
Placenta-on-a-Chip as an In Vitro Approach to Evaluate the Physiological and Structural Characteristics of the Human Placental Barrier upon Drug Exposure: A Systematic Review
 , ,
, ,  , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Study Selection
2.4. Data Extraction
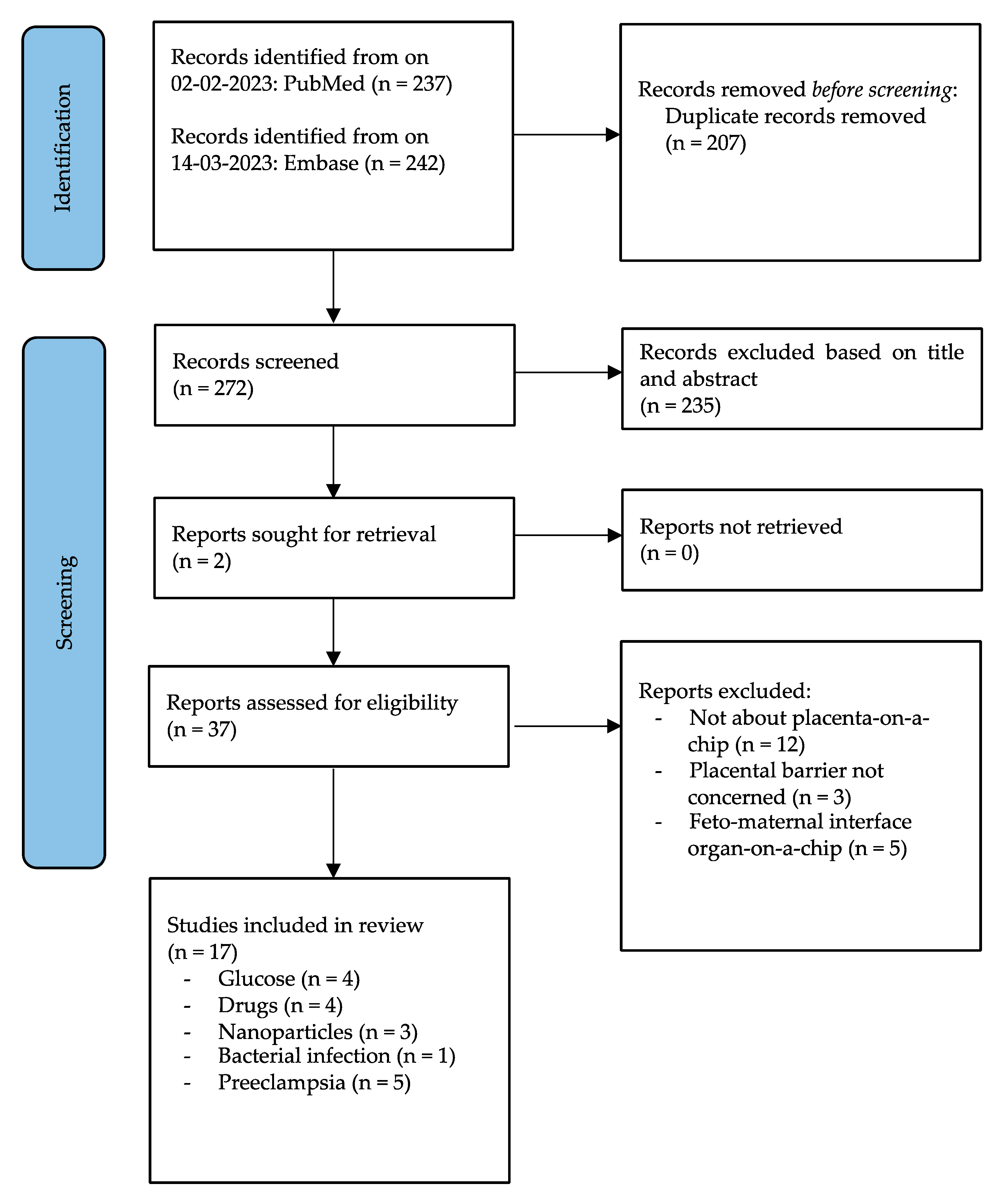
3. Results
3.1. Study Selection and Data Extraction
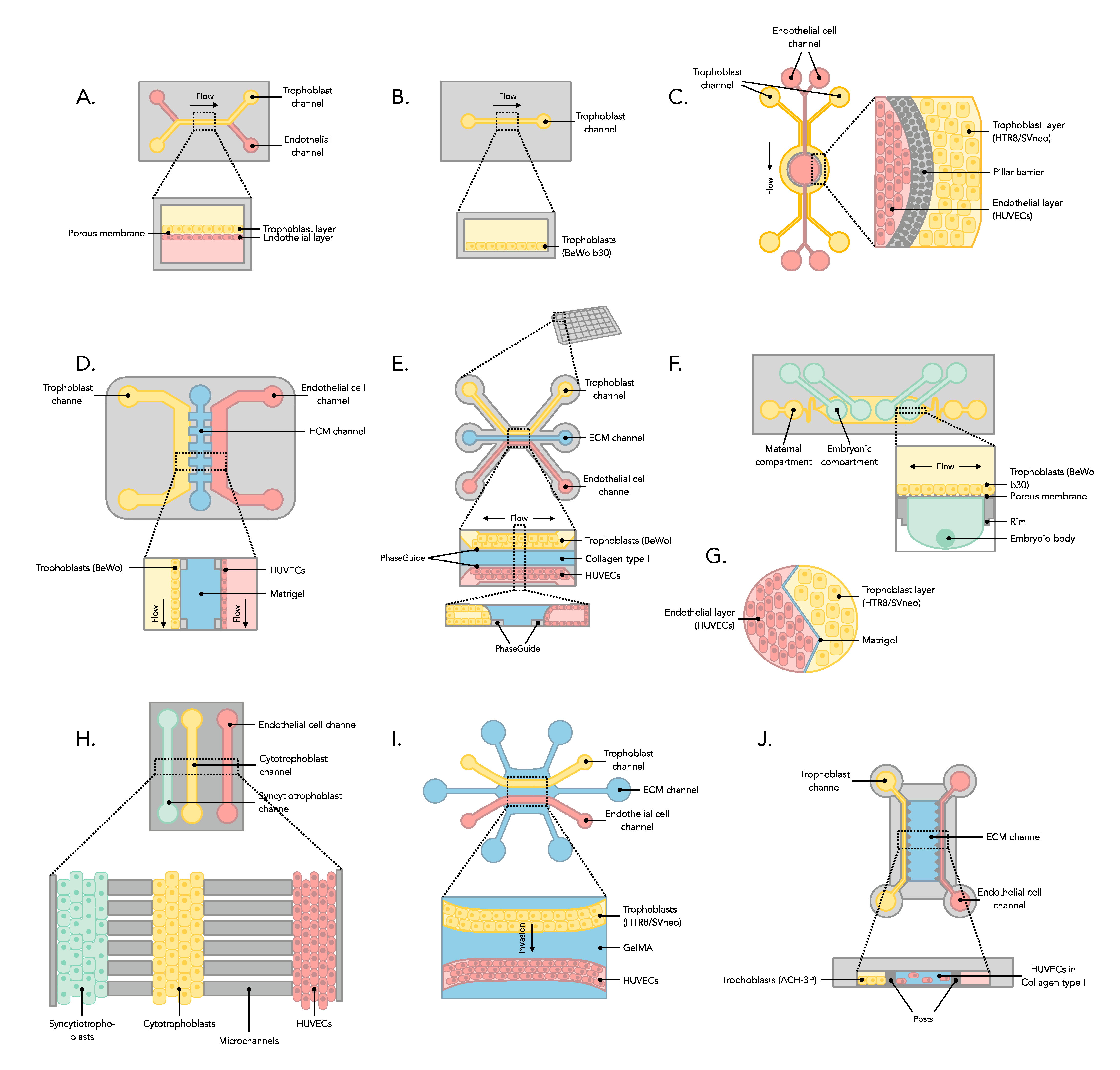
3.2. Manufacture of Placenta-on-a-Chip Microfluidic Device
3.3. Analysis of Experiments from Placenta-on-a-Chip
3.3.1. Glucose
3.3.2. Drugs
3.3.3. Nanoparticles
3.3.4. Bacterial Exposure
3.3.5. Preeclampsia
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eurocat—Use of Teratogenic Medication during Pregnancy. Available online: https://umcgresearch.org/w/eurocat-use-of-teratogenic-medication-during-pregnancy (accessed on 28 February 2023).
- Blundell, C.; Yi, Y.S.; Ma, L.; Tess, E.R.; Farrell, M.J.; Georgescu, A.; Aleksunes, L.M.; Huh, D. Placental Drug Transport-on-a-Chip: A Microengineered In Vitro Model of Transporter-Mediated Drug Efflux in the Human Placental Barrier. Adv. Healthc. Mater. 2018, 7, 1700786. [Google Scholar] [CrossRef]
- Pemathilaka, R.L.; Reynolds, D.E.; Hashemi, N.N. Drug transport across the human placenta: Review of placenta-on-a-chip and previous approaches. Interface Focus 2019, 9, 20190031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosavati, B.; Oleinikov, A.; Du, E. 3D microfluidics-assisted modeling of glucose transport in placental malaria. Sci. Rep. 2022, 12, 15278. [Google Scholar] [CrossRef] [PubMed]
- Gude, N.M.; Roberts, C.T.; Kalionis, B.; King, R.G. Growth and function of the normal human placenta. Thromb. Res. 2004, 114, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Rubinchik-Stern, M.; Eyal, S. Drug Interactions at the Human Placenta: What is the Evidence? Front. Pharmacol. 2012, 3, 126. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.K.; Campbell, J.P. Placental structure, function and drug transfer. Contin. Educ. Anaesth. Crit. Care Pain 2015, 15, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Shetty, S.; Gogate, A.; Gogate, S.; Malet, P. A reproducible modified method for direct preparation of chorionic villi cytogenetic analysis. Methods Cell Sci. 2003, 25, 149–154. [Google Scholar] [CrossRef]
- Kojima, J.; Ono, M.; Kuji, N.; Nishi, H. Human Chorionic Villous Differentiation and Placental Development. Int. J. Mol. Sci. 2022, 23, 8003. [Google Scholar] [CrossRef]
- Abostait, A.; Tyrrell, J.; Abdelkarim, M.; Shojaei, S.; Tse, W.H.; El-Sherbiny, I.M.; Keijzer, R.; Labouta, H.I. Placental Nanoparticle Uptake-On-a-Chip: The Impact of Trophoblast Syncytialization and Shear Stress. Mol. Pharm. 2022, 19, 3757–3769. [Google Scholar] [CrossRef]
- Miller, R.K.; Genbacev, O.; Turner, M.A.; Aplin, J.D.; Caniggia, I.; Huppertz, B. Human placental explants in culture: Approaches and assessments. Placenta 2005, 26, 439–488. [Google Scholar] [CrossRef]
- Hutson, J.R.; Garcia-Bournissen, F.; Davis, A.; Koren, G. The human placental perfusion model: A systematic review and development of a model to predict in vivo transfer of therapeutic drugs. Clin. Pharmacol. Ther. 2011, 90, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, P.; Patel, B.G.; Patel, B.K. Drug use in pregnancy; a point to ponder. Indian J. Pharm. Sci. 2009, 71, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bode, C.J.; Jin, H.; Rytting, E.; Silverstein, P.S.; Young, A.M.; Audus, K.L. In vitro models for studying trophoblast transcellular transport. Methods Mol. Med. 2006, 122, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Prast, J.; Saleh, L.; Husslein, H.; Sonderegger, S.; Helmer, H.; Knöfler, M. Human chorionic gonadotropin stimulates trophoblast invasion through extracellularly regulated kinase and AKT signalling. Endocrinology 2008, 149, 979–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, R.E.; Metz, T.D.; Ward, R.M.; McKnite, A.M.; Enioutina, E.Y.; Sherwin, C.M.; Watt, K.M.; Job, K.M. Drug exposure during pregnancy: Current understanding and approaches to measure maternal-fetal drug exposure. Front. Pharmacol. 2023, 14, 1111601. [Google Scholar] [CrossRef] [PubMed]
- PRISMA. Available online: https://prisma-statement.org/ (accessed on 19 March 2023).
- Blundell, C.; Tess, E.R.; Schanzer, A.S.; Coutifaris, C.; Su, E.J.; Parry, S.; Huh, D. A microphysiological model of the human placental barrier. Lab Chip 2016, 16, 3065–3073. [Google Scholar] [CrossRef] [Green Version]
- Mosavati, B.; Oleinikov, A.V.; Du, E. Development of an Organ-on-a-Chip-Device for Study of Placental Pathologies. Int. J. Mol. Sci. 2020, 21, 8755. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Romero, R.; Han, Y.M.; Kim, H.C.; Kim, C.J.; Hong, J.S.; Huh, D. Placenta-on-a-chip: A novel platform to study the biology of the human placenta. J. Matern. Fetal Neonatal Med. 2016, 29, 1046–1054. [Google Scholar] [CrossRef]
- Pemathilaka, R.L.; Alimoradi, N.; Reynolds, D.E.; Hashemi, N.N. Transport of Maternally Administered Pharmaceutical Agents Across the Placental Barrier In Vitro. ACS Appl. Bio Mater. 2022, 5, 2273–2284. [Google Scholar] [CrossRef]
- Pemathilaka, R.L.; Caplin, J.D.; Aykar, S.S.; Montazami, R.; Hashemi, N.N. Placenta-on-a-Chip: In Vitro Study of Caffeine Transport across Placental Barrier Using Liquid Chromatography Mass Spectrometry. Glob. Chall. 2019, 2, 1800112. [Google Scholar] [CrossRef] [Green Version]
- Richardson, L.S.; Kammala, A.K.; Costantine, M.M.; Fortunato, S.J.; Radnaa, E.; Kim, S.; Taylor, R.N.; Han, A.; Menon, R. Testing of drugs using human feto-maternal interface organ-on-chips provide insights into pharmacokinetics and efficacy. Lab Chip 2022, 22, 4574–4592. [Google Scholar] [CrossRef]
- Yin, F.; Zhu, Y.; Zhang, M.; Yu, H.; Chen, W.; Qin, J. A 3D human placenta-on-a-chip model to probe nanoparticle exposure at the placental barrier. Toxicol. Vitr. 2019, 54, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Boos, J.A.; Misun, P.M.; Brunoldi, G.; Furer, L.A.; Aengenheister, L.; Modena, M.; Rousset, N.; Buerki-Thurnherr, T.; Hierlemann, A. Microfluidic Co-Culture Platform to Recapitulate the Maternal-Placental-Embryonic Axis. Adv. Biol. 2021, 5, e2100609. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, F.; Wang, H.; Wang, L.; Yuan, J.; Qin, J. Placental Barrier-on-a-Chip: Modeling Placental Inflammatory Responses to Bacterial Infection. ACS Biomater. Sci. Eng. 2018, 4, 3356–3363. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.; Jeon, T.J.; Kim, S.M. Trophoblast Migration with Different Oxygen Levels in a Gel-Patterned Microfluidic System. Micromachines 2022, 13, 2216. [Google Scholar] [CrossRef]
- Cho, H.; Fuwad, A.; Ko, G.; Kim, G.B.; Jeon, T.; Kim, S.Y. A PDMS-Based Interdigitated Platform for Trophoblast Invasion Study Under Oxygen Stress Conditions. Biochip J. 2021, 15, 362–370. [Google Scholar] [CrossRef]
- Rabussier, G.; Bünter, I.; Bouwhuis, J.; Soragni, C.; van Zijp, T.; Ng, C.P.; Dormansky, K.; De Windt, L.J.; Vulto, P.; Murdoch, C.E.; et al. Healthy and diseased placenta barrier on-a-chip models suitable for high-throughput studies. Acta Biomater. 2022, 164, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanpour, S.M.; Richards, C.; Pienaar, D.; Sesperez, K.; Aboulkheyr Es, H.; Nikolic, V.N.; Karadzov Orlic, N.; Mikovic, Z.; Stefanovic, M.; Cakic, Z.; et al. A placenta-on-a-chip model to determine the regulation of FKBPL and galectin-3 in preeclampsia. Cell. Mol. Life Sci. 2023, 80, 44. [Google Scholar] [CrossRef]
- Pu, Y.; Gingrich, J.; Veiga-Lopez, A. A 3-dimensional microfluidic platform for modeling human extravillous trophoblast invasion and toxicological screening. Lab Chip 2021, 21, 546–557. [Google Scholar] [CrossRef]
- Murchison, A.C.; Odanga, J.J.; Treadwell, M.L.; Breathwaite, E.K.; Weaver, J.R.; Lee, J.B. Human Placenta-Derived ECM Supports Tri-Lineage Differentiation of Human Induced Pluripotent Stem Cells. Int. J. Stem Cells 2020, 13, 432–438. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, S.; Liu, B.; Miao, Y.; Hu, Z. Use of extracellular matrix hydrogen from human placenta to restore hair-inductive potential of dermal papilla cells. Regen. Med. 2019, 14, 741–751. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, H.; Qin, H.; Yang, J.; Wang, Y.; Jiang, S.; Pan, Y. Vascular Endothelial Growth Factor expression in peripheral blood of patients with pregnancy induced hypertension syndrome and its clinical significance. Pak. J. Med. Sci. 2014, 30, 634–637. [Google Scholar] [CrossRef]
- Mitchell, A.A.; Gilboa, S.M.; Werler, M.M.; Kelley, K.E.; Louik, C.; Hernández-Díaz, S.; National Birth Defects Prevention Study. Medication use during pregnancy, with particular focus on prescription drugs: 1976–2008. Am. J. Obstet. Gynecol. 2011, 205, e1–e51. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, M.A.; Fernando, R.C.; Gardner, L.; Hollinshead, M.S.; Burton, G.J.; Moffett, A.; Turco, M.Y. Establishment and differentiation of long-term trophoblast organoid cultures from the human placenta. Nat. Protoc. 2020, 15, 3441–3463. [Google Scholar] [CrossRef]
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.S.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal-fetal interactions during human placentation. Nature 2018, 563, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karvas, R.M.; Khan, S.A.; Verma, S.; Yin, Y.; Kulkarni, D.; Dong, C.; Park, K.M.; Chew, B.; Sane, E.; Fischer, L.A.; et al. Stem-cell-derived trophoblast organoids model human placental development and susceptibility to emerging pathogens. Cell Stem Cell 2022, 29, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Meinhardt, G.; Saleh, L.; Kunihs, V.; Gamperl, M.; Kaindl, U.; Ellinger, A.; Burkard, T.R.; Fiala, C.; Pollheimer, J.; et al. Self-Renewing Trophoblast Organoids Recapitulate the Developmental Program of the Early Human Placenta. Stem Cell Rep. 2018, 11, 537–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, J.; Wei, W.; Chen, Z.; Lin, B.; Zhao, W.; Luo, Y.; Zhang, X. Engineered Liver-on-a-Chip Platform to Mimic Liver Functions and Its Biomedical Applications: A Review. Micromachines 2019, 10, 676. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, E.; Rasponi, M. Liver-Heart on chip models for drug safety. APL Bioeng. 2021, 5, 031505. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, X.; Cribbin, E.M.; Kim, A.M.; Li, J.J.; Yong, K.T. Liver-on-a-chip: Considerations, advances, and beyond. Biomicrofluidics 2022, 16, 061502. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Yang, Y.; Yan, J.; Xiong, Y.; Wang, W.; Lei, J.; Jiang, T. Recapitulating essential pathophysiological characteristics in lung-on-a-chip for disease studies. Front. Immunol. 2023, 14, 1093460. [Google Scholar] [CrossRef]
- Yang, X.; Li, K.; Zhang, X.; Liu, C.; Guo, B.; Wen, W.; Gao, X. Nanofiber membrane supported lung-on-a-chip microdevice for anti-cancer drug testing. Lab Chip 2018, 18, 486–495. [Google Scholar] [CrossRef]
- Chen, S.L.; Chou, H.C.; Lin, K.C.; Yang, J.W.; Xie, R.H.; Chen, C.Y.; Liu, X.Y.; Chung, J.H.Y.; Chen, G.Y. Investigation of the role of the autophagic protein LC3B in the regulation of human airway epithelium cell differentiation in COPD using a biomimetic model. Mater. Today 2021, 13, 100182. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.; Choi, Y.M.; Cho, S.; Gao, G.; Kim, D.; Kim, J.; Choi, H.; Lee, S.H.; Jang, J. Modular assembly of bioprinted perfusable blood vessel and tracheal epithelium for studying inflammatory respiratory diseases. Biofabrication 2022, 15, 014101. [Google Scholar] [CrossRef] [PubMed]
- Sundarakrishnan, A.; Zukas, H.; Coburn, J.; Bertini, B.T.; Liu, Z.; Georgakoudi, I.; Baugh, L.; Dasgupta, Q.; Black, L.D.; Kaplan, D.L. Bioengineered in Vitro Tissue Model of Fibroblast Activation for Modeling Pulmonary Fibrosis. ACS Biomater. Sci. Eng. 2019, 5, 2417–2429. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Huh, D.; Hamilton, G.; Ingber, D.E. Human gut-on-a-chip inhabited by microbial flora that experiences intestinal peristalsis-like motions and flow. Lab Chip 2012, 12, 2165–2174. [Google Scholar] [CrossRef]
- Kasendra, M.; Tovaglieri, A.; Sontheimer-Phelps, A.; Jalili-Firoozinezhad, S.; Bein, A.; Chalkiadaki, A.; Scholl, W.; Zhang, C.; Rickner, H.; Richmond, C.A.; et al. Development of a primary human Small Intestine-on-a-Chip using biopsy-derived organoids. Sci. Rep. 2018, 8, 2871. [Google Scholar] [CrossRef] [Green Version]
- Olivier, E.; Wakx, A.; Fouyet, S.; Dutot, M.; Rat, P. JEG-3 placental cells in toxicology studies: A promising tool to reveal pregnancy disorders. Anat. Cell Biol. 2021, 54, 83–92. [Google Scholar] [CrossRef]
- van Meer, B.J.; de Vries, H.; Firth, K.S.A.; van Weerd, J.; Tertoolen, L.G.J.; Karperien, H.B.J.; Jonkheijm, P.; Denning, C.; IJzerman, A.P.; Mummery, C.L. Small molecule absorption by PDMS in the context of drug response bioassays. Biochem. Biophys. Res. Commun. 2017, 482, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Nunes, P.S.; Ohlsson, P.D.; Ordeig, O.; Kutter, J.P. Cyclic olefin polymers: Emerging materials for lab-on-a-chip applications. Microfluid. Nanofluidics 2010, 9, 145–161. [Google Scholar] [CrossRef]
- Morley, L.C.; Beech, D.J.; Walker, J.J.; Simpson, N.A.B. Emerging concepts of shear stress in placental development and function. Mol. Hum. Reprod. 2019, 25, 329–339. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Placenta-on-a-Chip | Placenta |
|---|---|
| “Placenta-on-a-chip”[Mesh] placenta-on-chip*[tiab], placenta model*[tiab], placental model*[tiab], placenta in vitro model*[tiab], “Lab-On-a-chip devices”[Mesh] on-a-chip*[tiab], nanochip*[tiab], microchip*[tiab], microfluidic*[tiab] | “Placenta”[Mesh] placenta*[tiab], placentome*[tiab], decidua*[tiab], deciduoma*[tiab], trophoblast*[tiab], cytotrophoblast*[tiab], syncytiotrophoblast*[tiab] |
| Placenta-on-a-Chip | Placenta |
|---|---|
| ‘placenta-on-a-chip’/exp placenta-on-chip*:ab,ti, placenta model*:ab,ti, placental model*:ab,ti, placenta in vitro model*:ab,ti, ‘lab on a chip’/exp on-a-chip*:ab,ti, nanochip*:ab,ti, microchip*:ab,ti, microfluidic*:ab,ti | ‘placenta’/exp placenta*:ab,ti, placentome*:ab,ti, decidua*:ab,ti, deciduoma*:ab,ti, trophoblast*:ab,ti, cytotrophoblast*:ab,ti, syncytiotrophoblast*:ab,ti |
| Type of Testing | Aim | Cultured Cells | Properties | Chip Type | Main Findings | Reference |
|---|---|---|---|---|---|---|
| Glucose | Placental glucose transport | BeWo b30 cells (Trophoblast cell line) HPVECs (Endothelial cell line) | Substance tested: Glucose concentration: Maternal channel: 10 mM Fetal channel: 5.5 mM Exposure 2 h Placental barrier: Flow rate: 100 μL/h Porous membrane: polycarbonate, pore size 1 μm ECM: Fibronectin coating (0.1 mg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Blundell et al. [18] |
| Glucose | Placental glucose transport | JEG-3 (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Glucose concentration: Maternal channel: 25 mM Fetal channel: 6.1 mM Exposure 68 h Placental barrier: Flow rate: 30 μL/h Porous membrane: Vitrified Collagen-I (2.43 mg/mL) ECM: Fibronectin coating (40 mg/mL, upper channel) and gelatin coating (1.5%, lower channel) | In-house manufactured using soft lithography, PDMS-based |
| Lee et al. [20] |
| Glucose | Placental glucose transport | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Glucose concentration: Maternal channel: 7.2 mM Fetal channel: 5.6 mM Exposure 2 h Placental barrier: Flow rate: 50 μL/h Porous membrane: polycarbonate, pore size 0.4 μm ECM: Collagen type I coating (concentration n.r.) | In-house manufactured using soft lithography, PDMS-based |
| Mosavati et al. [19] |
| Glucose | Placental glucose transport under influence of placental malaria | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Glucose concentration: Maternal channel: 8.9 mM Fetal channel: 5.4 mM Exposure 2 h Placental barrier: Flow rate: n/a. Shear stress: 0–1.41 dyne/cm2 ECM: Collagen type I gel (0.4 mg/mL), pore size 2–3 μm | 3-lane OrganoPlate (MIMETAS) |
| Mosavati et al. [4] |
| Type of Testing | Aim | Cultured Cells | Properties | Chip Type | Main Findings | Reference |
|---|---|---|---|---|---|---|
| Naltrexone/ 6ß-Naltrexol | Placental drug transport and fetal drug exposure | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Naltrexone concentration: 293 mM 6ß-Naltrexol concentration: 291 mM Exposure 8 h Placental barrier: Flow rate: 50 μL/h Porous membrane: polyethylene terephthalate, pore size: 0.4 μm ECM: Entactin collagen IV-laminin coating (10 μg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Pemathilaka et al. [21] |
| Caffeine | Rate of placental caffeine transport | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Caffeine concentration: 1.3 mM Exposure 7.5 h Placental barrier: Flow rate: 50 μL/h Porous membrane: polyethylene terephthalate, pore size: 0.4 μm ECM: Entactin collagen IV-laminin coating (10 μg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Pemathilaka et al. [22] |
| Glyburide | Active placental drug transport | BeWo b30 cells (Trophoblast cell line) HPVECs (Endothelial cell line) | Substance tested: Heparin size: 3000–15,000 kDa Exposure 5 h Glyburide concentration: 1.0 × 10−3 mM (BODIPY-conjugated Glyburide) Exposure 3 h Placental barrier: Flow rate: 100 μL/h Porous membrane: polycarbonate, pore size: 1 μm ECM: Fibronectin coating (0.1 mg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Blundell et al. [2] |
| Rosuvastatin/ Pravastatin | Placental drug transport, efficacy and kinetics of statins | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Heparin (0.1 mg/mL) Pravastatin concentration: 4.7 × 10−4 mM Rosuvastatin concentration: 4.2 × 10−4 mM Exposure 4–24 h Placental barrier: Array of 24 microchannels, 5 μm in height, 30 μm in width, 300/600 μm in length, connecting syncytiotrophoblast, cytotrophoblast and endothelial cell compartments ECM: Collagen type I coating (10 μg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Richardson et al. [23] |
| Type of Testing | Aim | Cultured Cells | Properties | Chip Type | Main Findings | Reference |
|---|---|---|---|---|---|---|
| Titanium dioxide nanoparticles | Placental responses to nanoparticles | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Particles: TiO2-nanoparticles: 50 or 200 μg/mL Size: 50 nm (diameter) Exposure 24 h Placental barrier: Flow rate: 20 μL/h Shear stress: 0.03 dyne/cm2 ECM barrier: Matrigel ECM: Chitosan coating (2%) | In-house manufactured using soft lithography, PDMS-based |
| Yin et al. [24] |
| Carboxyl-modified polystyrene microparticles (PS-MPs) | Systemic (in)direct embryotoxicity | BeWo b30 cells (Trophoblast cell line) Embryoid bodies | Particles: PS-MPs concentration: 1, 10 or 100 μg/mL Size: 500 nm (diameter) Exposure 72 h Placental barrier: Flow rate: 276 μL/h Shear stress: 0.0003 dyne/cm2 Porous membrane: polyethylene terephthalate, pore size: 3 μm ECM: Collagen type I coating (2%) | In-house manufactured using soft lithography, PDMS-based |
| Boos et al. [25] |
| CSA-conjugated PEGylated liposomal nanocarriers | Trophoblast cell uptake under dynamic conditions and after chemically induced syncytialization | BeWo b30 cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Particles: Liposome concentration: 5 × 106–5 × 108 particles/mL Size: 93.7 nm (hydrodynamic diameter conjugated liposomes) 48.8/49.3 nm (hydrodynamic diameter unconjugated liposomes) Exposure 48/72 h Placental barrier: Flow rate: 1373/4 μL/h Shear stress: 0.025 dyne/cm2 ECM: Gelatin coated (concentration not reported) | iBidi μ-Slide I0.4 |
| Abostait et al. [10] |
| Type of Testing | Aim | Cultured Cells | Properties | Chip Type | Main Findings | Reference |
|---|---|---|---|---|---|---|
| Bacterial infection (E. coli) | Placental inflammatory responses with bacterial infection | BeWo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Particles: E. coli (rod-shaped bacterium 0.5 μm in width, 2 μm in length) Exposure 6 h Placental barrier: Flow rate: 10–40 μL/h Porous membrane: polyethylene terephthalate, pore size: 0.4 μm ECM: Collagen type I coating (0.1 mg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Zhu et al. [26] |
| Type of Testing | Aim | Cultured Cells | Properties | Chip Type | Main Findings | Reference |
|---|---|---|---|---|---|---|
| Preeclampsia (TNF-) | Assess effects inflammatory conditions in preeclampsia | ACH-3P (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: TNF- concentration: 1.0 × 10−5 g/L Exposure 24/72 h Placental barrier: Flow rate: pressure gradient ECM barrier: Collagen type I gel (2.5 mg/mL) | AIM Biotech idenTx chip |
| Ghorbanpour et al. [30] |
| Cell invasion | Trophoblast invasion with intraluminal flow | HTR8/SVneo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Substance tested: Folic acid concentration: 2.3 × 10−4 mM Exposure 24/48/72 h Placental barrier: Flow rate: 0.6, 3, 6 and 60 μL/h Shear stress: 0.046, 0.228, and 0.457 dyne/cm2 Barrier pillar spacing: 3 μm ECM: Fibronectin coating (200 μg/mL) | SynVivo 3D microfluidic chip |
| Pu et al. [31] |
| Oxygen levels | Effect of trophoblast invasion through oxygen level control | HTR8/Svneo (Trophoblast cell line) HUVECs (Endothelial cell line) | Conditions tested: Normoxic condition: 21% O2 Hypoxic conditions: 3% O2 Exposure 12/24/26 h Placental barrier: Flow rate: static system ECM barrier: Matrigel ECM: Fibronectin coating (40 μg/mL) | In-house manufactured using soft lithography, PDMS-based |
| Cho et al. [28] |
| Oxygen levels | Effect of oxygen level on trophoblast migration | HTR8/SVneo cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Conditions tested: Normoxic condition: 21% O2 Hypoxic conditions: 3% O2 Exposure 6/8 days Placental barrier: Flow rate: static system ECM barrier: Gelatin-methacrylate (GelMA) ECM: Fibronectin coating (50 ng/mL) | In-house manufactured using soft lithography, PDMS-based |
| Ko et al. [27] |
| Oxygen levels | Understand the underlying mechanism of preeclampsia | BeWo b30 cells (Trophoblast cell line) HUVECs (Endothelial cell line) | Conditions tested: Normoxic condition: 20% O2 Hypoxic conditions: 1% O2 Exposure 24/48/72 h Placental barrier: Flow rate: static system ECM barrier: Collagen-I/collagen-IV mixture (3:1 ratio) | 3-lane Organo Plate (MIMETAS) |
| Rabussier et al. [29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elzinga, F.A.; Khalili, B.; Touw, D.J.; Prins, J.R.; Olinga, P.; Leuvenink, H.G.D.; van Goor, H.; Gordijn, S.J.; Nagelkerke, A.; Mian, P. Placenta-on-a-Chip as an In Vitro Approach to Evaluate the Physiological and Structural Characteristics of the Human Placental Barrier upon Drug Exposure: A Systematic Review. J. Clin. Med. 2023, 12, 4315. https://doi.org/10.3390/jcm12134315
Elzinga FA, Khalili B, Touw DJ, Prins JR, Olinga P, Leuvenink HGD, van Goor H, Gordijn SJ, Nagelkerke A, Mian P. Placenta-on-a-Chip as an In Vitro Approach to Evaluate the Physiological and Structural Characteristics of the Human Placental Barrier upon Drug Exposure: A Systematic Review. Journal of Clinical Medicine. 2023; 12(13):4315. https://doi.org/10.3390/jcm12134315
Chicago/Turabian StyleElzinga, Femke A., Behrad Khalili, Daan J. Touw, Jelmer R. Prins, Peter Olinga, Henri G. D. Leuvenink, Harry van Goor, Sanne J. Gordijn, Anika Nagelkerke, and Paola Mian. 2023. "Placenta-on-a-Chip as an In Vitro Approach to Evaluate the Physiological and Structural Characteristics of the Human Placental Barrier upon Drug Exposure: A Systematic Review" Journal of Clinical Medicine 12, no. 13: 4315. https://doi.org/10.3390/jcm12134315
APA StyleElzinga, F. A., Khalili, B., Touw, D. J., Prins, J. R., Olinga, P., Leuvenink, H. G. D., van Goor, H., Gordijn, S. J., Nagelkerke, A., & Mian, P. (2023). Placenta-on-a-Chip as an In Vitro Approach to Evaluate the Physiological and Structural Characteristics of the Human Placental Barrier upon Drug Exposure: A Systematic Review. Journal of Clinical Medicine, 12(13), 4315. https://doi.org/10.3390/jcm12134315