
Immune System Dysfunction and Inflammation in Hemodialysis Patients: Two Sides of the Same Coin

 ,
,  and
and
Abstract
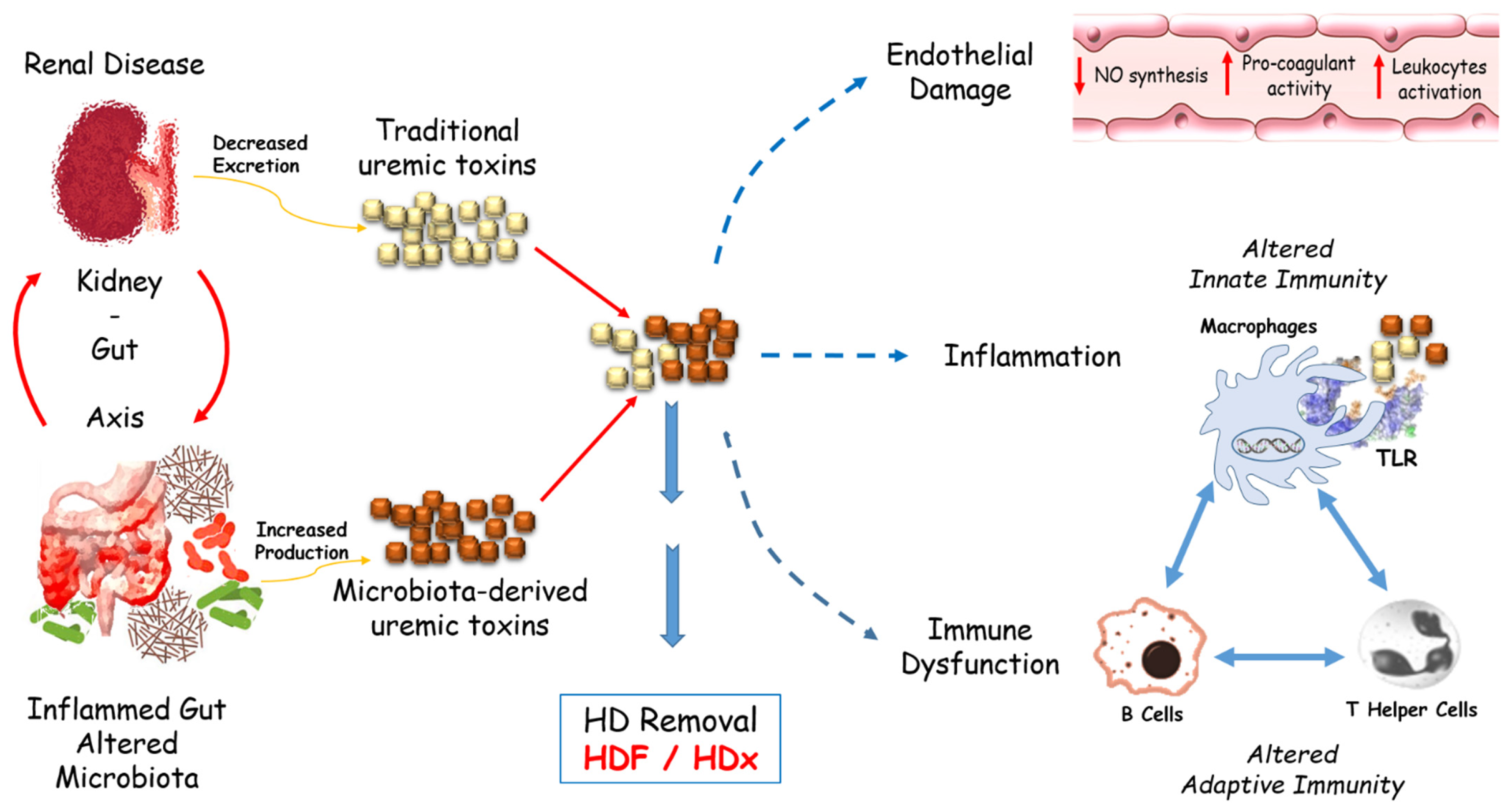
:1. Introduction
2. Platelets and Coagulation System
3. Innate Immune System
3.1. Complement
3.2. Neutrophils and Monocyte Macrophages
4. Acquired Immune System
T and B Cells
5. Inflammation and Oxidative Stress
6. Uremic Toxin Involvement
7. Future Perspectives
8. Conclusions
Funding
Conflicts of Interest
References
- Liyanage, T.; Ninomiya, T.; Jha, V.; Neal, B.; Patrice, H.M.; Okpechi, I.; Zhao, M.H.; Lv, J.; Garg, A.X.; Knight, J.; et al. Worldwide access to treatment for end-stage kidney disease: A systematic review. Lancet 2015, 385, 1975–1982. [Google Scholar] [CrossRef]
- Monardo, P.; Lacquaniti, A.; Campo, S.; Bucca, M.; Casuscelli di Tocco, T.; Rovito, S.; Ragusa, A.; Santoro, A. Updates on hemodialysis techniques with a common denominator: The personalization of the dialytic therapy. Semin. Dial. 2021, 34, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, L. Inflammation and Cardiovascular Disease Associated With Hemodialysis for End-Stage Renal Disease. Front. Pharmacol. 2022, 13, 800950. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, F.; Zoccali, C.; SIR SIN Study Investigators. Clinical policies on the management of chronic kidney disease patients in Italy. Nephrol. Dial. Transpl. 2008, 23, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Jofré, R.; Rodriguez-Benitez, P.; López-Gómez, J.M.; Pérez-Garcia, R. Inflammatory Syndrome in Patients on Hemodialysis. J. Am. Soc. Nephrol. 2006, 17, S274–S280. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. New York Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Canaud, B.; Kooman, J.P.; Selby, N.M.; Taal, M.W.; Francis, S.; Maierhofer, A.; Kopperschmidt, P.; Collins, A.; Kotanko, P. Dialysis-Induced Cardiovascular and Multiorgan Morbidity. Kidney Int. Rep. 2020, 5, 1856–1869. [Google Scholar] [CrossRef]
- Ratner, B.D. The biocompatibility manifesto: Biocompatibility for the twenty-first century. J. Cardiovasc. Transl. Res. 2011, 4, 523–527. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.F. Definitions in biomaterials. In Proceedings of the a Consensus Conference of the European Society for Biomaterials, Chester, UK, 3–5 March 1986; Elsevier: Amsterdam, The Netherlands, 1987; Volume 4. [Google Scholar]
- Williams, D.F. On the mechanisms of biocompatibility. Biomaterials 2008, 29, 2941–2953. [Google Scholar] [CrossRef]
- Doherty, P.J. (Ed.) Biomaterial-tissue interfaces. In Proceedings of the Ninth European Conference on Biomaterials, Chester, UK, 9–11 September 1991; Elsevier: Amsterdam, The Netherlands, 1991; Volume 10. [Google Scholar]
- Williams, D.F. Biocompatibility pathways and mechanisms for bioactive materials: The bio-activity zone. Bioact. Mater 2022, 10, 306–322. [Google Scholar] [CrossRef]
- Kokubo, K.; Kurihara, Y.; Kobayashi, K.; Tsukao, H.; Kobayashi, H. Evaluation of the Biocompatibility of Dialysis Membranes. Blood Purif. 2015, 40, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Rai, V.; Agrawal, D.K. Transcriptomic Analysis Identifies Differentially Expressed Genes Associated with Vascular Cuffing and Chronic Inflammation Mediating Early Thrombosis in Arteriovenous Fistula. Biomedicines 2022, 10, 433. [Google Scholar] [CrossRef] [PubMed]
- Lyu, B.; Chan, M.R.; Yevzlin, A.S.; Gardezi, A.; Astor, B.C. Arteriovenous Access Type and Risk of Mortality, Hospitalization, and Sepsis Among Elderly Hemodialysis Patients: A Target Trial Emulation Approach. Am. J. Kidney Dis. 2022, 79, 69–78. [Google Scholar] [CrossRef]
- Kameshwar, K.; Damasiewicz, M.J.; Polkinghorne, K.R.; Kerr, P.G. A pilot study comparing the efficiency of a novel asymmetric cellulose triacetate (ATA) dialyser membrane (Solacea-190H) to a standard high flux polysulfone dialyser membrane (FX-80) in the setting of extended hours haemodialysis. Nephrology 2022, 27, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Abdelrasoul, A.; Westphalen, H.; Saadati, S.; Shoker, A. Hemodialysis biocompatibility mathematical models to predict the inflammatory biomarkers released in dialysis patients based on hemodialysis membrane characteristics and clinical practices. Sci. Rep. 2021, 11, 23080. [Google Scholar] [CrossRef]
- Westphalen, H.; Saadati, S.; Eduok, U.; Abdelrasoul, A.; Shoker, A.; Choi, P.; Doan, H.; Ein-Mozaffari, F. Case studies of clinical hemodialysis membranes: Influences of membrane morphology and biocompatibility on uremic blood-membrane interactions and inflammatory biomarkers. Sci. Rep. 2020, 10, 14808. [Google Scholar] [CrossRef]
- Craddock, P.R.; Fehr, J.; Brigham, K.L.; Kronenberg, R.S.; Jacob, H.S. Complement and leukocyte-mediated pulmonary dysfunction in hemodialysis. N. Engl. J. Med. 1977, 296, 769–774. [Google Scholar] [CrossRef]
- Poppelaars, F.; Faria, B.; Gaya da Costa, M.; Franssen, C.; van Son, W.J.; Berger, S.P.; Daha, M.R.; Seelen, M.A. The Complement System in Dialysis: A Forgotten Story? Front. Immunol. 2018, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Westphalen, H.; Abdelrasoul, A.; Shoker, A. Protein adsorption phenomena in hemodialy-sis membranes: Mechanisms, influences of clinical practices, modeling, and challenges. Colloid Interface Sci. Commun. 2021, 40, 100348. [Google Scholar] [CrossRef]
- Frank, R.D.; Weber, J.; Dresbach, H.; Thelen, H.; Weiss, C.; Floege, J. Role of contact system activation in hemodialyzer-induced thrombogenicity. Kidney Int. 2001, 60, 1972–1981. [Google Scholar] [CrossRef] [Green Version]
- Marney, A.M.; Ma, J.; Luther, J.M.; Ikizler, T.A.; Brown, N.J. Endogenous bradykinin contributes to increased plasminogen activator inhibitor 1 antigen following hemodialysis. J. Am. Soc. Nephrol. 2009, 20, 2246–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butani, L.; Calogiuri, G. Hypersensitivity reactions in patients receiving hemodialysis. Am. Coll. Allergy Asthma Immunol. 2017, 118, 680–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, S.C.; Derebail, V.K.; Poulton, C.J.; Bunch, D.C.; Roy-Chaudhury, P.; Key, N.S. Hemodialysis-Related Complement and Contact Pathway Activation and Cardiovascular Risk: A Narrative Review. Kidney Med. 2021, 3, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Coughlin, S.R. Protease-activated receptors in hemostasis, thrombosis and vascular biology. J. Thromb. Haemost. 2005, 3, 1800–1814. [Google Scholar] [CrossRef] [PubMed]
- Martorell, L.; Martínez-González, J.; Rodríguez, C.; Gentile, M.; Calvayrac, O.; Badimon, L. Thrombin and protease-activated receptors (PARs) in atherothrombosis. Thromb. Haemost. 2008, 99, 305–315. [Google Scholar] [CrossRef]
- Sabry, A.; Taha, M.; Nada, M.; Al Fawzan, F.; Alsaran, K. Anticoagulation therapy during haemodialysis: A comparative study between two heparin regimens. Blood Coagul. Fibrinolysis 2009, 20, 57–62. [Google Scholar] [CrossRef]
- Tennankore, K.K.; d’Gama, C.; Faratro, R.; Fung, S.; Wong, E.; Chan, C.T. Adverse technical events in home hemodialysis. Am. J. Kidney Dis. 2015, 65, 116–121. [Google Scholar] [CrossRef]
- Richtrova, P.; Rulcova, K.; Mares, J.; Reischig, T. Evaluation of Three Different Methods to Prevent Dialyzer Clotting Without Causing Systemic Anticoagulation Effect. Artif. Organs 2011, 35, 83–88. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, X.; Wang, S.; Li, H. High Neutrophil-to-Lymphocyte Ratio and Platelet-to-Lymphocyte Ratio Are Associated with Poor Survival in Patients with Hemodialysis. BioMed Res. Int. 2021, 9958081. [Google Scholar] [CrossRef]
- Abdelmaguid, A.; Roberts, L.N.; Tugores, L.; Joslin, J.R.; Hunt, B.J.; Parmar, K.; Nebres, D.; Naga, S.S.; Khalil, E.S.; Bramham, K. Evaluation of novel coagulation and platelet function assays in patients with chronic kidney disease. J. Thromb. Haemost. 2022, 20, 845–856. [Google Scholar] [CrossRef]
- Daugirdas, J.T.; Bernardo, A.A. Hemodialysis effect on platelet count and function and hemodialysis-associated thrombocytopenia. Kidney Int. 2012, 82, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafiranye, F.; Kyaw, W.; Olafiranye, O. Resolution of dialyzer membrane-associated thrombocytopenia with use of cellulose triacetate membrane: A case report. Case Rep. Med. 2011, 2011, 134295. [Google Scholar] [CrossRef] [PubMed]
- De Prada, L.; Lee, J.; Gillespie, A.; Benjamin, J. Thrombocytopenia associated with one type of polysulfone hemodialysis membrane: A report of 5 cases. Am. J. Kidney Dis. 2013, 61, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Claudio-Gonzalez, I.; Ravindranathan, D.; Kempton, C.L.; Bailey, J.L.; Wall, S.M. Thrombocytopenia Induced by Polysulfone Dialysis Membranes. Am. J. Case Rep. 2021, 22, e932045. [Google Scholar] [CrossRef]
- Zawada, A.M.; Melchior, P.; Erlenkötter, A.; Delinski, D.; Stauss-Grabo, M.; Kennedy, J.P. Polyvinylpyrrolidone in hemodialysis membranes: Impact on platelet loss during hemodialysis. Hemodial. Int. 2021, 25, 498–506. [Google Scholar] [CrossRef]
- Zhu, L.; Song, H.; Wang, J.; Xue, L. Polysulfone hemodiafiltration membranes with enhanced anti-fouling and hemocompatibility modified by poly(vinyl pyrrolidone) via in situ cross-linked polymerization. Mater Sci. Eng. C Mater Biol. Appl. 2017, 74, 159–166. [Google Scholar] [CrossRef]
- Gubensek, J.; Strobl, K.; Harm, S.; Weiss, R.; Eichhorn, T.; Buturovic-Ponikvar, J.; Weber, V.; Hartmann, J. Influence of citrate concentration on the activation of blood cells in an in vitro dialysis setup. PLoS ONE 2018, 13, e0199204. [Google Scholar] [CrossRef] [Green Version]
- Orsag, A.; Bozic-Mijovski, M.; Hudoklin, S.; Simcic, S.; Gubensek, J. Biocompatibility Parameters with Standard and Increased Dose of Citrate in Hemodialysis-A Randomized Trial. J. Clin. Med. 2021, 10, 2987. [Google Scholar] [CrossRef]
- Inoshita, H.; Ohsawa, I.; Kusaba, G.; Ishii, M.; Onda, K.; Horikoshi, S.; Ohi, H.; Tomino, Y. Complement in patients receiving maintenance hemodialysis: Functional screening and quantitative analysis. BMC Nephrol. 2010, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Zoccali, C.; Moissl, U.; Chazot, C.; Mallamaci, F.; Tripepi, G.; Arkossy, O.; Wabel, P.; Stuard, S. Chronic Fluid Overload and Mortality in ESRD. J. Am. Soc. Nephrol. 2017, 28, 2491–2497. [Google Scholar] [CrossRef] [Green Version]
- Melchior, P.; Erlenkötter, A.; Zawada, A.M.; Delinski, D.; Schall, C.; Stauss-Grabo, M.; Kennedy, J.P. Complement activation by dialysis membranes and its association with secondary membrane formation and surface charge. Artif. Organs 2021, 45, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Stepniewska, J.; Dolegowska, B.; Golembiewska, E.; Marchelek-Mysliwiec, M.; Domanski, M.; Ciechanowski, K.; Zair, L. The activation of complement system in different types of renal replacement therapy. J. Physiol. Pharmacol. 2020, 71, 275–281. [Google Scholar] [CrossRef]
- Cheung, A.K.; Chang, T.I.; Cushman, W.C.; Furth, S.L.; Hou, F.F.; Ix, J.H.; Knoll, G.A.; Muntner, P.; Pecoits-Filho, R.; Sarnak, M.J. Executive summary of the KDIGO 2021 Clinical Practice Guideline for the Management of Blood Pressure in Chronic Kidney Disease. Kidney Int. 2021, 99, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.L.; Pilely, K.; Lund, K.P.; Warming, P.E.; Plesner, L.L.; Iversen, K.K.; Garred, P. Hemodialysis leads to plasma depletion of lectin complement pathway initiator molecule ficolin-2. Hemodial. Int. 2021, 25, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Mares, J.; Richtrova, P.; Hricinova, A.; Tuma, Z.; Moravec, J.; Lysak, D.; Matejovic, M. Proteomic profiling of blood-dialyzer interactome reveals involvement of lectin complement pathway in hemodialysis-induced inflammatory response. Proteom. Clin. Appl. 2010, 4, 829–838. [Google Scholar] [CrossRef]
- Kim, H.J.; Seong, E.Y.; Lee, W.; Kim, S.; Ahn, H.S.; Yeom, J.; Kim, K.; Kwon, C.H.; Song, S.H. Comparative analysis of therapeutic effects between medium cut-off and high flux dialyzers using metabolomics and proteomics: Exploratory, prospective study in hemodialysis. Sci. Rep. 2021, 11, 17335. [Google Scholar] [CrossRef]
- Huang, S.; Sandholm, K.; Jonsson, N.; Nilsson, A.; Wieslander, A.; Grundström, G.; Hancock, V.; Ekdahl, K.N. Low concentrations of citrate reduce complement and granulocyte activation in vitro in human blood. Clin. Kidney J. 2015, 8, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Kourtzelis, I.; Markiewski, M.M.; Doumas, M.; Rafail, S.; Kambas, K.; Mitroulis, I.; Panagoutsos, S.; Passadakis, P.; Vargemezis, V.; Magotti, P.; et al. Complement anaphylatoxin C5a contributes to hemodialysis-associated thrombosis. Blood 2010, 116, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Ning, J.P. Synthesis and biocompatibility of an argatroban-modified polysulfone membrane that directly inhibits thrombosis. J. Mater. Sci. Mater. Med. 2018, 29, 66. [Google Scholar] [CrossRef]
- Ekdahl, K.N.; Soveri, I.; Hilborn, J.; Fellström, B.; Nilsson, B. Cardiovascular disease in haemodialysis: Role of the intravascular innate immune system. Nat. Rev. Nephrol. 2017, 13, 285–296. [Google Scholar] [CrossRef]
- Fukushi, T.; Yamamoto, T.; Yoshida, M.; Fujikura, E.; Miyazaki, M.; Nakayama, M. Enhanced neutrophil apoptosis accompanying myeloperoxidase release during hemodialysis. Sci. Rep. 2020, 10, 21747. [Google Scholar] [CrossRef] [PubMed]
- Bieber, S.; Muczynski, K.A.; Lood, C. Neutrophil Activation and Neutrophil Extracellular Trap Formation in Dialysis Patients. Kidney Med. 2020, 2, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Koga, Y.; Fujieda, H.; Meguro, H.; Ueno, Y.; Aoki, T.; Miwa, K.; Kainoh, M. Biocompatibility of Polysulfone Hemodialysis Membranes and Its Mechanisms: Involvement of Fibrinogen and Its Integrin Receptors in Activation of Platelets and Neutrophils. Artif. Organs 2018, 42, E246–E258. [Google Scholar] [CrossRef]
- Campillo, S.; Bohorquez, L.; Gutiérrez-Calabrés, E.; García-Ayuso, D.; Miguel, V.; Griera, M.; Calle, Y.; de Frutos, S.; Rodríguez-Puyol, M.; Rodríguez-Puyol, D.; et al. Indoxyl sulfate- and P-cresol-induced monocyte adhesion and migration is mediated by integrin-linked kinase-dependent podosome formation. Exp. Mol. Med. 2022, 54, 226–238. [Google Scholar] [CrossRef]
- Ziegler-Heitbrock, L.; Ancuta, P.; Crowe, S.; Dalod, M.; Grau, V.; Hart, D.N.; Leenen, P.J.; Liu, Y.J.; MacPherson, G.; Randolph, G.J.; et al. Nomenclature of monocytes and dendritic cells in blood. Blood 2010, 116, e74–e80. [Google Scholar] [CrossRef]
- Carmona, A.; Agüera, M.L.; Luna-Ruiz, C.; Buendía, P.; Calleros, L.; García-Jerez, A.; Rodríguez-Puyol, M.; Arias, M.; Arias-Guillen, M.; de Arriba, G.; et al. Markers of endothelial damage in patients with chronic kidney disease on hemodialysis. Am. J. Physiology. Ren. Physiol. 2017, 312, F673–F681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges Bonan, N.; Schepers, E.; Pecoits-Filho, R.; Dhondt, A.; Pletinck, A.; De Somer, F.; Vanholder, R.; Van Biesen, W.; Moreno-Amaral, A.; Glorieux, G. Contribution of the uremic milieu to an increased pro-inflammatory monocytic phenotype in chronic kidney disease. Sci. Rep. 2019, 9, 10236. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, V.; Jeron, A.; Shah, A.; Bruder, D.; Mertens, P.R.; Gorny, X. Hemodialysis-related changes in phenotypical features of monocytes. Sci. Rep. 2018, 8, 13964. [Google Scholar] [CrossRef] [Green Version]
- Ariza, F.; Merino, A.; Carracedo, J.; Alvarez de Lara, M.A.; Crespo, R.; Ramirez, R.; Martín-Malo, A.; Aljama, P. Post-dilution high convective transport improves microinflammation and endothelial dysfunction independently of the technique. Blood Purif. 2013, 35, 270–278. [Google Scholar] [CrossRef]
- Bolasco, P.; Spiga, P.; Arras, M.; Murtas, S.; La Nasa, G. Could there be Haemodynamic Stress Effects on Pro-Inflammatory CD14+CD16+ Monocytes during Convective-Diffusive Treatments? A Prospective Randomized Controlled Trial. Blood Purif. 2019, 47, 385–394. [Google Scholar] [CrossRef]
- Fiedler, R.; Neugebauer, F.; Ulrich, C.; Wienke, A.; Gromann, C.; Storr, M.; Böhler, T.; Seibert, E.; Girndt, M. Randomized controlled pilot study of 2 weeks’ treatment with high cutoff membrane for hemodialysis patients with elevated C-reactive protein. Artif. Organs 2012, 36, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Girndt, M.; Lengler, S.; Kaul, H.; Sester, U.; Sester, M.; Köhler, H. Prospective crossover trial of the influence of vit-amin E-coated dialyzer membranes on T-cell activation and cytokine induction. Am. J. Kidney Dis. 2000, 35, 95–104. [Google Scholar] [CrossRef]
- Nockher, W.A.; Wiemer, J.; Scherberich, J.E. Haemodialysis monocytopenia: Differential sequestration kinetics of CD14+CD16+ and CD14++ blood monocyte subsets. Clin. Exp. Immunol. 2001, 123, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Sester, U.; Sester, M.; Heine, G.; Kaul, H.; Girndt, M.; Köhler, H. Strong depletion of CD14(+) CD16(+) monocytes during haemodialysis treatment. Nephrol. Dial. Transpl. 2001, 16, 1402–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansouri, L.; Nopp, A.; Jacobson, S.H.; Hylander, B.; Lundahl, J. Hemodialysis Patients Display a Declined Proportion of Th2 and Regulatory T Cells in Parallel with a High Inter-feron-γ Profile. Nephron 2017, 136, 254–260. [Google Scholar] [CrossRef]
- Meijers, R.W.; Litjens, N.H.; de Wit, E.A.; Langerak, A.W.; van der Spek, A.; Baan, C.C.; Weimar, W.; Betjes, M.G. Uremia causes premature ageing of the T cell compartment in end-stage renal disease patients. Immun. Ageing 2012, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Betjes, M.G.H.; Meijers, R.W.J.; de Wit, L.E.A.; Litjens, N.H.R. A killer on the road: Circulating CD4(+) CD28 null T cells as cardiovascular risk factor in ESRD patients. J. Nephrol. 2012, 25, 183–191. [Google Scholar] [CrossRef]
- Litjens, N.H.R.; van Druningen, C.J.; Betjes, M.G.H. Progressive loss of renal function is associated with activation and depletion of naive T lymphocytes. Clin. Immunol. 2006, 118, 83–91. [Google Scholar] [CrossRef]
- Crépin, T.; Legendre, M.; Carron, C.; Vachey, C.; Courivaud, C.; Rebibou, J.M.; Ferrand, C.; Laheurte, C.; Vauchy, C.; Gaiffe, E.; et al. Uraemia-induced immune senescence and clinical outcomes in chronic kidney disease patients. Nephrol. Dial. Transplant. 2020, 35, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Lisowska, K.A.; Storoniak, H.; Dębska-Ślizień, A. T cell subpopulations and cytokine levels in hemodialysis patients. Hum. Immunol. 2022, 83, 134–143. [Google Scholar] [CrossRef]
- Borges, A.; Borges, M.; Fernandes, J.; Nascimento, H.; Sameiro-Faria, M.; Miranda, V.; Reis, F.; Belo, L.; Costa, E.; Santos-Silva, A. Apoptosis of peripheral CD4(+) T-lymphocytes in end-stage renal disease patients under hemodialysis and rhEPO therapies. Ren. Fail. 2011, 33, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Lisowska, K.A.; Pindel, M.; Pietruczuk, K.; Kuźmiuk-Glembin, I.; Storoniak, H.; Dębska-Ślizień, A.; Witkowski, J.M. The influence of a single hemodialysis procedure on human T lymphocytes. Sci. Rep. 2019, 9, 5041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisowska, K.A.; Dębska-Ślizień, A.; Jasiulewicz, A.; Heleniak, Z.; Bryl, E.; Witkowski, J.M. Hemodialysis affects phenotype and proliferation of CD4-positive T lymphocytes. J. Clin. Immunol. 2012, 32, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, F.; Cao, X.; Chen, X.; Zhang, Z.; Ding, X.; Zou, J.; Shen, B. Decreased Peripheral Naïve T Cell Number and Its Role in Predicting Cardiovascular and Infection Events in Hemodialysis Patients. Front. Immunol. 2021, 12, 644627. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Chung, B.H.; Jeon, E.J.; Kim, B.M.; Choi, B.S.; Park, C.W.; Kim, Y.S.; Cho, S.G.; Cho, M.L.; Yang, C.W. B cell-associated immune profiles in patients with end-stage renal disease (ESRD). Exp. Mol. Med. 2012, 44, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Esposito, P.; Rampino, T.; Gregorini, M.; Gabanti, E.; Bianzina, S.; Dal Canton, A. Mechanisms underlying sCD40 production in hemodialysis patients. Cell. Immunol. 2012, 278, 10–15. [Google Scholar] [CrossRef]
- Nowak, K.L.; Chonchol, M. Does inflammation affect outcomes in dialysis patients? Semin. Dial. 2018, 31, 388–397. [Google Scholar] [CrossRef]
- Kakuta, T.; Komaba, H.; Takagi, N.; Takahashi, Y.; Suzuki, H.; Hyodo, T.; Nagaoka, M.; Tanaka, R.; Iwao, S.; Ishida, M.; et al. A Prospective Multicenter Randomized Controlled Study on Interleukin-6 Removal and Induction by a new Hemodialyzer With Improved Biocompatibility in Hemodialysis Patients: A Pilot Study. Ther. Apher. Dial. 2016, 20, 569–578. [Google Scholar] [CrossRef]
- Kamińska, J.; Stopiński, M.; Mucha, K.; Jędrzejczak, A.; Gołębiowski, M.; Niewczas, M.A.; Pączek, L.; Foroncewicz, B. IL 6 but not TNF is linked to coronary artery calcification in patients with chronic kidney disease. Cytokine 2019, 120, 9–14. [Google Scholar] [CrossRef]
- Sepe, V.; Rampino, T.; Libetta, C. Arterial ‘inflammaging’ drives vascular calcification in children on dialysis. Kidney Int. 2019, 96, 522. [Google Scholar] [CrossRef]
- Li, P.; Schmidt, I.M.; Sabbisetti, V.; Tio, M.C.; Opotowsky, A.R.; Waikar, S.S. Plasma Endothelin-1 and Risk of Death and Hospitalization in Patients Undergoing Maintenance Hemodialysis. Clin. J. Am. Soc. Nephrol. 2020, 15, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, A.; Akazaki, S.; Nagano, Y.; Ueda, A.; Chang-Il Lee, M.; Aoyagi, K.; Oowada, S.; Sato, K. Hemodialysis raises oxidative stress through carbon-centered radicals despite improved biocompatibility. J. Clin. Biochem. Nutr. 2021, 69, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Duni, A.; Liakopoulos, V.; Roumeliotis, S.; Peschos, D.; Dounousi, E. Oxidative Stress in the Pathogenesis and Evolution of Chronic Kidney Disease: Untangling Ariadne’s Thread. Int. J. Mol. Sci. 2019, 20, 3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wann, J.G.; Hsu, Y.H.; Yang, C.C.; Lin, C.S.; Tai, D.W.; Chen, J.S.; Hsiao, C.W.; Chen, C.F. Neutrophils in acidotic haemodialysed patients have lower intracellular pH and inflamed state. Nephrol. Dial. Transpl. 2007, 22, 2613–2622. [Google Scholar] [CrossRef] [Green Version]
- Morena, M.; Cristol, J.P.; Senécal, L.; Leray-Moragues, H.; Krieter, D.; Canaud, B. Oxidative stress in hemodialysis patients: Is NADPH oxidase complex the culprit? Kidney Int. Suppl. 2002, 80, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, M.; Hung, A.; Kovalchuk, O.; Bitzer, M.; Mokrzycki, M.H. The initial vascular access type contributes to inflammation in incident hemodialysis patients. Int. J. Nephrol. 2012, 2012, 917465. [Google Scholar] [CrossRef]
- Crespo-Montero, R.; Gómez-López, V.E.; Guerrero-Pavón, F.; Carmona-Muñoz, A.; Romero-Saldaña, M.; Ranchal-Sanchez, A.; Aljama-García, P. Influence of Tunneled Hemodialysis-Catheters on Inflammation and Mortality in Dialyzed Patients. Int. J. Environ. Res. Public Health 2021, 18, 7605. [Google Scholar] [CrossRef]
- Suvakov, S.; Jerotic, D.; Damjanovic, T.; Milic, N.; Pekmezovic, T.; Djukic, T.; Jelic-Ivanovic, Z.; Savic Radojevic, A.; Pljesa-Ercegovac, M.; Matic, M.; et al. Markers of Oxidative Stress and Endothelial Dysfunction Predict Haemodialysis Patients Survival. Am. J. Nephrol. 2019, 50, 115–125. [Google Scholar] [CrossRef]
- Diaz-Ricart, M.; Torramade-Moix, S.; Pascual, G.; Palomo, M.; Moreno-Castano, A.B.; Martinez-Sanchez, J.; Vera, M.; Cases, A.; Escolar, G. Endothelial Damage, Inflammation and Immunity in Chronic Kidney Disease. Toxins 2020, 12, 361. [Google Scholar] [CrossRef]
- Falconi, C.A.; Junho, C.; Fogaça-Ruiz, F.; Vernier, I.; da Cunha, R.S.; Stinghen, A.; Carneiro-Ramos, M.S. Uremic Toxins: An Alarming Danger Concerning the Cardiovascular System. Front. Physiol. 2021, 12, 686249. [Google Scholar] [CrossRef]
- Sun, Y.; Johnson, C.; Zhou, J.; Wang, L.; Li, Y.F.; Lu, Y. Uremic toxins are conditional danger- or homeosta-sis-associated molecular patterns. Front. Biosci. Landmark 2018, 23, 348–387. [Google Scholar]
- Addi, T.; Poitevin, S.; McKay, N.; El Mecherfi, K.E.; Kheroua, O.; Jourde-Chiche, N. Mechanisms of tissue factor induction by the uremic toxin indole-3 acetic acid through aryl hydrocarbon receptor/nuclear factor-kappa B signaling pathway in human endothelial cells. Arch. Toxicol. 2019, 93, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Düsing, P.; Zietzer, A.; Goody, P.R.; Hosen, M.R.; Kurts, C.; Nickenig, G.; Jansen, F. Vascular pathologies in chronic kidney disease: Pathophysiological mechanisms and novel therapeutic approaches. J. Mol. Med. 2021, 99, 335–348. [Google Scholar] [CrossRef]
- Rapa, S.F.; Prisco, F.; Popolo, A.; Iovane, V.; Autore, G.; Di Iorio, B.R.; Dal Piaz, F.; Paciello, O.; Nishijima, F.; Marzocco, S. Pro-Inflammatory Effects of Indoxyl Sulfate in Mice: Impairment of Intestinal Homeostasis and Immune Response. Int. J. Mol. Sci. 2021, 22, 1135. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rysz, J.; Franczyk, B.; Ławiński, J.; Olszewski, R.; Ciałkowska-Rysz, A.; Gluba Brzózka, A.J.T. The Impact of CKD on Uremic Toxins and Gut Microbiota. Toxins 2021, 13, 252. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, S.; Li, S.; Zhao, L.; Hao, Y.; Qin, J.; Zhang, L.; Zhang, C.; Bian, W.; Zuo, L.; et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut 2020, 69, 2131–2142. [Google Scholar] [CrossRef]
- Lau, W.L.; Chang, Y.; Vaziri, N.D. The consequences of altered microbiota in immune-related chronic kidney disease. Nephrol. Dial. Transpl. 2021, 27, 1791–1798. [Google Scholar] [CrossRef]
- De Mauri, A.; Carrera, D.; Vidali, M.; Bagnati, M.; Rolla, R.; Riso, S.; Torreggiani, M.; Chiarinotti, D. Compliance, Adherence and Concordance Differently Predict the Improvement of Uremic and Microbial Toxins in Chronic Kidney Disease on Low Protein Diet. Nutrients 2022, 14, 487. [Google Scholar] [CrossRef]
- Calaf, R.; Cerini, C.; Génovésio, C.; Verhaeghe, P.; Jourde-Chiche, N.; BergéLegranc, D. Determination of uremic so-lutes in biological fluids of chronic kidney disease by HPLC assay. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2011, 879, 2281–2286. [Google Scholar] [CrossRef]
- Dellepiane, S.; Marengo, M.; D’Arezzo, M.; Donati, G.; Fabbrini, P.; Lacquaniti, A.; Ronco, C.; Cantaluppi, V. The Next Evolution of HemoDialysis eXpanded: From a Delphi Questionnaire-Based Approach to the Real Life of Italian Dialysis Units. Blood Purif. 2022, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jang, M.J.; Jeon, J.; Lee, J.E.; Huh, W.; Choi, B.S.; Park, C.W.; Chin, H.J.; Kang, C.L.; Kim, D.K.; et al. Cardiovascular Risk Comparison between Expanded Hemodialysis Using Theranova and Online Hemodiafiltration (CARTOON): A Multicenter Randomized Controlled Trial. Sci. Rep. 2021, 11, 10807. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Xiao, L.; Xu, B.; Xu, X.X.; Liu, F.Y.; Sun, L. Effects of vitamin E-coated dialyzer on oxidative stress and inflammation status in hemodialysis patients: A systematic review and meta-analysis. Ren. Fail. 2014, 36, 722–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepe, V.; Gregorini, M.; Rampino, T.; Esposito, P.; Coppo, R.; Galli, F.; Libetta, C. Vitamin e-loaded membrane dialyzers reduce hemodialysis inflammaging. BMC Nephrol. 2019, 20, 412. [Google Scholar] [CrossRef]
- Bensaadi, S.; Arous, O.; Kerdjoudj, H.; Amara, M. Evaluating molecular weight of PVP on characteristics of CTA membrane dialysis. J. Environ. Chem. Eng. 2016, 4, 1545–1554. [Google Scholar] [CrossRef]
- Venkatesh, K.; Arthanareeswaran, G.; Suresh Kumar, P.; Kweon, J. Fabrication of Zwitterion TiO2 Nano-material-Based Nanocomposite Membranes for Improved Antifouling and Antibacterial Properties and Hemocompatibility and Reduced Cytotoxicity. ACS Omega 2021, 6, 20279–20291. [Google Scholar] [CrossRef]
- Meyer, J.M.; Steer, D.; Weber, L.A.; Zeitone, A.A.; Thakuria, M.; Ho, C.H.; Aslam, S.; Mullon, C.; Kossmann, R.J. Safety of a Novel Dialyzer Containing a Fluorinated Polyurethane Surface-Modifying Macromolecule in Patients with End-Stage Kidney Disease. Blood Purif. 2021, 50, 959–967. [Google Scholar] [CrossRef]


{kind=link}
| Pathway | Mechanisms | Potential Biomarkers | Potential Therapy |
|---|---|---|---|
| Complement System | Activation of the complement response, inducing pro-coagulant state, releasing inflammatory mediators from immune cells with vascular endothelial damage and atherosclerosis Reduction in expression of complement inhibitors due to dialyzer absorption (CFH, an inhibitor of C3 convertase and C3b, ficolin-2) [45,46,47] | Ficolin-2 reduction C3a increase C5a increase C5b increase Soluble C5b9 increase C3d/C3 ratio increase | HDF/MCO/ HDx [48] Anticoagulant citrate [49] Polysulfone grafted with argatroban [51] |
| Innate Immunity | Decrease in neutrophils due to activation and apoptosis [53] Monocyte (CD14++/CD16+) Mo2 and Mo3 phenotypes attach to endothelial cells, contributing to inflammation and endothelial damage | HMGB1 Calprotectin NETs [54] Monocyte subpopulations | Online HDF [61,62] |
| Acquired Immunity | Reduced and not-functional naïve T cells, Th2 and regulatory T cells, showing a pro-inflammatory phenotype [67] Altered B lymphocytes with increased high differentiated forms and a reduction in naïve cells [77] | T-cell lymphopenia increased CD4+/CD8+ Increased soluble CD40 | No data |
| Coagulation System and Platelet Activity | Pro-thrombotic status mediated by activated intrinsic and extrinsic pathways leading to pro-inflammatory effects and endothelial cell damage Platelet dysfunction with atypical activation | D-dimer, β-TG TAT | Anticoagulant citrate [39,40] Polyvinylpyrrolidone [37,38] |
| Oxidative Stress | Loss of balance between pro-oxidant factors and antioxidant mechanisms | ROS dosage [80,81] Oxidized lipids AOPP | Vitamin-E-coated filter [106] |
| Uremic Toxin | Endothelial dysfunction Inflammation Immune dysfunction | FLC, microbiota-derived uremic toxins, lipoprotein-associated phospholipase A2 | HDF/MCO/HDx [103,104] |
| Future Perspectives | |||
| Materials | Mechanisms | Potential Biomarkers | Effects |
| Vitamin-E-coated filter | Enhanced ROS scavenger activity | ROS NO IDO1 [106] | Anti-inflammatory antioxidative |
| PVP and PEG filters | Enhanced membrane morphology and reduced adsorption/adhesion of macromolecules [107] | No data | Anti-inflammatory antithrombotic |
| PES filter grafted with zwitterion | Anti-fouling activity and high biocompatibility in terms of protein absorption and platelet adhesion [108] | No data | Anti-inflammatory antithrombotic |
| Endexo | Reduced adhesion and activation of blood proteins and platelets [109] | No data | Anti-inflammatory antithrombotic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campo, S.; Lacquaniti, A.; Trombetta, D.; Smeriglio, A.; Monardo, P. Immune System Dysfunction and Inflammation in Hemodialysis Patients: Two Sides of the Same Coin. J. Clin. Med. 2022, 11, 3759. https://doi.org/10.3390/jcm11133759
Campo S, Lacquaniti A, Trombetta D, Smeriglio A, Monardo P. Immune System Dysfunction and Inflammation in Hemodialysis Patients: Two Sides of the Same Coin. Journal of Clinical Medicine. 2022; 11(13):3759. https://doi.org/10.3390/jcm11133759
Chicago/Turabian StyleCampo, Susanna, Antonio Lacquaniti, Domenico Trombetta, Antonella Smeriglio, and Paolo Monardo. 2022. "Immune System Dysfunction and Inflammation in Hemodialysis Patients: Two Sides of the Same Coin" Journal of Clinical Medicine 11, no. 13: 3759. https://doi.org/10.3390/jcm11133759
APA StyleCampo, S., Lacquaniti, A., Trombetta, D., Smeriglio, A., & Monardo, P. (2022). Immune System Dysfunction and Inflammation in Hemodialysis Patients: Two Sides of the Same Coin. Journal of Clinical Medicine, 11(13), 3759. https://doi.org/10.3390/jcm11133759