effects by Thyroid Hormones in Chronic Lymphocytic Leukemia" Journal of Clinical Medicine 10, no. 8: 1766. https://doi.org/10.3390/jcm10081766
APA StyleAbadi, U., Weisz, A., Kidron, D., Katzav, A., Hercbergs, A., Davis, P. J., Ellis, M. H., & Ashur-Fabian, O. (2021). αvβ3 Integrin Expression and Mitogenic Effects by Thyroid Hormones in Chronic Lymphocytic Leukemia. Journal of Clinical Medicine, 10(8), 1766. https://doi.org/10.3390/jcm10081766