
Epitope Mapping of Pathogenic Autoantigens on Sjögren’s Syndrome-Susceptible Human Leukocyte Antigens Using In Silico Techniques


Abstract
:1. Introduction
2. Materials and Methods
2.1. In Silico Binding Affinities of Peptides for HLA-DR3 and Other Risk Alleles
2.2. PDB Structures of HLA-DR3 and Predicted Peptide Docking
2.3. Homology Determination for Viral and Bacterial Peptides
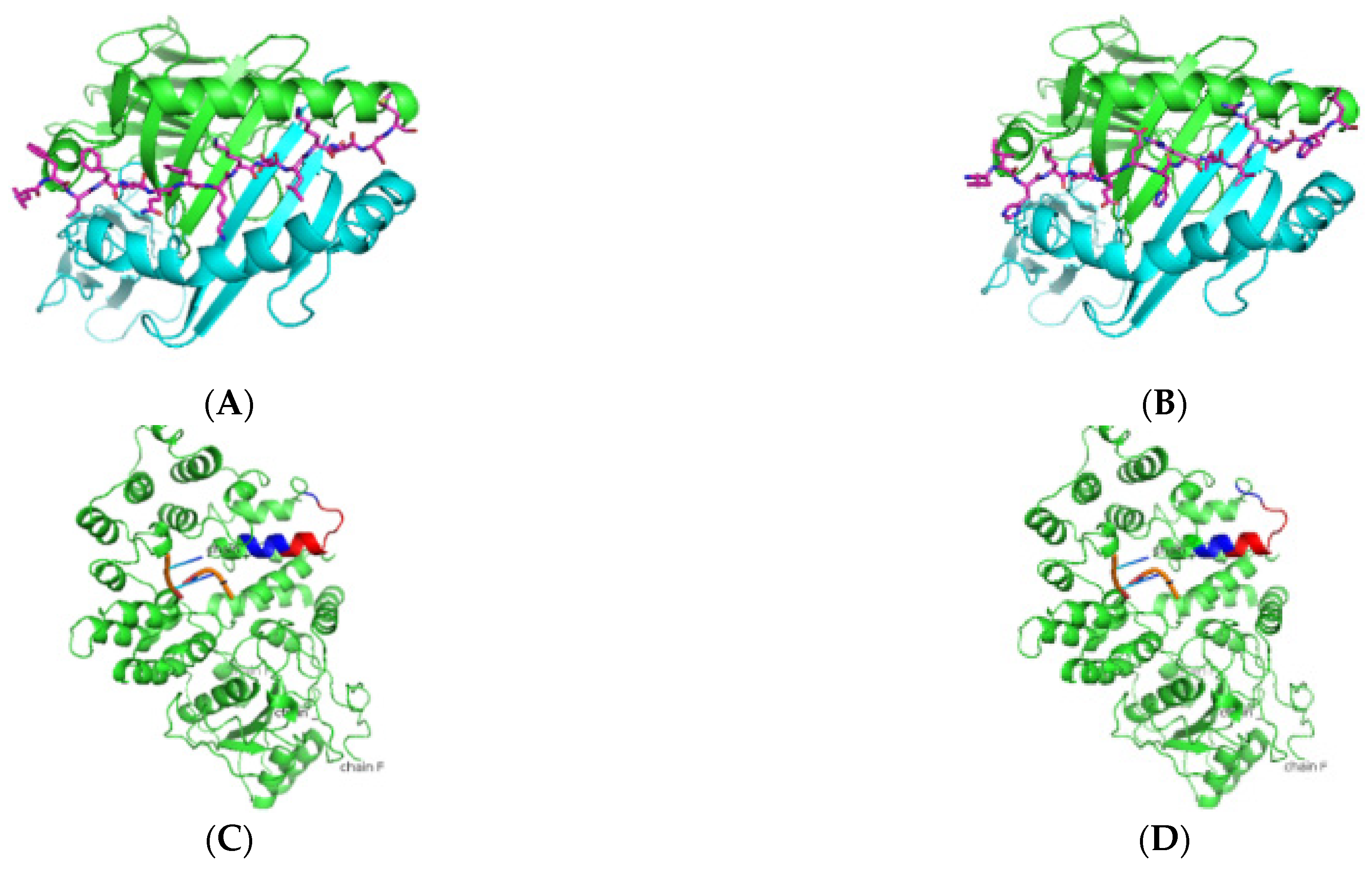
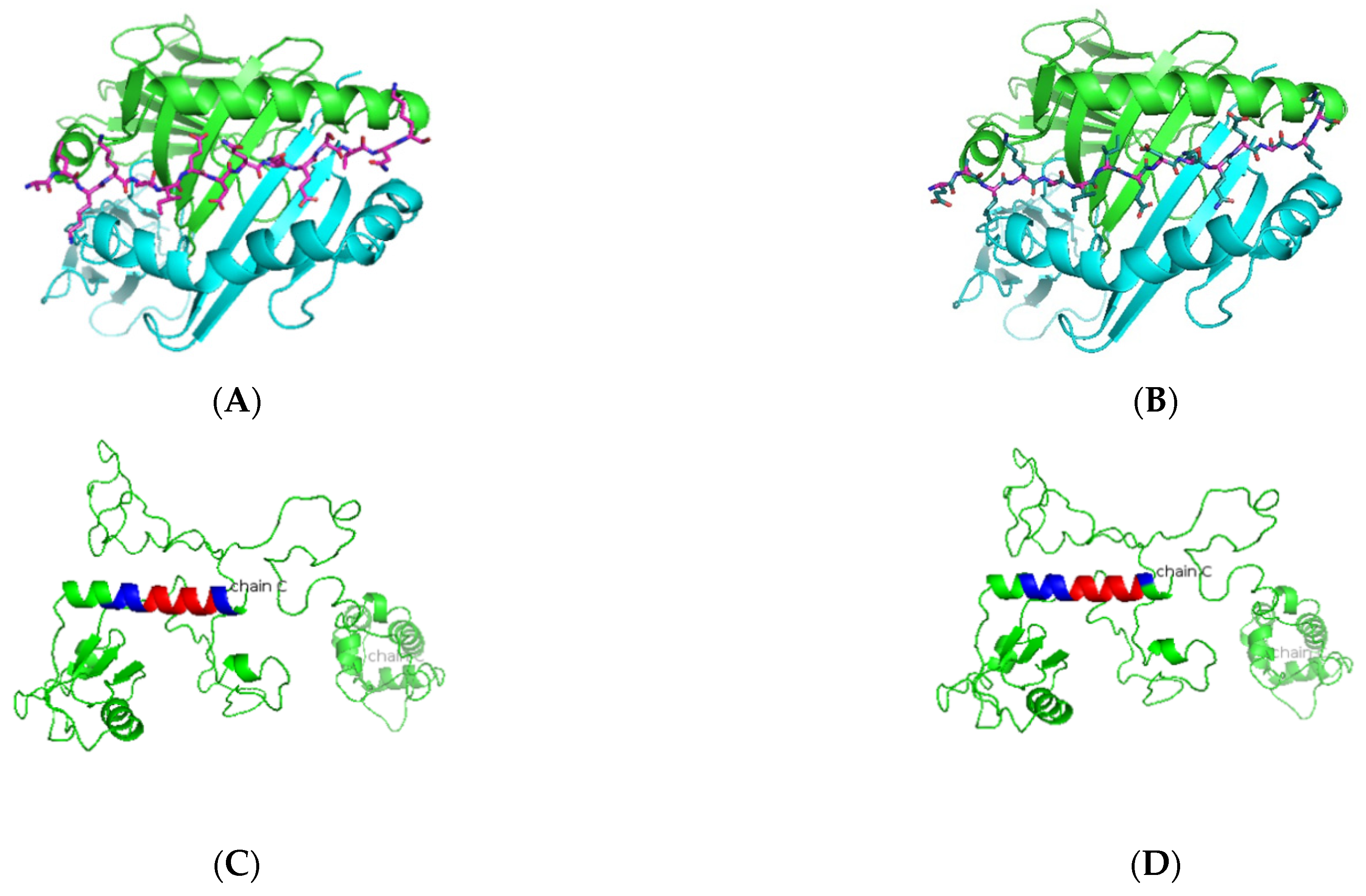
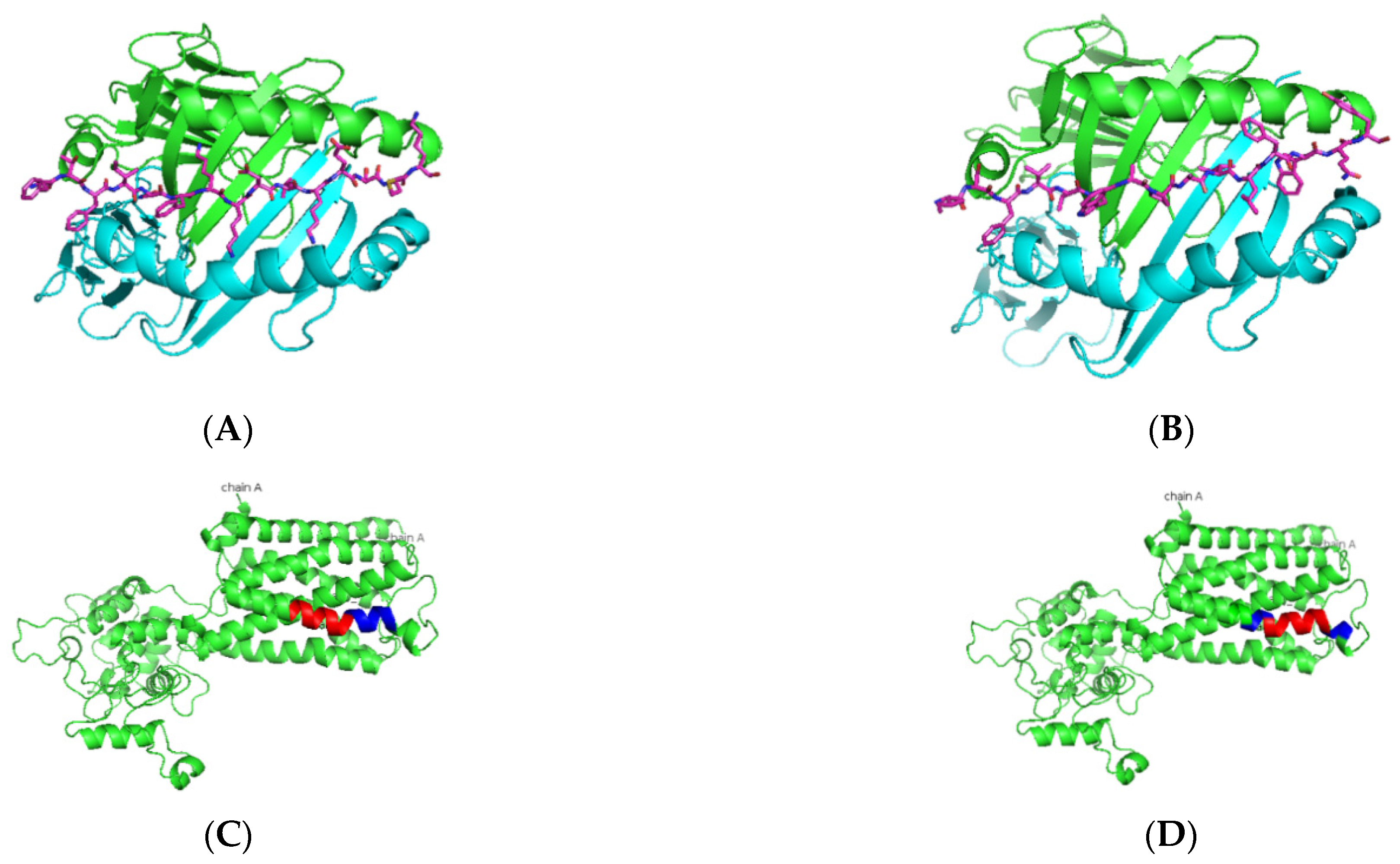
3. Results
3.1. In Silico Antigenic Mapping of High-Risk Autoantigens Presented on HLA-DR3
3.2. Elucidating the Nature of Predicted Peptides Presented on Other Risk Alleles
3.3. Homology of Predicted Peptides Binding to HLA-DRB1*03:01 to Viral and Bacterial Proteins
3.4. Homology of Predicted Peptides Binding to Other Risk HLA Alleles to Viral and Bacterial Proteins
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, C.Q.; Sharma, A.; Lee, B.H.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expression in the salivary gland during development and onset of xerostomia in Sjogren’s syndrome-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Arthritis Res. Ther. 2009, 11, R56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmick, C.G.; Felson, D.; Lawrence, R.C.; Gabriel, S.; Hirsch, R.; Kwoh, C.K.; Liang, M.H.; Kremers, H.M.; Mayes, M.D.; Merkel, P.A.; et al. Estimates of the prevalence of arthritis and other rheumatic conditions in the United States: Part I. Arthritis Care Res. 2007, 58, 15–25. [Google Scholar] [CrossRef]
- Vivino, F.B.; Bunya, V.Y.; Massaro-Giordano, G.; Johr, C.R.; Giattino, S.L.; Schorpion, A.; Shafer, B.; Peck, A.; Sivils, K.; Rasmussen, A.; et al. Sjogren’s syndrome: An update on disease pathogenesis, clinical manifestations and treatment. Clin. Immunol. 2019, 203, 81–121. [Google Scholar] [CrossRef]
- Thomas, E.; Hay, E.M.; Hajeer, A.; Silman, A.J. Sjogren’s syndrome: A community-based study of prevalence and impact. Rheumatology 1998, 37, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Burbelo, P.D.; Ambatipudi, K.; Alevizos, I. Genome-wide association studies in Sjögren’s syndrome: What do the genes tell us about disease pathogenesis? Autoimmun. Rev. 2014, 13, 756–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haugen, A.J.; Peen, E.; Hultén, B.; Johannessen, A.C.; Brun, J.G.; Halse, A.K.; Haga, H. Estimation of the prevalence of primary Sjögren’s syndrome in two age-different community-based populations using two sets of classification criteria: The Hordaland Health Study. Scand. J. Rheumatol. 2008, 37, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Teos, L.Y.; Alevizos, I. Genetics of Sjögren’s syndrome. Clin. Immunol. 2017, 182, 41–47. [Google Scholar] [CrossRef]
- Ice, J.A.; Li, H.; Adrianto, I.; Lin, P.C.; Kelly, J.; Montgomery, C.; Lessard, C.J.; Moser, K.L. Genetics of Sjögren’s syndrome in the genome-wide association era. J. Autoimmun. 2012, 39, 57–63. [Google Scholar] [CrossRef] [Green Version]
- the International Multiple Sclerosis Genetics Consortium Class II HLA interactions modulate genetic risk for multiple sclerosis. Nat. Genet. 2015, 47, 1107–1113. [CrossRef]
- Fernando, M.M.A.; Stevens, C.R.; Walsh, E.C.; De Jager, P.L.; Goyette, P.; Plenge, R.M.; Vyse, T.J.; Rioux, J.D. Defining the Role of the MHC in Autoimmunity: A Review and Pooled Analysis. PLoS Genet. 2008, 4, e1000024. [Google Scholar] [CrossRef]
- Cobb, B.L.; Lessard, C.J.; Harley, J.B.; Moser, K.L. Genes and Sjögren’s Syndrome. Rheum. Dis. Clin. N. Am. 2008, 34, 847–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fye, K.H.; Terasaki, P.I.; Daniels, T.E.; Opelz, G.; Talal, N. Relationship of Hla-Dw3 and Hla-B8 to Sjögren’S Syndrome. Arthritis Care Res. 1978, 21, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Dendrou, C.A.; Petersen, J.; Rossjohn, J.; Fugger, L. HLA variation and disease. Nat. Rev. Immunol. 2018, 18, 325–339. [Google Scholar] [CrossRef]
- Rudolph, M.G.; Stanfield, R.L.; Wilson, I.A. How Tcrs Bind Mhcs, Peptides, and Coreceptors. Annu. Rev. Immunol. 2006, 24, 419–466. [Google Scholar] [CrossRef]
- Muraro, P.; Robins, H.; Malhotra, S.; Howell, M.; Phippard, D.; Desmarais, C.; Sousa, A.D.P.A.; Griffith, L.M.; Lim, N.; Nash, R.A.; et al. T cell repertoire following autologous stem cell transplantation for multiple sclerosis. J. Clin. Investig. 2014, 124, 1168–1172. [Google Scholar] [CrossRef]
- Gebe, J.; Swanson, E.; Kwok, W.W. HLA Class II peptide-binding and autoimmunity. Tissue Antigens 2002, 59, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Rischmueller, M.; Lester, S.; Chen, Z.; Champion, G.; Berg, R.V.D.; Beer, R.; Coates, T.; Mccluskey, J.; Gordon, T. HLA class II phenotype controls diversification of the autoantibody response in primary Sjögren’s syndrome (pSS). Clin. Exp. Immunol. 1998, 111, 365–371. [Google Scholar] [CrossRef]
- Mann, D.L.; Moutsopoulos, H.M. HLA DR alloantigens in different subsets of patients with Sjogren’s syndrome and in family members. Ann. Rheum. Dis. 1983, 42, 533–536. [Google Scholar] [CrossRef]
- Bolstad, A.I.; Wassmuth, R.; Haga, H.J.; Jonsson, R. HLA markers and clinical characteristics in Caucasians with primary Sjögren’s syndrome. J. Rheumatol. 2001, 28, 1554–1562. [Google Scholar]
- Varela-Calvino, R.; Calviño-Sampedro, C.; Gomez-Touriño, I.; Cordero, O.J. Apportioning Blame: Autoreactive CD4+ and CD8+ T Cells in Type 1 Diabetes. Arch. Immunol. Ther. Exp. 2017, 65, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.; Nicholson, M.J.; Pyrdol, J.; Wucherpfennig, K.W. Unconventional topology of self peptide-major histocompatibility complex binding by a human autoimmune T cell receptor. Nat. Immunol. 2005, 6, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Chused, T.M.; Kassan, S.S.; Opelz, G.; Moutsopoulos, H.M.; Terasaki, P.I. Sjögren’s Syndrome Associated with HLA-Dw3. N. Engl. J. Med. 1977, 296, 895–897. [Google Scholar] [CrossRef] [PubMed]
- Manthorpe, R.; Morling, N.; Platz, P.; Ryder, L.P.; Svejgaard, A.; Thomsen, M. Hla-D Antigen Frequencies in Sjögren’s Syndrome: Differences between the primary and secondary form. Scand. J. Rheumatol. 1981, 10, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Reveille, J.D.; Wilson, R.W.; Provost, T.T.; Bias, W.B.; Arnett, F.C. Primary Sjögren’s Syndrome and Other Autoimmune Diseases in Families. Ann. Intern. Med. 1984, 101, 748–756. [Google Scholar] [CrossRef]
- Brito-Zerón, P.; Baldini, C.; Bootsma, H.; Bowman, S.J.; Jonsson, R.; Mariette, X.; Sivils, K.; Theander, E.; Tzioufas, A.; Ramos-Casals, M. Sjögren syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16047. [Google Scholar] [CrossRef]
- Anaya, J.-M.; Mantilla, R.D.; Correa, P.A. Immunogenetics of primary Sjögren’s syndrome in Colombians. Semin. Arthritis Rheum. 2005, 34, 735–743. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, P.; Gohlke, H.; Price, D.J.; Klebe, G.; Brooks, C.L. Assessing Scoring Functions for Protein−Ligand Interactions. J. Med. Chem. 2004, 47, 3032–3047. [Google Scholar] [CrossRef]
- Ewing, T.J.; Makino, S.; Skillman, A.G.; Kuntz, I.D. DOCK 4.0: Search strategies for automated molecular docking of flexible molecule databases. J. Comput. Aided Mol. Des. 2001, 15, 411–428. [Google Scholar] [CrossRef]
- Harley, J.B.; Reichlin, M.; Arnett, F.C.; Alexander, E.L.; Bias, W.B.; Provost, T.T. Gene Interaction at HLA-DQ Enhances Autoantibody Production in Primary Sjögren’s Syndrome. Science 1986, 232, 1145–1147. [Google Scholar] [CrossRef]
- Gottenberg, J.-E.; Busson, M.; Loiseau, P.; Cohen-Solal, J.; Lepage, V.; Charron, D.; Sibilia, J.; Mariette, X. In primary Sjögren’s syndrome, HLA class II is associated exclusively with autoantibody production and spreading of the autoimmune response. Arthritis Care Res. 2003, 48, 2240–2245. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Tapias, P.; Rojas-Villarraga, A.; Maier-Moore, S.; Anaya, J.-M. HLA and Sjögren’s syndrome susceptibility. A meta-analysis of worldwide studies. Autoimmun. Rev. 2012, 11, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Quelvennec, E.; Alizadeh, M.; Guggenbuhl, P.; Birebent, B.; Perdriger, A.; Grosbois, B.; Pawlotsky, P.Y.; Semana, G. DRB1*15 and DRB1*03 extended haplotype interaction in primary Sjögren’s syndrome genetic susceptibility. Clin. Exp. Rheumatol. 1998, 16, 725–728. [Google Scholar]
- Paisansinsup, T.; Deshmukh, U.S.; Chowdhary, V.R.; Luthra, H.S.; Fu, S.M.; David, C.S. HLA class II influences the immune response and antibody diversification to Ro60/Sjögren’s syndrome-A: Heightened antibody responses and epitope spreading in mice expressing HLA-DR molecules. J. Immunol. 2002, 168, 5876–5884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, E.A.; Moustakas, A.K.; Bui, J.; Nouv, R.; Papadopoulos, G.K.; Kwok, W.W. The Binding of Antigenic Peptides to HLA-DR Is Influenced by Interactions between Pocket 6 and Pocket 9. J. Immunol. 2009, 183, 3249–3258. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, M.E.; Terasaki, P.I.; Graw, R.; Chused, T.M. Increased Frequency of HL-A8 in Sjogren’s Syndrome. Tissue Antigens 1975, 6, 342–346. [Google Scholar] [CrossRef]
- Moutsopoulos, H.M.; Mann, D.L.; Johnson, A.H.; Chused, T.M. Genetic Differences between Primary and Secondary Sicca Syndrome. N. Engl. J. Med. 1979, 301, 761–763. [Google Scholar] [CrossRef]
- Molina, R.; Provost, T.T.; Arnett, F.C.; Bias, W.B.; Hochberg, M.C.; Wilson, R.W.; Alexander, E.L. Primary Sjögren’s syndrome in men. Am. J. Med. 1986, 80, 23–31. [Google Scholar] [CrossRef]
- Kang, H.I.; Fei, H.M.; Saito, I.; Sawada, S.; Chen, S.L.; Yi, D.; Chan, E.; Peebles, C.; Bugawan, T.L.; Erlich, H.A. Comparison of HLA class II genes in Caucasoid, Chinese, and Japanese patients with primary Sjögren’s syndrome. J. Immunol. 1993, 150, 3615–3623. [Google Scholar]
- Moriuchi, J.; Ichikawa, Y.; Takaya, M.; Shimizu, H.; Uchiyama, M.; Sato, K.; Tsuji, K.; Arimori, S. Familial Sjögren’s syndrome in the Japanese: Immunogenetic and serological studies. Clin. Exp. Rheumatol. 1986, 4, 237–241. [Google Scholar]
- Miyagawa, S.; Shinohara, K.; Nakajima, M.; Kidoguchi, K.-I.; Fujita, T.; Fukumoto, T.; Yoshioka, A.; Dohi, K.; Shirai, T. Polymorphisms of HLA class II genes and autoimmune responses to Ro/SS-A-La/SS-B among Japanese subjects. Arthritis Care Res. 1998, 41, 927–934. [Google Scholar] [CrossRef]
- Hernández-Molina, G.; Vargas-Alarcón, G.; Rodríguez-Pérez, J.M.; Martínez-Rodríguez, N.; Lima, G.; Sánchez-Guerrero, J. High-resolution HLA analysis of primary and secondary Sjögren’s syndrome: A common immunogenetic background in Mexican patients. Rheumatol. Int. 2014, 35, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J.-M.; Correa, P.; Mantilla, R.D.; Arcos-Burgos, M. TAP, HLA-DQB1, and HLA-DRB1 polymorphism in Colombian patients with primary Sjögren’s syndrome. Semin. Arthritis Rheum. 2002, 31, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Roitberg-Tambur, A.; Friedmann, A.; Safirman, C.; Markitziu, A.; Ben-Chetrit, E.; Rubinow, A.; Moutsopoulos, H.M.; Stavropoulos, E.; Skopouli, F.N.; Margalit, H.; et al. Molecular analysis of HLA class II genes in primary sjo¨gren’s syndrome: A study of Israeli Jewish and Greek Non-Jewish patients. Hum. Immunol. 1993, 36, 235–242. [Google Scholar] [CrossRef]
- Manoussakis, M.N.; Georgopoulou, C.; Zintzaras, E.; Spyropoulou, M.; Stavropoulou, A.; Skopouli, F.N.; Moutsopoulos, H.M. Sjögren’s syndrome associated with systemic lupus erythematosus: Clinical and laboratory profiles and comparison with primary Sjögren’s syndrome. Arthritis Care Res. 2004, 50, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Lope, B.; García, M.T.C.; de Ramón Garrido, E. Immunogenetics of the Sjogren’s syndrome in southern Spain. An. Med. Interna 1994, 11, 56–61. [Google Scholar]
- Vitali, C.; Tavoni, A.; Rizzo, G.; Neri, R.; D’Ascanio, A.; Cristofani, R.; Bombardieri, S. HLA antigens in Italian patients with primary Sjogren’s syndrome. Ann. Rheum. Dis. 1986, 45, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Morling, N.; Andersen, V.; Fugger, L.; Georgsen, J.; Halberg, P.; Oxholm, P.; Odum, N.; Svejgaard, A. Immunogenetics of rheumatoid arthritis and primary Sjögren’s syndrome: DNA polymorphism of HLA class II genes. Dis. Markers 1991, 9, 289–296. [Google Scholar]
- Kerttula, T.O.; Collin, P.; Polvi, A.; Korpela, M.; Partanen, J.; Mäki, M. Distinct immunologic features of finnish Sjögren’s syndrome patients with HLA alleles DRB1*0301, DQA1*0501, and DQB1*0201. Alterations in circulating T cell receptor γ/δ subsets. Arthritis Care Res. 1996, 39, 1733–1739. [Google Scholar] [CrossRef]
- Nakken, B.; Jonsson, R.; Brokstad, K.A.; Omholt, K.; Nerland, A.H.; Haga, H.J.; Halse, A.-K. Associations of MHC Class II Alleles in Norwegian Primary Sjögren’s Syndrome Patients: Implications for Development of Autoantibodies to the Ro52 Autoantigen. Scand. J. Immunol. 2001, 54, 428–433. [Google Scholar] [CrossRef]
- Pease, C.T.; Shattles, W.; Charles, P.J.; Venables, P.J.; Maini, R.N. Clinical, serological, and HLA phenotype subsets in Sjögren’s syndrome. Clin. Exp. Rheumatol. 1989, 7, 185–190. [Google Scholar] [PubMed]
- Kacem, H.H.; Kaddour, N.; Adyel, F.; Bahloul, Z.; Ayadi, H. HLA-DQB1 CAR1/CAR2, TNFa IR2/IR4 and CTLA-4 polymorphisms in Tunisian patients with rheumatoid arthritis and Sjogren’s syndrome. Rheumatology 2001, 40, 1370–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scofield, R.; Frank, M.B.; Neas, B.R.; Horowitz, R.M.; Hardgrave, K.L.; Fujisaku, A.; McArthur, R.; Harley, J.B. Cooperative Association of T Cell β Receptor and HLA-DQ Alleles in the Production of Anti-Ro in Systemic Lupus Erythematosus. Clin. Immunol. Immunopathol. 1994, 72, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-Y.; Huang, W.; She, J.-X.; Baxter, F.; Volpe, R.; MacLaren, N.K. HLA-DRB1108, DRB1103/DRB310101, and DRB310202 Are Susceptibility Genes for Graves’ Disease in North American Caucasians, Whereas DRB1107 Is Protective1. J. Clin. Endocrinol. Metab. 1999, 84, 3182–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitlin, A.A.; Heward, J.M.; Newby, P.R.; Carr-Smith, J.D.; Franklyn, J.A.; Gough, S.C.L.; Simmonds, M. Analysis of HLA class II genes in Hashimoto’s thyroiditis reveals differences compared to Graves’ disease. Genes Immun. 2008, 9, 358–363. [Google Scholar] [CrossRef]
- Castro-Santos, P.; Olloquequi, J.; Verdugo, R.A.; Gutiérrez, M.A.; Pinochet, C.; Quiñones, L.A.; Díaz-Peña, R. HLA-DRB1*07:01 and *08:02 Alleles Confer a Protective Effect Against ACPA-Positive Rheumatoid Arthritis in a Latin American Admixed Population. Biology 2020, 9, 467. [Google Scholar] [CrossRef]
- Jacobi, T.; Massier, L.; Klöting, N.; Horn, K.; Schuch, A.; Ahnert, P.; Engel, C.; Löffler, M.; Burkhardt, R.; Thiery, J.; et al. HLA Class II Allele Analyses Implicate Common Genetic Components in Type 1 and Non–Insulin-Treated Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2020, 105, e245–e254. [Google Scholar] [CrossRef]
- de Holanda, M.I.; Klumb, E.; Imada, A.; Lima, L.A.; Alcântara, I.; Gregório, F.; Christiani, L.F.; Martins, C.O.; Timoner, B.E.; Motta, J.; et al. The prevalence of HLA alleles in a lupus nephritis population. Transpl. Immunol. 2018, 47, 37–43. [Google Scholar] [CrossRef]
- Dawson, L.; Tobin, A.; Smith, P.; Gordon, T. Antimuscarinic antibodies in Sjögren’s syndrome: Where are we, and where are we going? Arthritis Care Res. 2005, 52, 2984–2995. [Google Scholar] [CrossRef]
- Yang, L.; Ju, J.; Zhang, W.; Lv, F.; Pang, C.; Yang, G.; Wang, Y. Effects of Muscarinic Acetylcholine 3 Receptor208-227Peptide Immunization on Autoimmune Response in Nonobese Diabetic Mice. Clin. Dev. Immunol. 2013, 2013, 485213. [Google Scholar] [CrossRef]
- Kurien, B.T.; Dsouza, A.; Igoe, A.; Lee, Y.J.; Maier-Moore, J.S.; Gordon, T.; Jackson, M.; Scofield, R.H. Immunization with 60 kD Ro peptide produces different stages of preclinical autoimmunity in a Sjögren’s syndrome model among multiple strains of inbred mice. Clin. Exp. Immunol. 2013, 173, 67–75. [Google Scholar] [CrossRef]
- Espinosa, A.; Zhou, W.; Ek, M.; Hedlund, M.; Brauner, S.; Popovic, K.; Horvath, L.; Wallerskog, T.; Oukka, M.; Nyberg, F.; et al. The Sjögren’s Syndrome-Associated Autoantigen Ro52 Is an E3 Ligase That Regulates Proliferation and Cell Death. J. Immunol. 2006, 176, 6277–6285. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, U.S.; Lewis, J.E.; Gaskin, F.; Kannapell, C.C.; Waters, S.T.; Lou, Y.-H.; Tung, K.S.K.; Fu, S.M. Immune Responses to Ro60 and Its Peptides in Mice. I. The Nature of the Immunogen and Endogenous Autoantigen Determine the Specificities of the Induced Autoantibodies. J. Exp. Med. 1999, 189, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Szczerba, B.M.; Kaplonek, P.; Wolska, N.; Podsiadlowska, A.; Rybakowska, P.; Dey, P.; Rasmussen, A.; Grundahl, K.; Hefner, K.S.; Stone, D.U.; et al. Interaction between innate immunity and Ro52-induced antibody causes Sjögren’s syndrome-like disorder in mice. Ann. Rheum. Dis. 2015, 75, 617–622. [Google Scholar] [CrossRef]
- Scofield, R.H.; Asfa, S.; Obeso, D.; Jonsson, R.; Kurien, B.T. Immunization with Short Peptides from the 60-kDa Ro Antigen Recapitulates the Serological and Pathological Findings as well as the Salivary Gland Dysfunction of Sjögren’s Syndrome. J. Immunol. 2005, 175, 8409–8414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scofield, R.H.; Pierce, P.G.; James, J.A.; Kaufman, K.M.; Kurien, B.T. Immunization with Peptides from 60 kDa Ro in Diverse Mouse Strains. Scand. J. Immunol. 2002, 56, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Scofield, R.H.; Kaufman, K.M.; Baber, U.; James, J.A.; Harley, J.B.; Kurien, B.T. Immunization of mice with human 60-kd Ro peptides results in epitope spreading if the peptides are highly homologous between human and mouse. Arthritis Rheum. 1999, 42, 1017–1024. [Google Scholar] [CrossRef]
- Topfer, F.; Gordon, T.; McCluskey, J. Intra- and intermolecular spreading of autoimmunity involving the nuclear self-antigens La (SS-B) and Ro (SS-A). Proc. Natl. Acad. Sci. USA 1995, 92, 875–879. [Google Scholar] [CrossRef] [Green Version]
- Igoe, A.; Scofield, R.H. Autoimmunity and infection in Sjögren’s syndrome. Curr. Opin. Rheumatol. 2013, 25, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Witas, R.; Gupta, S.; Nguyen, C.Q. Contributions of Major Cell Populations to Sjögren’s Syndrome. J. Clin. Med. 2020, 9, 3057. [Google Scholar] [CrossRef]
- Pearson, M.N.; Bailey, A.M. Viruses of Botrytis. Adv. Virus Res. 2013, 86, 249–272. [Google Scholar] [CrossRef]
- Hammond, J.; Reinsel, M.; Grinstead, S.; Lockhart, B.; Jordan, R.; Mollov, D. A Mixed Infection of Helenium Virus S with Two Distinct Isolates of Butterbur Mosaic Virus, One of Which Has a Major Deletion in an Essential Gene. Front. Microbiol. 2020, 11, 612936. [Google Scholar] [CrossRef] [PubMed]
- Gulder, T.A.M.; Neff, S.; Schüz, T.; Winkler, T.; Gees, R.; Böhlendorf, B. The myxocoumarins A and B from Stigmatella aurantiaca strain MYX-030. Beilstein J. Org. Chem. 2013, 9, 2579–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunze, B.; Reichenbach, H.; Augustiniak, H.; Höfle, G. Isolation and identification of althiomycin from Cystobacter fuscus (Myxobacterales). J. Antibiot. 1982, 35, 635–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyksma, S.; Pjevac, P.; Ovanesov, K.; Mussmann, M. Evidence for H2consumption by unculturedDesulfobacteralesin coastal sediments. Environ. Microbiol. 2017, 20, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Palleroni, N.J. The Pseudomonas Story. Environ. Microbiol. 2010, 12, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, M.J.; Myers, K.S.; Donohue, T.J.; Noguera, D.R. Medium-Chain Fatty Acid Synthesis by “Candidatus Weimeria bifida” gen. nov., sp. nov., and “Candidatus Pseudoramibacter fermentans” sp. nov. Appl. Environ. Microbiol. 2020, 86, e02242-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wan, X.; Wu, X.; Zhang, C.; Liu, J.; Hou, S. Eubacterium rectale contributes to colorectal cancer initiation via promoting colitis. Gut Pathog. 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Abdel-Wahab, N.M.; Hassan, H.M.; Abdelmohsen, U.R. Saccharopolyspora: An underexplored source for bioactive natural products. J. Appl. Microbiol. 2019, 128, 314–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creemers-Schild, D.; Gronthoud, F.; Spanjaard, L.; Visser, L.; Brouwer, C.; Kuijper, E. Fusobacterium necrophorum, an emerging pathogen of otogenic and paranasal infections? New Microbes New Infect. 2014, 2, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khelaifia, S.; Croce, O.; Lagier, J.-C.; Robert, C.; Couderc, C.; Di Pinto, F.; Davoust, B.; Djossou, F.; Raoult, D.; Fournier, P.-E. Noncontiguous finished genome sequence and description of Virgibacillus massiliensis sp. nov., a moderately halophilic bacterium isolated from human gut. New Microbes New Infect. 2015, 8, 78–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentini, F.; Evangelisti, M.; Arpinelli, M.; Di Nardo, G.; Borro, M.; Simmaco, M.; Villa, M.P. Gut microbiota composition in children with obstructive sleep apnoea syndrome: A pilot study. Sleep Med. 2020, 76, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Barandouzi, Z.A.; Starkweather, A.R.; Henderson, W.; Gyamfi, A.; Cong, X.S. Altered Composition of Gut Microbiota in Depression: A Systematic Review. Front. Psychiatry 2020, 11, 541. [Google Scholar] [CrossRef] [PubMed]
- van der Meulen, T.A.; Harmsen, H.J.; Vila, A.V.; Kurilshikov, A.; Liefers, S.C.; Zhernakova, A.; Fu, J.; Wijmenga, C.; Weersma, R.K.; de Leeuw, K.; et al. Shared gut, but distinct oral microbiota composition in primary Sjögren’s syndrome and systemic lupus erythematosus. J. Autoimmun. 2018, 97, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Szymula, A.; Rosenthal, J.; Szczerba, B.M.; Bagavant, H.; Fu, S.M.; Deshmukh, U.S. T cell epitope mimicry between Sjögren’s syndrome Antigen A (SSA)/Ro60 and oral, gut, skin and vaginal bacteria. Clin. Immunol. 2014, 152, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrêa, J.D.; Calderaro, D.C.; Ferreira, G.A.; Mendonça, S.M.S.; Fernandes, G.R.; Xiao, E.; Teixeira, A.L.; Leys, E.J.; Graves, D.T.; Silva, T.A. Subgingival microbiota dysbiosis in systemic lupus erythematosus: Association with periodontal status. Microbiome 2017, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mandl, T.; Marsal, J.; Olsson, P.; Ohlsson, B.; Andréasson, K. Severe intestinal dysbiosis is prevalent in primary Sjögren’s syndrome and is associated with systemic disease activity. Arthritis Res. Ther. 2017, 19, 237. [Google Scholar] [CrossRef]
- De Paiva, C.S.; Jones, D.B.; Stern, M.E.; Bian, F.; Moore, Q.L.; Corbiere, S.; Streckfus, C.F.; Hutchinson, D.S.; Ajami, N.J.; Petrosino, J.F.; et al. Altered Mucosal Microbiome Diversity and Disease Severity in Sjögren Syndrome. Sci. Rep. 2016, 6, 23561. [Google Scholar] [CrossRef] [PubMed]
- Tsunawaki, S.; Nakamura, S.; Ohyama, Y.; Sasaki, M.; Ikebe-Hiroki, A.; Hiraki, A.; Kadena, T.; Kawamura, E.; Kumamaru, W.; Shinohara, M.; et al. Possible function of salivary gland epithelial cells as nonprofessional antigen-presenting cells in the de-velopment of Sjögren’s syndrome. J. Rheumatol. 2002, 29, 1884–1896. [Google Scholar]
- Geluk, A.; Van Meijgaarden, K.E.; Southwood, S.; Oseroff, C.; Drijfhout, J.W.; De Vries, R.R.; Ottenhoff, T.H.; Sette, A. HLA-DR3 molecules can bind peptides carrying two alternative specific submotifs. J. Immunol. 1994, 152, 5742–5748. [Google Scholar]
- Becerra-Artiles, A.; Cruz, J.; Leszyk, J.D.; Sidney, J.; Sette, A.; Shaffer, S.A.; Stern, L.J. Naturally processed HLA-DR3-restricted HHV-6B peptides are recognized broadly with polyfunctional and cytotoxic CD4 T-cell responses. Eur. J. Immunol. 2019, 49, 1167–1185. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.; Yusuf, N.; Smith, E.L.; Whettlock, E.M.; Naran, K.; Arif, S.; Peakman, M. Proinsulin peptide promotes autoimmune diabetes in a novel HLA-DR3-DQ2-transgenic murine model of spontaneous disease. Diabetologia 2019, 62, 2252–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.W.; Osman, R.; Menconi, F.; Concepcion, E.; Tomer, Y. Cepharanthine blocks TSH receptor peptide presentation by HLA-DR3: Therapeutic implications to Graves’ disease. J. Autoimmun. 2020, 108, 102402. [Google Scholar] [CrossRef] [PubMed]
- Li, C.W.; Menconi, F.; Osman, R.; Mezei, M.; Jacobson, E.M.; Concepcion, E.; David, C.S.; Kastrinsky, D.B.; Ohlmeyer, M.; Tomer, Y. Identifying a Small Molecule Blocking Antigen Presentation in Autoimmune Thyroiditis. J. Biol. Chem. 2016, 291, 4079–4090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, A.; Endreffy, E.; Petri, I.; Kovacs, L.; Pokorny, G. HLA class II allele polymorphism in Hungarian patients with primary Sjögren’s syndrome. Scand. J. Rheumatol. 2006, 35, 75–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country of Origin/Population | HLA Alleles Connotation | Auto-Antibodies Identified | References |
|---|---|---|---|
| U.S.A./American Caucasian | HLA-B8 | ND * | [36] |
| U.S.A./American Caucasian | HLA-Dw3 | ND * | [22] |
| U.S.A./American Caucasian | HLA-Dw3-HLA-B8 | ND * | [12] |
| U.S.A./American Caucasian | HLA-DRw3-HLA-B8 | Antinuclear antibodies Ro60 | [37] |
| U.S.A./American Caucasian | HLA-DRw3-HLA-B8 | Ro52 | [18] |
| U.S.A./American Caucasian | HLA-DRw52 | SS-A | [38] |
| Japan/Japanese population | HLA-DRB1*0301 | SS-A and SS-B | [39] |
| HLA-DRB3*0101 | |||
| HLA-DQA1*0501/DQB1*0201 | |||
| Japan/Japanese population | HLA-DRB1*0405 | SS-A and SS-B | [39] |
| HLA-DRB4*0101 | |||
| HLA-DQA1*0301/DQB1*0401 | |||
| Japan/Japanese population | HLA-DRw53 | Ro/SS-A and La/SS-B | [40] |
| Japan/Japanese population | HLA-DRB1*8032/DQA1*0103/DQB1*0601 | Ro/SS-A and La/SS-B | [39,41] |
| HLA-DRB1*8032 | |||
| HLA-DRB1*0405-DRB4*0101 | |||
| HLA-DQA1*0301 | |||
| HLA-DQB1*0401 | |||
| China/Chinese population | HLA-DRB1*0803 | SS-A and SS-B | [39] |
| HLA-DQA1*0103/DQB1*0601 | |||
| Mexico/Mexican population | HLA-DRB1*01:01 | Ro/SS-A and La/SS-B | [42] |
| HLA-B*35:01 | |||
| Colombia/Mestizo Colombian population | HLA-DRB1*0301 | Ro/SS-A and La/SS-B | [26,32,43] |
| HLA-DQB1*0201 | |||
| Israel/Israeli Jewish/Greek | HLA-DQA1*001 | SS-A, and SS-B | [44] |
| HLA-DQA1*0201/DQB1*0501-Jewish | |||
| HLA-DQA1*0501-Greek | |||
| Greece/Greek population | HLA-DRB1*0301 | Ro/SSA and anti-La/SSB | [45] |
| Spain/Spanish population | HLA-Cw7 | Ro/SSA and anti-La/SSB | [46] |
| HLA- DRB1*0301 | |||
| HLA-DR11 | |||
| France/French population | HLA-DRB1*1501 | ND * | [33] |
| HLA- DRB1*0301 | |||
| HLA-DQB1*0201 | |||
| HLA-DQB1*0602 | |||
| France/French population | HLA-DRB1*0301 | anti-SSA and/or anti-SSB | [31] |
| HLA-DQB1*02 | |||
| Italy/Italian population | HLA-DRB1*0301 | anti-Ro/SSA | [47] |
| Denmark/Danish population | HLA-Dw2 | ND * | [23] |
| Denmark/Danish population | HLA-DQA1*0501 | anti-SSA and/or anti-SSB | [48] |
| HLA-DQB1*0201 | |||
| HLA-DQA1*0301 | |||
| Finland/Finnish population | HLA-DRB1*0301 | anti-SS-A/Ro and anti-SS-B/La | [49] |
| HLA-DQA1*0501 | |||
| HLA-DQB1*0201 | |||
| Norway/Norwegian Caucasian population | HLA-DRB1*0301 | Ro/SSA and La/SSB | [50] |
| Norway/Norwegian Caucasian population | HLA-DRB1*0301 | anti-La/SSB strong positive association with DQA1*0501 anti-Ro/SSA and anti-La/SSB autoantibody response was positively associated with DRB1*03, DQB1*02 and DRB1*03/DRB1*15-DQB1*02/DQB1*0602 | [19] |
| HLA-DQB1*02 | |||
| HLA-DQA1*0501 | |||
| United Kingdom/British Caucasian population | HLA-DRB1*0301 | Ro/SSA and La/SSB | [51] |
| HLA-DRw52 | |||
| Australia/Australian population | HLA-DRB1*0301 | Ro/SSA and La/SSB | [16] |
| HLA-DQA1*0501 | |||
| HLA-DQB1*02 | |||
| Tunisian population | HLA-DQB1 CAR1/CAR2 | ND * | [52] |
| European and African American population | HLA-DQB1*0201 | SSA | [53] |
| HLA-DQA1*0101 |
| Disease | Protective HLA Class II Allele | References |
|---|---|---|
| Graves’ disease | HLA-DRB1*07 | [54] |
| HLA-DQB1*02 | ||
| HLA-DQA1*02 | ||
| Hashimoto’s thyroiditis | HLA-DRB1*07 | [55] |
| HLA-DQB1*02 | ||
| HLA-DQA1*02 | ||
| Rheumatoid arthritis | HLA-DRB1*0103 | [56] |
| HLA-DRB1*07 | ||
| HLA-DRB1*1201 | ||
| HLA-DRB1*1301 | ||
| HLA-DRB1*1501 | ||
| Multiple sclerosis | DRB1*14-DQB1*06-DQA1*0102 | [9] |
| Type 1 diabetes | DRB1*14-DQB1*06-DQA1*0102 | [57] |
| DRB1*15-DQB1*06-DQA1*01 | ||
| Systemic lupus erythematosus | DR4 | [58] |
| DR5 | ||
| DR11 | ||
| DR14 |
| Peptide | Amino Acids | Amino Acid Sequence | In Vivo Confirmation | References | HLA-DR3 | IC50 |
|---|---|---|---|---|---|---|
| M3R | 205–237 | LFWQYFVGKRTVPPGECFIQFLSEPTITFGTAI | NOD/LtJ mice | [59] | GECFIQFLSEPTITF | 473 |
| 208–227 | QYFVGKRTVPPGECFIQFLS | Immunization of young female NOD/LtJ mice on autoimmune response | [60] | QYFVGKRTVPPGECF | 8607 | |
| Part of second extracellular loop | ||||||
| 213–228 | KRTVPPGECFIQFLSE | BALB/c | [61] | KRTVPPGECFIQFLS | 50,000 | |
| 514–527 | NTFCDSCIPKTFWN | BALB/c | [61] | NTFCDSCIPKTFWNL | 6549 | |
| MTLHSNSTTSPLFPNISSSWVHSPSEAGLP, N1 | C57BL/6j (B6) mice (M3R+/+) M3R−/− mice Rag1−/− mice | [62] | PNISSSWVHSPSEAG | 4760 | ||
| VHSPSEAGLPLGTVSQLDSYNISGTSGNFS, N2 | LPLGTVSQLDSYNIS | 6028 | ||||
| NISQTSGNFSSNDTSSDPLGGHTIWQV, N3 | TSGNFSSNDTSSDPL | 6471 | ||||
| FTTYIIMNRWALGNLACDLW, Extracellular loop 1 | FTTYIIMNRWALGNL | 955 | ||||
| QYFVGKRTVPPGECFIQFLSEP, Extracellular loop 2 | QYFVGKRTVPPGECF | 8607 | ||||
| VLVNTFCDSCIPKTYWNLGY, Extracellular loop 3 | VLVNTFCDSCIPKTY | 5219 | ||||
| H 441–465 | PAGGTDCSLPMIWAQKTNTPADVFI | SJL/L (H-2s) A/J(H-2a) | [63] | TDCSLPMIWAQKTNT | 2068 | |
| H 316–335 | KARIHPFHILIALETYKTGH | SJL/L (H-2s) BALB/c (H-2d) A/J(H-2a) | [63] | IHPFHILIALETYKT | 1485 | |
| H 306–325 | EKLCNEKLLKKARIHPFHIL | SJL/L (H-2s) | [63] | EKLLKKARIHPFHIL | 1721 | |
| H 26–45 | QVTDMNRLHRFLCFGSEGGT | SJL/L (H-2s) | [63] | QVTDMNRLHRFLCFG | 2266 | |
| H 401–425 | MVVTREKDSYVVAFSDEMVPCPVT | SJL/L (H-2s) A/J(H-2a) | [63] | REKDSYVVAFSDEMV | 2879 | |
| H 481–505 | IALREYRKKMDIPAKLIVCGMSTNG | SJL/L (H-2s) | [63] | REYRKKMDIPAKLIV | 622 | |
| H 201–225 | YITKGWKEVHELYKEKALSVETEKL | BALB/c (H-2d) | [63] | VHELYKEKALSVETE | 2191 | |
| H 241–265 | ELEVIHLIEEHRLLTNHLKS | BALB/c (H-2d) A/J(H-2a) | [63] | VIHLIEEHRLLTNHL | 130 | |
| Ro52 | Full peptide | Full protein | New Zealand Mixed Mice (NZMZ) 2758 | [64] | NPWLILSEDRRQVRL | 28 |
| Ro60 | 480–494 | AIALREYRKKMDIPA | Animals were immunized with peptide Ro480–494 | [65,66] | AIALREYRKKMDIPA | 1876 |
| 274–290 | QEMPLTALLRNLGKMT | Animals were immunized with peptide Ro274–290 | [65,66] | EMPLTALLRNLGKMT | 1598 | |
| 274–290 | Human QEMPLTALLRNLGKMT | Amino acid sequences of the human 60-kd Ro peptides used for immunization of BALB/c mice | [67] | EMPLTALLRNLGKMT | 1598 | |
| Mouse QEMPLTALLRNLGKMT | ||||||
| 413–428 | Human VAFSDEMVPCPVTTDM | |||||
| Mouse VAFACDMVPFPVTTDM | ||||||
| Rabbit VAFSDEMVPCPLTTDM | ||||||
| 480–495 | Human AIALREYRKKMDIPA | VAFSDEMVPCPVTTD | 20,917 | |||
| Mouse AVALREYRKKMDIPA | AIALREYRKKMDIPA | 1876 | ||||
| La | 1–107 | GYVDISLLVSFNKMKKLTTDGKLIARALKSSSVVELDLEGTRIRRKKPLGERPKDEEERTVYVELLPKNVTH | [68] | MKKLTTDGKLIARAL | 136 | |
| 243–345 | KAKKRAQKDGVGQAASEVSKESRDLEFCSTEEEKETDRKGDSLSKVKRKHKKKHKERHKMGEEVIPLRVLSKTEWMDLKKEYLALQKASMASLKKTISQ | SKTEWMDLKKEYLAL | 922 | |||
| 111–242 | EQAAKAIEFLNNPPEEAPRKPGIFPKTVKNKPIPSLRVAEEKKKKKKKKGRIKKEESVQAKESAVDSSSSGVCKATKRPRTASEGSEAETPEAPKQPAKKKKKRDKVEASSLPEARAGKRERCSAEDEDCL | SSSGVCKATKRPRTA | 561 |
| Allele | Start | End | Length | Core Sequence | Peptide Sequence | IC50 | Percentile Rank | Adjusted Rank |
|---|---|---|---|---|---|---|---|---|
| HLA-DRB1*03:01 | 297 | 311 | 15 | LSEDRRQVR | NPWLILSEDRRQVRL | 28.00 | 0.10 | 0.10 |
| HLA-DRB1*03:01 | 296 | 310 | 15 | LILSEDRRQ | ANPWLILSEDRRQVR | 29.00 | 0.11 | 0.11 |
| HLA-DRB1*03:01 | 298 | 312 | 15 | LSEDRRQVR | PWLILSEDRRQVRLG | 29.00 | 0.11 | 0.11 |
| HLA-DRB1*03:01 | 299 | 313 | 15 | LSEDRRQVR | WLILSEDRRQVRLGD | 29.00 | 0.11 | 0.11 |
| HLA-DRB1*03:01 | 300 | 314 | 15 | LSEDRRQVR | LILSEDRRQVRLGDT | 29.00 | 0.11 | 0.11 |
| HLA-DRB1*03:01 | 301 | 315 | 15 | LSEDRRQVR | ILSEDRRQVRLGDTQ | 90.00 | 0.91 | 0.91 |
| HLA-DRB1*03:01 | 302 | 316 | 15 | LSEDRRQVR | LSEDRRQVRLGDTQQ | 93.00 | 0.95 | 0.95 |
| HLA-DRB1*03:01 | 197 | 211 | 15 | LEKDEREQL | LQELEKDEREQLRIL | 129.00 | 1.60 | 1.60 |
| HLA-DRB1*03:01 | 196 | 210 | 15 | LEKDEREQL | QLQELEKDEREQLRI | 137.00 | 1.60 | 1.60 |
| HLA-DRB1*03:01 | 198 | 212 | 15 | LEKDEREQL | QELEKDEREQLRILG | 137.00 | 1.60 | 1.60 |
| Allele | Start | End | Length | Core Sequence | Peptide Sequence | IC50 | Percentile Rank | Adjusted Rank |
|---|---|---|---|---|---|---|---|---|
| HLA-DRB1*03:01 | 126 | 140 | 15 | FKKDLKESM | FTFIQFKKDLKESMK | 75.00 | 0.70 | 0.70 |
| HLA-DRB1*03:01 | 127 | 141 | 15 | FKKDLKESM | TFIQFKKDLKESMKC | 75.00 | 0.70 | 0.70 |
| HLA-DRB1*03:01 | 125 | 139 | 15 | LFTFIQFKK | LFTFIQFKKDLKESM | 76.00 | 0.74 | 0.74 |
| HLA-DRB1*03:01 | 244 | 258 | 15 | LIEEHRLVR | VIHLIEEHRLVREHL | 78.00 | 0.76 | 0.76 |
| HLA-DRB1*03:01 | 128 | 142 | 15 | FKKDLKESM | FIQFKKDLKESMKCG | 79.00 | 0.77 | 0.77 |
| HLA-DRB1*03:01 | 245 | 259 | 15 | LIEEHRLVR | IHLIEEHRLVREHLL | 80.00 | 0.77 | 0.77 |
| HLA-DRB1*03:01 | 129 | 143 | 15 | FKKDLKESM | IQFKKDLKESMKCGM | 81.00 | 0.79 | 0.79 |
| HLA-DRB1*03:01 | 242 | 256 | 15 | LIEEHRLVR | LEVIHLIEEHRLVRE | 82.00 | 0.82 | 0.82 |
| HLA-DRB1*03:01 | 243 | 257 | 15 | LIEEHRLVR | EVIHLIEEHRLVREH | 82.00 | 0.82 | 0.82 |
| HLA-DRB1*03:01 | 241 | 255 | 15 | ELEVIHLIE | ELEVIHLIEEHRLVR | 83.00 | 0.83 | 0.83 |
| Allele | Start | End | Length | Core Sequence | Peptide Sequence | IC50 | Percentile Rank | Adjusted Rank |
|---|---|---|---|---|---|---|---|---|
| HLA-DRB1*03:01 | 328 | 15 | 15 | IIEDQQESL | ALKKIIEDQQESLNK | 49.00 | 0.36 | 0.36 |
| HLA-DRB1*03:01 | 327 | 15 | 15 | IIEDQQESL | EALKKIIEDQQESLN | 50.00 | 0.37 | 0.37 |
| HLA-DRB1*03:01 | 326 | 15 | 15 | KEALKKIIE | KEALKKIIEDQQESL | 51.00 | 0.38 | 0.38 |
| HLA-DRB1*03:01 | 329 | 15 | 15 | IIEDQQESL | LKKIIEDQQESLNKW | 51.00 | 0.38 | 0.38 |
| HLA-DRB1*03:01 | 330 | 15 | 15 | IIEDQQESL | KKIIEDQQESLNKWK | 54.00 | 0.40 | 0.40 |
| HLA-DRB1*03:01 | 91 | 15 | 15 | ISEDKTKIR | AELMEISEDKTKIRR | 131.00 | 1.60 | 1.60 |
| Allele | Start | End | Length | Core Sequence | Peptide Sequence | IC50 | Percentile Rank | Adjusted Rank |
|---|---|---|---|---|---|---|---|---|
| HLA-DRB1*03:01 | 192 | 206 | 15 | AWVISFVLW | AWVISFVLWAPAILF | 120.00 | 1.40 | 1.40 |
| HLA-DRB1*03:01 | 195 | 209 | 15 | VLWAPAILF | ISFVLWAPAILFWQY | 121.00 | 1.40 | 1.40 |
| HLA-DRB1*03:01 | 193 | 207 | 15 | VLWAPAILF | WVISFVLWAPAILFW | 123.00 | 1.50 | 1.50 |
| HLA-DRB1*03:01 | 194 | 208 | 15 | VLWAPAILF | VISFVLWAPAILFWQ | 123.00 | 1.50 | 1.50 |
| HLA-DRB1*03:01 | 196 | 210 | 15 | VLWAPAILF | SFVLWAPAILFWQYF | 125.00 | 1.50 | 1.50 |
| HLA-DRB1*03:01 | 375 | 389 | 15 | ILNSTKLPS | STILNSTKLPSSDNL | 169.00 | 2.30 | 2.30 |
| HLA-DRB1*03:01 | 374 | 388 | 15 | ILNSTKLPS | HSTILNSTKLPSSDN | 170.00 | 2.30 | 2.30 |
| HLA-DRB1*03:01 | 371 | 385 | 15 | LPGHSTILN | LPGHSTILNSTKLPS | 171.00 | 2.30 | 2.30 |
| HLA-DRB1*03:01 | 372 | 386 | 15 | ILNSTKLPS | PGHSTILNSTKLPSS | 171.00 | 2.30 | 2.30 |
| HLA-DRB1*03:01 | 373 | 387 | 15 | ILNSTKLPS | GHSTILNSTKLPSSD | 171.00 | 2.30 | 2.30 |
| HLA-DRB1*03:01 | 548 | 562 | 15 | FRTTFKMLL | NKTFRTTFKMLLLCQ | 198.00 | 2.70 | 2.70 |
| HLA-DRB1*03:01 | 546 | 560 | 15 | FRTTFKMLL | LCNKTFRTTFKMLLL | 199.00 | 2.70 | 2.70 |
| HLA-DRB1*03:01 | 549 | 563 | 15 | FRTTFKMLL | KTFRTTFKMLLLCQC | 199.00 | 2.70 | 2.70 |
| HLA-DRB1*03:01 | 547 | 561 | 15 | FRTTFKMLL | CNKTFRTTFKMLLLC | 200.00 | 2.70 | 2.70 |
| Allele | Start | End | Length | Core Sequence | Peptide Sequence | IC50 | Percentile Rank | Adjusted Rank |
|---|---|---|---|---|---|---|---|---|
| HLA-DRB1*03:01 | 1318 | 1332 | 15 | SHDLQRFLS | SHDLQRFLSDFRDLM | 12.00 | 0.01 | 0.01 |
| HLA-DRB1*03:01 | 1319 | 1333 | 15 | FLSDFRDLM | HDLQRFLSDFRDLMS | 12.00 | 0.01 | 0.01 |
| HLA-DRB1*03:01 | 1320 | 1334 | 15 | FLSDFRDLM | DLQRFLSDFRDLMSW | 12.00 | 0.01 | 0.01 |
| HLA-DRB1*03:01 | 1322 | 1336 | 15 | FLSDFRDLM | QRFLSDFRDLMSWIN | 12.00 | 0.01 | 0.01 |
| HLA-DRB1*03:01 | 363 | 377 | 15 | FLADFRDLT | LQRFLADFRDLTSWV | 26.00 | 0.06 | 0.06 |
| HLA-DRB1*03:01 | 360 | 374 | 15 | SYRLQRFLA | SYRLQRFLADFRDLT | 27.00 | 0.07 | 0.07 |
| HLA-DRB1*03:01 | 361 | 375 | 15 | FLADFRDLT | YRLQRFLADFRDLTS | 27.00 | 0.07 | 0.07 |
| HLA-DRB1*03:01 | 362 | 376 | 15 | FLADFRDLT | RLQRFLADFRDLTSW | 27.00 | 0.07 | 0.07 |
| HLA-DRB1*03:01 | 364 | 378 | 15 | FLADFRDLT | QRFLADFRDLTSWVT | 28.00 | 0.10 | 0.10 |
| HLA-DRB1*03:01 | 1323 | 1337 | 15 | FLSDFRDLM | RFLSDFRDLMSWING | 36.00 | 0.16 | 0.16 |
| HLA-DRB1*03:01 | 1324 | 1338 | 15 | FLSDFRDLM | FLSDFRDLMSWINGI | 37.00 | 0.17 | 0.17 |
| HLA-DRB1*03:01 | 365 | 379 | 15 | FLADFRDLT | RFLADFRDLTSWVTE | 83.00 | 0.83 | 0.83 |
| HLA-DRB1*03:01 | 366 | 380 | 15 | FLADFRDLT | FLADFRDLTSWVTEM | 85.00 | 0.83 | 0.83 |
| Allele | Core Sequence | Peptide Sequence | IC50 |
|---|---|---|---|
| HLA-DRB1*01:01 | LKNLRPNRQ | RFLLKNLRPNRQLAN | 44.00 |
| RFLLKNLRP | CRQRFLLKNLRPNRQ | 52.00 | |
| HLA-DRB1*15:01 | TGPLRPFFS | CAFTGPLRPFFSPGF | 122.00 |
| LRPFFSPGF | AFTGPLRPFFSPGFN | 123.00 | |
| HLA -DRB1*04:05 | EAGMVSFYN | LDYEAGMVSFYNITD | 39.00 |
| MVSFYNITD | DYEAGMVSFYNITDH | 39.00 | |
| HLA-DRB4*01:01 | LKNLRPNRQ | RFLLKNLRPNRQLAN | 102.00 |
| RFLLKNLRP | CRQRFLLKNLRPNRQ | 110.00 | |
| HLA-DRB3*01:01 | KRADWKEVI | IAIKRADWKEVIIVL | 229.00 |
| EVEIAIKRA | EVEIAIKRADWKEVI | 247.00 |
| Allele | Core Sequence | Peptide Sequence | IC50 |
|---|---|---|---|
| HLA-DRB1*01:01 | LFTFIQFKK | LFTFIQFKKDLKESM | 286.00 |
| FKKDLKESM | FTFIQFKKDLKESMK | 289.00 | |
| HLA-DRB1*15:01 | IQEIKSFSQ | CEVIQEIKSFSQEGR | 238.00 |
| VIQEIKSFS | GRGCEVIQEIKSFSQ | 251.00 | |
| HLA-DRB1*04:05 | LRLSHLKPS | HKDLLRLSHLKPSSE | 75.00 |
| LSHLKPSSE | DLLRLSHLKPSSEGK | 75.00 | |
| HLA-DRB4*01:01 | TYYIKEQKL | EGGTYYIKEQKLGLE | 228.00 |
| KDLLRLSHL | SHKDLLRLSHLKPSS | 237.00 | |
| HLA-DRB3*01:01 | LFTFIQFKK | LFTFIQFKKDLKESM | 286.00 |
| FKKDLKESM | FTFIQFKKDLKESMK | 289.00 |
| Allele | Core Sequence | Peptide Sequence | IC50 |
|---|---|---|---|
| HLA-DRB1*01:01 | FNVIVEALS | TDFNVIVEALSKSKA | 52.00 |
| DFNVIVEAL | NRLTTDFNVIVEALS | 60.00 | |
| HLA- DRB1*15:01 | LHILFSNHG | REDLHILFSNHGEIK | 34.00 |
| DLHILFSNH | QTCREDLHILFSNHG | 37.00 | |
| HLA-DRB1*04:05 | FNVIVEALS | LTTDFNVIVEALSKS | 66.00 |
| NRLTTDFNV | NRLTTDFNVIVEALS | 67.00 | |
| HLA-DRB4*01:01 | EIMIKFNRL | VPLEIMIKFNRLNRL | 75.00 |
| IKFNRLNRL | PLEIMIKFNRLNRLT | 77.00 | |
| HLA-DRB3*01:01 | DLDDQTCRE | DLDDQTCREDLHILF | 142.00 |
| CREDLHILF | LDDQTCREDLHILFS | 143.00 |
| Allele | Core Sequence | Peptide Sequence | IC50 |
|---|---|---|---|
| HLA-DRB1*01:01 | IAFLTGILA | VVFIAFLTGILALVT | 9.00 |
| LTGILALVT | FIAFLTGILALVTII | 12.00 | |
| HLA- DRB1*15:01 | IIGNILVIV | VTIIGNILVIVSFKV | 14.00 |
| ILVIVSFKV | IIGNILVIVSFKVNK | 14.00 | |
| HLA-DRB1*04:05 | VPPGECFIQ | VPPGECFIQFLSEPT | 7.00 |
| FIQFLSEPT | PPGECFIQFLSEPTI | 7.00 | |
| HLA-DRB4*01:01 | LVTIIGNIL | GILALVTIIGNILVI | 88.00 |
| IGVISMNLF | ADLIIGVISMNLFTT | 98.00 | |
| HLA-DRB3*01:01 | GECFIQFLS | GECFIQFLSEPTITF | 124.00 |
| FLSEPTITF | ECFIQFLSEPTITFG | 127.00 |
| Allele | Core Sequence | Peptide Sequence | IC50 |
|---|---|---|---|
| HLA-DRB1*01:01 | FQKIKSMAA | NGRFQKIKSMAASRR | 3.00 |
| IKLLQAQKL | MREKGIKLLQAQKLV | 5.00 | |
| HLA- DRB1*15:01 | WRRLKAQMI | LDRWRRLKAQMIEKR | 68.00 |
| EVLDRWRRL | NEVLDRWRRLKAQMI | 71.00 | |
| HLA-DRB1*04:05 | FRSSLSSAQ | HDAFRSSLSSAQADF | 38.00 |
| HDAFRSSLS | REAHDAFRSSLSSAQ | 39.00 | |
| HLA-DRB4*01:01 | KMREKGIKL | KMREKGIKLLQAQKL | 5.00 |
| IKLLQAQKL | MREKGIKLLQAQKLV | 5.00 | |
| HLA-DRB3*01:01 | IQETRTYLL | IQETRTYLLDGSCMV | 25.00 |
| YLLDGSCMV | QETRTYLLDGSCMVE | 25.00 |
| Protein | Predicted Peptide | Virus | Protein | Homology with Sequence (Percentage) |
|---|---|---|---|---|
| Human Ro52 | LEKDEREQL | Miniopterus schreibersii polyomavirus 1 | Large T antigen | 88.9% |
| Micromonas pusilla virus PL1 | Uncharacterized | 77.8% | ||
| Miniopterus schreibersii polyomavirus 1 | Small T antigen | 88.9% | ||
| Mouse Ro52 | MEMDLTMQR | Wiseana iridescent virus (WIV) (Insect iridescent virus type 9) | 70% | |
| Mouse Ro52 | KELAEKMEM | Mimivirus LCMiAC02 | Uncharacterized | 77.8% |
| Mouse Ro60 | LFTFIQFKK | Botrytis virus X (isolate Botrytis cinerea/New Zealand/Howitt/2006) (BOTV-X) | RNA replication | 100% |
| Human Ro60 | LFTFIQFKK | Botrytis virus X (isolate Botrytis cinerea/New Zealand/Howitt/2006) (BOTV-X)B19:B22 | RNA replication protein | 100% |
| Human M3R | AWVISFVLW | Pseudomonas phage PaMx74 | Putative membrane protein | 75% |
| Human M3R | LPGHSTILN | Pepper mild mottle virus (strain Spain) (PMMV-S) | Replicase large subunit | 88.9% |
| Odontoglossum ringspot virus (isolate Korean Cy) (ORSV-Cy) | Replicase large | 88.9% | ||
| Tobacco mild green mosaic virus (TMGMV) (TMV strain U2) | Replicase large subunit | 88.9% | ||
| Turnip vein-clearing virus (TVCV) | Replicase large subunit | 88.9% | ||
| Youcai mosaic virus (YoMV) | Replicase large subunit | 88.9% | ||
| Hoya necrotic spot virus | Methyltransferase/RNA helicase | 88.9% | ||
| Odontoglossum ringspot virus | Methyltransferase/RNA helicase | 88.9% | ||
| Virgaviridae sp. | Replication-associated protein | 88.9% | ||
| Tobacco mild green mosaic virus (TMGMV) (TMV strain U2) | Replicase large subunit | 88.9% | ||
| Brugmansia mild mottle virus | Methyltransferase/RNA helicase | 88.9% | ||
| Streptocarpus flower break virus | Methyltransferase/RNA helicase | 88.9% | ||
| Ribgrass mosaic virus (RMV) | Methyltransferase/RNA helicase | 88.9% | ||
| Wasabi mottle virus | Methyltransferase/RNA helicase | 88.9% | ||
| Piper chlorosis virus | Replicase large subunit | 88.9% | ||
| Human La | KEALKKIIE | Helenium virus S (HelVS) | Helicase | 88.9% |
| Arthrobacter phage Boersma | DNA polymerase I | 100% | ||
| Human/Mouse α-fodrin | SYRLQRFLA | Uncultured Caudovirales phage | Uncharacterized protein | 88.9% |
| Human Ro52 | LSEDRRQVR | Stigmatella aurantiaca (strain DW4/3-1) | Peptidase, M20 family | 100% |
| Cystobacter fuscus DSM 2262 | Acetylornithine deacetylase | 100% | ||
| Stigmatella aurantiaca (strain DW4/3-1) | Peptidase, M20/M25/M40 family | 100% | ||
| Human Ro52 | LEKDEREQL | Geobacter sp. (strain M21) | Endopeptidase La | 100% |
| Seonamhaeicola marinus | RNA polymerase sigma factor | 100% | ||
| Mouse Ro52 | MEMDLTMQR | Sulfuriferula nivalis | Phytoene synthase | 88.9% |
| Corallococcus exercitus | Phytoene/squalene synthase | 88.9% | ||
| Corallococcus aberystwythensis | Phytoene/squalene synthase | 88.9% | ||
| Corallococcus sp. CA047B | Phytoene/squalene synthase | 88.9% | ||
| Corallococcus exercitus | Phytoene/squalene synthase | 88.9% | ||
| Mouse Ro52 | KELAEKMEM | Arenicella xantha | RNA pol sigma factor | 100% |
| Gamma proteobacterium SS-5 | RNA pol sigma factor | 100% | ||
| Granulosicoccus antarcticus | RNA pol sigma factor | 100% | ||
| Gammaproteobacteria bacterium | RNA pol sigma factor | 100% | ||
| Granulosicoccus sp. | RNA pol sigma factor | 100% | ||
| Candidatus Methyloumidiphilum | RNA pol sigma factor | 100% | ||
| Gammaproteobacteria bacterium | Fumarate flavoprotein | 100% | ||
| Tindallia magadiensis | RNA pol sigma factor | 100% | ||
| Oceanospirillales bacterium | RNA pol sigma factor | 100% | ||
| Cyanobacterium sp. IPPAS | RNA pol sigma factor | 100% | ||
| Cyanobacterium sp. HL-69 | RNA pol sigma factor | 100% | ||
| Culicoidibacter larvae | RNA pol sigma factor | 100% | ||
| Chromobacterium violaceum | RNA pol sigma factor | 100% | ||
| Cyanobacterium stanieri | RNA pol sigma factor | 100% | ||
| Clostridium cellulovorans | RNA pol sigma factor | 100% | ||
| Anaerolineaceae bacterium | RNA pol sigma factor | 100% | ||
| Pseudobythopirellula maris | RNA pol sigma factor | 100% | ||
| Bacteroidetes bacterium | RNA pol sigma factor | 100% | ||
| Epulopiscium sp. | RNA pol sigma factor | 100% | ||
| Betaproteobacteria bacterium | RNA pol sigma factor | 100% | ||
| Fulvivirga imtechensis AK7 | 100% | |||
| Human M3R | AWVISFVLW | Planctomycetes bacterium | Uncharacterized protein | 88.9% |
| Mouse M3R | VLWAPAILF | Desulfobacterales bacterium | Site-2 protease family protein | 88.9% |
| Human La | KEALKKIIE | Candidatus Dojkabacteria bacterium | Uncharacterized protein | 100% |
| Hydrogenimonas sp. | Anthranilate phosphoribosyltransferase | 100% | ||
| candidate division WOR-3 bacterium | Uncharacterized protein | 100% | ||
| Mouse La | QRYWQKILV | Planctomycetes bacterium SM23_25 | Uncharacterized protein | 88.9% |
| Mouse La | ILVDRQAKL | Pseudomonas sp. NFR16 | Uncharacterized | 100.0% |
| Pseudomonas sp. Bc-h | Uncharacterized | 100.0% | ||
| Pseudomonas sp. GV021 | Uncharacterized | 100.0% | ||
| Pseudomonas abietaniphila | Uncharacterized | 100.0% | ||
| Pseudomonas graminis | DUF2914 family | 100.0% | ||
| Pseudomonas graminis | Uncharacterized | 100.0% | ||
| Pseudomonas graminis | Uncharacterized | 100.0% | ||
| Pseudomonas graminis | DUF2914 domain | 100.0% | ||
| Pseudomonas graminis | Uncharacterized | 100.0% | ||
| Pseudomonas sp. | DUF2914 domain | 100.0% | ||
| Pseudomonas sp. NFACC02 | Uncharacterized | 100.0% | ||
| Pseudomonas sp. LP_7_YM | DUF2914 domain | 100.0% | ||
| Pseudomonas sp. M47T1 | Uncharacterized | 100.0% | ||
| Pseudomonas eucalypticola | DUF2914 domain | 100.0% | ||
| Pseudomonas sp. K1S02-6 | DUF2914 domain | 100.0% | ||
| Human/Mouse α-fodrin | FLSDFRDLM | Cocleimonas flava | Uncharacterized | 88.9% |
| Verrucomicrobiales bacterium | Uncharacterized | 88.9% | ||
| Planctomycetaceae bacterium | SH3 domain | 88.9% |
| Allele | SjS Protein | Core Sequence | Virus | Viral Protein | Homology |
|---|---|---|---|---|---|
| HLA-DRB4*01:01 | Ro52 | RFLLKNLRP | Human Immunodeficiency Virus | Glycoprotein 120 | 100.0% |
| Serratia phage 2050H2 | Uncharacterized | 87.5% | |||
| Klebsiella phage 31 | Endopeptidase Rz | 87.5% | |||
| Escherichia phage ECA2 | Endopeptidase | 87.5% | |||
| Leclercia phage 10164RH | Uncharacterized | 87.5% | |||
| Citrobacter phage SH1 | Endopeptidase | 87.5% | |||
| Citrobacter phage phiCFP-1 | Uncharacterized | 87.5% | |||
| Serratia phage SALSA | Endopeptidase | 87.5% | |||
| Citrobacter phage SH2 | Endopeptidase Rz | 87.5% | |||
| Klebsiella phage KPP-5 | Endopeptidase | 87.5% | |||
| Leclercia phage 10164-302 | Uncharacterized | 87.5% | |||
| Enterobacter phage E-2 | Endopeptidase | 87.5% | |||
| Klebsiella phage NL_ZS_3 | Endopeptidase Rz | 87.5% | |||
| Serratia phage SM9-3Y | I-spanin | 87.5% | |||
| Escherichia phage LL2 | I-spanin | 87.5% | |||
| Salmonella phage phiSG-JL2 | Gp18.5 | 87.5% | |||
| Yersinia phage phiYeO3-12 | Endopeptidase | 87.5% | |||
| Enterobacter phage E-4 | Endopeptidase Rz | 87.5% | |||
| Enterobacter phage E-3 | Endopeptidase | 87.5% | |||
| Yersinia phage phiYe-F10 | Uncharacterized | 87.5% | |||
| Klebsiella phage | endopeptidase | 87.5% | |||
| HLA-DRB1*01:01 | Ro60 | LFTFIQFKK | Botrytis virus X | RNA replication protein | |
| 100% | Ro60 | TYYIKEQKL | Streptomyces phage | Ro-like RNA binding protein | 88.9% |
| Streptomyces phage | Ro-like RNA binding protein | 88.9% | |||
| Streptomyces phage Beuffert | Ro-like RNA binding protein | 88.9% | |||
| Pyramimonas orientalis virus | Uncharacterized protein | 69.2% | |||
| KDLLRLSHL | Botrytis virus X | RNA replication | 100% | ||
| HLA-DRB1*01:01 | La | DFNVIVEAL | Salmonella phage SPFM12 | Uncharacterized | 88.9% |
| HLA-DRB3*01:01 | La | DLDDQTCRE | Leviviridae sp. | RNA replicase beta chain | 64.3% |
| HLA-DRB1*01:01 | M3R | IAFLTGILA | Bacillus phage 031MP004 | Uncharacterized | 100% |
| Bacillus phage 055SW001 | Uncharacterized | 100% | |||
| Bacillus phage 022DV001 | Uncharacterized | 100% | |||
| Bacillus phage 031MP002 | Uncharacterized | 100% | |||
| Bacillus phage 031MP003 | Uncharacterized | 100% | |||
| HLA- DRB1*15:01 | M3R | IIGNILVIV | Human immunodeficiency virus 1 | Protein Vpu | 100% |
| Allele | SjS Protein | Core Sequence | Bacteria | Bacterial Protein | Homology |
|---|---|---|---|---|---|
| HLA -DRB1*04:05 | Ro52 | EAGMVSFYN | Legionella moravica | Ankyrin | 88.9% |
| Legionella sp. Km535 | Ankyrin repeat domain-containing protein | 88.9% | |||
| Ro52 | MVSFYNITD | Legionella moravica | Ankyrin | 88.9% | |
| Legionella sp. Km535 | Ankyrin repeat domain-containing protein | 88.9% | |||
| HLA-DRB4*01:01 | Ro60 | TYYIKEQKL | Helicobacter sp. 11S03491-1 | Protoporphyrinogen oxidase | 100% |
| Fusobacterium | Uncharacterized | 100% | |||
| Fusobacterium | Uncharacterized | 100% | |||
| HLA-DRB3*01:01 | La | CREDLHILF | Virgibacillus massiliensis | Uncharacterized | 100% |
| HLA-DRB4*01:01 | M3R | LVTIIGNIL | Uncultured | Uncharacterized | 88.9% |
| HLA-DRB1*01:01 | Alpha Fodrin | IKLLQAQKL | Eubacteriaceae | Glycosyltransferase | 100.0% |
| Candidatus Pseudoramibacter | Glycosyltransferase | 100.0% | |||
| HLA- DRB1*15:01 | Alpha Fodrin | EVLDRWRRL | Desulfonatronum sp. | Thioredoxin | 88.9% |
| Thermoleophilaceae bacterium | Proline RNA ligase | 88.9% | |||
| Thermoleophilaceae bacterium | Proline tRNA ligase | 88.9% | |||
| Nonomuraea nitratireducens | DUF885 family protein | 88.9% | |||
| Nonomuraea phyllanthi | DUF885 domain-containing protein | 88.9% | |||
| Firmicutes bacterium | Biotin protein ligase | 88.9% | |||
| Firmicutes bacterium | Biotin protein ligase | 88.9% | |||
| Streptomyces malaysiensis | Putative non-ribosomal peptide synthetase | 100.0% | |||
| Streptomyces malaysiensis | Non-ribosomal peptide synthetase | 100.0% | |||
| Streptomyces malaysiensis | Carrier domain-containing protein | 100.0% | |||
| Aquisphaera giovannonii | Phosphomannomutase/phosphoglucomutase | 100.0% | |||
| Streptomycetaceae bacterium | Uncharacterized protein | 88.9% | |||
| Curtobacterium sp. MCPF17_047 | Uncharacterized protein | 100.0% | |||
| Nitriliruptorales bacterium | DUF1932 domain-containing protein | 100.0% | |||
| Paracoccus homiensis | Acetyltransferase (GNAT) family protein | 100.0% | |||
| Actinophytocola xanthii | SnoaL-like domain-containing protein | 100.0% | |||
| Frigoribacterium sp. PhB160 | S-DNA-T family DNA segregation ATPase | 100.0% | |||
| Frigoribacterium sp. PhB107 | S-DNA-T family DNA segregation ATPase | 100.0% | |||
| Frigoribacterium sp. ACAM 257 | Cell division protein FtsK | 100.0% | |||
| Geodermatophilus sp. DF01_2 | Peptidase_M16_C domain-containing protein | 100.0% | |||
| Acidobacteria bacterium | Uncharacterized protein | 100.0% | |||
| Nitrosococcus oceani C-27 | Transposase | 88.9% | |||
| Nitrosococcus oceani (strain) | Y1_Tnp domain-containing protein | 88.9% | |||
| Dietzia sp. MeA6-2017 | Uncharacterized protein | 100.0% | |||
| Firmicutes bacterium | Bifunctional ligase/repressor BirA | 100.0% | |||
| Dietzia sp. oral taxon 368 | Uncharacterized protein | 100.0% | |||
| Saccharopolyspora sp. ASAGF58 | Uncharacterized protein | 100.0% | |||
| Saccharopolyspora spinosa | Uncharacterized protein | 100.0% | |||
| Chloroflexi bacterium | Biotin [acetyl-CoA-carboxylase] ligase | 100.0% | |||
| Actinobacteria bacterium 13 | Biotin [acetyl-CoA-carboxylase] ligase | 100.0% | |||
| Pelagibaca abyssi | Uncharacterized protein | 100.0% | |||
| Candidatus Kentron sp. LFY | Type III restriction enzyme | 88.9% | |||
| Planctomycetes bacterium | Diguanylate cyclase | 88.9% | |||
| Candidatus Kentron sp. LFY | Type III restriction enzyme, res subunit | 88.9% | |||
| Candidatus Solibacter sp. | 3-isopropylmalate dehydratase large subunit | 88.9% | |||
| Hyalangium minutum | Uncharacterized protein | 88.9% | |||
| Actinokineospora terrae | AraC-type DNA-binding protein | 88.9% | |||
| Actinokineospora cianjurensis | AraC-like DNA-binding protein | 88.9% | |||
| HLA-DRB4*01:01 | Alpha Fodrin | IKLLQAQKL | Eubacteriaceae Candidatus Pseudoramibacter | Glycosyltransferase Glycosyltransferase | 100.0% 100.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, S.; Li, D.; Ostrov, D.A.; Nguyen, C.Q. Epitope Mapping of Pathogenic Autoantigens on Sjögren’s Syndrome-Susceptible Human Leukocyte Antigens Using In Silico Techniques. J. Clin. Med. 2022, 11, 1690. https://doi.org/10.3390/jcm11061690
Gupta S, Li D, Ostrov DA, Nguyen CQ. Epitope Mapping of Pathogenic Autoantigens on Sjögren’s Syndrome-Susceptible Human Leukocyte Antigens Using In Silico Techniques. Journal of Clinical Medicine. 2022; 11(6):1690. https://doi.org/10.3390/jcm11061690
Chicago/Turabian StyleGupta, Shivai, Danmeng Li, David A. Ostrov, and Cuong Q. Nguyen. 2022. "Epitope Mapping of Pathogenic Autoantigens on Sjögren’s Syndrome-Susceptible Human Leukocyte Antigens Using In Silico Techniques" Journal of Clinical Medicine 11, no. 6: 1690. https://doi.org/10.3390/jcm11061690
APA StyleGupta, S., Li, D., Ostrov, D. A., & Nguyen, C. Q. (2022). Epitope Mapping of Pathogenic Autoantigens on Sjögren’s Syndrome-Susceptible Human Leukocyte Antigens Using In Silico Techniques. Journal of Clinical Medicine, 11(6), 1690. https://doi.org/10.3390/jcm11061690