

Cognitive Stress Regulation in Schizophrenia Patients and Healthy Individuals: Brain and Behavior
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Neurocognitive Abilities and Questionnaires
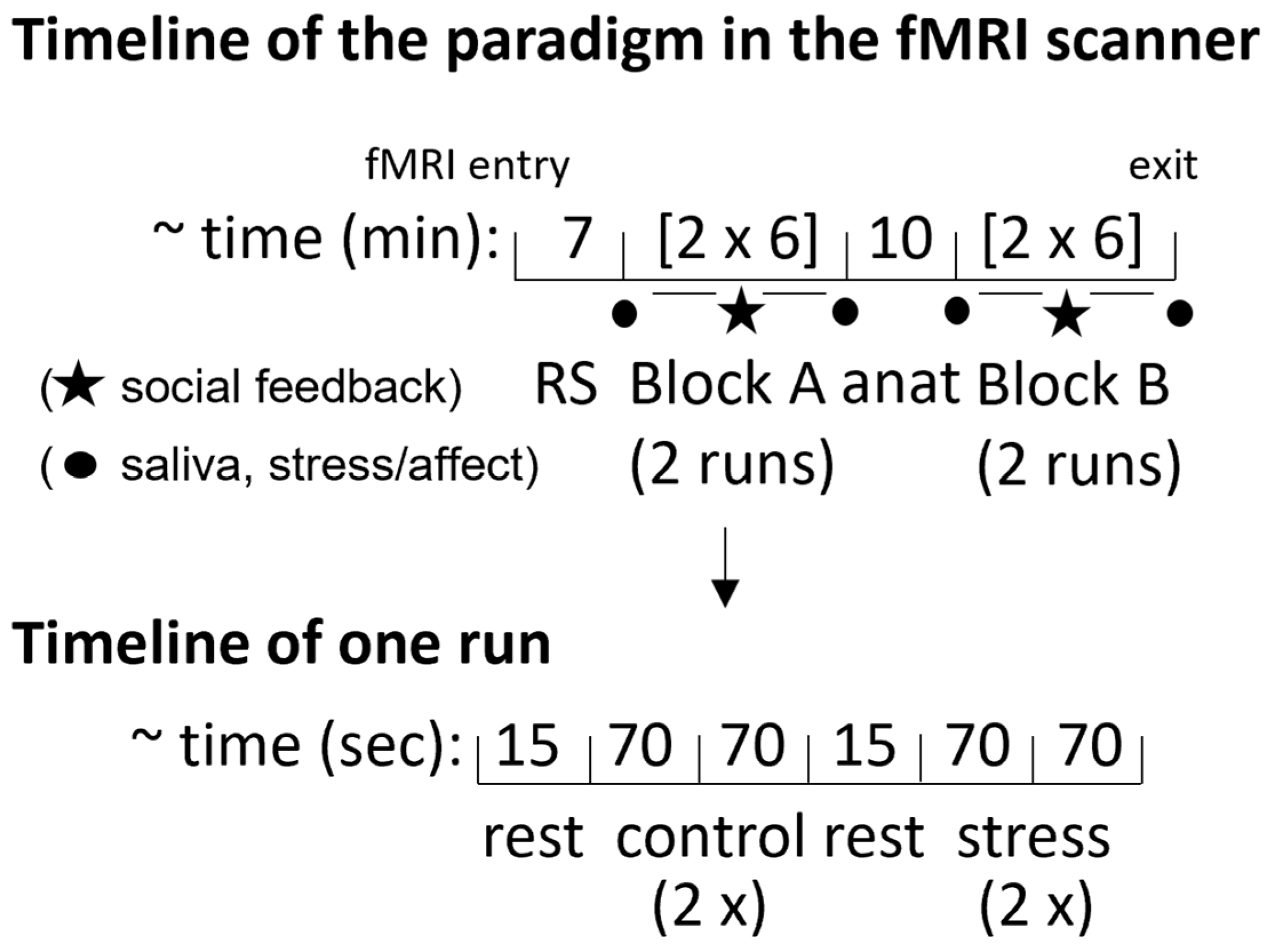
2.3. fMRI Stress Task
2.4. Statistical Analysis of Behavioral and Psychophysiological Data
2.5. Imaging Data Acquisition
2.6. fMRI Preprocessing and Analysis
2.7. Region-of-Interest Analysis
3. Results
3.1. Sample Description
3.2. Behavioral Results
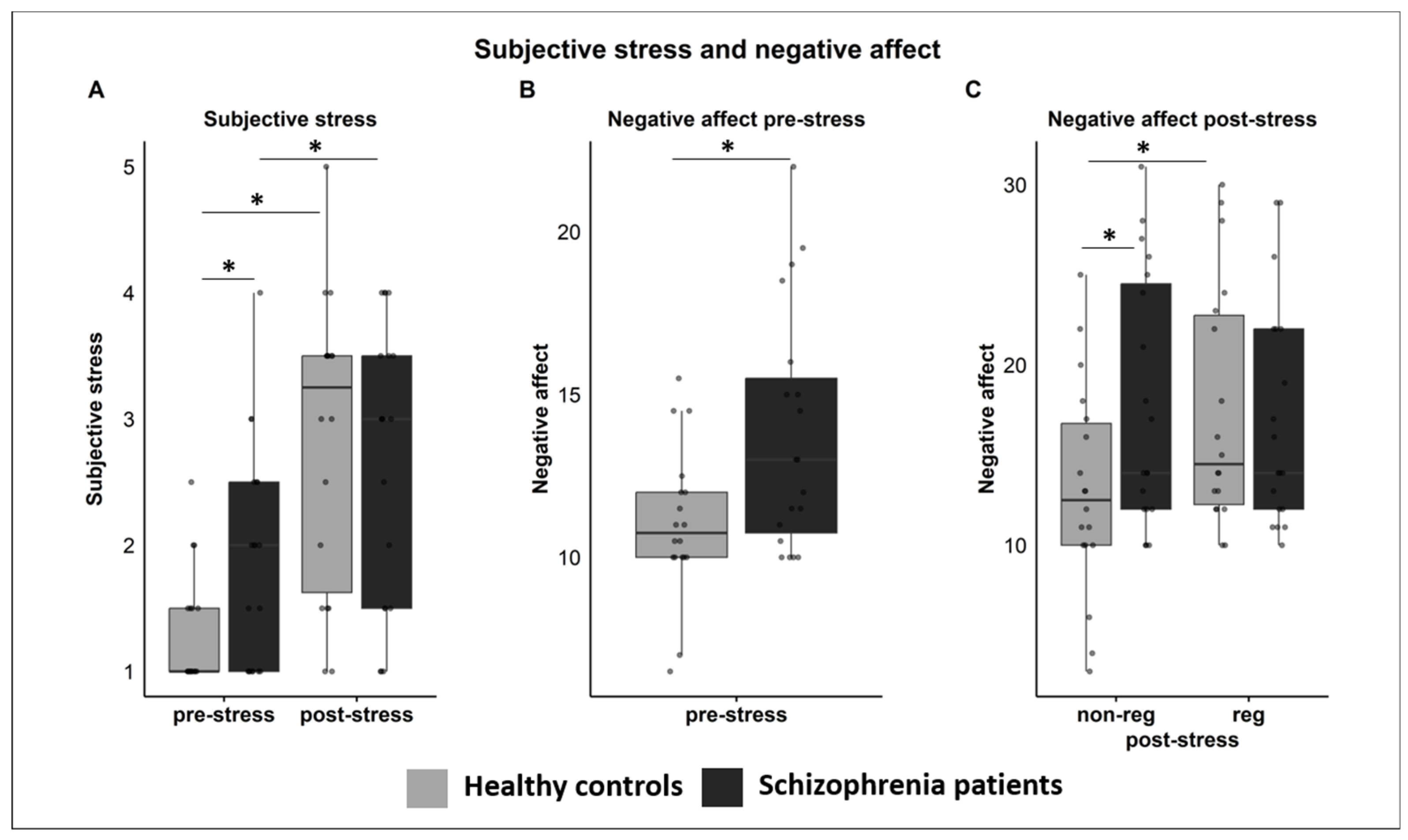
3.3. Subjective Ratings
3.3.1. Subjective Stress
3.3.2. Negative Affect
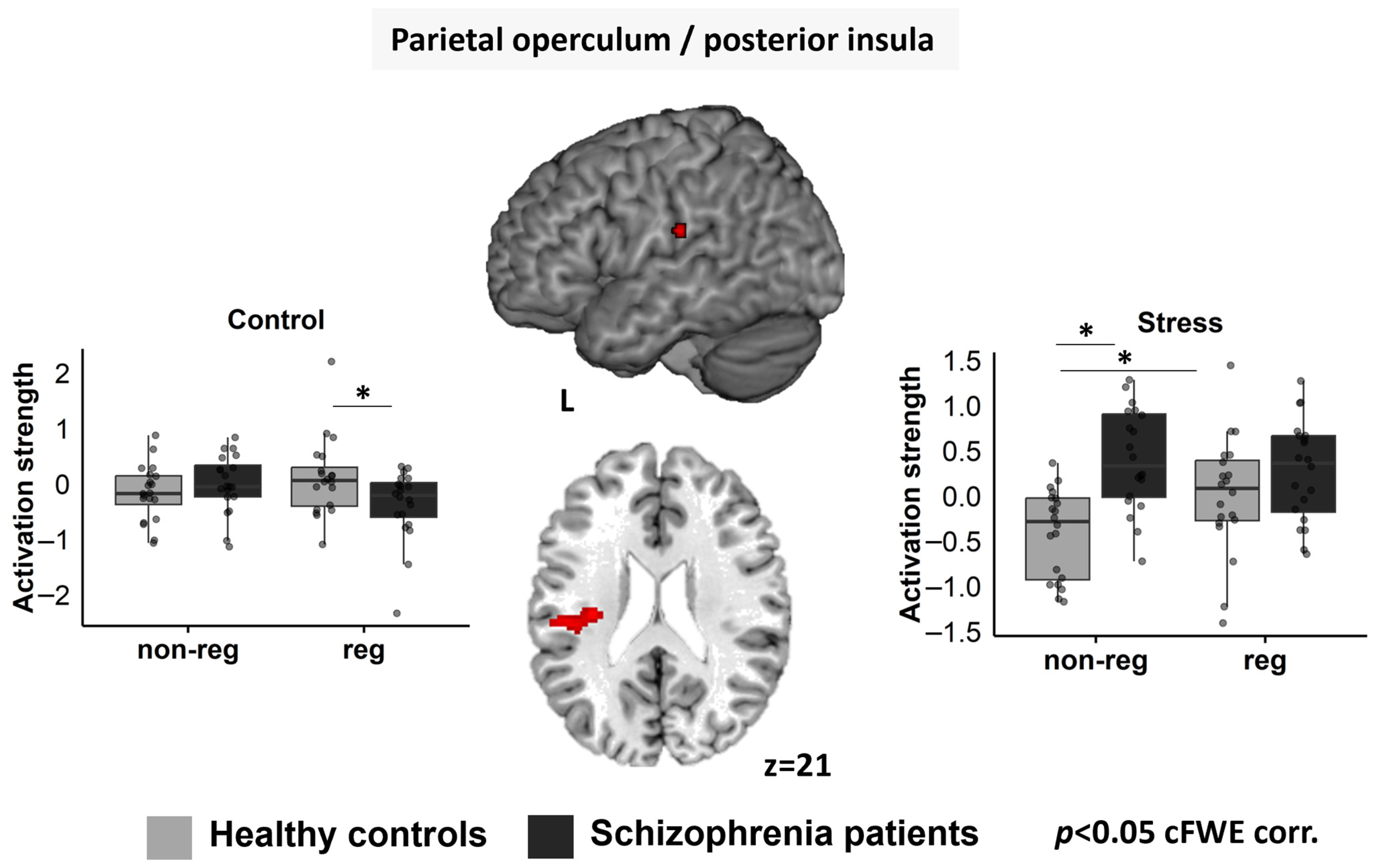
3.4. Whole-brain fMRI Data
Left pOP/pI
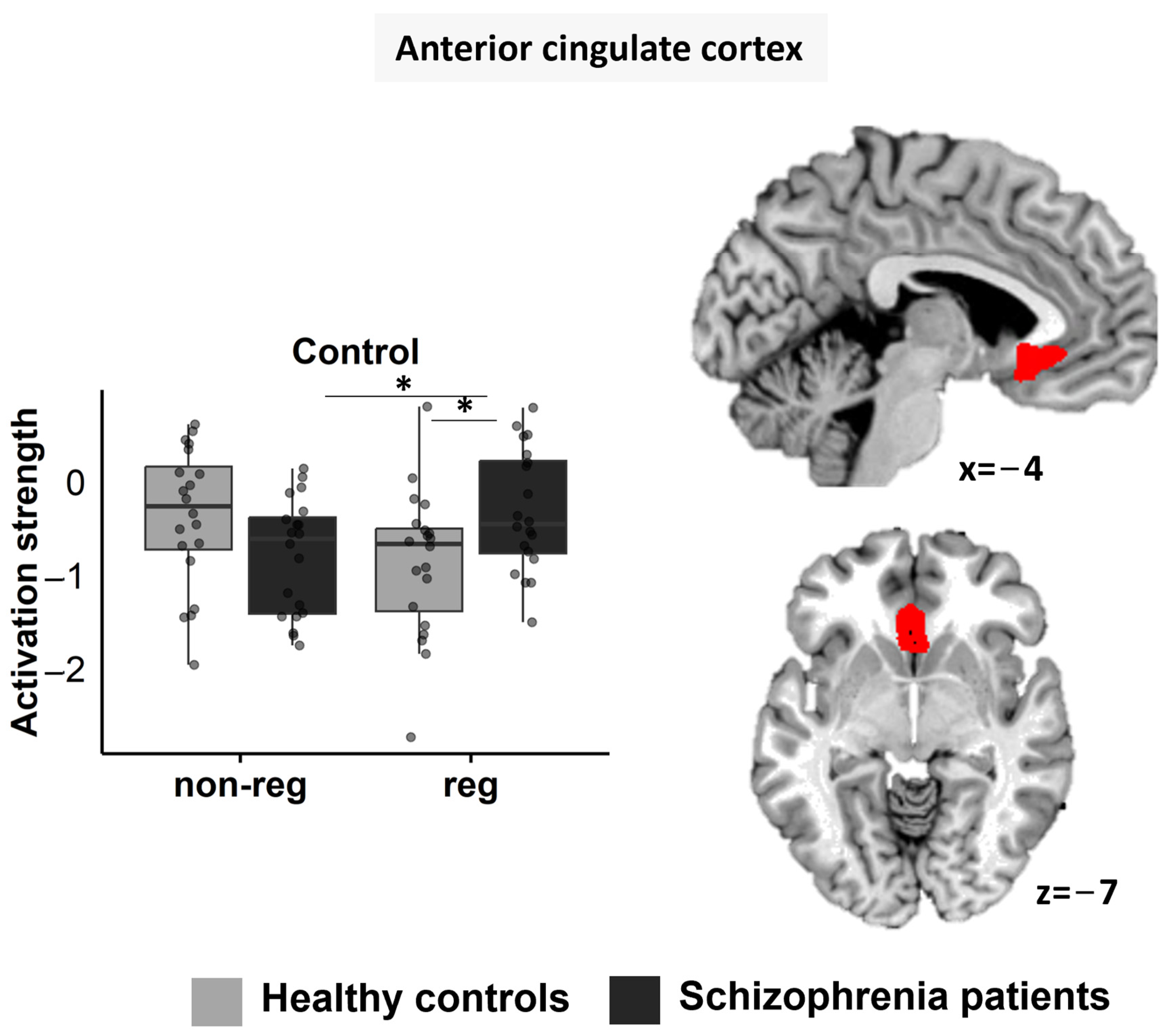
3.5. Region-of-Interest Analyses
3.5.1. ACC
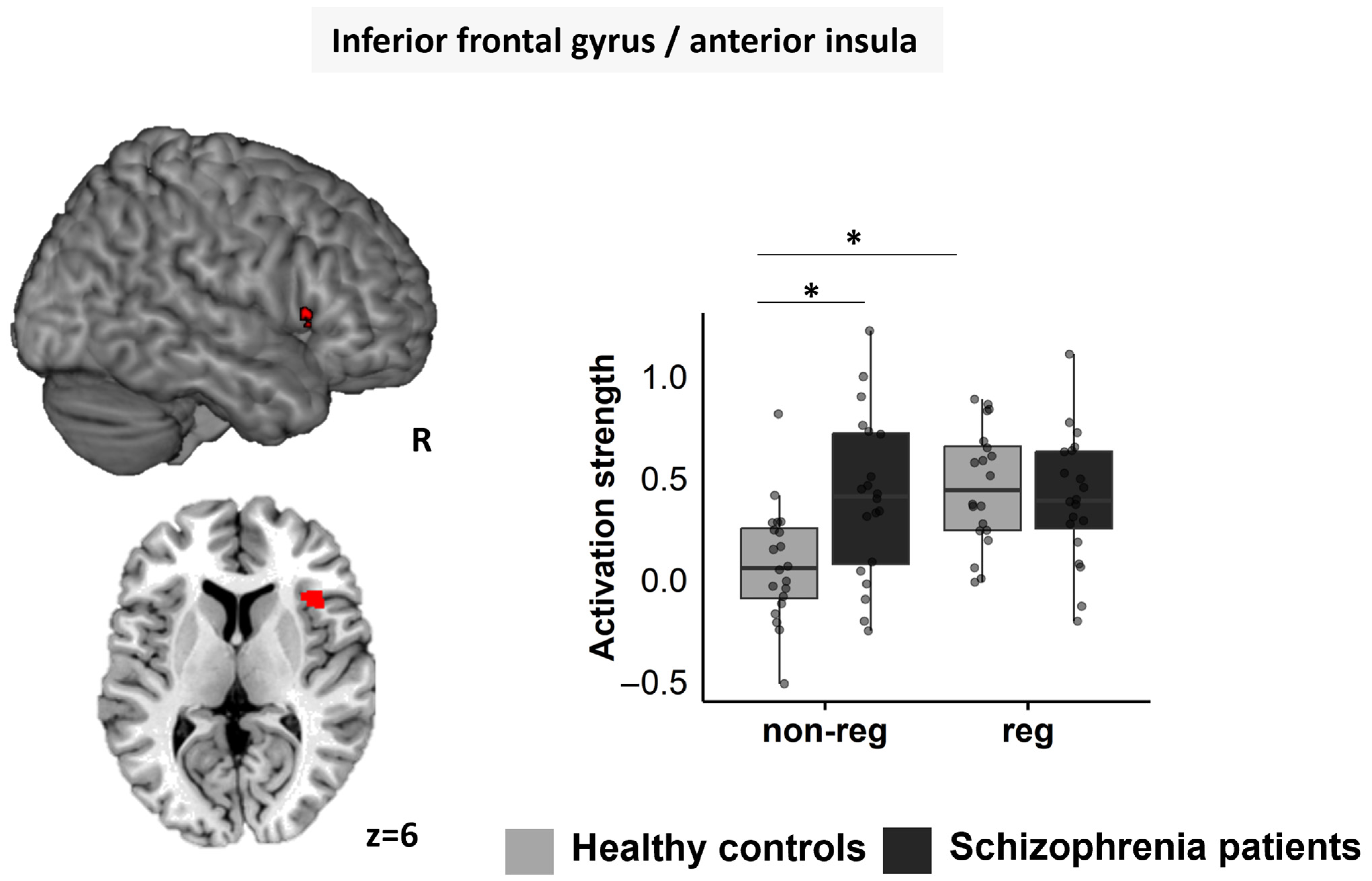
3.5.2. Right IFG/aI
4. Discussion
4.1. Group Differences in Non-Regulated Stress
4.2. Group Differences in Stress Regulation
4.3. Limitations and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirschbaum, C.; Pirke, K.M.; Hellhammer, D.H. The ‘Trier Social Stress Test’—A Tool for Investigating Psychobiological Stress Responses in a Laboratory Setting. Neuropsychobiology 1993, 28, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.F.; Diforio, D. Schizophrenia: A Neural Diathesis-Stress Model. J. Psychol. Rev. 1997, 104, 667–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myin-Germeys, I.; van Os, J.; Schwartz, J.E.; Stone, A.A.; Delespaul, P.A. Emotional Reactivity to Daily Life Stress in Psychosis. Arch. Gen. Psychiatry 2001, 58, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Aas, M.; Dazzan, P.; Mondelli, V.; Toulopoulou, T.; Reichenberg, A.; Di Forti, M.; Fisher, H.L.; Handley, R.; Hepgul, N.; Marques, T.; et al. Abnormal Cortisol Awakening Response Predicts Worse Cognitive Function in Patients with First-Episode Psychosis. J. Psychol. Med. 2011, 41, 463–476. [Google Scholar] [CrossRef] [Green Version]
- Castro, M.N.; Villarreal, M.F.; Bolotinsky, N.; Papávero, E.; Goldschmidt, M.G.; Costanzo, E.Y. Brain Activation Induced by Psychological Stress in Patients with Schizophrenia. Schizophr. Res. 2015, 168, 313–321. [Google Scholar] [CrossRef]
- Soliman, A.; O’Driscoll, G.A.; Pruessner, J.; Joober, R.; Ditto, B.; Streicker, E.; Goldberg, Y.; Caro, J.; Rekkas, P.V.; Dagher, A. Limbic Response to Psychosocial Stress in Schizotypy: A Functional Magnetic Resonance Imaging Study. J. Schizophr. Res. 2011, 131, 184–191. [Google Scholar] [CrossRef]
- Ochsner, K.N.; Ray, R.D.; Cooper, J.C.; Robertson, E.R.; Chopra, S.; Gabrieli, J.D.E.; Gross, J.J. For Better or for Worse: Neural Systems Supporting the Cognitive down- and up-Regulation of Negative Emotion. Neuroimage 2004, 23, 483–499. [Google Scholar] [CrossRef]
- Berking, M.; Wupperman, P. Emotion Regulation and Mental Health: Recent Findings, Current Challenges, and Future Directions. Curr. Opin. Psychiatry 2012, 25, 128–134. [Google Scholar] [CrossRef]
- Raio, C.M.; Orederu, T.A.; Palazzolo, L.; Shurick, A.A.; Phelps, E.A. Cognitive Emotion Regulation Fails the Stress Test. Proc. Natl. Acad. Sci. USA 2013, 110, 15139–15144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogler, L.; Gur, R.C.; Derntl, B. Sex Differences in Cognitive Regulation of Psychosocial Achievement Stress: Brain and Behavior. Hum. Brain Mapp. 2015, 36, 1028–1042. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.W.; Sparks, A.; Mitchell, P.B.; Weickert, C.S.; Green, M.J. Lack of Cortico-Limbic Coupling in Bipolar Disorder and Schizophrenia during Emotion Regulation. Transl. Psychiatry 2012, 2, e90. [Google Scholar] [CrossRef] [Green Version]
- Perry, Y.; Henry, J.D.; Grisham, J.R. The Habitual Use of Emotion Regulation Strategies in Schizophrenia. Br. J. Clin. Psychol. 2011, 50, 217–222. [Google Scholar] [CrossRef]
- van der Meer, L.; Swart, M.; van der Velde, J.; Pijnenborg, G.; Wiersma, D.; Bruggeman, R.; Aleman, A. Neural Correlates of Emotion Regulation in Patients with Schizophrenia and Non-Affected Siblings. PLoS ONE 2014, 9, e99667. [Google Scholar] [CrossRef] [Green Version]
- Wittchen, H.-U.; Zaudig, M.; Fydrich, T. Strukturiertes Klinisches Interview Für DSM-IV; Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Schmidt, K.H.; Metzler, P. Wortschatztest (WST); Beltz Test GmbH: Weinheim, Germany, 1992. [Google Scholar]
- Reitan, R. Trail Making Test: Manual for Administration, Scoring and Interpretation; Indiana University: Bloomington, IN, USA, 1956. [Google Scholar]
- Aster, M.; Neubauer, A.; Horn, R. Wechsler Intelligenztest Für Erwachsene (WIE). Deutschsprachige Bearbeitung Und Adaption Des WAIS-III von David Wechsler; Harcourt: Frankfurt, Germany, 2006. [Google Scholar]
- Kälin, W. Deutsche 24-Item Kurzform Des “Coping Inventory for Stressful Situations” (CISS); von Endler, N.S., Parker, J.D.A., Eds.; Universität, Institut für Psychologie: Bern, Germany, 1995. [Google Scholar]
- Abler, B.; Kessler, H. Emotion Regulation Questionnaire–Eine Deutschsprachige Fassung Des ERQ von Gross Und John. Diagnostica 2009, 55, 144–152. [Google Scholar] [CrossRef]
- Hautzinger, M.; Keller, F.; Kühner, C. Beck Depression Inventar II (BDI 2); Harcourt Test Service: Frankfurt, Germany, 2006. [Google Scholar]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bul. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Chen, J.; Patil, K.R.; Weis, S.; Sim, K.; Nickl-Jockschat, T.; Zhou, J.; Aleman, A.; Sommer, I.E.; Liemburg, E.J.; Hoffstaedter, F.; et al. Neurobiological Divergence of the Positive and Negative Schizophrenia Subtypes Identified on a New Factor Structure of Psychopathology Using Non-Negative Factorization: An International Machine Learning Study. Biol. Psychiatry 2020, 87, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Dedovic, K.; Renwick, R.; Khalili-Mahani, N.; Engert, V.; Lupien, S.J.; Pruessner, J.C. The Montreal Imaging Stress Task: Using Functional Imaging to Investigate the Effects of Perceiving and Processing Psychosocial Stress in the Human Brain. J. Psychiatry Neurosci. 2005, 30, 319–325. [Google Scholar]
- Dedovic, K.; Rexroth, M.; Wolff, E.; Duchesne, A.; Scherling, C.; Beaudry, T.; Lue, S.D.; Lord, C.; Engert, V.; Pruessner, J.C. Neural Correlates of Processing Stressful Information: An Event-Related FMRI Study. Brain Res. 2009, 1293, 49–60. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and Validation of Brief Measures of Positive and Negative Affect: The PANAS Scales. J. Pers. Soc. Psychol. 1988, 54, 1063–1070. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Unified Segmentation. Neuroimage 2005, 26, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, S.B.; Stephan, K.E.; Mohlberg, H.; Grefkes, C.; Fink, G.R.; Amunts, K.; Zilles, K. A New SPM Toolbox for Combining Probabilistic Cytoarchitectonic Maps and Functional Imaging Data. Neuroimage 2005, 25, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Kogler, L.; Müller, V.I.; Chang, A.; Eickhoff, S.B.; Fox, P.T.; Gur, R.C.; Derntl, B. Psychosocial versus Physiological Stress-Meta-Analyses on the Deactivations and Activations of the Neural Correlates of Stress Reactions. Neuroimage 2015, 119, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Tischler, L.; Petermann, F.; Trail Making Test (TMT). Zeitschrift für Psychiatrie, Psychologie und Psychotherapie. Z. Psychiatr. Psychol. Psychother. 2010, 58, 79–81. [Google Scholar] [CrossRef]
- Rokita, K.I.; Dauvermann, M.R.; Mothersill, D.; Holleran, L.; Bhatnagar, P.; McNicholas, Á.; McKernan, D.; Morris, D.W.; Kelly, J.; Hallahan, B.; et al. Current psychosocial stress, childhood trauma and cognition in patients with schizophrenia and healthy participants. Schizophr. Res. 2021, 237, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Carol, E.E.; Spencer, R.L.; Mittal, V.A. Acute Physiological and Psychological Stress Response in Youth at Clinical High-Risk for Psychosis. Front. Psychiatry 2021, 12, 641762. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Amunts, K.; Mohlberg, H.; Zilles, K. The Human Parietal Operculum. II. Stereotaxic Maps and Correlations with Functional Imaging Results. Cereb. Cortex 2006, 16, 268–279. [Google Scholar] [CrossRef]
- Chang, M.; Womer, F.Y.; Bai, C.; Zhou, Q.; Wei, S.; Jiang, X.; Geng, H.; Zhou, Y.; Tang, Y.; Wang, F. Voxel-Based Morphometry in Individuals at Genetic High Risk for Schizophrenia and Patients with Schizophrenia during Their First Episode of Psychosis. PLoS ONE 2016, 11, e0163749. [Google Scholar] [CrossRef] [Green Version]
- Schilbach, L.; Hoffstaedter, F.; Müller, V.I.; Cieslik, E.C.; Goya-Maldonado, R.; Trost, S.; Sorg, C.; Riedl, V.; Jardri, R.; Sommer, I.; et al. Transdiagnostic Commonalities and Differences in Resting State Functional Connectivity of the Default Mode Network in Schizophrenia and Major Depression. NeuroImage Clin. 2015, 10, 326–335. [Google Scholar] [CrossRef]
- Morawetz, C.; Bode, S.; Derntl, B.; Heekeren, H.R. The Effect of Strategies, Goals and Stimulus Material on the Neural Mechanisms of Emotion Regulation: A Meta-Analysis of FMRI Studies. Neurosci. Biobehav. Rev. 2017, 725, 111–128. [Google Scholar] [CrossRef]
- Massey, S.H.; Stern, D.; Alden, E.C.; Petersen, J.E.; Cobia, D.J.; Wang, L.; Csernansky, J.G.; Smith, M.J. Cortical Thickness of Neural Substrates Supporting Cognitive Empathy in Individuals with Schizophrenia. Schizophr. Res. 2017, 179, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolls, E.T.; Cheng, W.; Gilson, M.; Gong, W.; Deco, G.; Lo, C.Z.; Yang, A.C.; Tsai, S.; Liu, M.; Lin, C.; et al. Beyond the Disconnectivity Hypothesis of Schizophrenia. Cereb Cortex 2019, 30, 1213–1233. [Google Scholar] [CrossRef] [PubMed]
- Radua, J.; Borgwardt, S.; Crescini, A.; Mataix-Cols, D.; Meyer-Lindenberg, A. Multimodal Meta-Analysis of Structural and Functional Brain Changes in First Episode Psychosis and the Effects of Antipsychotic Medication. Neurosci. Biobehav. Rev. 2012, 36, 2325–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohi, K.; Hashimoto, R.; Yasuda, Y.; Nemoto, K.; Ohnishi, T. Impact of the Genome Wide Supported NRGN Gene on Anterior Cingulate Impact of the Genome Wide Supported NRGN Gene on Anterior Cingulate Morphology in Schizophrenia. PLoS ONE 2012, 7, e29780. [Google Scholar] [CrossRef] [Green Version]
- Rietschel, M.; Mattheisen, M.; Degenhardt, F.; Investigators, G.; Mu, T.W.; Czerski, P.M.; Giegling, I.; Strengman, E.; Schmael, C.; Mors, O.; et al. Association between Genetic Variation in a Region on Chromosome 11 and Schizophrenia in Large Samples from Europe. Mol. Psychiatry 2012, 17, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Marwood, L.; Wise, T.; Perkins, A.M.; Cleare, A.J. Meta-Analyses of the Neural Mechanisms and Predictors of Response to Psychotherapy in Depression and Anxiety. Neurosci. Biobehav. Rev. 2018, 95, 61–72. [Google Scholar] [CrossRef]
- Wada, M.; Noda, Y.; Iwata, Y.; Tsugawa, S.; Yoshida, K.; Tani, H.; Hirano, Y.; Koike, S.; Sasabayashi, D.; Katayama, H.; et al. Dopaminergic dysfunction and excitatory/inhibitory imbalance in treatment-resistant schizophrenia and novel neuromodulatory treatment. Mol. Psychiatry 2022, 27, 2950–2967. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- WHO. Gender in Mental Health Research; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Hodes, G.E.; Epperson, C.N. Sex Differences in Vulnerability and Resilience to Stress Across the Life Span. Biol. Psychiatry 2019, 86, 421–432. [Google Scholar] [CrossRef]
- Gur, R.E.; Moore, T.M.; Rosen, A.F.G.; Barzilay, R.; Roalf, D.R.; Calkins, M.E.; Ruparel, K.; Scott, J.C.; Almasy, L.; Satterthwaite, T.D.; et al. Burden of Environmental Adversity Associated with Psychopathology, Maturation, and Brain Behavior Parameters in Youths. JAMA Psychiatry 2019, 76, 966–975. [Google Scholar] [CrossRef]
- King, A.; Liberzon, I. Assessing the neuroendocrine stress response in the functional neuroimaging context. Neuroimage 2009, 47, 1116–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boucsein, W.; Fowles, D.; Christie, M.; Grimnes, S.; Ben-Shakhar, G.; Roth, W.T.; Dawson, M.E.; Filion, D.L. Publication recommendations for electrodermal measurements. Psychophysiology 2012, 49, 1017–1034. [Google Scholar] [CrossRef] [PubMed]
- Benedek, M.; Kaernbach, C. A continuous measure of phasic electrodermal activity. J. Neurosci. Methods 2010, 190, 80–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemmerich, W. StatistikGuru: Rechner zur Adjustierung des α-Niveaus. 2016. Available online: https://statistikguru.de/rechner/adjustierung-des-alphaniveaus.html (accessed on 23 March 2020).
- Eickhoff, S.B.; Schleicher, A.; Zilles, K.; Amunts, K. The human parietal operculum. I. Cytoarchitectonic mapping of subdivisions. Cereb. Cortex 2006, 16, 254–267. [Google Scholar] [CrossRef]
- Geyer, S.; Schormann, T.; Mohlberg, H.; Zilles, K. Areas 3a, 3b, and 1 of human primary somatosensory cortex. Part 2. Spatial normalization to standard anatomical space. Neuroimage 2000, 11, 684–696. [Google Scholar] [CrossRef]
- Caspers, S.; Geyer, S.; Schleicher, A.; Mohlberg, H.; Amunts, K.; Zilles, K. The human inferior parietal cortex: Cytoarchitectonic parcellation and interindividual variability. Neuroimage 2006, 33, 430–448. [Google Scholar] [CrossRef]
- Bludau, S.; Eickhoff, S.B.; Mohlberg, H.; Caspers, S.; Laird, A.R.; Fox, P.T.; Schleicher, A.; Zilles, K.; Amunts, K. Cytoarchitecture, probability maps and functions of the human frontal pole. Neuroimage 2014, 93, 260–275. [Google Scholar] [CrossRef] [Green Version]
- Amunts, K.; Schleicher, A.; Bürgel, U.; Mohlberg, H.; Uylings, H.B.; Zilles, K. Broca’s region revisited: Cytoarchitecture and intersubject variability. J. Comp. Neurol. 1999, 412, 319–341. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SZP n = 20 | HC n = 20 | ||||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | p-Value | |
| Age (years) | 36.75 | 10.33 | 33.75 | 10.30 | 0.363 |
| Working memory (raw score) | 13.55 | 4.03 | 18.11 a | 4.11 | 0.001 |
| Verbal intelligence (raw score) | 29.32 a | 8.49 | 33.05 | 3.50 | 0.088 |
| Processing speed (TMT-A, s) | 27.21 | 7.36 | 19.56 | 8.89 | 0.005 |
| Cognit. flexibility (TMT-B, s) | 55.58 | 33.37 | 30.70 | 8.97 | 0.004 |
| BDI-II | 9.85 | 5.91 | 2.85 | 3.08 | <0.001 |
| Emotion regulation | |||||
| Suppression | 4.03 b | 1.28 | 4.24 a | 1.34 | 0.631 |
| Reappraisal | 4.42 b | 0.86 | 4.50 a | 1.40 | 0.830 |
| Stress coping | |||||
| Task oriented stress coping | 3.53 a | 0.64 | 3.60 | 0.65 | 0.753 |
| Emotion oriented stress coping | 2.54 a | 0.65 | 2.49 | 0.68 | 0.821 |
| Distraction | 2.62 a | 0.63 | 2.18 | 0.84 | 0.071 |
| Social diversion | 3.12 a | 0.83 | 3.24 | 0.77 | 0.644 |
| Avoidance | 2.87 a | 0.64 | 2.71 | 0.58 | 0.412 |
| Highest educational degree | 0.075 | ||||
| Elementary school | 0 | 0 | |||
| Secondary modern school | 0 | 0 | |||
| Junior high school | 2 | 3 | |||
| Vocational diploma | 0 | 10 | |||
| Secondary school | 5 | 0 | |||
| Academic studies | 6 | 3 | |||
| Training | 7 | 4 | |||
| Doctorate | 0 | 0 | |||
| Clinical parameters | |||||
| Age of onset (years) | 27.20 | 7.86 | |||
| Duration of illness (years) | 9.53 | 8.66 | |||
| Number of episodes | 3.28 b | 2.47 | |||
| Global symptoms | 26.95 | 7.54 | |||
| Positive symptoms | 15.50 | 5.93 | |||
| Negative symptoms | 11.80 | 4.63 | |||
| Negative dimension | 4.34 | 1.60 | |||
| Positive dimension | 4.86 | 1.98 | |||
| Affective dimension | 5.47 | 1.67 | |||
| Cognitive dimension | 4.97 | 2.20 | |||
| Global functioning | 51.53 | 13.97 | |||
| CPZ-equivalents | 536.30 b | 275.77 | |||
| OLZ-equivalents | 20.26 b | 9.00 | |||
| SZP | HC | |||
|---|---|---|---|---|
| Pre-Stress | Post-Stress | Pre-Stress | Post-Stress | |
| Subjective stress | ||||
| Non-regulation | 1.68 (0.88) | 2.74 (1.24) | 1.33 (0.59) | 2.78 (1.22) |
| Regulation | 2.05 (1.18) | 2.58 (1.26) | 1.28 (0.67) | 2.89 (1.28) |
| Negative affect | ||||
| Non-regulation | 13.47 (4.10) | 17.79 (7.00) | 10.11(3.71) | 13.06 (5.90) |
| Regulation | 14.11 (4.65) | 17.05 (6.23) | 12.00 (3.22) | 17.50 (6.72) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kogler, L.; Regenbogen, C.; Müller, V.I.; Kohn, N.; Schneider, F.; Gur, R.C.; Derntl, B. Cognitive Stress Regulation in Schizophrenia Patients and Healthy Individuals: Brain and Behavior. J. Clin. Med. 2023, 12, 2749. https://doi.org/10.3390/jcm12072749
Kogler L, Regenbogen C, Müller VI, Kohn N, Schneider F, Gur RC, Derntl B. Cognitive Stress Regulation in Schizophrenia Patients and Healthy Individuals: Brain and Behavior. Journal of Clinical Medicine. 2023; 12(7):2749. https://doi.org/10.3390/jcm12072749
Chicago/Turabian StyleKogler, Lydia, Christina Regenbogen, Veronika I. Müller, Nils Kohn, Frank Schneider, Ruben C. Gur, and Birgit Derntl. 2023. "Cognitive Stress Regulation in Schizophrenia Patients and Healthy Individuals: Brain and Behavior" Journal of Clinical Medicine 12, no. 7: 2749. https://doi.org/10.3390/jcm12072749
APA StyleKogler, L., Regenbogen, C., Müller, V. I., Kohn, N., Schneider, F., Gur, R. C., & Derntl, B. (2023). Cognitive Stress Regulation in Schizophrenia Patients and Healthy Individuals: Brain and Behavior. Journal of Clinical Medicine, 12(7), 2749. https://doi.org/10.3390/jcm12072749