
The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis




Abstract
:1. Introduction
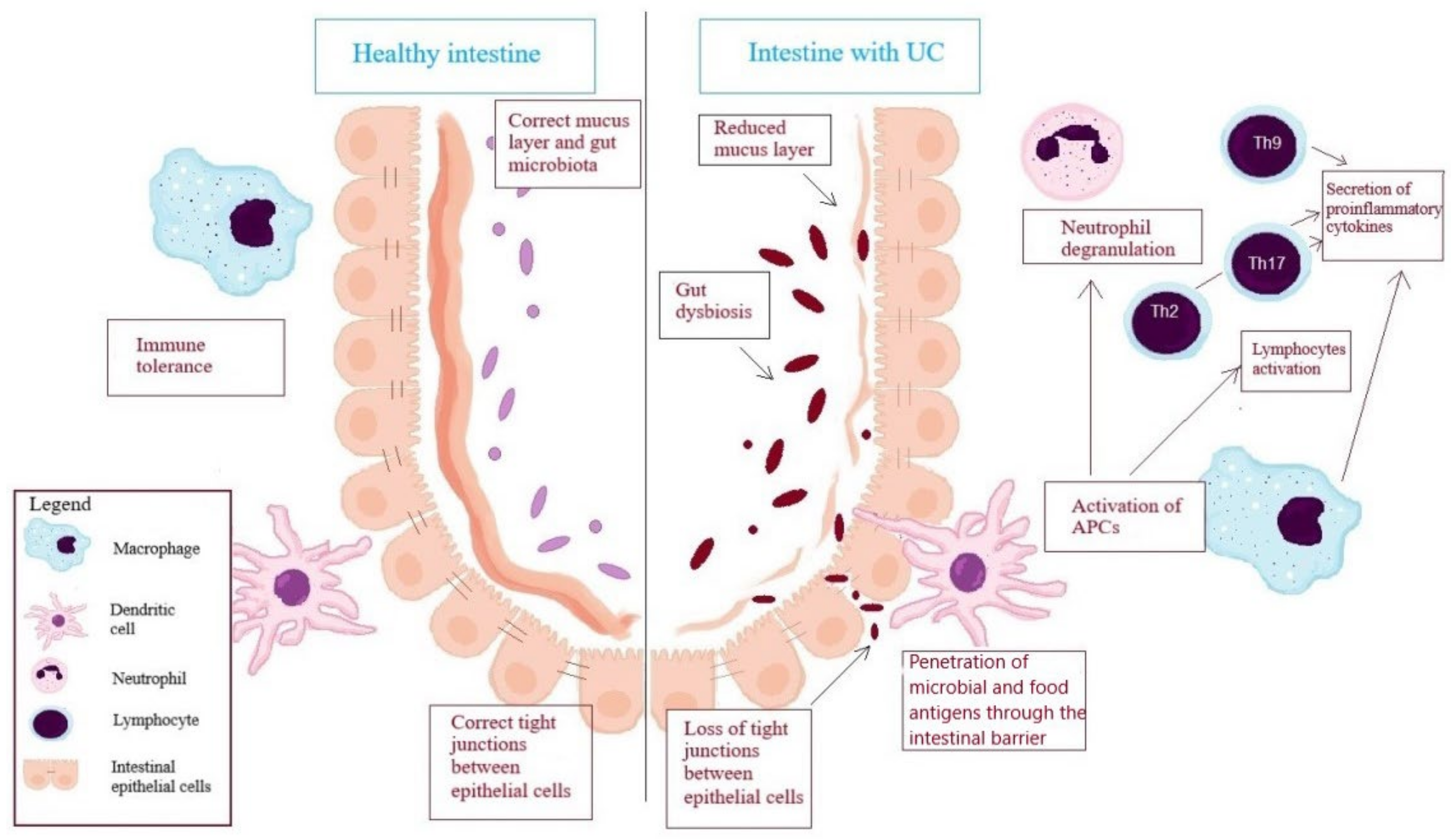
2. Immune Cells in the Pathogenesis of Ulcerative Colitis
2.1. Cells of Innate Immunity
2.2. Antigen-Presenting Cells (APC)
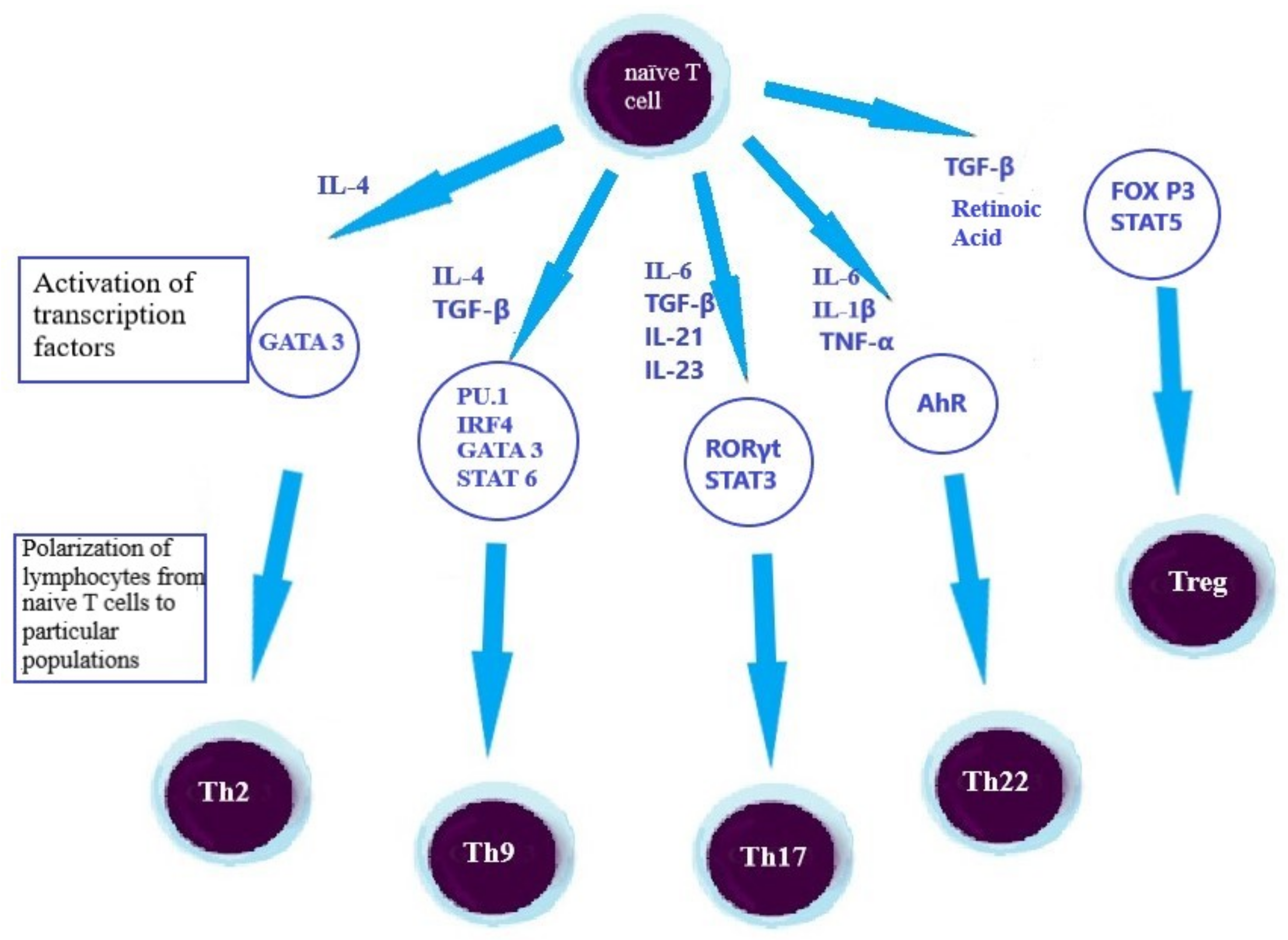
2.3. Lymphocytes, as an Element of the Adaptive Immune Response in the UC’s Pathogenesis
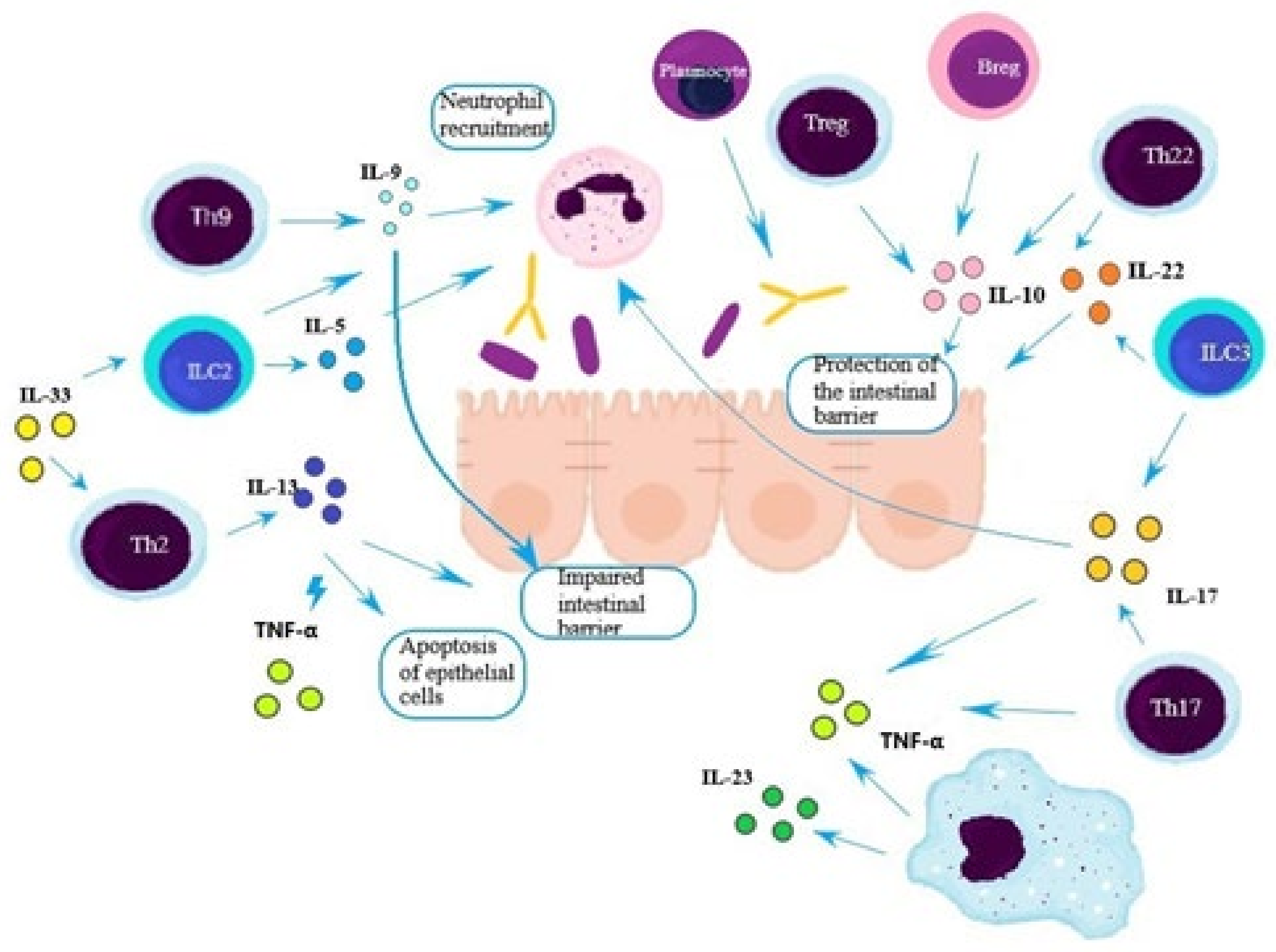
2.4. Th2 Lymphocytes
2.5. Th9 Lymphocytes
2.6. Th17 Lymphocytes
2.7. Th22 Lymphocytes
2.8. Treg Lymphocytes
2.9. B Lymphocytes/Plasmocytes
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Porter, R.J.; Kalla, R.; Ho, G.-T. Ulcerative colitis: Recent advances in the understanding of disease pathogenesis. F100 Res. 2020, 9, 294. [Google Scholar] [CrossRef] [PubMed]
- Teigen, L.M.; Geng, Z.; Sadowsky, M.J.; Vaughn, B.P.; Hamilton, M.J.; Khortus, A. Dietary factors in sulfur metabolism and pathogenesis of ulcerative colitis. Nutrients 2019, 11, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.; Bäumler, A.J. Dysbiotic Proteobacteria expansion: A microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Kmieć, Z.; Cyman, M.; Ślebioda, T.J. Cells of the innate and adaptive immunity and their interactions in inflammatory bowel disease. Adv. Med. Sci. 2017, 62, 1–16. [Google Scholar] [CrossRef]
- McCauley, H.A.; Guasch, G. Three cheers for the goblet cell: Maintaing homeostasis in mucosal epithelia. Trends Mol. Med. 2015, 21, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, A.; Goggolidou, P. Ulcerative colitis: Understanding its cellular pathology could provide insight into novel therapies. J. Infalmm. 2020, 17, 15. [Google Scholar] [CrossRef] [Green Version]
- Wejman, J.; Bartnik, W. Atlas Kliniczno-Patologiczny Nieswoistych Chorób Zapalnych Jelit; Wydawnictwa Medyczne Termedia: Warsaw, Poland, 2011; pp. 53–54, 65–73. [Google Scholar]
- Shohan, M.; Elahi, S.; Shirzad, H.; Rafieian-Kopaei, M.; Bagheri, N.; Soltani, E. Th9 cells: Probable players in ulcerative colitis pathogenesis. Int. Rev. Immunol. 2018, 37, 192–205. [Google Scholar] [CrossRef]
- Shohan, M.; Sabzevary-Ghahfarokhi, M.; Bagheri, N.; Shirzad, H.; Rahimian, G.; Soltani, A.; Ghatreh-Samani, M.; Deris, F.; Tahmasbi, K.; Shahverdi, E.; et al. Intensified Th9 response is associated with the immunopathogenesis of active ulcerative colitis. Immunol. Investig. 2018, 47, 700–711. [Google Scholar] [CrossRef]
- Polińska, B.; Matowicka-Karna, J.; Kemona, H. Cytokiny w nieswoistych zapalnych chorobach jelit. Postepy Hig. Med. Dosw. 2009, 63, 389–394. [Google Scholar]
- Szulc-Dąbrowska, L.; Gieryńska, M.; Depczyńska, D.; Schollenberger, A.; Toka, F.N. Limfocyty Th17 w zakażeniach bakteryjnych. Postepy Hig. Med. Dosw. 2015, 69, 398–417. [Google Scholar] [CrossRef]
- Lee, S.H.; Kwon, J.E.; Cho, M.-L. Immunological pathogenesis of inflammatory bowel disease. Intest. Res. 2018, 16, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Geremia, A.; Biancheri, P.; Allan, P.; Corazza, G.R.; Di Sabatino, A. Innate and adaptive immunity in inflammatory bowel disease. Autoimmun. Rev. 2013, 13, 3–10. [Google Scholar] [CrossRef]
- Miller, A.M. Role of IL-33 in inflammation and disease. J. Inflamm. 2011, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, Z.H.; Bogdanovski, D.A.; Barratt-Stopper, P.; Paglinco, S.R.; Antonioli, L.; Rolandelli, R.H. Crohn’s disease and ulcerative colitis show unique cytokine profiles. Cureus 2017, 9, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinallo, V.; Marafini, I.; Di Fusco, D.; Laudisi, F.; Franze, E.; Di Grazia, A.; Figliuzzi, M.M.; Caprioli, F.; Stolfi, C.; Monteleone, I.; et al. Neutrophil extracellular traps sustain inflammatory signals in ulcerative colitis. J. Crohn’s Colitis 2019, 13, 772–784. [Google Scholar] [CrossRef]
- Matoszka, N.; Działo, J.; Tokarz-Deptuła, B.; Deptuła, W. NET i NEToza—Nowe zjawisko w immunologii. Postepy Hig. Med. Dosw. 2012, 66, 437–445. [Google Scholar] [CrossRef]
- Forkel, M.; van Tol, S.; Höög, C.; Michaëlsson, J.; Almer, S.; Mjösberg, J. Distinct alterations in the composition of mucosal innate lymphoid cells in newly diagnosed and established Crohn’s disease and ulcerative colitis. J. Crohns Colitis 2019, 13, 67–78. [Google Scholar] [CrossRef]
- Ordas, I.; Eckmann, L.; Talamini, M.; Baumgart, D.; Sandborn, W.J. Ulcerative Colitis. Lancet 2012, 380, 1606–1619. [Google Scholar] [CrossRef] [Green Version]
- Forkel, M.; Mjösberg, J. Dysregulation of group 3 innate lymphoid cells in the pathogenesis of inflammatory bowel disease. Curr. Allergy Asthma Rep. 2016, 16, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, A.L.; Al-Hassi, H.O.; Rigby, R.J.; Bell, S.J.; Emmanuel, A.V.; Knight, S.C.; Kamm, M.A.; Stagg, A.J. Characteristic of Intestinal Dendritic Cells in Inflammatory Bowel Diseases. Gastroenterology 2005, 129, 50–65. [Google Scholar] [CrossRef]
- Kappeler, A.; Mueller, C. The role of activated cytotoxic T cells in inflammatory bowel disease. Histol. Histopathol. 2000, 15, 167–172. [Google Scholar] [PubMed]
- Rabe, H.; Malmquist, M.; Barkman, C.; Östman, S.; Gjertsson, I.; Saalman, R.; Wold, A.E. Distinct patterns of naive, activated and memory T and B cells in blood of patients with ulcerative colitis or Crohn’s disease. Clin. Expl. Immunol. 2019, 197, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerlach, K.; Hwang, Y.; Nikolaev, A.; Atreya, R.; Dornhoff, H.; Steiner, S.; Lehr, H.-A.; Wirtz, S.; Vieth, M.; Waisman, A.; et al. Th9 cells that express the trancription factor PU.1 drive T cel-mediated colitis via IL-9 receptor signaling in intestinal epithelial cells. Nat. Immunol. 2014, 15, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Bamias, G.; Cominelli, F. Role of Th2 immunity in intestinal inflammation. Curr. Opin. Gastroenterol. 2015, 31, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Heller, F.; Fromm, A.; Gitter, A.H.; Mankertz, J.; Schulzke, J.-D. Epithelial apoptosis is a prominent feature of the epithelial barier disturbance in intedtinal inflammation: Effect of pro-inflammatory interlekin-13 on the epithelial cell function. Mucosal. Immunol. 2008, 1, S58–S61. [Google Scholar] [CrossRef] [Green Version]
- Raju, P.; Shashikanth, N.; Tsai, P.-Y.; Pongkorpsakol, P.; Chanez-Paredes, S.; Steinhagen, P.R.; Kuo, W.-T.; Singh, G.; Tsukita, S.; Turner, J.R. Inactivation of paracellular cation-selective claudin-2 channels attenuates immune-mediated experimental colitis in mice. J. Clin. Investig. 2020, 130, 5197–5208. [Google Scholar] [CrossRef]
- Seidelin, J.B.; Coskun, M.; Kvist, P.H.; Holm, T.L.; Holgersen, K.; Nielsen, O.H. IL-33 promotes GATA-3 polarization of gut-derived T cells in experimental and ulcerative colitis. J. Gastroenterol. 2015, 50, 180–190. [Google Scholar] [CrossRef]
- Travers, J.; Rochman, M.; Miracle, C.E.; Habel, J.E.; Brusilovsky, M.; Caldwell, J.M.; Rymer, J.K.; Rothenberg, M.E. Chromatin regulates IL-33 release and extracellular cytokine activity. Nat. Commun. 2018, 9, 3244. [Google Scholar] [CrossRef]
- He, Z.; Chen, L.; Furtado, G.C.; Lira, S.A. Interleukin 33 regulates gene expression in intestinal epithelial independently of its nuclear localization. Cytokine 2018, 111, 146–153. [Google Scholar] [CrossRef]
- Akimoto, M.; Takenaga, K. Role of the IL-33/ST2L axis in colorectal cancer progression. Cell Immunol. 2019, 343, 103740. [Google Scholar] [CrossRef] [PubMed]
- Hodzic, Z.; Schill, E.M.; Bolock, A.M.; Good, M. IL-33 and the intestine: The good, the bad and the inflammatory. Cytokine 2017, 100, 1–10. [Google Scholar] [CrossRef]
- Bessa, J.; Meyer, C.A.; de Vera Mudry, M.C.; Schliht, S.; Smith, S.H.; Iglesias, A.; Cote-Sierra, J. Altered subcellular localization of IL-33 leads to non-resolving lethal inflammation. J. Autoimmun. 2014, 55, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Luettig, J.; Rosenthal, R.; Barmeyer, C.; Schulzke, J.D. Claudin-2 as a mediator of leaky gut barier during intestinal inflammation. Tissue Barriers 2015, 3, e977176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Lu, Q.; Liu, Y.; Shi, Z.; Hu, L.; Zheng, Z.; Tu, Y.; Xiao, Z.; Xu, Q. Th17 zells in Inflammatory Bowel Disease: Cytokines, plasticity, therapies. J. Immunol. Res. 2021, 2021, 8816041. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulan, F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Chen, K.; Ayanoglu, G.; et al. IL-23 independent IL-17 production regulates intestinal epithelial permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.M.; Davenport, M.; Wolff, M.J.; Wiens, K.E.; Abidi, W.M.; Poles, M.A.; Cho, I.; Ullman, T.; Mayer, L.; Loke, P. IL-22-producing CD4+ cells are depleted in actively inflamed colitis tissue. Mucosal. Immunol. 2014, 7, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Targeting immune cell circuits and trafficking in inflammatory bowel disease. Nat. Immunol. 2019, 20, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Woźniak-Stolarska, B.; Sajewicz, Z.; Błachut, K. Serum concentration of interleukin-10 in inflammatory bowel disease. Adv. Clin. Exp. Med. 2003, 12, 717–721. [Google Scholar]
- Asselin, C.; Gendron, F.-P. Shuttling of information between the mucosal and luminal environment drives intestinal homeostasis. FEBS Lett. 2014, 588, 4148–4157. [Google Scholar] [CrossRef]
- Maśliński, W.; Kontny, E. Podstawy Immunologii dla Reumatologów; Wydawnictwo Narodowego Instytutu Geriatrii, Reumatologii i Rehabilitacji im: Warsaw, Poland, 2015; pp. 77–91. [Google Scholar]
- Mizoguchi, A.; Bhan, A.K. Immunobiology of B cells in inflammatory bowel disease. In Crohn’s Disease and Ulcerative Colitis; Baumgart, D., Ed.; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, Y.; Zhang, M.; Wang, H.; Jiang, Y.; Gao, P. Ulcerative colitis is chracterized by a decrease in regulatory B cells. J. Crohns Colitis. 2016, 10, 1212–1223. [Google Scholar] [CrossRef] [Green Version]
- Uo, M.; Hisamatsu, T.; Miyoshi, J.; Kaito, D.; Yoneno, K.; Kitazume, M.T.; Mori, M.; Sugita, A.; Koganei, K.; Matsuoka, K.; et al. Mucosal CXCR4+ IgG plasma cells contribute to pathogenesis of human ulcerative colitis through FcγR-mediated CD14 macrophage activation. Gut 2013, 62, 1734–1744. [Google Scholar] [CrossRef] [PubMed]
- Castro-Dopico, T.; Dennison, T.W.; Ferdinand, J.R.; Clare, S.; Parkes, M.; Clatworthy, M.R. Anti-comensal IgG drives intestinal inflammation and type 17 immunity in ulcerative colitis. Immunity 2019, 50, 1099–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Dopico, T.; Clatworthy, M.R. IgG and Fcγ receptors in intestinal immunity and inflammation. Front. Immunol. 2019, 10, 805. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cytokine | Expression of Cytokine in UC | Cells Secreting Cytokine | Influence on the Inflammation | Function |
|---|---|---|---|---|
| IL-1β | Increased | Macrophages | pro-inflammatory | Stimulates T cells to secrete pro-inflammatory cytokines, induces chemotactic factors, influences the proliferation of B cells, systemically causes an increase of temperature and acute phase proteins |
| IL-4 | Th2 | anti-inflammatory | Stimulates humoral immune response, inhibits the secretion of pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), activates M2 macrophages, activates transcription factors responsible for differentiation of T CD4+ cells to Th9 cells | |
| IL-5 | Increased | Th2 | Decreases the activity of Th1 cells and cellular immune response, stimulates the maturation of eosinophils and basophils | |
| IL-6 | Increased | Macrophages, dendritic cells | pro-inflammatory | Activates transcription factors responsible for differentiation of T CD4+ cells to Th17 cells, inhibits the secretion of TNF-α |
| IL-9 | Increased | Th9, Th2 | pro-inflammatory | Activates mast cells, neutrophils and eosinophils, influences the expression of proteins creating tight junctions in the intestinal barrier |
| IL-10 | Increased | Treg, Th2, Th17, Breg | anti-inflammatory | Inhibits the secretion of tissue metalloproteinases, tissue factor and cyclooxygenase 2, suppresses the expression of transcription factor NF-κB, activates macrophages M2 |
| IL-13 | Increased | Th2 | anti-inflammatory | Decreases the activity of Th1 cells and cellular immune response, inhibits the secretion of pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), influences the expression of proteins creating tight junctions in the intestinal barrier and the epithelial cells apoptosis, activates macrophages M2 |
| IL-17 | Increased | Th17, monocytes, neutrophils, T CD8+, NK cells | pro-inflammatory | Stimulates monocytes, epithelial and endothelial cells to secrete pro-inflammatory cytokines (IL-1β, TNF-α) and chemokines responsible for leukocytes and neutrophils migration to inflamed tissues, in the absence of IL-23 supports the intestinal barrier through occludin regulation in tight junctions |
| IL-21 | Th17, Th2 | Decreases the activity of Th1 cells and thus the cellular immune response, increases the expression of IL-23 receptor, stimulates proliferation and maturation of B, T CD8+ and NK cells | ||
| IL-22 | Th17, Th22, Th1 | Induces the secretion of antimicrobial peptides, IL-10 and mucus, mediates in the tissue repair processes | ||
| IL-23 | Increased | Macrophages, dendritic cells | pro-inflammatory | Takes part in the differentiation of T CD4+ cells to Th17 cells |
| IL-25 | Th2 | Decreases the activity of Th1 cells and cellular immune response | ||
| IL-33 | Treg, macrophages, dendritic cells, mast cells | Enhances the secretion of IL-4, IL-5, IL-13, increases the accumulation of Th2, stimulates pathogenic Th2 and Th17 response, induces tissue repair through coordination of Treg | ||
| IL-35 | Treg | anti-inflammatory | Suppress the differentiation of Th17 | |
| TGF-β | Treg | anti-inflammatory | Stimulates epithelial repair, decreases expression of IL-33 and Th22, stimulates differentiation of Th17 in the presence of IL-6 and Treg in the absence of IL-6, activates the transcription factors responsible for differentiation of T CD4+ cells to Th9 cells | |
| IFN-γ | Th1, dendritic cells, macrophages | Enhances transcytosis and paracellular transport, activates macrophages | ||
| TNF-α | Increased | Th17, macrophages, dendritic cells | pro-inflammatory | Takes part in cell apoptosis, stimulates lymphocytes and activates other immune cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kałużna, A.; Olczyk, P.; Komosińska-Vassev, K. The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis. J. Clin. Med. 2022, 11, 400. https://doi.org/10.3390/jcm11020400
Kałużna A, Olczyk P, Komosińska-Vassev K. The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis. Journal of Clinical Medicine. 2022; 11(2):400. https://doi.org/10.3390/jcm11020400
Chicago/Turabian StyleKałużna, Aleksandra, Paweł Olczyk, and Katarzyna Komosińska-Vassev. 2022. "The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis" Journal of Clinical Medicine 11, no. 2: 400. https://doi.org/10.3390/jcm11020400
APA StyleKałużna, A., Olczyk, P., & Komosińska-Vassev, K. (2022). The Role of Innate and Adaptive Immune Cells in the Pathogenesis and Development of the Inflammatory Response in Ulcerative Colitis. Journal of Clinical Medicine, 11(2), 400. https://doi.org/10.3390/jcm11020400