
Enhanced Efficacy of Doxorubicin by microRNA-499-Mediated Improvement of Tumor Blood Flow

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Experimental Section
2.1. Preparation of Tetraethylenepentamine-Based Polycation Liposomes (TEPA-PCL) and Lipoplex
2.2. Cell Culture
2.3. Experimental Animals
2.4. Evaluation of Tumor Blood Flow
2.5. Accumulation of Doxorubicin in the Tumor
2.6. Therapeutic Experiment
2.7. Statistical Analysis
3. Results and Discussion
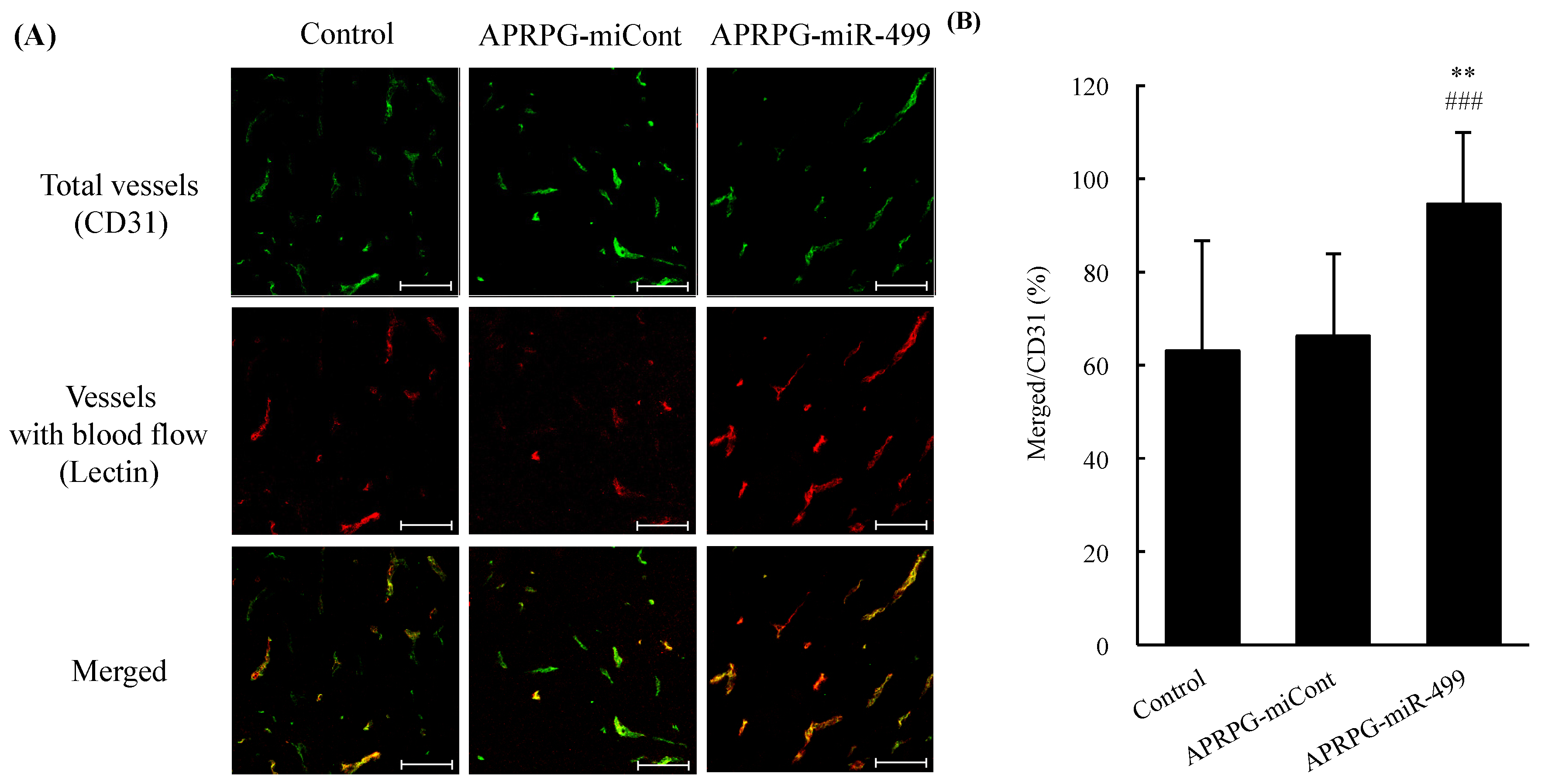
3.1. Amelioration of Incomplete Blood Flow in Tumors Treated with miR-499

3.2. Improvement of Doxorubicin (DOX) Accumulation in Tumors Treated with miR-499

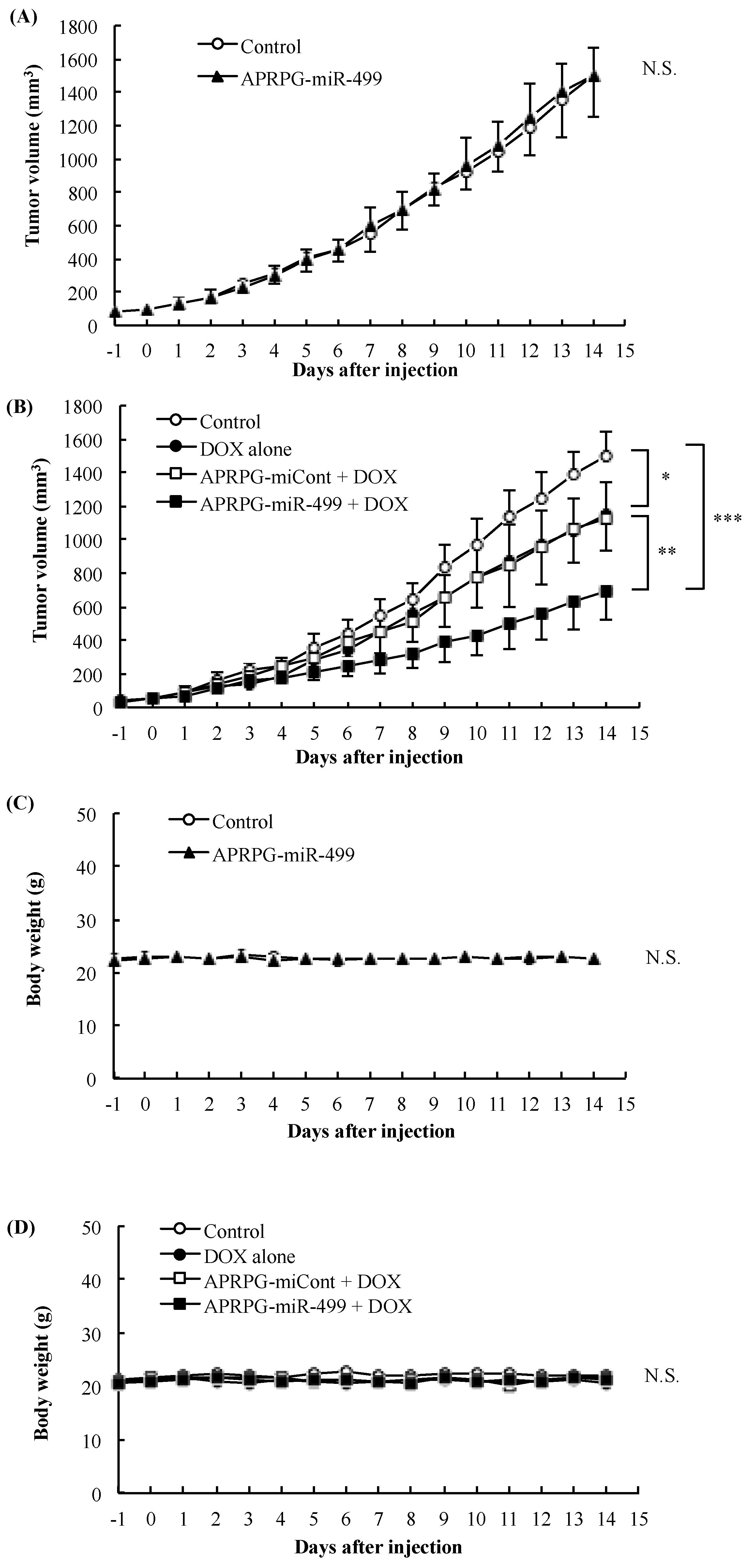
3.3. Combination Therapy with miR-499 and DOX

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wilson, K.D.; Hu, S.; Venkatasubrahmanyam, S.; Fu, J.-D.; Sun, N.; Abilez, O.J.; Baugh, J.J.A.; Jia, F.; Ghosh, Z.; Li, R.A.; et al. Dynamic microRNA expression programs during cardiac differentiation of human embryonic stem cells: Role for miR-499. Circ. Cardiovasc. Genet. 2010, 3, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Liu, J.J.; Liu, F.; Liu, W.H.; Wang, Y.S.; Zhu, B.; Yu, B. MiR-499 induces cardiac differentiation of rat mesenchymal stem cells through Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2012, 420, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Drzewiecka, A.W.; Ratajewski, M.; Wagner, W.; Dastych, J. HIF-1α is up-regulated in activated mast cells by a process that involves calcineurin and NFAT. J. Immunol. 2008, 181, 1665–1672. [Google Scholar] [CrossRef]
- Zhang, X.; Gaspard, J.P.; Chung, D.C. Regulation of vascular endothelial growth factor by the Wnt and K-ras pathways in colonic neoplasia. Cancer Res. 2001, 61, 6050–6054. [Google Scholar] [PubMed]
- Ando, H.; Asai, T.; Koide, H.; Okamoto, A.; Maeda, N.; Tomita, K.; Dewa, T.; Minamino, T.; Oku, N. Advanced cancer therapy by integrative antitumor actions via systemic administration of miR-499. J. Control Release 2014, 181, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Matsushita, S.; Kenjo, E.; Tsuzuku, T.; Yonenaga, N.; Koide, H.; Hatanaka, K.; Dewa, T.; Nango, M.; Maeda, N.; et al. Dicetyl phosphate-tetraethylenepentamine-based liposomes for systemic siRNA delivery. Bioconj. Chem. 2011, 22, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Okamoto, A.; Yokota, M.; Shimizu, K.; Asai, T.; Dewa, T.; Oku, N. Development of a miR-92a delivery system for anti-angiogenesis-based cancer therapy. J. Gene Med. 2013, 15, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Okamoto, A.; Yokota, M.; Asai, T.; Dewa, T.; Oku, N. Polycation liposomes as a vector for potential intracellular delivery of microRNA. J. Gene Med. 2013, 15, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Yang, L.; Ling, X.; Yang, R.; Yang, X.; Zhang, L.; Fang, W.; Xie, C.; Huang, D.; Zhou, Y.; et al. Sequence variation in mature microRNA-499 confers unfavorable prognosis of lung cancer patients treated with platinum-based chemotherapy. Clin. Cancer Res. 2015, 21, 1602–1163. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [PubMed]
- Folkman, J.; Klagsbrun, M. Angiogenic factors. Science 1987, 235, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Barriers to drug delivery in solid tumors. Sci. Am. 1994, 271, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Wildiers, H.; Guetens, G.; Boeck, G.D.; Verbeken, E.; Landuyt, B.; Landuyt, W.; de Bruijn, E.A.; van Oosterom, A.T. Effect of antivascular endothelial growth factor treatment on the intratumoral uptake of CPT-11. Br. J. Cancer 2003, 88, 1979–1986. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, D.; Jain, R.K. Tumor microvasculature and microenvironment: Targets for anti-angiogenesis and normalization. Microvasc. Res. 2007, 74, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Tong, R.T.; Boucher, Y.; Kozin, S.V.; Winkler, F.; Hicklin, D.J.; Jain, R.K. Vascular normalization by vascular endothelial growth factor receptor 2 blockade induces a pressure gradient across the vasculature and improves drug penetration in tumors. Cancer Res. 2004, 64, 3731–3736. [Google Scholar] [CrossRef] [PubMed]
- Saranadasa, M.; Wang, E.S. Vascular endothelial growth factor inhibition: Conflicting roles in tumor growth. Cytokine 2011, 53, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Klement, G.; Huang, P.; Mayer, B.; Green, S.K.; Man, S.; Bohlen, P.; Hicklin, D.; Kerbel, R.S. Differences in therapeutic indexes of combination metronomic chemotherapy and an anti-VEGFR-2 antibody in multidrug-resistant human breast cancer xenografts. Clin. Cancer Res. 2002, 8, 221–232. [Google Scholar] [PubMed]
- Jain, R.K. Normalization of tumor vasculature: An emerging concept in antiangiogenic therapy. Science 2005, 307, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.I.; Sessa, W.C. Antiangiogenic therapy: Creating a unique “window” of opportunity. Cancer Cell 2004, 6, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Kozin, S.V.; Tong, R.T.; Chae, S.S.; Booth, M.F.; Garkavtsev, I.; Xu, L.; Hicklin, D.J.; Fukumura, D.; di Tomaso, E.; et al. Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response to radiation: Role of oxygenation, angiopoietin-1, and matrix metalloproteinases. Cancer Cell 2004, 6, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, H.; Fehrenbacher, L.; Novotny, W.; Cartwright, T.; Hainsworth, J.; Heim, W.; Berlin, J.; Baron, A.; Griffing, S.; Holmgren, E.; et al. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N. Engl. J. Med. 2004, 350, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Chen, L. Recombinant human endostatin improves anti-tumor efficacy of paclitaxel by normalizing tumor vasculature in Lewis lung carcinoma. J. Cancer Res. Clin. Oncol. 2010, 136, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Datta, M.; Via, L.E.; Kamoun, W.S.; Liu, C.; Chen, W.; Seano, G.; Weiner, D.M.; Schimel, D.; England, K.; Martin, J.D.; et al. Anti-vascular endothelial growth factor treatment normalizes tuberculosis granuloma vasculature and improves small molecule delivery. Proc. Natl. Acad. Sci. USA 2015, 112, 1827–1832. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okamoto, A.; Asai, T.; Ryu, S.; Ando, H.; Maeda, N.; Dewa, T.; Oku, N. Enhanced Efficacy of Doxorubicin by microRNA-499-Mediated Improvement of Tumor Blood Flow. J. Clin. Med. 2016, 5, 10. https://doi.org/10.3390/jcm5010010
Okamoto A, Asai T, Ryu S, Ando H, Maeda N, Dewa T, Oku N. Enhanced Efficacy of Doxorubicin by microRNA-499-Mediated Improvement of Tumor Blood Flow. Journal of Clinical Medicine. 2016; 5(1):10. https://doi.org/10.3390/jcm5010010
Chicago/Turabian StyleOkamoto, Ayaka, Tomohiro Asai, Sho Ryu, Hidenori Ando, Noriyuki Maeda, Takehisa Dewa, and Naoto Oku. 2016. "Enhanced Efficacy of Doxorubicin by microRNA-499-Mediated Improvement of Tumor Blood Flow" Journal of Clinical Medicine 5, no. 1: 10. https://doi.org/10.3390/jcm5010010
APA StyleOkamoto, A., Asai, T., Ryu, S., Ando, H., Maeda, N., Dewa, T., & Oku, N. (2016). Enhanced Efficacy of Doxorubicin by microRNA-499-Mediated Improvement of Tumor Blood Flow. Journal of Clinical Medicine, 5(1), 10. https://doi.org/10.3390/jcm5010010